Dictyostelium: An Important Source of Structural and Functional Diversity in Drug Discovery


Abstract
:1. Introduction
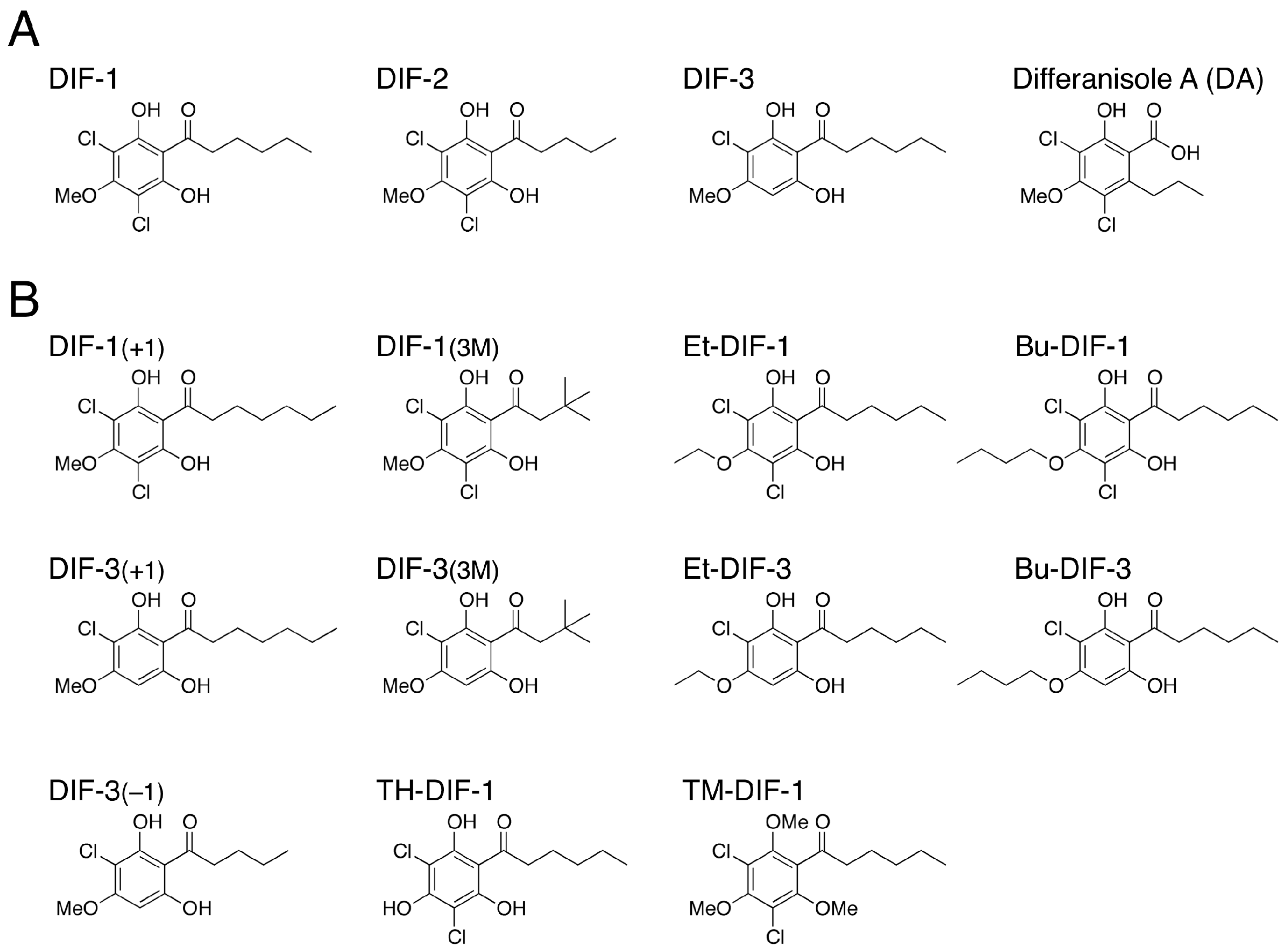
2. Biological and Pharmacological Activities of DIF-1 and Its Derivatives
2.1. Functions of DIF-1, DIF-2 and DIF-3 in D. discoideum
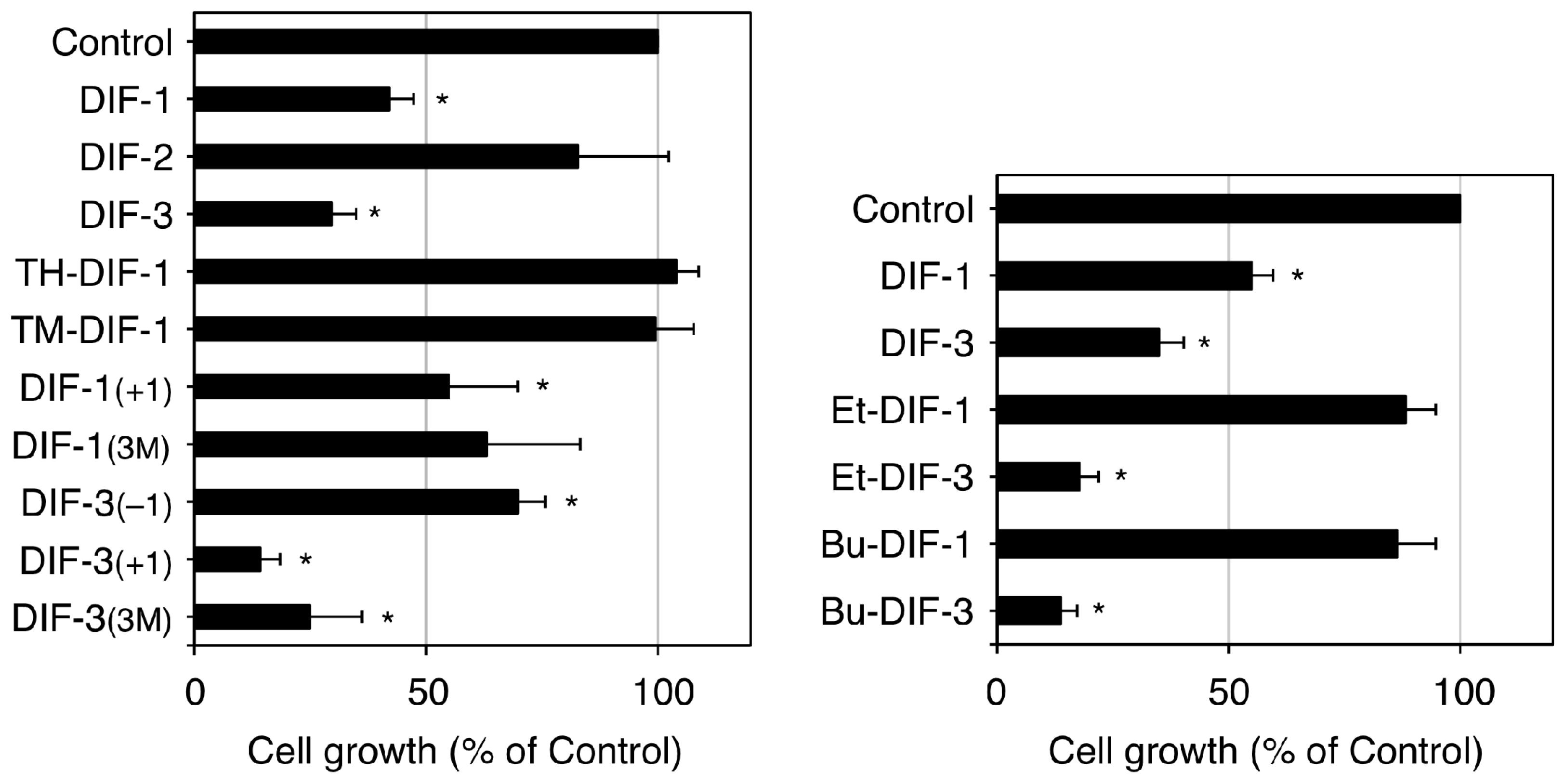
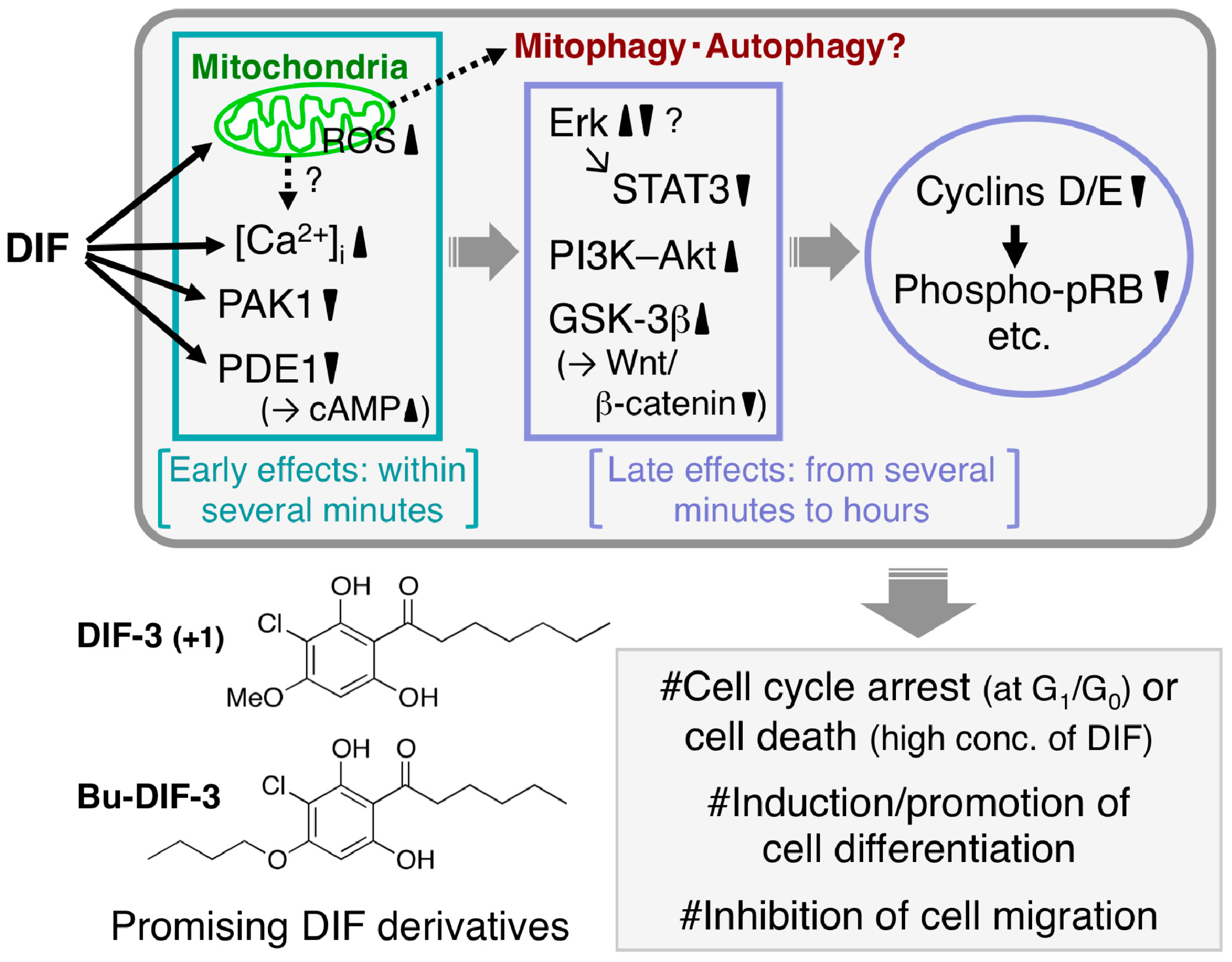
2.2. Discovery of the Antitumor Activities of DIFs
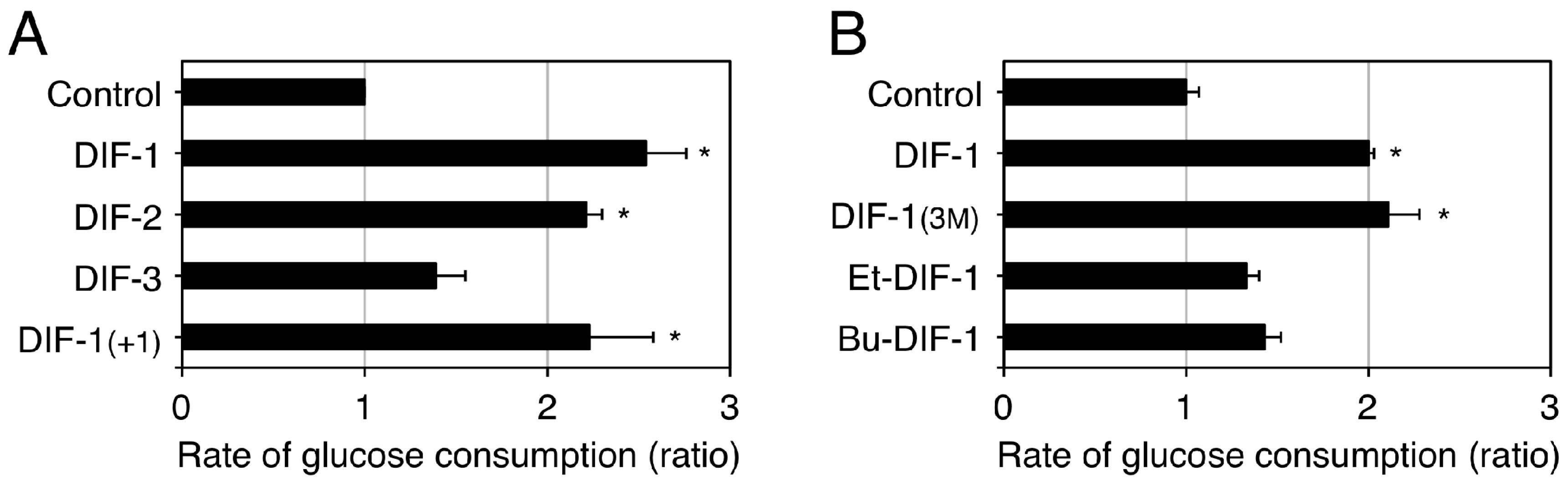
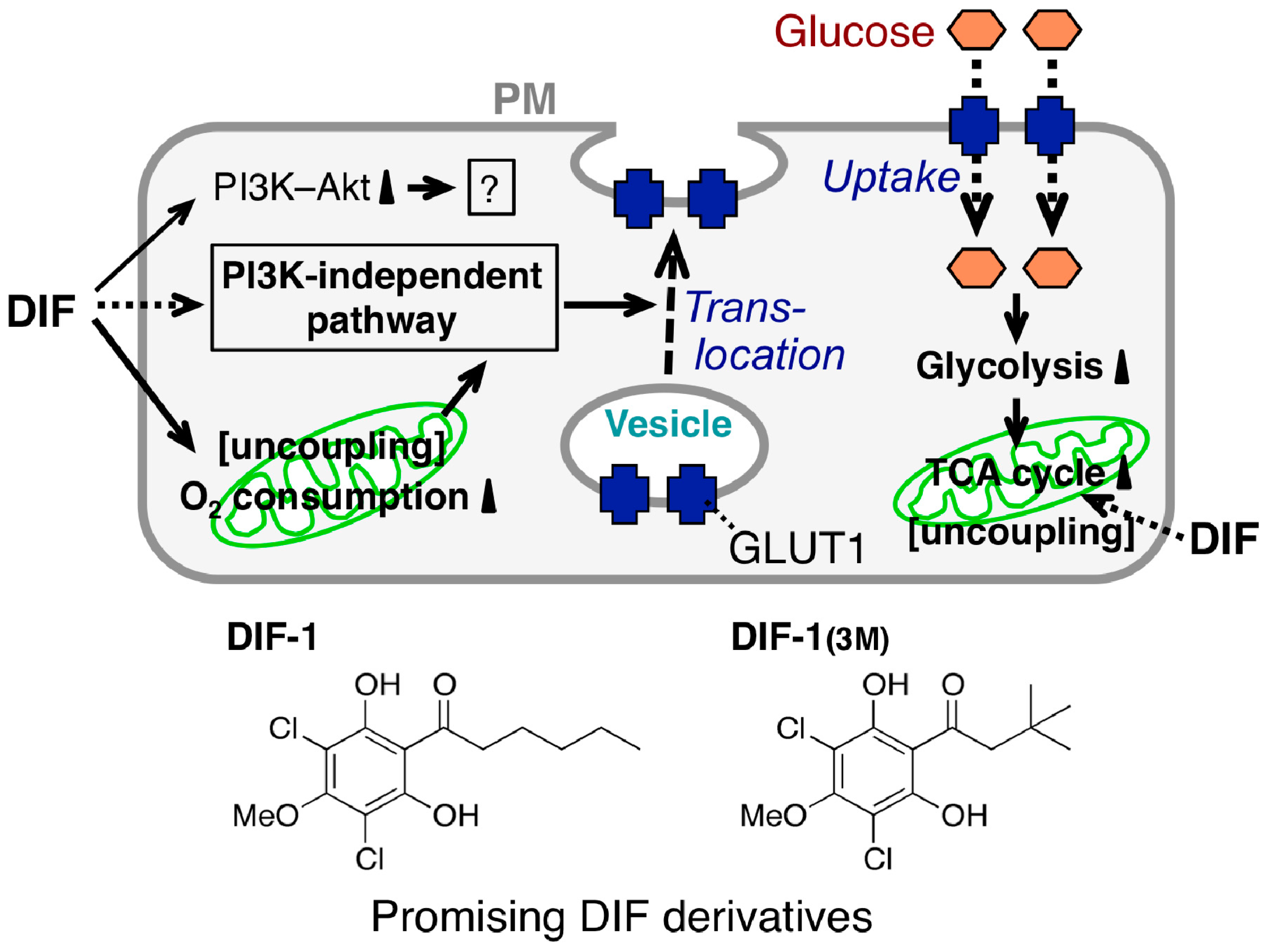
2.3. Glucose Uptake-Promoting Activity of DIF-1
2.4. Other Biological Activities of DIFs
2.4.1. Differentiation-Inducing and -Promoting Activities
2.4.2. Anti-Meiotic Activity
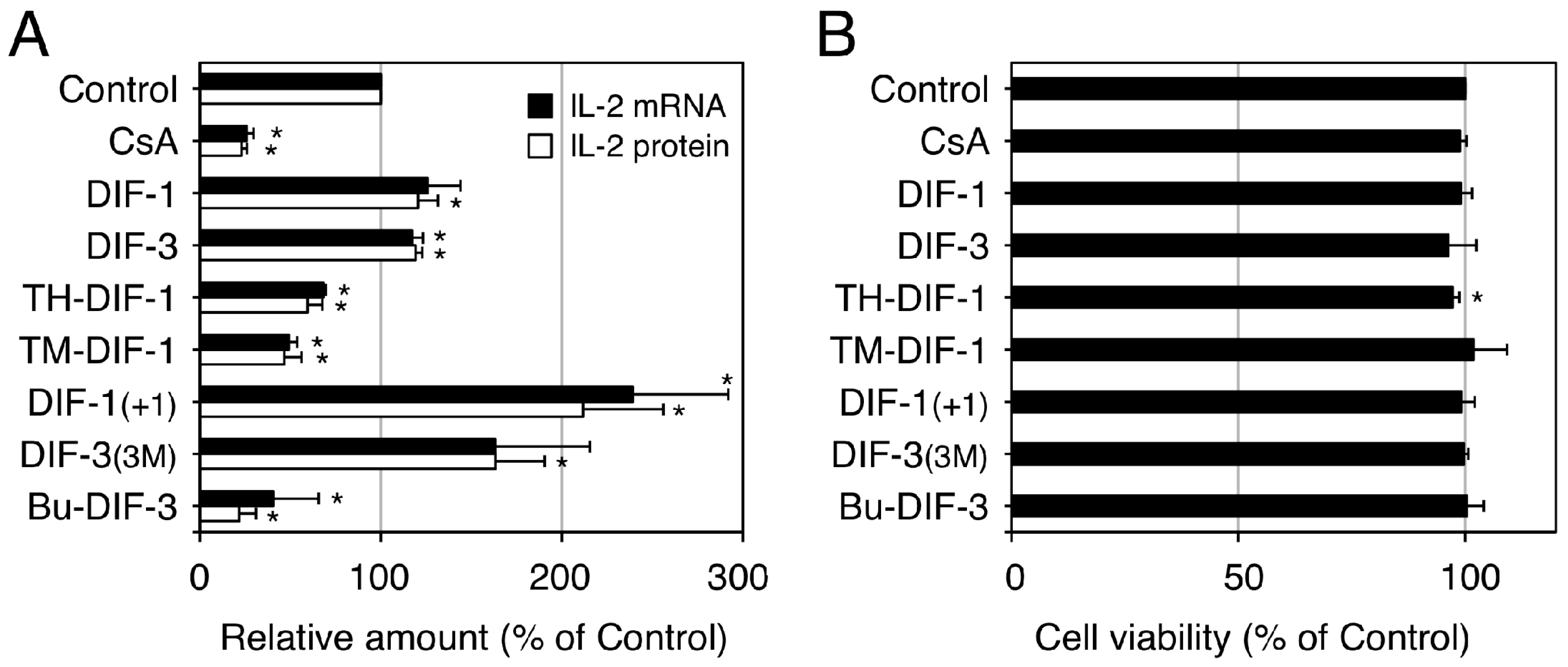
2.4.3. Immunomodulatory Activities in Jurkat T Cells
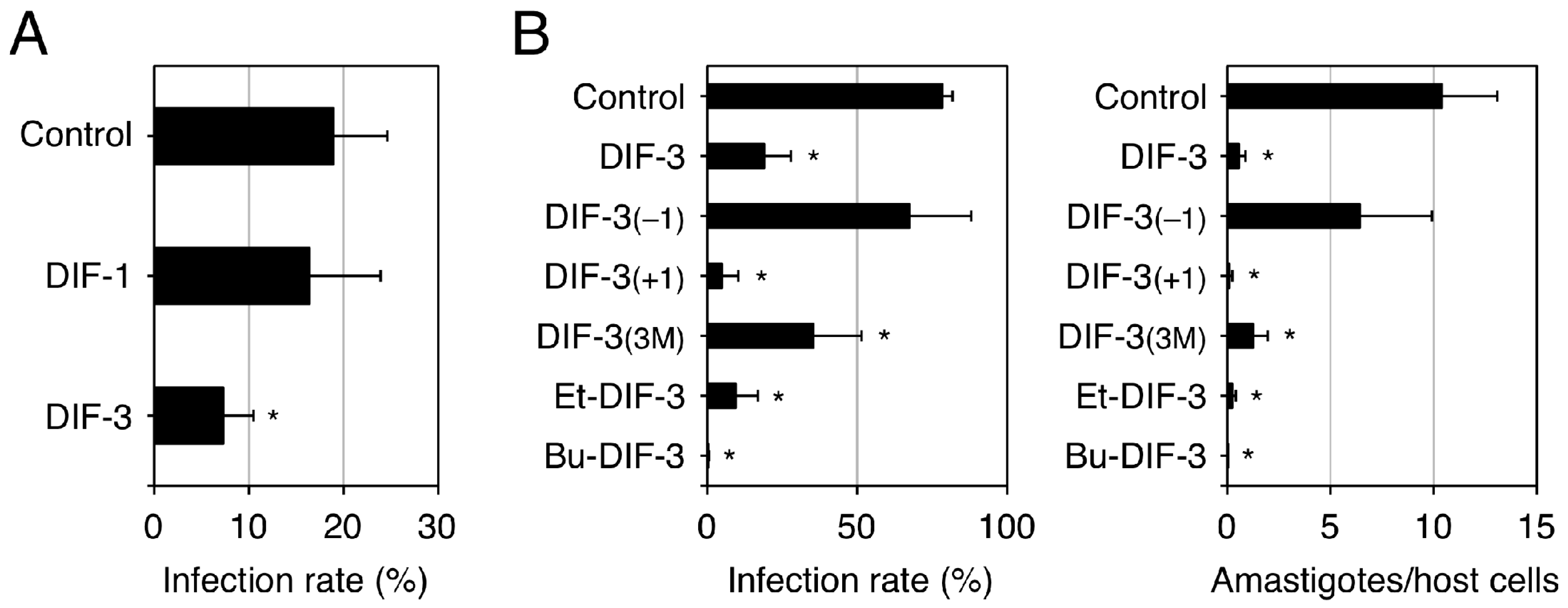
2.4.4. Anti-Trypanosoma Activity
2.4.5. Anti-β-Amyloid Activity
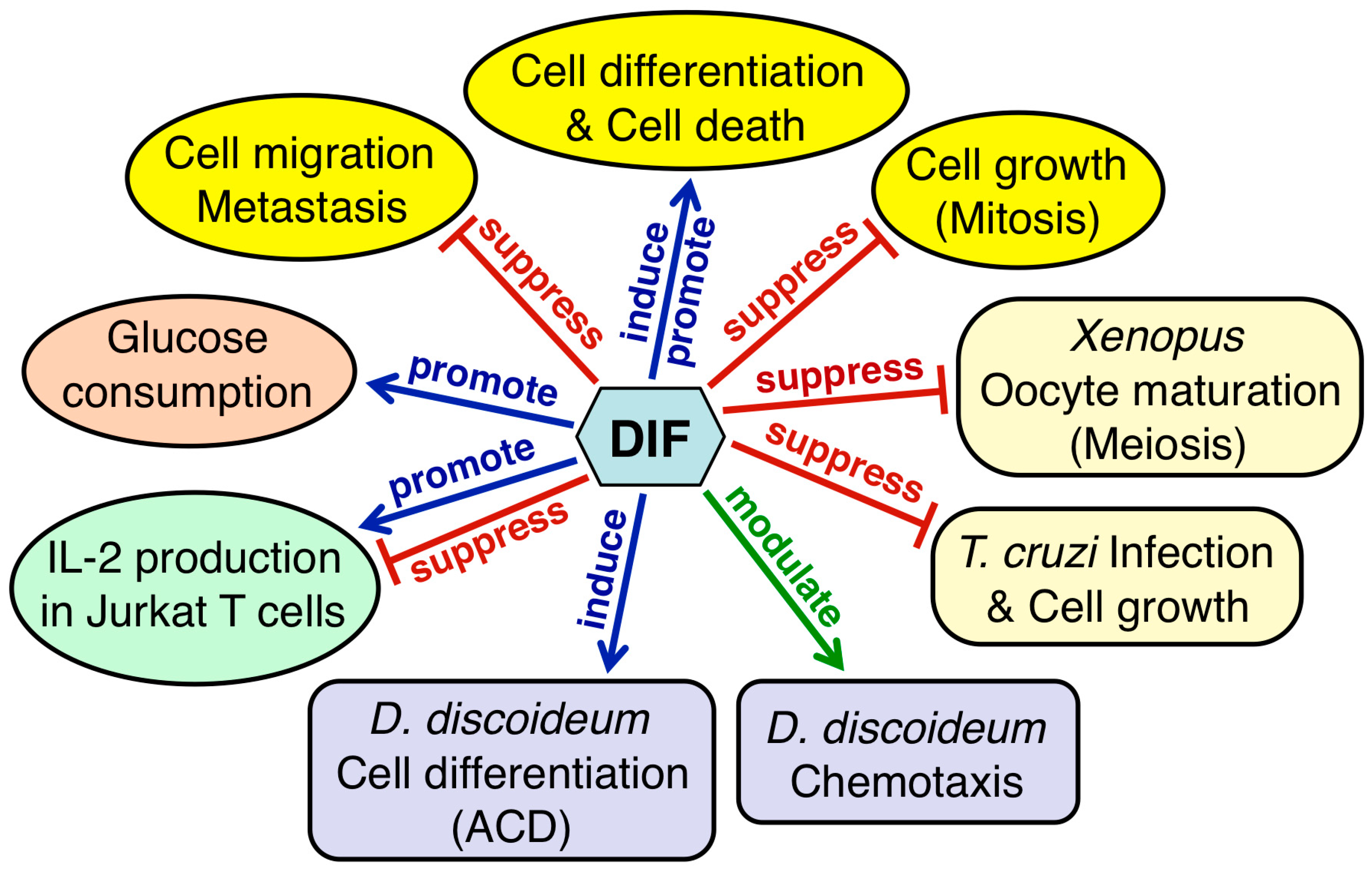
2.4.6. Conclusions on the Activities of DIFs
3. Novel Biologically Active Compounds Found in Cellular Slime Molds
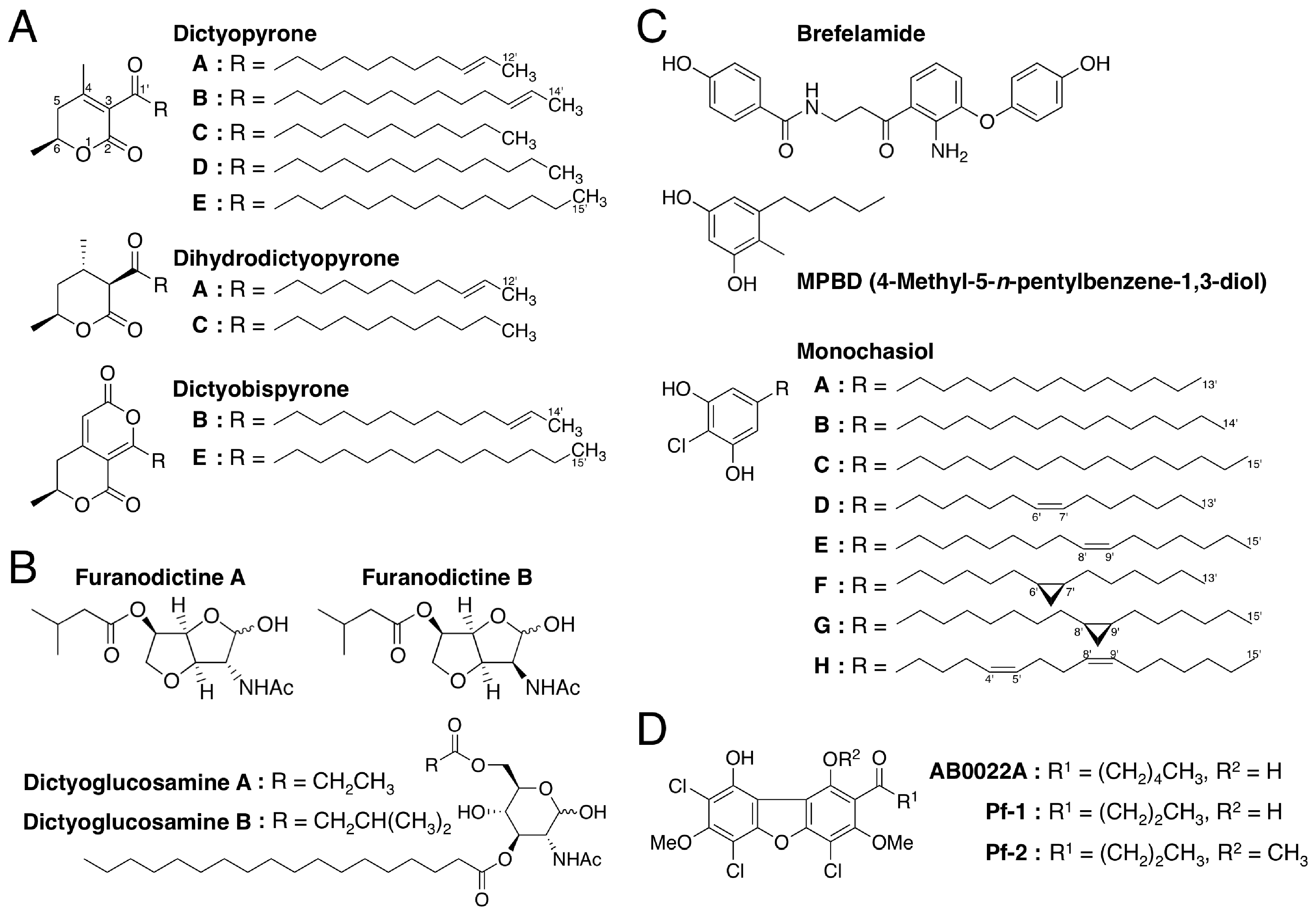
3.1. Dictyopyrones
3.2. Amino Sugar Derivatives: Furanodictines and Dictyoglucosamines
3.3. Brefelamide
3.4. MPBD
3.5. Monochasiols
3.6. Dibenzofurans: AB0022A, Pf-1 and Pf-2
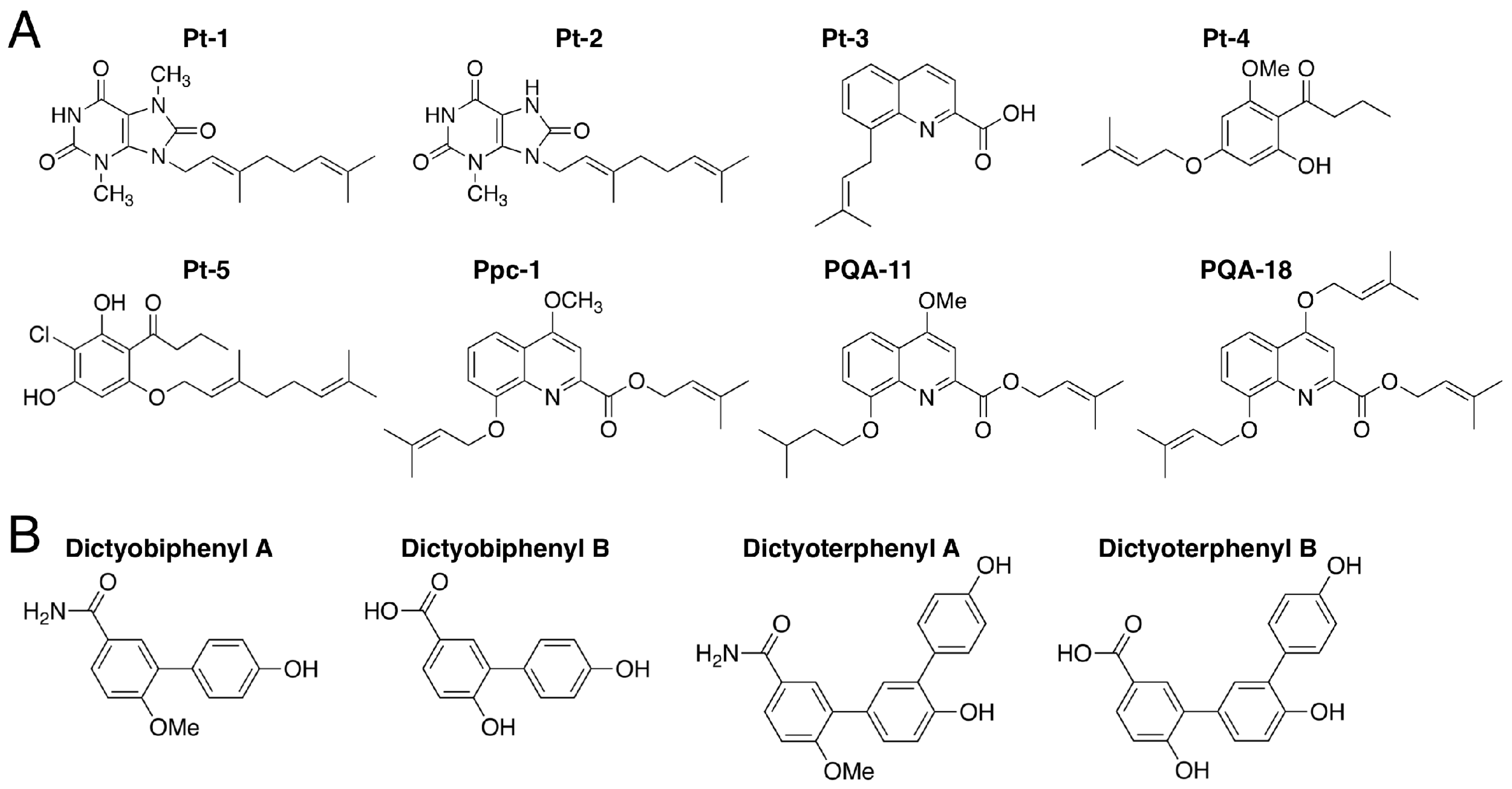
3.7. Prenylated and Geranylated Aromatic Compounds: Pt-1–5 and Ppc-1
3.8. Dictyobiphenyls and Dictyoterphenyls
4. Conclusions
5. Patents
Funding
Acknowledgments
Conflicts of Interest
References
- Clardy, J.; Fischbach, M.A.; Walsh, C.T. New antibiotics from bacterial natural products. Nat. Biotech. 2006, 24, 1541–1550. [Google Scholar] [CrossRef] [PubMed]
- Challinor, V.L.; Bode, H.B. Biozctive natural products from novel microbial sources. Ann. N. Y. Acad. Sci. 2015, 1354, 82–97. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2018, in press. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.D.; Wright, G.D. Antibacterial drug discovery in the resistance era. Nature 2016, 529, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Adl, S.M.; Simpson, A.G.; Farmer, M.A.; Andersen, R.A.; Anderson, O.R.; Barta, J.R.; Bowser, S.S.; Brugerolle, G.; Fensome, R.A.; Fredericq, S.; et al. The new higher level classification of eukaryotes with emphasis on the taxonomy of protists. J. Eukaryot. Microbiol. 2005, 52, 399–451. [Google Scholar] [CrossRef] [PubMed]
- Eichinger, L.; Pachebat, J.A.; Glöckner, G.; Rajandream, M.A.; Sucgang, R.; Berriman, M.; Song, J.; Olsen, R.; Szafranski, K.; Xu, Q.; et al. The genome of the social amoeba Dictyostelium discoideum. Nature 2005, 435, 43–57. [Google Scholar] [CrossRef]
- Adl, S.M.; Simpson, A.G.; Lane, C.E.; Lukeš, J.; Bass, D.; Bowser, S.S.; Brown, M.W.; Burki, F.; Dunthorn, M.; Hampl, V.; et al. The revised classification of eukaryotes. J. Eukaryot. Microbiol. 2012, 59, 429–493. [Google Scholar] [CrossRef]
- Raper, K.B. Dictyostelium discoideum, A new species of slime mold from decaying forest leaves. J. Agric. Res. 1935, 55, 289–316. [Google Scholar]
- Newell, P.C. The development of the cellular slime mould Dictyostelium discoideum: A model system for the study of cellular differentiation. Essays Biochem. 1971, 7, 87–126. [Google Scholar]
- Firtel, R.A.; Meili, R. Dictyostelium: A model for regulated cell movement during morphogenesis. Curr. Opin. Genet. Dev. 2000, 10, 421–427. [Google Scholar] [CrossRef]
- Calvo-Garrido, J.; Carilla-Latorre, S.; Kubohara, Y.; Santos, N.; Mesquita, A.; Soldati, T.; Golstein, P.; Escalante, R. Autophagy in Dictyostelium: Genes and pathways, cell death and infection. Autophagy 2010, 6, 686–701. [Google Scholar] [CrossRef] [PubMed]
- Loomis, W.F. Cell signaling during development of Dictyoslteium. Dev. Biol. 2014, 391, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Loomis, W.F. Genetic control of morphogenesis in Dictyostelium. Dev. Biol. 2015, 402, 146–161. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.M.; Veltman, D.; Kay, R.R. Chemotaxis of a model organism: Progress with Dictyostelium. Curr. Opin. Cell Biol. 2015, 36, 7–12. [Google Scholar] [CrossRef]
- Bretschneider, T.; Othmer, H.G.; Weijer, C.J. Progress and perspectives in signal transduction, actin dynamics and movement at the cell and tissue level: Lessons from Dictyostelium. Interface Focus 2016, 6, 20160047. [Google Scholar] [CrossRef]
- Huber, R.J. Using the social amoeba Dictyostelium to study the functions of proteins linked to neuronal ceroid lipofuscinosis. J. Biomed. Sci. 2016, 23, 83. [Google Scholar] [CrossRef]
- Mesquita, A.; Cardenal-Muñoz, E.; Dominguez, E.; Muñoz-Braceras, S.; Nuñez-Corcuera, B.; Phillips, B.A.; Tábara, L.C.; Xiong, Q.; Coria, R.; Eichinger, L.; et al. Autophagy in Dictyostelium: Mechanisms, regulation and disease in a simple biomedical model. Autophagy 2017, 13, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Stuelten, C.H.; Parent, C.A.; Montell, D.J. Cell motility in cancer invasion and metastasis: Insights from simple model organisms. Nat. Rev. Cancer 2018, 18, 296–312. [Google Scholar] [CrossRef]
- Domínguez-Martín, E.; Hernández-Elvira, M.; Vincent, O.; Coria, R.; Escalante, R. Unfolding the Endoplasmic Reticulum of a Social Amoeba: Dictyostelium discoideum as a New Model for the Study of Endoplasmic Reticulum Stress. Cells 2018, 7, 56. [Google Scholar] [CrossRef]
- Dannat, K.; Tillner, J.; Winckler, T.; Weiss, M.; Eger, K.; Dingermann, T. Effects of medicinal compounds on the differentiation of the eukaryotic microorganism dictyostelium discoideum: Can this model be used as a screening test for reproductive toxicity in humans? Pharmazie 2003, 58, 204–210. [Google Scholar]
- Ludtmann, M.H.; Boeckeler, K.; Williams, R.S. Molecular pharmacology in a simple model system: Implicating MAP kinase and phosphoinositide signalling in bipolar disorder. Semin. Cell Dev. Biol. 2011, 22, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.C.; Williams, R.S. New experimental therapies for status epilepticus in preclinical development. Epilepsy Behav. 2015, 49, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Rzepnikowska, W.; Flis, K.; Muñoz-Braceras, S.; Menezes, R.; Escalante, R.; Zoladek, T. Yeast and other lower eukaryotic organisms for studies of Vps13 proteins in health and disease. Traffic 2017, 18, 711–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swart, A.L.; Harrison, C.F.; Eichinger, L.; Steinert, M.; Hilbi, H. Acanthamoeba and Dictyostelium as cellular models for Legionella infection. Front. Cell. Infect. Microbiol. 2018, 8, 61. [Google Scholar] [CrossRef]
- Kikuchi, H.; Saito, Y.; Komiya, J.; Takaya, Y.; Honma, S.; Nakahata, N.; Ito, A.; Oshima, Y. Furanodictine A and B: Amino sugar analogues produced by cellular slime mold Dictyostelium discoideum showing neuronal differentiation activity. J. Org. Chem. 2001, 66, 6982–6987. [Google Scholar] [CrossRef]
- Kikuchi, H.; Komiya, J.; Sito, Y.; Sekiya, J.; Honma, S.; Nakahata, N.; Oshima, Y. The isolation and synthesis of two novel N-acetyl glucosamine derivatives from Dictyostelium cellular slime molds which exhibit neurite outgrowth activity. Tetrahedron Lett. 2002, 43, 1477–1480. [Google Scholar] [CrossRef]
- Kikuchi, H.; Saito, Y.; Sekiya, J.; Okano, Y.; Saito, M.; Nakahata, H.; Kubohara, Y.; Oshima, Y. Isolation and synthesis of a new aromatic compound, brefelamide, from Dictyostelium cellular slime molds and its inhibitory effect on proliferation of astrocytoma cells. J. Org. Chem. 2005, 70, 8854–8858. [Google Scholar] [CrossRef]
- Kikuchi, H.; Oshima, Y.; Ichimura, A.; Gokan, N.; Hasegawa, A.; Hosaka, K.; Kubohara, Y. Anti-leukemic activities of Dictyostelium secondary metabolites: A novel aromatic metabolite, 4-methyl-5-n-pentylbenzene-1,3-diol, isolated from Dictyostelium mucoroides suppresses cell growth in human leukemia K562 and HL-60 cells. Life Sci. 2006, 80, 160–165. [Google Scholar] [CrossRef]
- Kikuchi, H.; Nakamura, K.; Kubohara, Y.; Gokan, N.; Hosaka, K.; Maeda, Y.; Oshima, Y. Dihydrodictyopyrone A and C: New members of dictyopyrone family isolated from Dictyostelium cellular slime molds. Tetrahedron Lett. 2007, 48, 5905–5909. [Google Scholar] [CrossRef]
- Kikuchi, H.; Ishiko, S.; Nakamura, K.; Kubohara, Y.; Oshima, Y. Novel prenylated and geranylated aromatic compounds isolated from Polysphondylium cellular slime molds. Tetrahedron 2010, 66, 6000–6007. [Google Scholar] [CrossRef]
- Kikuchi, H.; Matsuo, Y.; Katou, Y.; Kubohara, Y.; Oshima, Y. Isolation, synthesis and biological activity of biphenyl and m-terphenyl-type compounds from Dictyostelium cellular slime molds. Tetrahedron 2012, 68, 8884–8889. [Google Scholar] [CrossRef]
- Kikuchi, H.; Kubohara, Y.; Nguyen, V.H.; Katou, Y.; Oshima, Y. Novel chlorinated dibenzofurans Isolated from the cellular slime mold, Polysphondylium filamentosum and their biological activities. Bioorg. Med. Chem. 2013, 21, 4628–4633. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, H.; Ito, I.; Takahashi, K.; Ishigaki, H.; Iizumi, K.; Kubohara, Y.; Oshima, Y. Isolation, synthesis and biological activity of chlorinated alkylresorcinols from Dictyostelium cellular slime molds. J. Nat. Prod. 2017, 80, 2716–2722. [Google Scholar] [CrossRef] [PubMed]
- Barnett, R.; Stallforth, P. Natural products form social amoebae. Biol. Chem. 2018, 24, 4202–4214. [Google Scholar]
- Sucgang, R.; Kuo, A.; Tian, X.; Salerno, W.; Parikh, A.; Feasley, C.L.; Dalin, E.; Tu, H.; Huang, E.; Barry, K.; et al. Comparative genomics of the social amoebae Dictyostelium discoideum and Dictyostelium purpureum. Genome Biol. 2011, 12, R20. [Google Scholar] [CrossRef] [PubMed]
- Morris, H.R.; Taylor, G.W.; Masento, M.S.; Jermyn, K.A.; Kay, R.R. Chemical structure of the morphogen differentiation inducing factor from Dictyostelium discoideum. Nature 1987, 328, 811–814. [Google Scholar] [CrossRef] [PubMed]
- Morris, H.R.; Masento, M.S.; Taylor, G.W.; Jermyn, K.A.; Kay, R.R. Structure elucidation of two differentiation inducing factors (DIF-2 and DIF-3) from the cellular slime mould Dictyostelium discoideum. Biochem. J. 1988, 249, 903–906. [Google Scholar] [CrossRef] [PubMed]
- Masento, M.S.; Morris, H.R.; Taylor, G.W.; Johnson, S.J.; Skapski, A.C.; Kay, R.R. Differentiation-inducing factor from the slime mould Dictyostelium discoideum and its analogues. Biochem. J. 1988, 256, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Kay, R.R.; Berks, M.; Traynor, D. Morphogen hunting in Dictyostelium. Development 1989, 107, 81–90. [Google Scholar] [PubMed]
- Kay, R.R.; Flatman, P.; Thompson, C.R.L. DIF signalling and cell fate. Semin. Cell Dev. Biol. 1999, 10, 577–585. [Google Scholar] [CrossRef]
- Wurster, B.; Kay, R.R. New roles for DIF? Effects on early development in Dictyostelium. Dev. Biol. 1990, 140, 189–195. [Google Scholar] [CrossRef]
- Morandini, P.; Offer, J.; Traynor, D.; Nayler, O.; Neuhaus, D.; Taylor, G.W.; Kay, R.R. The proximal pathway of metabolism of the chlorinated signal molecule differentiation-inducing factor-1 (DIF-1) in the cellular slime mould Dictyostelium. Biochem. J. 1995, 306, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Cornillon, S.; Foa, C.; Davoust, J.; Buonavista, N.; Gross, J.D.; Golstein, P. Programmed cell death in Dictyostelium. J. Cell Sci. 1994, 107, 2691–2704. [Google Scholar] [PubMed]
- Giusti, C.; Tresse, E.; Luciani, M.F.; Golstein, P. Autophagic cell death: Analysis in Dictyostelium. Biochim Biophys Acta 2009, 1793, 1422–1431. [Google Scholar] [CrossRef]
- Luciani, M.F.; Kubohara, Y.; Kikuchi, H.; Oshima, Y.; Golstein, P. Autophagic or necrotic cell death triggered by distinct motifs of the differentiation factor DIF-1. Cell Death Differ. 2009, 16, 564–570. [Google Scholar] [CrossRef]
- Luciani, M.F.; Giusti, C.; Harms, B.; Oshima, Y.; Kikuchi, H.; Kubohara, Y.; Golstein, P. Atg1 allows second-signaled autophagic cell death in Dictyostelium. Autophagy 2011, 7, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Giusti, C.; Luciani, M.F.; Ravens, S.; Gillet, A.; Golstein, P. Autophagic cell death in Dictyostelium requires the receptor histidine kinase DhkM. Mol. Biol. Cell 2010, 21, 1825–1835. [Google Scholar] [CrossRef]
- Kuwayama, H.; Kubohara, Y. Differentiation-inducing factor-1 and -2 function also as modulators for Dictyostelium chemotaxis. PLoS ONE 2009, 4, e6658. [Google Scholar] [CrossRef]
- Kuwayama, H.; Kikuchi, H.; Oshima, Y.; Kubohara, Y. Artificial compounds differentially control Dictyostelium chemotaxis and cell differentiation. Cell Struct. Funct. 2011, 36, 21–26. [Google Scholar] [CrossRef]
- Kuwayama, H.; Kubohara, Y. Differentiation-inducing factor 2 modulates chemotaxis via the histidine kinase DhkC-dependent pathway in Dictyostelium discoideum. FEBS Lett. 2016, 590, 760–768. [Google Scholar] [CrossRef]
- Insall, R.; Kay, R.R. A specific DIF binding protein in Dictyostelium. EMBO J. 1990, 9, 3323–3328. [Google Scholar] [CrossRef] [PubMed]
- Kubohara, Y.; Maeda, M.; Okamoto, K. Analysis of the maturation process of prestalk cells in Dictyostelium discoideum. Exp. Cell Res. 1993, 207, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Kubohara, Y.; Okamoto, K. Cytoplasmic Ca2+ and H+ concentrations determine cell fate in Dictyostelium discoideum. FASEB J. 1994, 8, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.H.; Schaap, P. The prokaryote messenger c-di-GMP triggers stalk cell differentiation in Dictyostelium. Nature 2012, 488, 680–683. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Luciani, M.F.; Giusti, C.; Golstein, P. c-di-GMP induction of Dictyostelium cell death requires the polyketide DIF-1. Mol. Biol. Cell 2015, 26, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Kuwayama, H.; Kikuchi, H.; Oshima, Y.; Kubohara, Y. Glutathione S-transferase 4 is a putative DIF-binding protein that regulates the size of fruiting bodies in Dictyostelium discoideum. Biochem. Biophys. Rep. 2016, 8, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Kubohara, Y.; Kikuchi, H.; Nguyen, V.H.; Kuwayama, H.; Oshima, Y. Evidence that differentiation-inducing factor-1 controls chemotaxis and cell differentiation, at least in part, via mitochondria in D. discoideum. Biol. Open 2017, 6, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Kay, R.R. The biosynthesis of differentiation-inducing factor, a chlorinated signal molecule regulating Dictyostelium development. J. Biol. Chem. 1998, 273, 2669–2675. [Google Scholar] [CrossRef]
- Oka, H.; Asahi, K.; Morishima, H.; Sanada, M.; Shiratori, K.; Iimura, Y.; Sakurai, T.; Uzawa, J.; Iwadare, S.; Takahashi, N. Differanisole A, a new differentiation inducing substance. J. Antibiot. 1985, 38, 1100–1102. [Google Scholar] [CrossRef]
- Kubohara, Y.; Okamoto, K.; Tanaka, Y.; Asahi, K.; Sakurai, A.; Takahashi, N. Differanisole A, an inducer of the differentiation of Friend leukemic cells, induces stalk cell differentiation in Dictyostelium discoideum. FEBS Lett. 1993, 322, 73–75. [Google Scholar] [CrossRef]
- Asahi, K.; Sakurai, A.; Takahashi, N.; Kubohara, Y.; Okamoto, K.; Tanaka, Y. DIF-1, morphogen of Dictyostelium discoideum, induces the erythroid differentiation in murine and human leukemia cells. Biochem. Biophys. Res. Commun. 1995, 208, 1036–1039. [Google Scholar] [CrossRef]
- Kubohara, Y.; Saito, Y.; Tatemoto, K. Differentiation-inducing factor of D. discoideum raises intracellular calcium concentration and suppresses cell growth in rat pancreatic AR42J cells. FEBS Lett. 1995, 359, 119–122. [Google Scholar] [CrossRef]
- Kubohara, Y. DIF-1, putative morphogen of D. discoideum, suppresses cell growth and promotes retinoic acid-induced cell differentiation in HL-60. Biochem. Biophys. Res. Commun. 1997, 236, 418–422. [Google Scholar] [CrossRef]
- Kubohara, Y. Effects of differentiation-inducing factors (DIFs) of Dictyostelium discoideum on the human leukemia K562 cells: DIF-3 is the most potent anti-leukemic agent. Eur. J. Pharmacol. 1999, 381, 57–62. [Google Scholar] [CrossRef]
- Kanai, M.; Konda, Y.; Nakajima, T.; Izumi, Y.; Nanakin, A.; Kanda, N.; Kubohara, Y.; Chiba, T. Differentiation-inducing factor-1 (DIF-1) inhibits STAT3 activity involved in gastric cancer cell proliferation via MEK-ERK dependent pathway. Oncogene 2003, 22, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Yanaga, F.; Taba, Y.; Miwa, Y.; Kubohara, Y.; Watanabe, Y.; Hirata, M.; Morimoto, S.; Sasaguri, T. Dictyostelium differentiation-inducing factor-3 activates glycogen synthase kinase-3b and degrades cyclin D1 in mammalian cells. J. Biol. Chem. 2003, 278, 9663–9670. [Google Scholar] [CrossRef] [PubMed]
- Kubohara, Y.; Kimura, C.; Tatemoto, K. Putative morphogen, DIF, of Dictyostelium discoideum induces apoptosis in rat pancreatic AR42J cells. Develop. Growth Differ. 1995, 37, 711–716. [Google Scholar] [CrossRef]
- Fujimaki, S.; Kubohara, Y.; Kobayashi, I.; Kojima, I. Caspase-independent apoptosis induced by differentiation-inducing factor of Dictyostelium discoideum in INS-1 cells. Eur. J. Pharmacol. 2001, 421, 93–100. [Google Scholar] [CrossRef]
- Dubois, A.; Ginet, C.; Furstoss, N.; Belaid, A.; Hamouda, M.A.; El Manaa, W.; Cluzeau, T.; Marchetti, S.; Ricci, J.E.; Jacquel, A.; et al. Differentiation inducing factor 3 mediates its anti-leukemic effect through ROS-dependent DRP1-mediated mitochondrial fission and induction of caspase-independent cell death. Oncotarget 2016, 7, 26120–26136. [Google Scholar] [CrossRef] [Green Version]
- Miwa, Y.; Sasaguri, T.; Kosaka, C.; Taba, Y.; Ishida, A.; Abumiya, T.; Kubohara, Y. DIF-1, a morphogen of Dictyostelium, induces G1 arrest and differentiation of vascular smooth muscle cells. Circ. Res. 2000, 86, 68–75. [Google Scholar] [CrossRef]
- Kubohara, Y.; Kikuchi, H.; Oshima, Y. Exploitation of the derivatives of Dictyostelium differentiation-inducing factor-1, which promote glucose consumption in mammalian cells. Life Sci. 2008, 83, 608–612. [Google Scholar] [CrossRef] [PubMed]
- Kubohara, Y.; Komachi, M.; Homma, Y.; Kikuchi, H.; Oshima, Y. Derivatives of Dictyostelium differentiation-inducing factors inhibit lysophosphatidic acid–stimulated migration of murine osteosarcoma LM8 cells. Biochem. Biophys. Res. Commun. 2015, 463, 800–805. [Google Scholar] [CrossRef] [PubMed]
- Gokan, N.; Kikuchi, H.; Nakamura, K.; Oshima, Y.; Hosaka, K.; Kubohara, Y. Structural requirements of Dictyostelium differentiation-inducing factors for their stalk-cell-inducing activity in Dictyostelium cells and anti-proliferative activity in K562 human leukemic cells. Biochem. Pharmacol. 2005, 70, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Oladimeji, P.; Kubohara, Y.; Kikuchi, H.; Oshima, Y.; Rusch, C.; Skerl, R.; Diakonova, M. A derivative of differentiation-inducing factor-3 inhibits PAK1 activity and breast cancer cell proliferation. Int. J. Cancer Clinic. Res. 2015, 2, 1–6. [Google Scholar] [CrossRef]
- Takahashi, K.; Kikuchi, H.; Nguyen, V.H.; Oshima, Y.; Ishigaki, H.; Nakajima-Shimada, J.; Kubohara, Y. Biological activities of novel derivatives of differentiation-inducing factor-3 from Dictyostelium discoideum. Biol. Pharm. Bull. 2017, 40, 1941–1947. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Murata, T.; Tagawa, T.; Takahashi, K.; Ishikawa, R.; Abe, Y.; Hosaka, K.; Kubohara, Y. Calmodulin-dependent cyclic nucleotide phosphodiesterase (PDE1) is a pharmacological target of differentiation-inducing factor-1, an anti-tumor agent isolated from Dictyostelium. Cancer Res. 2004, 64, 2568–2571. [Google Scholar] [CrossRef] [PubMed]
- Kubohara, Y.; Kikuchi, H.; Matsuo, Y.; Oshima, Y.; Homma, Y. Mitochondria are the target organelle of differentiation-inducing factor-3, an anti-tumor agent isolated from Dictyostelium discoideum. PLoS ONE 2013, 8, e72118. [Google Scholar] [CrossRef]
- Kubohara, Y.; Hosaka, K. The putative morphogen, DIF-1, of Dictyostelium discoideum activates Akt/PKB in human leukemia K562 cells. Biochem. Biophys. Res. Commun. 1999, 263, 790–796. [Google Scholar] [CrossRef]
- Akaishi, E.; Narita, T.; Kawai, S.; Miwa, Y.; Sasaguri, T.; Hosaka, K.; Kubohara, Y. Differentiation-inducing factor-1-induced growth arrest of K562 leukemia cells involves the reduction of ERK1/2 activity. Eur. J. Pharmacol. 2004, 485, 21–29. [Google Scholar] [CrossRef]
- Takahashi-Yanaga, F.; Sasaguri, T. Drug development targeting the glycogen synthase kinase-3 β(GSK-3β)-mediated signal transduction pathway: Inhibitors of the Wnt/β-catenin signaling pathway as novel anticancer drugs. J. Pharmacol. Sci. 2009, 109, 179–183. [Google Scholar] [CrossRef]
- Yasmin, T.; Takahashi-Yanaga, F.; Mori, J.; Miwa, Y.; Hirata, M.; Watanabe, Y.; Morimoto, S.; Sasaguri, T. Differentiation-inducing factor-1 suppresses gene expression of cyclin D1 in tumor cells. Biochem. Biophys. Res. Commun. 2005, 338, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Arioka, M.; Takahashi-Yanaga, F.; Kubo, M.; Igawa, K.; Tomooka, K.; Sasaguri, T. Anti-tumor effects of differentiation-inducing factor-1 in malignant melanoma: GSK-3-mediated inhibition of cell proliferation and GSK-3-independent suppression of cell migration and invasion. Biochem. Pharmacol. 2017, 138, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Kubokura, N.; Takahashi-Yanaga, F.; Arioka, M.; Yoshihara, T.; Igawa, K.; Tomooka, K.; Morimoto, S.; Nakatsu, Y.; Tsuzuki, T.; Nakabeppu, Y.; et al. Differentiation-inducing factor-3 inhibits intestinal tumor growth in vitro and in vivo. J. Pharmacol. Sci. 2015, 127, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Omata, W.; Shibata, H.; Nagasawa, M.; Kojima, I.; Kikuchi, H.; Oshima, Y.; Hosaka, K.; Kubohara, Y. Dictyostelium differentiation-inducing factor-1 induces glucose transporter 1 translocation and promotes glucose uptake in mammalian cells. FEBS J. 2007, 274, 3392–3404. [Google Scholar] [CrossRef] [PubMed]
- Kawaharada, R.; Nakamura, A.; Takahashi, K.; Kikuchi, H.; Oshima, Y.; Kubohara, Y. Oral administration of Dictyostelium differentiation-inducing factor 1 lowers blood glucose levels in streptozotocin-induced diabetic rats. Life Sci. 2016, 155, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Seya, K.; Kanemaru, K.; Matsuki, M.; Hongo, K.; Kitahara, H.; Kikuchi, H.; Oshima, Y.; Kubohara, Y.; Okumura, K.; Motomura, S.; et al. Br-DIF-1 accelerates dimethyl sulphoxide-induced differentiation of P19CL6 embryonic carcinoma cells into cardiomyocytes. Br. J. Pharmacol. 2012, 165, 870–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzaki, E.; Takahashi-Yanaga, F.; Miwa, Y.; Hirata, M.; Watanabe, Y.; Sato, N.; Morimoto, S.; Hirofuji, T.; Maeda, K.; Sasaguri, T. Differentiation-inducing factor-1 alters canonical Wnt signaling and suppresses alkaline phosphatase expression in osteoblast-like cell lines. J. Bone Miner. Res. 2006, 21, 1307–1316. [Google Scholar] [CrossRef]
- Kubohara, Y.; Hanaoka, Y.; Akaishi, E.; Kobayashi, H.; Maeda, M.; Hosaka, K. DIF-1, an anti-tumor agent found in Dictyostelium discoideum, inhibits progesterone-induced oocyte maturation in Xenopus laevis. Eur. J. Pharmacol. 2003, 460, 93–98. [Google Scholar] [CrossRef]
- Takahashi, K.; Murakami, M.; Hosaka, K.; Kikuchi, H.; Oshima, Y.; Kubohara, Y. Regulation of IL-2 production in Jurkat cells by Dictyostelium-derived factors. Life Sci. 2009, 85, 438–443. [Google Scholar] [CrossRef]
- Takahashi, K.; Murakami, M.; Kikuchi, H.; Oshima, Y.; Kubohara, Y. Derivatives of Dictyostelium differentiation-inducing factors promote mitogen-activated IL-2 production via AP-1 in Jurkat cells. Life Sci. 2011, 88, 480–485. [Google Scholar] [CrossRef]
- Clayton, J. Chagas disease 101. Nature 2010, 465, S4–S5. [Google Scholar] [CrossRef] [PubMed]
- Cançado, J.R. Long term evaluation of etiological treatment of Chagas disease with benznidazole. Rev. Inst. Med. Trop. São Paulo 2002, 44, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coura, J.R.; de Castro, S.L. A critical review of Chagas disease chemotherapy. Mem. Inst. Oswaldo Cruz 2002, 97, 3–24. [Google Scholar] [CrossRef]
- Nakajima-Shimada, J.; Hatabu, T.; Hosoi, Y.; Onizuka, Y.; Kikuchi, H.; Oshima, Y.; Kubohara, Y. Derivatives of Dictyostelium discoideum differentiation-inducing factor-3 suppress the activities of Trypanosoma cruzi in vitro and in vivo. Biochem. Pharmacol. 2013, 85, 1603–1610. [Google Scholar] [CrossRef] [PubMed]
- Haass, C.; Schlossmacher, M.G.; Hung, A.Y.; Vigo-Pelfrey, C.; Mellon, A.; Ostaszewski, B.L.; Lieberburg, I.; Koo, E.H.; Schenk, D.; Teplow, D.B.; et al. Amyloid beta-peptide is produced by cultured cells during normal metabolism. Nature 1992, 359, 322–325. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J. The relationship between amyloid and tau. J. Mol. Neurosci. 2003, 20, 203–206. [Google Scholar] [CrossRef]
- Myre, M.A.; Washicosky, K.; Moir, R.D.; Tesco, G.; Tanzi, R.E.; Wasco, W. Reduced amyloidogenic processing of the amyloid beta-protein precursor by the small-molecule Differentiation Inducing Factor-1. Cell Signal. 2009, 21, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Kanno, T.; Kubohara, Y.; Goto, F.; Takata, K.; Hayashi, K. NMDA-receptor-dependent and -independent cytotoxic effects of D. discoideum differentiation-inducing factor-1 on rat cortical neurons. Develop. Growth Differ. 2001, 43, 709–716. [Google Scholar] [CrossRef]
- Matsuda, T.; Takahashi-Yanaga, F.; Yoshihara, T.; Maenaka, K.; Watanabe, Y.; Miwa, Y.; Morimoto, S.; Kubohara, Y.; Hirata, M.; Sasaguri, T. Dictyostelium differentiation-inducing factor-1 binds to mitochondrial malate dehydrogenase and inhibits its activity. J. Pharmacol. Sci. 2010, 112, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Takaya, Y.; Kikuchi, H.; Terui, Y.; Komiya, J.; Furukawa, K.I.; Seya, K.; Motomura, S.; Ito, A.; Oshima, Y. Novel acyl a-pyronoids, dictyopyrone A, B and C, from Dictyostelium cellular slime molds. J. Org. Chem. 2000, 65, 985–989. [Google Scholar] [CrossRef]
- Kikuchi, H.; Sasaki, K.; Sekiya, J.; Maeda, Y.; Amagai, A.; Kubohara, Y.; Oshima, Y. Structural requirements of dictyopyrones isolated from Dictyostelium spp. in the regulation of Dictyostelium development and in anti-leukemic activity. Bioorg. Med. Chem. 2004, 12, 3203–3214. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.H.; Kikuchi, H.; Sasaki, H.; Iizumi, K.; Kubohara, Y.; Oshima, Y. Production of novel bispyrone metabolites in the cellular slime mold Dictyostelium giganteum induced by zinc(II) ion. Tetrahedron 2017, 73, 583–588. [Google Scholar] [CrossRef]
- Arai, A.; Goto, Y.; Hasegawa, A.; Hosaka, K.; Kikuchi, H.; Oshima, Y.; Tanaka, S.; Kubohara, Y. Dictyopyrones, novel a-pyronoids isolated from Dictyostelium spp., promote stalk cell differentiation in Dictyostelium discoideum. Differentiation 2005, 73, 377–384. [Google Scholar]
- Mareyala, H.B.; Baseeruddin, M.; Reddy, K.S. Formala synthesis of furanodictine B from d-glucose. Tetrahedron Asymmetry 2004, 15, 3457–3460. [Google Scholar] [CrossRef]
- Matsuura, D.; Mitsui, T.; Sengoku, T.; Takahashi, M.; Yoda, H. Stereo-divergent synthesis of new amino sugars, furanodictines A and B, starting from d-glucuronolactone. Tetrahedron 2008, 64, 11686–11696. [Google Scholar] [CrossRef]
- Ogata, M.; Hattori, T.; Takeuchi, R.; Usui, T. Novel and facile synthesis of furanodictines A and B based on transformation of 2-acetamido-2-deoxy-d-glucose into 3,6-anhydro hexofuranoses. Carbohydr. Res. 2010, 345, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Honma, S.; Kouno, K.; Takasaka, S.; Mitazaki, S.; Abe, S.; Kikuchi, H.; Oshima, Y.; Yoshida, M. Effect of brefelamide on proliferation of 1321N1 human astrocytoma cells induced by glial cell line-derived neurotrophic factor. Pharmazie 2018, 73, 22–28. [Google Scholar]
- Zhang, J.; Yamada, O.; Kida, S.; Matsushita, Y.; Murase, S.; Hattori, T.; Kubohara, Y.; Kikuchi, H.; Oshima, Y. Identification of brefelamide as a novel inhibitor of osteopontin that suppresses invasion of A549 lung cancer cells. Oncol. Rep. 2016, 36, 2357–2364. [Google Scholar] [CrossRef]
- Pascapurnama, D.N.; Labayo, H.K.M.; Dapat, I.; Nagarajegowda, D.D.; Zhao, J.; Zhang, J.; Yamada, O.; Kikuchi, H.; Egawa, S.; Oshima, Y.; et al. Induction of osteopontin by dengue virus-3 infection in THP-1 cells—Inhibition of the synthesis by brefelamide and its derivative. Front. Microbiol. 2017, 8, 521. [Google Scholar] [CrossRef]
- Saito, T.; Taylor, G.W.; Yang, J.C.; Neuhaus, D.; Stetsenko, D.; Kato, A.; Kay, R.R. Identification of new differentiation inducing factors from Dictyostelium discoideum. Biochim. Biophys. Acta 2006, 1760, 754–761. [Google Scholar] [CrossRef]
- Murata, C.; Ogura, T.; Narita, S.; Kondo, A.P.; Iwasaki, N.; Saito, T.; Usuki, T. Synthesis and SAR of 4-methyl5-pentylbenzene-1,3-diol (MPBD), produced by Dictyostelium discoideum. Bioorg. Med. Chem. Lett. 2016, 26, 1428–1433. [Google Scholar] [CrossRef] [PubMed]
- Sawada, T.; Aono, M.; Asakawa, S.; Ito, A.; Awano, K. Structure determination and total synthesis of a novel antibacterial substance, AB0022A, produced by a cellular slime mold. J. Antibiot. 2000, 53, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Kikuchi, H.; Ogura, M.; Homma, M.K.; Oshima, Y.; Homma, Y. Weight loss by Ppc-1, a novel small molecule mitochondrial uncoupler derived from slime mold. PLoS ONE 2015, 10, e0117088. [Google Scholar] [CrossRef] [PubMed]
- Ogura, M.; Kikuchi, H.; Suzuki, T.; Yamaki, J.; Homma, M.K.; Oshima, Y.; Homma, Y. Prenylated quinolinecarboxylic acid derivative suppresses immune response through inhibition of PAK2. Biochem. Pharmacol. 2016, 105, 55–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogura, M.; Kikuchi, H.; Shakespear, N.B.; Suzuki, T.; Yamaki, J.; Homma, M.K.; Oshima, Y.; Homma, Y. Prenylated quinolinecarboxylic acid derivative prevents neuronal cell death through inhibition of MKK4. Biochem. Pharmacol. 2018, in press. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DIF Species Examined | Target of DIF | Biological Activities | Reference |
|---|---|---|---|
| DIF-1, DIF-3 | PDE1 | Direct inhibition of PDE1 activity | [76] |
| DIF-1 | mMDH | Direct inhibition of mMDH activity | [100] |
| DIF-1, DIF-3, & their derivatives | Mitochondria | Uncoupling of mitochondrial activity | [72,77] |
| DIF-3 derivatives (e.g., DIF-3(+1)) | PAK1 | Direct inhibition of PAK1 activity | [74] |
| Compound | Source Organism | Biological Activities | Reference |
|---|---|---|---|
| Dictyopyrone A | D. discoideum D. rhizoposium D. longosporum | Promotion of morphogenesis & stalk cell differentiation, & inhibition of spore formation in D. discoideum | [100,101,103] |
| Anti-proliferative activity in K562 cells | [101] | ||
| Dictyopyrone B | D. discoideum D. rhizoposium D. magnum D. longosporum | Promotion of morphogenesis & stalk cell differentiation, & inhibition of spore formation in D. discoideum | [100,101,103] |
| Dictyopyrone C | D. longosporum | Promotion of morphogenesis in D. discoideum | [100,101] |
| Anti-proliferative activity in K562 cells | [101] | ||
| Dictyopyrone D | D. magnum | Promotion of morphogenesis in D. discoideum | [101] |
| Dihydrodictyopyrones A & C | D. firmibasis | N.D. | [29] |
| Dictyobispyrones B & E | D. giganteum | N.D. | [102] |
| Furanodictine A | D. discoideum | Promotion of NGF-induced neurite formation in PC-12 cells | [25] |
| Furanodictine B | D. discoideum | Induction of neurite formation in PC-12 cells | [25] |
| Promotion of NGF-induced neurite formation in PC-12 cells | [25] | ||
| Dictyoglucosamine A | D. purpureum | Induction of neurite formation in PC-12 cells | [26] |
| Dictyoglucosamine B | D. discoideum | Induction of neurite formation in PC-12 cells | [26] |
| Brefelamide (& derivatives) | D. brefeldianum D. giganteum | Anti-proliferative activity in 1321N1 cells | [27,107] |
| Anti-proliferative & anti-metastatic activities in A562 cells | [108] | ||
| Inhibition of GDNF secretion in astrocytoma cells | [27,107] | ||
| Anti-dengue viral activity | [109] | ||
| MPBD (& derivatives) | D. discoideum D. mucoroides | Promotion of cell differentiation in D. discoideum | [110] |
| Anti-proliferative activity in K562 and HL-60 cells | [28] | ||
| Antimicrobial activities vs. E. coli and B. subtilis | [111] | ||
| Monochasiol A | D. monochasioides | Inhibition of IL-2 production in Jurkat T cells | [33] |
| Monochasiols B–H | D. monochasioides | N.D. | [33] |
| AB0022A | D. purpureum | Antimicrobial activities vs. Gram-positive bacteria | [112] |
| Pf-1 | P. filamentosum | Anti-proliferative activity in K562 and HL-60 cells | [32] |
| Pf-2 | P. filamentosum | Stalk-cell-inducing activity in D. discoideum | [32] |
| Pt-1 | P. tenuissimum | Anti-proliferative activity in K562 cells | [30] |
| Pt-2, Pt-3, Pt-4 | P. tenuissimum | N.D. | [30] |
| Pt-5 | P. tenuissimum | Anti-proliferative activity in K562 cells | [30] |
| Ppc-1 (& derivatives) | P. pseudo-candidum | Anti-proliferative activity in K562 and HeLa cells | [30] |
| Promotion of glucose consumption in 3T3-L1 cells & mitochondrial uncoupling | [30,112] | ||
| Inhibition of IL-2 production in Jurkat T cells | [114] | ||
| Neuroprotective activities in vitro and in vivo | [115] | ||
| Dictyobiphenyls A & B | D. polycephalum | N.D. | [31] |
| Dictyoterphenyl A | D. polycephalum | Anti-proliferative activity in K562, HeLa and LM8 cells | [31] |
| Dictyoterphenyl B | D. polycephalum | N.D. | [31] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubohara, Y.; Kikuchi, H. Dictyostelium: An Important Source of Structural and Functional Diversity in Drug Discovery. Cells 2019, 8, 6. https://doi.org/10.3390/cells8010006
Kubohara Y, Kikuchi H. Dictyostelium: An Important Source of Structural and Functional Diversity in Drug Discovery. Cells. 2019; 8(1):6. https://doi.org/10.3390/cells8010006
Chicago/Turabian StyleKubohara, Yuzuru, and Haruhisa Kikuchi. 2019. "Dictyostelium: An Important Source of Structural and Functional Diversity in Drug Discovery" Cells 8, no. 1: 6. https://doi.org/10.3390/cells8010006
APA StyleKubohara, Y., & Kikuchi, H. (2019). Dictyostelium: An Important Source of Structural and Functional Diversity in Drug Discovery. Cells, 8(1), 6. https://doi.org/10.3390/cells8010006