Age-Related Alterations Affecting the Chondrogenic Differentiation of Synovial Fluid Mesenchymal Stromal Cells in an Equine Model



Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Equine SfMSCs
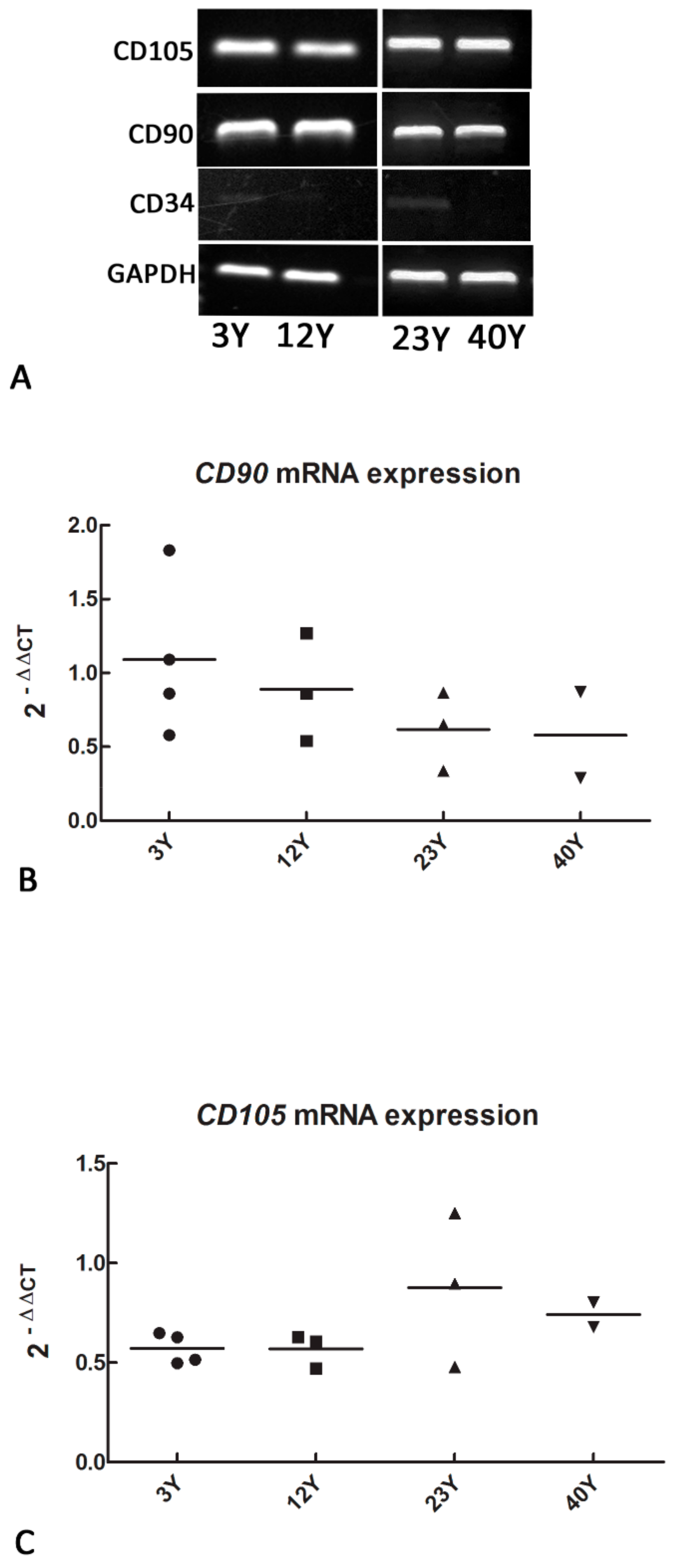
2.2. mRNA Expression of Mesenchymal and Self-Renewal Markers by RT-PCR and Real-Time PCR
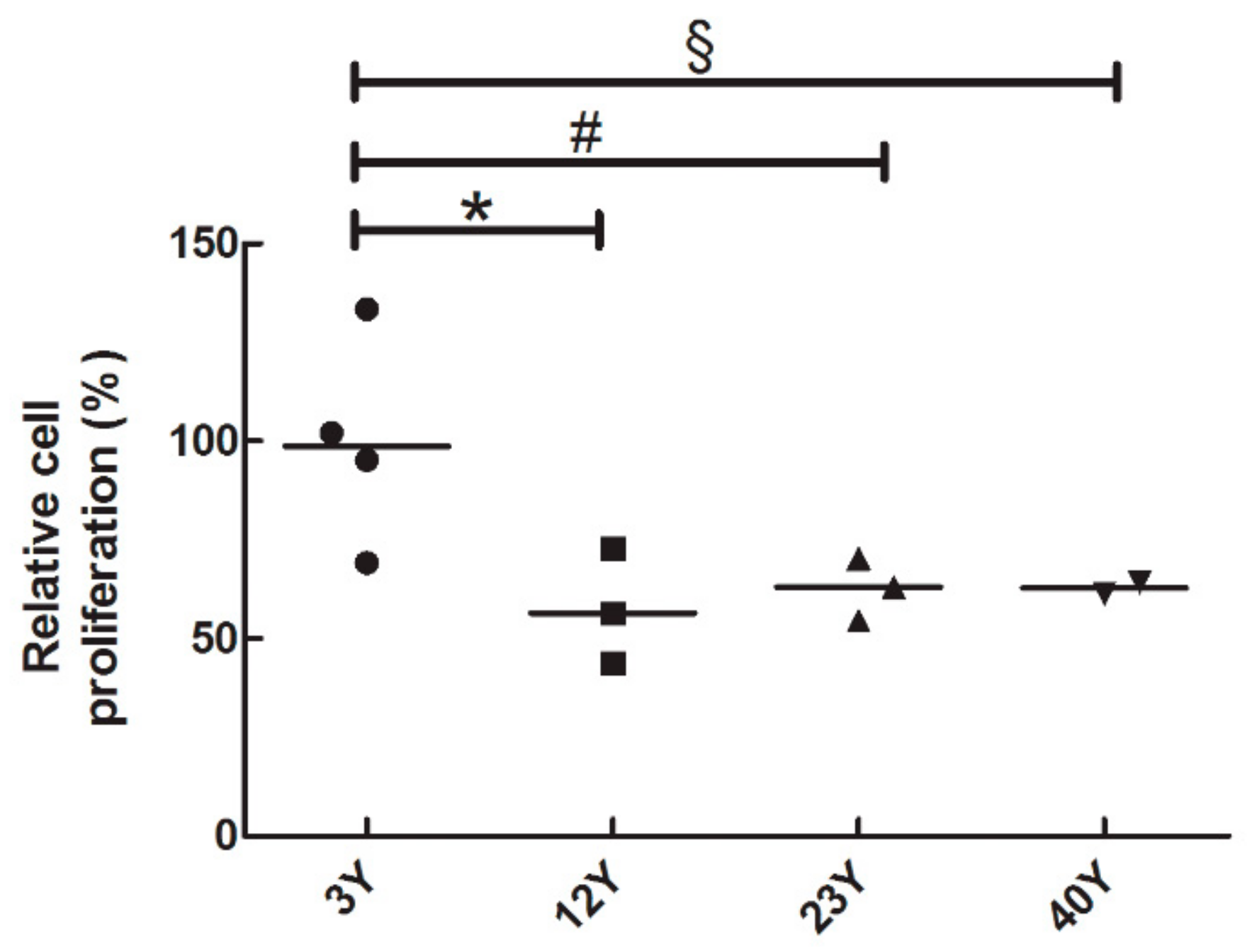
2.3. BrdU Proliferation Assay
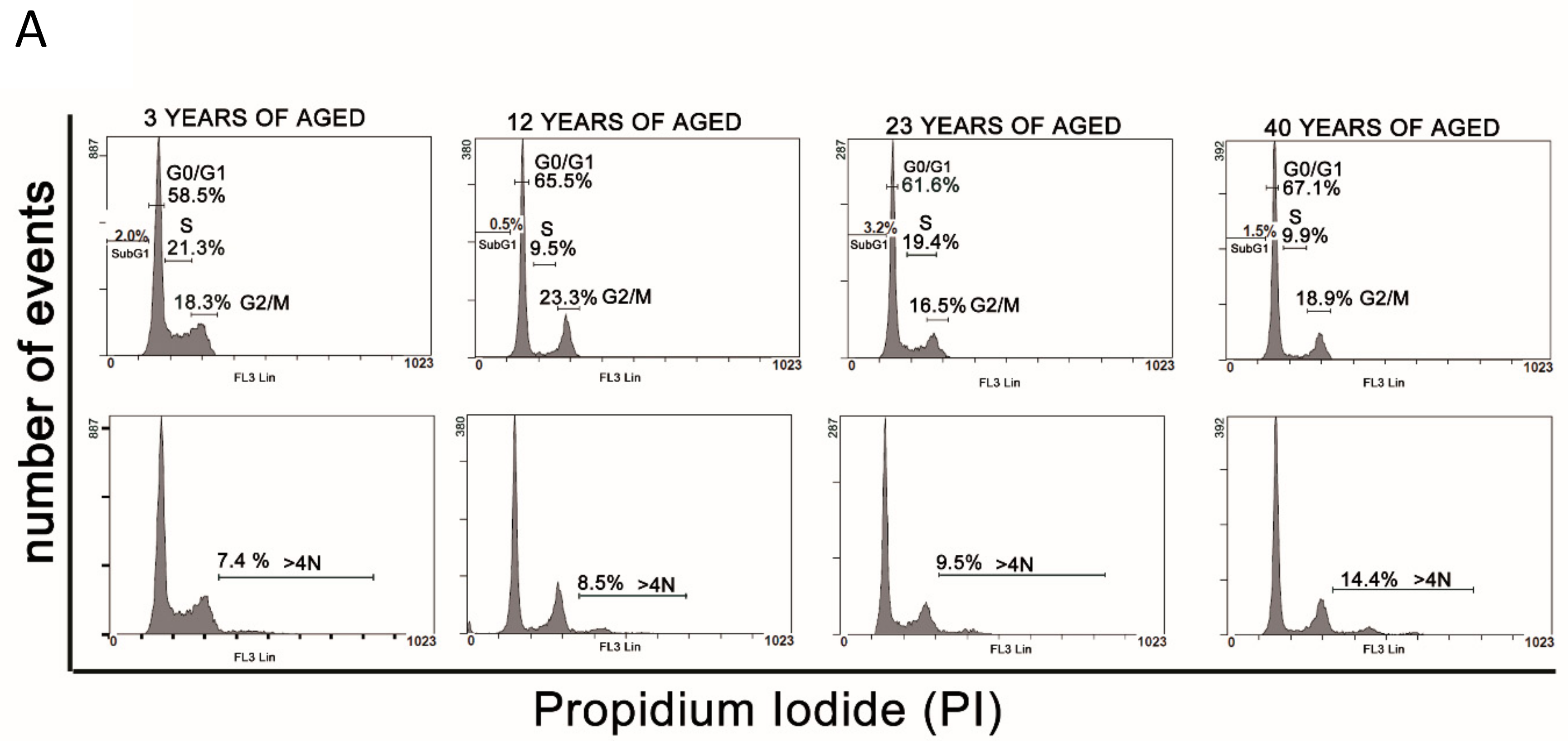
2.4. Cell Cycle Analysis
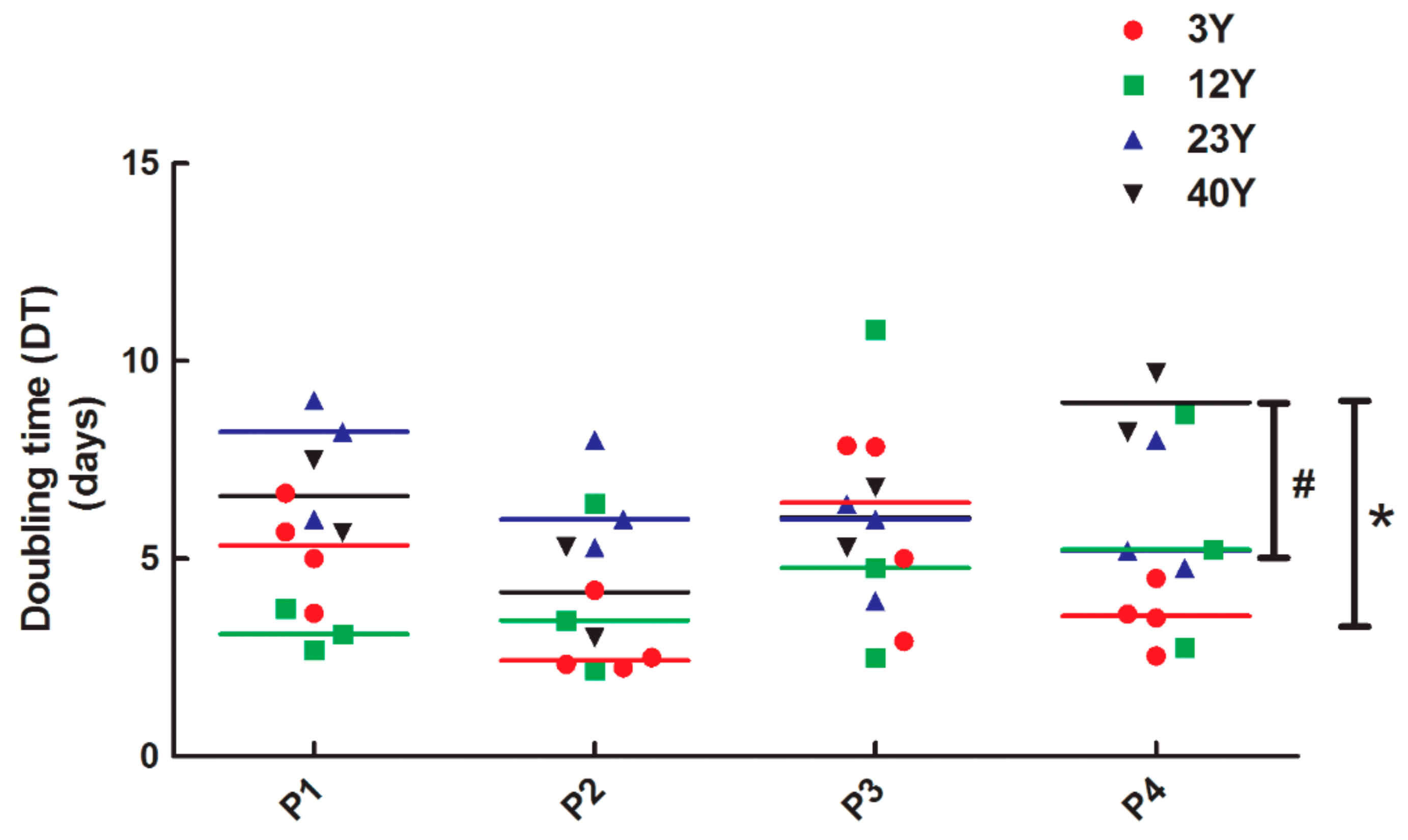
2.5. Doubling Time Assay
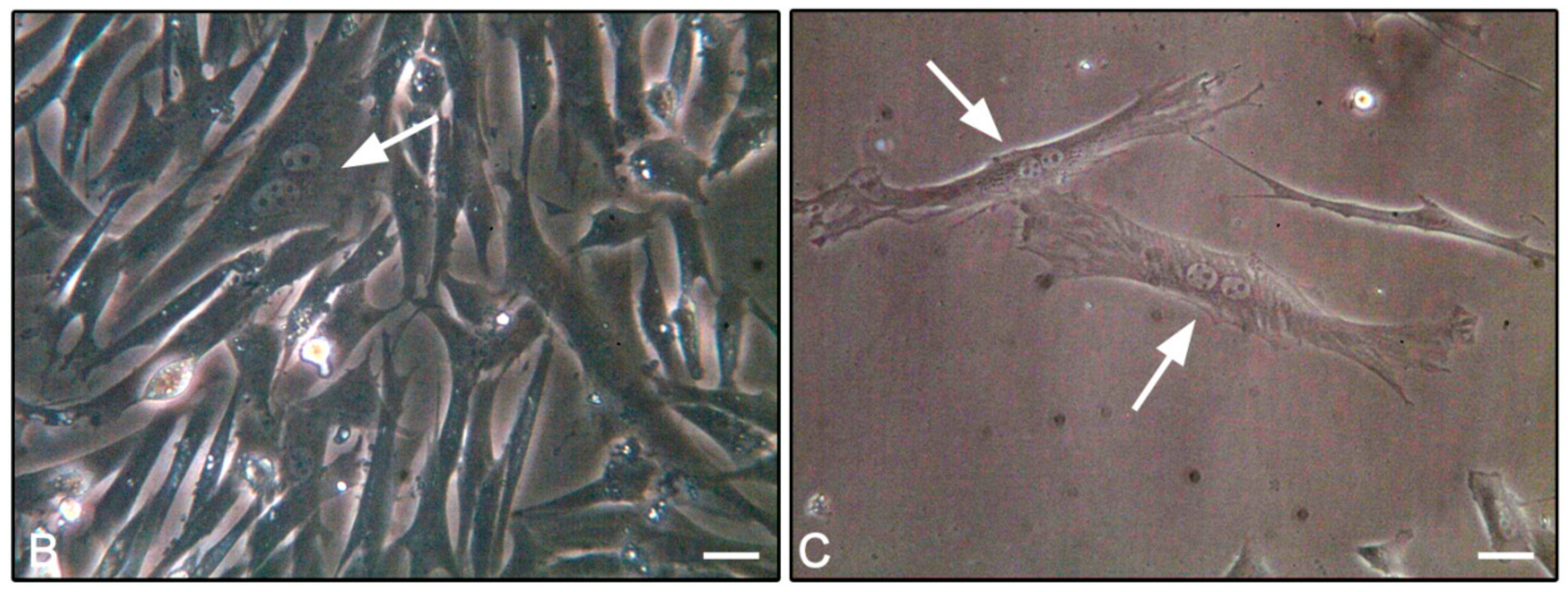
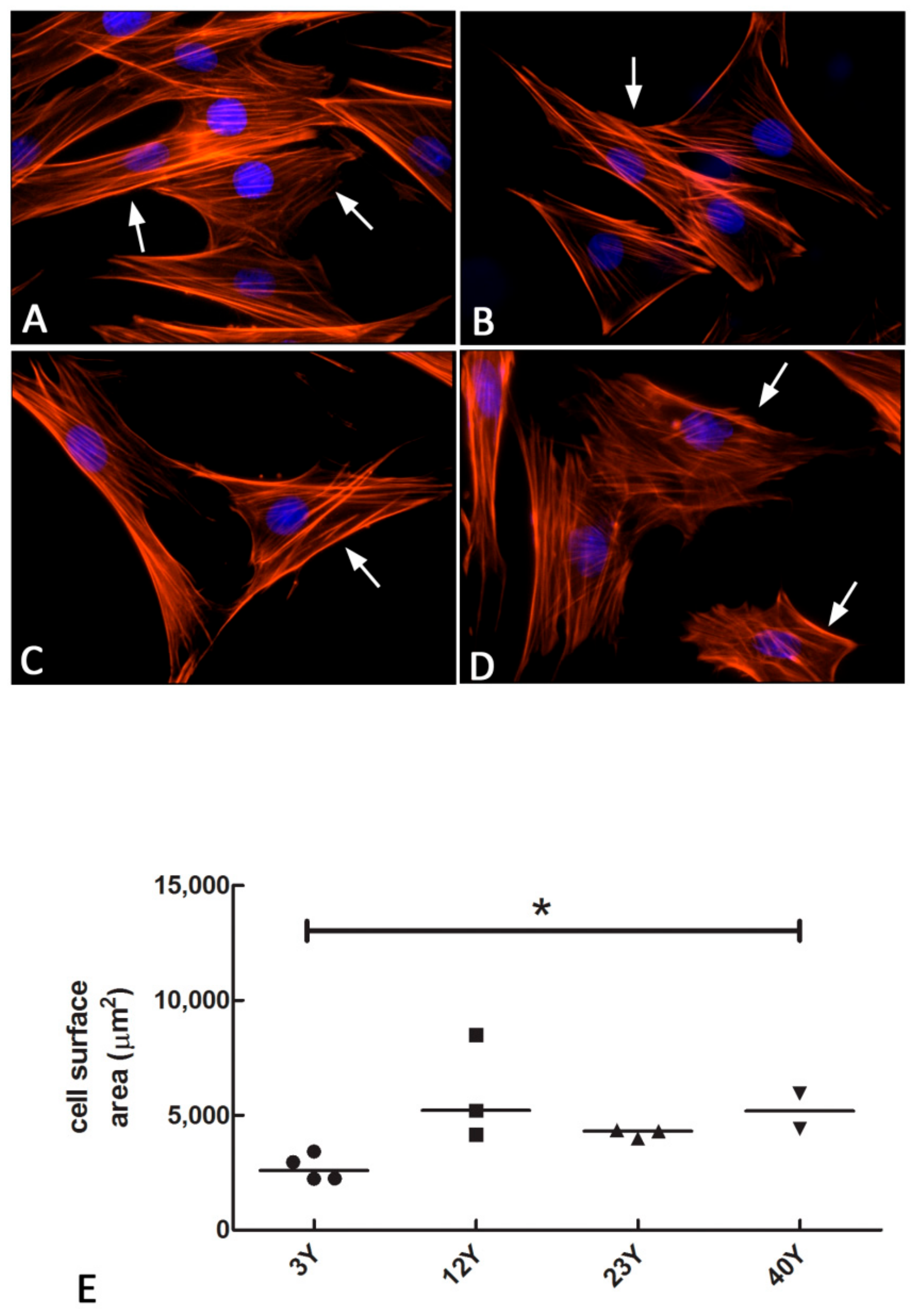
2.6. F-Actin Staining
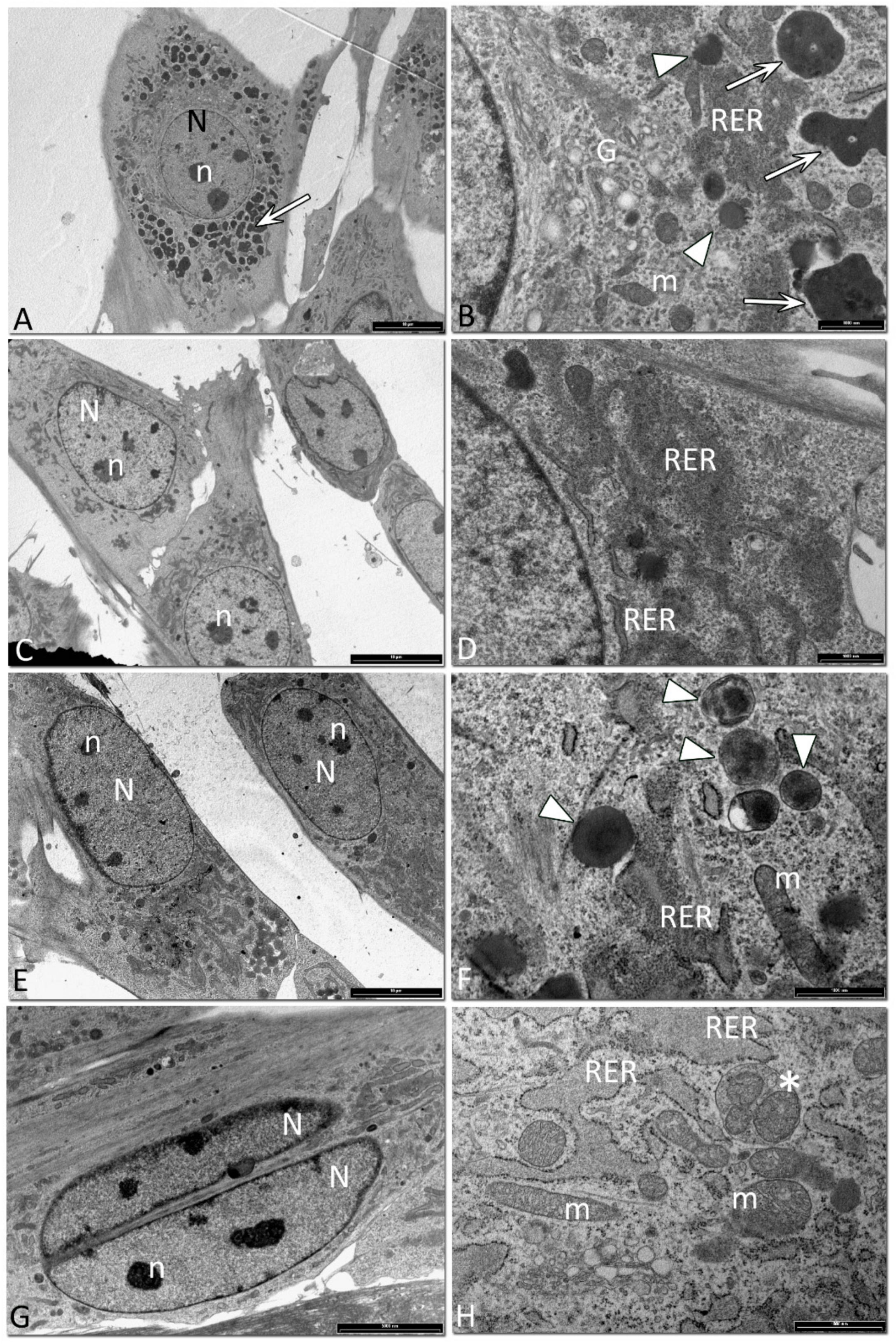
2.7. TEM Analysis
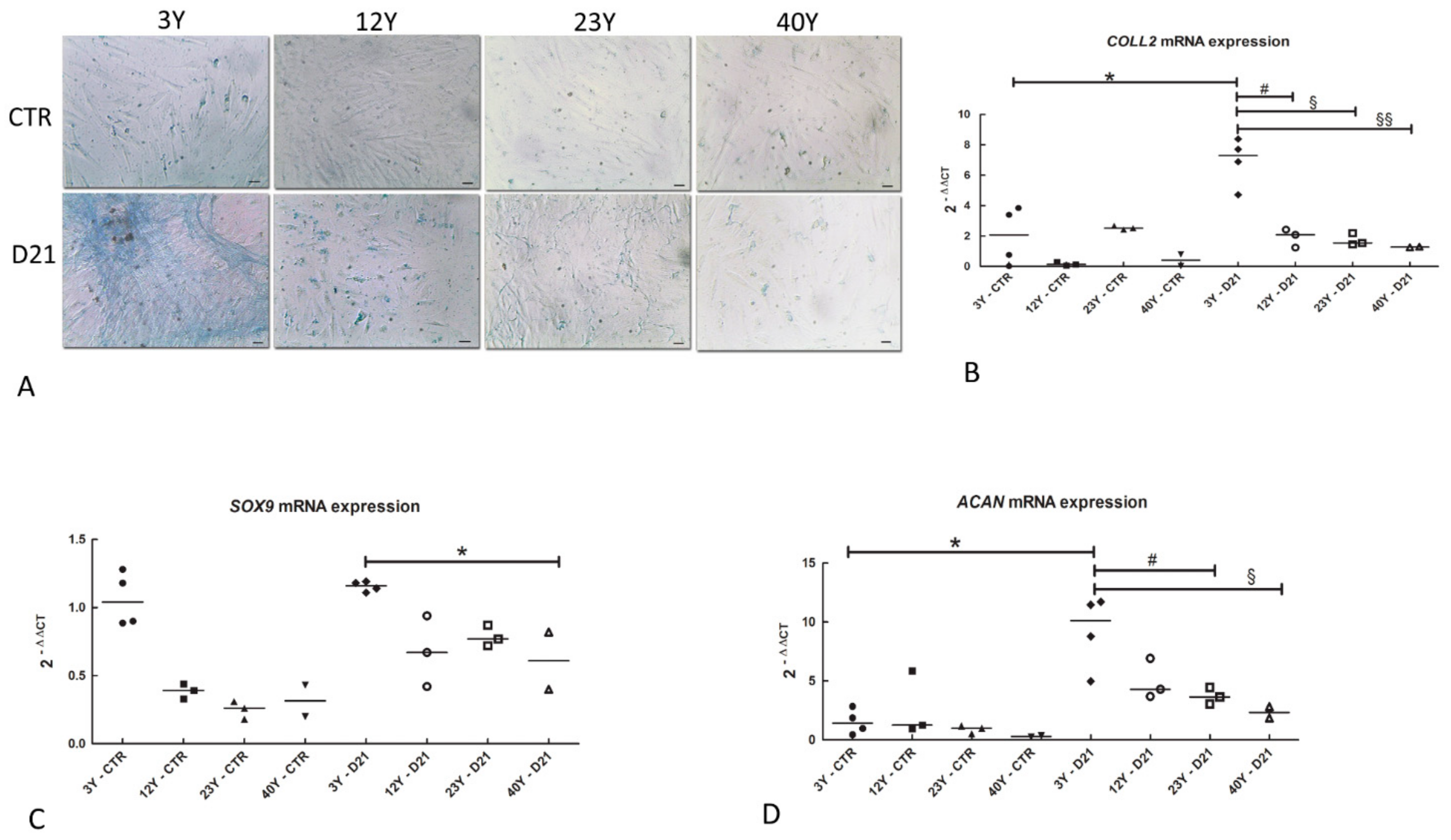
2.8. Alcian Blue
2.9. Chondrogenic Differentiation
2.10. Real-Time PCR
2.11. Statistical Analysis
3. Results
3.1. Mesenchymal and Self-Renewal Expression Markers
3.2. SfMSCs Proliferation Assay
3.3. Cell Cycle Analysis
3.4. F-Actin Staining
3.5. Ultrastructural Analysis of Equine SfMSCs
3.6. Chondrogenic Differentiation
4. Discussion
5. Conclusion
Author Contributions
Funding
Conflicts of Interest
References
- Huang, Z.Y.; Xie, H.Q.; Silini, A.; Parolini, O.; Zhang, Y.; Deng, L.; Huang, Y.C. Mesenchymal tem/Progenitor Cells Derived from Articular Cartilage, Synovial Membrane and Synovial Fluid for Cartilage Regeneration: Current Status and Future Perspectives. Stem Cell Rev. 2017, 13, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Berebichez-Fridman, R.; Montero-Olvera, P.R. Sources and Clinical Applications of Mesenchymal Stem Cells: State-of-the-Art Review. Sultan Qaboos Univ. Med. J. 2018, 18, e264–e277. [Google Scholar] [CrossRef] [PubMed]
- Fitzsimmons, R.E.B.; Mazurek, M.S.; Soos, A.; Simmons, C.A. Mesenchymal Stromal/Stem Cells in Regenerative Medicine and Tissue Engineering. Stem Cells Int. 2018, 2018, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Teti, G.; Cavallo, C.; Grigolo, B.; Giannini, S.; Facchini, A.; Mazzotti, A.; Falconi, M. Ultrastructural Analysis of Human Bone Marrow Mesenchymal Stem Cells During In Vitro Osteogenesis and Chondrogenesis. Microsc. Res. Tech. 2012, 75, 596–604. [Google Scholar] [CrossRef]
- Focaroli, S.; Teti, G.; Salvatore, V.; Durante, S.; Belmonte, M.M.; Giardino, R.; Mazzotti, A.; Bigi, A.; Falconi, M. Chondrogenic Differentiation of Human Adipose Mesenchimal Stem Cells: Influence of a Biomimetic Gelatin Genipin Crosslinked Porous Scaffold. Microsc. Res. Tech. 2014, 77, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Focaroli, S.; Teti, G.; Salvatore, V.; Orienti, I.; Falconi, M. Calcium/Cobalt Alginate Beads as Functional Scaffolds for Cartilage Tissue Engineering. Stem Cells Int. 2016, 2016, 2030478. [Google Scholar] [CrossRef]
- Teti, G.; Salvatore, V.; Focaroli, S.; Durante, S.; Mazzotti, A.; Dicarlo, M.; Mattioli-Belmonte, M.; Orsini, G. In Vitro osteogenic and odontogenic differentiation of human dental pulp stem cells seeded on carboxymethyl cellulose-hydroxyapatite hybrid hydrogel. Front. Physiol. 2015, 6, 297. [Google Scholar] [CrossRef]
- Teti, G.; Focaroli, S.; Salvatore, V.; Mazzotti, E.; Ingra, L.; Mazzotti, A.; Falconi, M. The Hypoxia-Mimetic Agent Cobalt Chloride Differently Affects Human Mesenchymal Stem Cells in Their Chondrogenic Potential. Stem Cells Int. 2018, 2018, 3237253. [Google Scholar] [CrossRef]
- Bunnell, B.A.; Flaat, M.; Gagliardi, C.; Patel, B.; Ripoll, C. Adipose-derived Stem Cells: Isolation, Expansion and Differentiation. Methods 2008, 45, 115–120. [Google Scholar] [CrossRef]
- Minguell, J.J.; Erices, A.; Conget, P. Mesenchymal Stem Cells. Exp. Biol. Med. 2001, 226, 507–520. [Google Scholar] [CrossRef]
- So, W.K.; Cheung, T.H. Molecular Regulation of Cellular Quiescence: A Perspective from Adult Stem Cells and Its Niches. Methods Mol. Biol. 2018, 1686, 1–25. [Google Scholar]
- Friedenstein, A.J.; Gorskaja, J.F.; Kulagina, N.N. Fibroblast Precursors in Normal and Irradiated Mouse Hematopoietic Organs. Exp. Hematol. 1976, 4, 267–274. [Google Scholar]
- Conget, P.A.; Allers, C.; Minguell, J.J. Identification of a Discrete Population of Human Bone Marrow-Derived Mesenchymal Cells Exhibiting Properties of Uncommitted Progenitors. J. Hematotherapy Stem Cell Res. 2001, 10, 749–758. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human Adipose Tissue Is a Source of Multipotent Stem Cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef]
- Dong, Q.; Wang, Y.; Mohabatpour, F.; Zheng, L.; Papagerakis, S.; Chen, D.; Papagerakis, P. Dental Pulp Stem Cells: Isolation, Characterization, Expansion, and Odontoblast Differentiation for Tissue Engineering. Methods Mol. Biol. 2019, 1922, 91–101. [Google Scholar]
- Ferretti, C.; Borsari, V.; Falconi, M.; Gigante, A.; Lazzarini, R.; Fini, M.; Di Primio, R.; Mattioli-Belmonte, M.; Primio, R. Human Periosteum-Derived Stem Cells for Tissue Engineering Applications: The Role of VEGF. Stem Cell Rev. 2012, 8, 882–890. [Google Scholar] [CrossRef]
- Mattioli-Belmonte, M.; Teti, G.; Salvatore, V.; Focaroli, S.; Orciani, M.; Dicarlo, M.; Fini, M.; Orsini, G.; Di Primio, R.; Falconi, M. Stem cell origin differently affects bone tissue engineering strategies. Front. Physiol. 2015, 6, 266. [Google Scholar] [CrossRef]
- Lu, L.-L.; Liu, Y.-J.; Yang, S.-G.; Zhao, Q.-J.; Wang, X.; Gong, W.; Han, Z.-B.; Xu, Z.-S.; Lu, Y.-X.; Liu, D.; et al. Isolation and characterization of human umbilical cord mesenchymal stem cells with hematopoiesis-supportive function and other potentials. Haematologica 2006, 91, 1017–1026. [Google Scholar]
- Merlo, B.; Teti, G.; Mazzotti, E.; Ingrà, L.; Salvatore, V.; Buzzi, M.; Cerqueni, G.; Dicarlo, M.; Lanci, A.; Castagnetti, C.; et al. Wharton’s Jelly Derived Mesenchymal Stem Cells: Comparing Human and Horse. Stem Cell Rev. 2018, 14, 574–584. [Google Scholar] [CrossRef]
- De Bari, C.; Dell’Accio, F.; Tylzanowski, P.; Luyten, F.P. Multipotent Mesenchymal Stem Cells from Adult Human Synovial Membrane. Arthritis Rheum. 2001, 44, 1928–1942. [Google Scholar] [CrossRef]
- Harvanova, D.; Tothova, T.; Sarissky, M.; Amrichova, J.; Rosocha, J. Isolation and Characterization of Synovial Mesenchymal Stem Cells. Folia Biol. 2011, 57, 119–124. [Google Scholar]
- Jones, E.A.; English, A.; Henshaw, K.; Kinsey, S.E.; Markham, A.F.; Emery, P.; McGonagle, D. Enumeration and Phenotypic Characterization of Synovial Fluid Multipotential Mesenchymal Progenitor Cells in Inflammatory and Degenerative Arthritis. Arthritis Rheum. 2004, 50, 817–827. [Google Scholar] [CrossRef]
- Morito, T.; Muneta, T.; Hara, K.; Ju, Y.J.; Mochizuki, T.; Makino, H.; Umezawa, A.; Sekiya, I. Synovial Fluid-Derived Mesenchymal Stem Cells Increase after Intra-Articular Ligament Injury in Humans. Rheumatology 2008, 47, 1137–1143. [Google Scholar] [CrossRef]
- Sekiya, I.; Ojima, M.; Suzuki, S.; Yamaga, M.; Horie, M.; Koga, H.; Tsuji, K.; Miyaguchi, K.; Ogishima, S.; Tanaka, H.; et al. Human Mesenchymal Stem Cells in Synovial Fluid Increase in the Knee with Degenerated Cartilage and Osteoarthritis. J. Orthop. Res. 2012, 30, 943–949. [Google Scholar] [CrossRef]
- Pei, M.; He, F.; Boyce, B.M.; Kish, V.L. Repair of Full-Thickness Femoral Condyle Cartilage Defects Using Allogeneic Synovial Cell-Engineered Tissue Constructs. Osteoarthr. Cartil. 2009, 17, 714–722. [Google Scholar] [CrossRef]
- Hunziker, E.B.; Rosenberg, L.C. Repair of Partial-Thickness Defects in Articular Cartilage: Cell Recruitment from the Synovial Membrane. J. Bone Jt. Surg. Am. 1996, 78, 721–733. [Google Scholar] [CrossRef]
- Murata, D.; Miyakoshi, D.; Hatazoe, T.; Miura, N.; Tokunaga, S.; Fujiki, M.; Nakayama, K.; Misumi, K. Multipotency of Equine Mesenchymal Stem Cells Derived from Synovial Fluid. Vet. J. 2014, 202, 53–61. [Google Scholar] [CrossRef]
- Yu, K.R.; Kang, K.S. Aging-Related Genes in Mesenchymal Stem Cells: A Mini-Review. Gerontology 2013, 59, 557–563. [Google Scholar] [CrossRef]
- Yang, Y.K. Aging of Mesenchymal Stem Cells: Implication in Regenerative Medicine. Regen. Ther. 2018, 9, 120–122. [Google Scholar] [CrossRef]
- Zaim, M.; Karaman, S.; Cetin, G.; Isik, S. Donor Age and Long-Term Culture Affect Differentiation and Proliferation of Human Bone Marrow Mesenchymal Stem Cells. Ann. Hematol. 2012, 91, 1175–1186. [Google Scholar] [CrossRef]
- Choudhery, M.S.; Badowski, M.; Muise, A.; Pierce, J.; Harris, D.T. Donor Age Negatively Impacts Adipose Tissue-Derived Mesenchymal Stem Cell Expansion and Differentiation. J. Transl. Med. 2014, 12, 8. [Google Scholar] [CrossRef]
- Lorenzini, A.; Maier, A.B. Influence of donor age and species longevity on replicative cellular senescence. In Cellular Ageing and Replicative Senescence; Ruttan, S.I.S., Hayflick, L., Eds.; Springer: Cham, Switzerland, 2016; pp. 49–70. [Google Scholar]
- Moran, C.J.; Ramesh, A.; Brama, P.A.; O’Byrne, J.M.; O’Brien, F.J.; Levingstone, T.J. The benefits and limitations of animal models for translational research in cartilage repair. J. Exp. Orthop. 2016, 3, 1. [Google Scholar] [CrossRef]
- Lonetti, A.; Cappellini, A.; Bertaina, A.; Locatelli, F.; Pession, A.; Buontempo, F.; Evangelisti, C.; Orsini, E.; Zambonin, L.; Neri, L.M.; et al. Improving Nelarabine Efficacy in T Cell Acute Lymphoblastic Leukemia by Targeting Aberrant Pi3k/Akt/Mtor Signaling Pathway. J. Hematol. Oncol. 2016, 9, 114. [Google Scholar] [CrossRef]
- Dominici, M.; le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Dzobo, K.; Turnley, T.; Wishart, A.; Rowe, A.; Kallmeyer, K.; van Vollenstee, F.A.; Thomford, N.E.; Dandara, C.; Chopera, D.; Pepper, M.S.; et al. Fibroblast-Derived Extracellular Matrix Induces Chondrogenic Differentiation in Human Adipose-Derived Mesenchymal Stromal/Stem Cells In Vitro. Int. J. Mol. Sci. 2016, 17, 1259. [Google Scholar] [CrossRef]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [Green Version]
- Hou, A.; Chen, P.; Tang, H.; Meng, H.; Cheng, X.; Wang, Y.; Zhang, Y.; Peng, J. Cellular Senescence in Osteoarthritis and Anti-Aging Strategies. Mech. Ageing Dev. 2018, 175, 83–87. [Google Scholar] [CrossRef]
- Lim, J.E.; Son, Y. Endogenous Stem Cells in Homeostasis and Aging. Tissue Eng. Regen. Med. 2017, 14, 679–698. [Google Scholar] [CrossRef]
- Mak, J.; Jablonski, C.L.; Leonard, C.A.; Dunn, J.F.; Raharjo, E.; Matyas, J.R.; Biernaskie, J.; Krawetz, R.J. Intra-Articular Injection of Synovial Mesenchymal Stem Cells Improves Cartilage Repair in a Mouse Injury Model. Sci. Rep. 2016, 6, 23076. [Google Scholar] [CrossRef]
- Neybecker, P.; Henrionnet, C.; Pape, E.; Mainard, D.; Galois, L.; Loeuille, D.; Gillet, P.; Pinzano, A. In Vitro and In Vivo Potentialities for Cartilage Repair from Human Advanced Knee Osteoarthritis Synovial Fluid-Derived Mesenchymal Stem Cells. Stem Cell Res. Ther. 2018, 9, 329. [Google Scholar] [CrossRef]
- Janes, J.G.; Kennedy, L.A.; Garrett, K.S.; Engiles, J.B. Common Lesions of the Distal End of the Third Metacarpal/Metatarsal Bone in Racehorse Catastrophic Breakdown Injuries. J. Vet. Diagn. Investig. 2017, 29, 431–436. [Google Scholar] [CrossRef]
- De Schauwer, C.; Meyer, E.; van de Walle, G.R.; van Soom, A. Markers of Stemness in Equine Mesenchymal Stem Cells: A Plea for Uniformity. Theriogenology 2011, 75, 1431–1443. [Google Scholar] [CrossRef]
- Fossett, E.; Khan, W.S.; Pastides, P.; Adesida, A.B. The Effects of Ageing on Proliferation Potential, Differentiation Potential and Cell Surface Characterisation of Human Mesenchymal Stem Cells. Curr. Stem Cell Res. Ther. 2012, 7, 282–286. [Google Scholar] [CrossRef]
- Tsai, C.C.; Hung, S.C. Functional roles of pluripotency transcription factors in mesenchymal stem cells. Cell Cycle 2012, 11, 3711–3712. [Google Scholar] [CrossRef] [Green Version]
- Davoli, T.; de Lange, T. The Causes and Consequences of Polyploidy in Normal Development and Cancer. Annu. Rev. Cell Dev. Biol. 2011, 27, 585–610. [Google Scholar] [CrossRef] [Green Version]
- Ullah, Z.; Kohn, M.J.; Yagi, R.; Vassilev, L.T.; DePamphilis, M.L. Differentiation of Trophoblast Stem Cells into Giant Cells Is Triggered by P57/Kip2 Inhibition of Cdk1 Activity. Genes Dev. 2008, 22, 3024–3036. [Google Scholar] [CrossRef]
- Ravid, K.; Lu, J.; Zimmet, J.M.; Jones, M.R. Roads to Polyploidy: The Megakaryocyte Example. J. Cell. Physiol. 2002, 190, 7–20. [Google Scholar] [CrossRef]
- Celton-Morizur, S.; Desdouets, C. Polyploidization of Liver Cells. Adv. Exp. Med. Biol. 2010, 676, 123–135. [Google Scholar]
- Vignery, A. Osteoclasts and Giant Cells: Macrophage-Macrophage Fusion Mechanism. Int. J. Exp. Pathol. 2000, 81, 291–304. [Google Scholar] [CrossRef]
- Liu, Z.; Yue, S.; Chen, X.; Kubin, T.; Braun, T. Regulation of Cardiomyocyte Polyploidy and Multinucleation by CyclinG1. Circ. Res. 2010, 106, 1498–1506. [Google Scholar] [CrossRef]
- Hixon, M.L.; Gualberto, A. Vascular Smooth Muscle Polyploidization from Mitotic Checkpoints to Hypertension. Cell Cycle 2003, 2, 105–110. [Google Scholar] [CrossRef]
- Gorla, G.R.; Malhi, H.; Gupta, S. Polyploidy Associated with Oxidative Injury Attenuates Proliferative Potential of Cells. J. Cell Sci. 2001, 114, 2943–2951. [Google Scholar]
- Estrada, J.C.; Torres, Y.; Benguria, A.; Dopazo, A.; Roche, E.; Carrera-Quintanar, L.; Perez, R.A.; Enríquez, J.A.; Torres, R.; Ramírez, J.C.; et al. Human Mesenchymal Stem Cell-Replicative Senescence and Oxidative Stress Are Closely Linked to Aneuploidy. Cell Death Dis. 2013, 4, e691. [Google Scholar] [CrossRef]
- Hwang, E.S.; Yoon, G.; Kang, H.T. A Comparative Analysis of the Cell Biology of Senescence and Aging. Cell Mol. Life Sci. 2009, 66, 2503–2524. [Google Scholar] [CrossRef]
- Wang, E.; Gundersen, D. Increased Organization of Cytoskeleton Accompanying the Aging of Human Fibroblasts in Vitro. Exp. Cell Res. 1984, 154, 191–202. [Google Scholar] [CrossRef]
- Revuelta, M.; Matheu, A. Autophagy in Stem Cell Aging. Aging Cell 2017, 16, 912–915. [Google Scholar] [CrossRef]
- Rubinsztein, D.C.; Marino, G.; Kroemer, G. Autophagy and Aging. Cell 2011, 146, 682–695. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Prat, L.; Martinez-Vicente, M.; Perdiguero, E.; Ortet, L.; Rodriguez-Ubreva, J.; Rebollo, E.; Ruiz-Bonilla, V.; Gutarra, S.; Ballestar, E.; Serrano, A.L.; et al. Autophagy Maintains Stemness by Preventing Senescence. Nature 2016, 529, 37–42. [Google Scholar] [CrossRef]
- Lee, W.S.; Yoo, W.H.; Chae, H.J. ER Stress and Autophagy. Curr. Mol. Med. 2015, 15, 735–745. [Google Scholar] [CrossRef]
- Taylor, R.C. Aging and the UPR (ER). Brain Res. 2016, 1648, 588–593. [Google Scholar] [CrossRef]
- Lemasters, J.J. Variants of Mitochondrial Autophagy: Types 1 and 2 Mitophagy and Micromitophagy (Type 3). Redox Biol 2014, 2, 749–754. [Google Scholar] [CrossRef]
- Szychlinska, M.A.; Stoddart, M.J.; D’Amora, U.; Ambrosio, L.; Alini, M.; Musumeci, G. Mesenchymal Stem Cell-Based Cartilage Regeneration Approach and Cell Senescence: Can We Manipulate Cell Aging and Function? Tissue Eng. Part B Rev. 2017, 23, 529–539. [Google Scholar] [CrossRef]
- Matjusaitis, M.; Chin, G.; Sarnoski, E.A.; Stolzing, A. Biomarkers to Identify and Isolate Senescent Cells. Ageing Res. Rev. 2016, 29, 1–12. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | Age (Years) | Gender | Donor’s Number | |
|---|---|---|---|---|
| 1 | Standardbred | 3 | M | 4 |
| 2 | Standardbred | 12 | F | 2 |
| 3 | Pleasure riding horse | 12 | M | 1 |
| 4 | Standardbred | 23 | F | 3 |
| 5 | Pleasure riding horse | 40 | M | 2 |
| Name | Forward | Reverse | bp |
|---|---|---|---|
| CD90 | 5′-ATGAGAATACCACCGCCACA-3′ | 5′-AGTTTGTCTCGGAGCACAGA-3′ | 262 |
| CD105 | 5′-TCAGGTCCCCAACACTAACC-3′ | 5′-AGTCTTGTTCGTGCTGAGGA-3′ | 148 |
| CD34 | 5′-CCTTGCCCAGTCTGAGGTTA-3′ | 5′-GTCTTGCGGGAATAGTGCTG-3′ | 172 |
| SOX9 | 5′-GAACAGCCCGTCTACACACA-3′ | 5′-GCCACTGATTCGCAACAAGG-3′ | 235 |
| COLL2 | 5′-CTGGCAAGCAAGGAGACAGA-3′ | 5′-CCATTAGCGCCATCTTTGCC-3′ | 292 |
| ACAN | 5′-TCATCTAGAGCCCACTGCCT-3′ | 5′-AGTCCACCGAGGTCCTCTAC-3′ | 234 |
| NANOG | 5′-TCTCTCCTCTGCCTTCCTCC -3′ | 5′-TCTGCTGGAGGCTGAGGTAT-3′ | 225 |
| OCT4 | 5′-GGTACGAGTGTGGTTCTGCA-3′ | 5′-ACCGAGGAGTACAGCGTAGT-3′ | 192 |
| SOX2 | 5′-GCCCTGCAGTACAACTCCAT-3′ | 5′-GACTTGACCACCGAACCCAT-3′ | 128 |
| GAPDH | 5′-TGCCCCAATGTTTGTGATGG-3′ | 5′-CACTGTGGTCATGAGTCCCT-3′ | 154 |
| G0/G1 (%) | S Phase (%) | G2/M (%) | 4N (%) | |
|---|---|---|---|---|
| 3Y Group | ||||
| Donor 1 | 56.6 ± 1.2 | 5.3 ± 2.6 | 28.6 ± 1.4 | 8.6 ± 1.2 a |
| Donor 2 | 58.0 ± 1.1 | 9.7 ± 1.4 | 26.9 ± 1.6 | 7.5 ± 0.9 a |
| Donor 3 | 58.5 ± 1.4 | 21.3 ± 0.9 | 18.3 ± 1.1 | 7.4 ± 0.9 a |
| Donor 4 | 48.6 ± 0.8 | 15.0 ± 0.5 | 28.2 ± 0.9 | 7.6 ± 0.3 a |
| 12Y Group | ||||
| Donor 1 | 64.1 ± 1.6 | 10.1 ± 1.1 | 23.3 ± 1.4 | 12.0 ± 1.2 |
| Donor 2 | 65.6 ± 1.6 | 9.5 ± 1.5 | 23.3 ± 1.1 | 8.5 ± 0.9 |
| Donor 3 | 59.9 ± 1.7 | 9.0 ± 1.6 | 22.6 ± 1.2 | 8.2 ± 0.5 |
| 23Y Group | ||||
| Donor 1 | 68.7 ± 1.6 | 4.9 ± 0.9 | 25.5 ± 1.4 | 10.5 ± 0.6 |
| Donor 2 | 61.6 ± 0.5 | 9.4 ± 0.2 | 16.5 ± 0.6 | 9.5 ± 0.3 |
| Donor 3 | 84.7 ± 0.8 | 7.8 ± 0.3 | 5.2 ± 0.7 | 10.7 ± 0.3 |
| 40Y Group | ||||
| Donor 1 | 67.1 ± 1.1 | 9.9 ± 1.5 | 18.9 ± 1.2 | 14.4 ± 0.7 |
| Donor 2 | 74.3 ± 1.2 | 8.1 ± 2.4 | 16.1 ± 1.5 | 18.5 ± 1.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazzotti, E.; Teti, G.; Falconi, M.; Chiarini, F.; Barboni, B.; Mazzotti, A.; Muttini, A. Age-Related Alterations Affecting the Chondrogenic Differentiation of Synovial Fluid Mesenchymal Stromal Cells in an Equine Model. Cells 2019, 8, 1116. https://doi.org/10.3390/cells8101116
Mazzotti E, Teti G, Falconi M, Chiarini F, Barboni B, Mazzotti A, Muttini A. Age-Related Alterations Affecting the Chondrogenic Differentiation of Synovial Fluid Mesenchymal Stromal Cells in an Equine Model. Cells. 2019; 8(10):1116. https://doi.org/10.3390/cells8101116
Chicago/Turabian StyleMazzotti, Eleonora, Gabriella Teti, Mirella Falconi, Francesca Chiarini, Barbara Barboni, Antonio Mazzotti, and Aurelio Muttini. 2019. "Age-Related Alterations Affecting the Chondrogenic Differentiation of Synovial Fluid Mesenchymal Stromal Cells in an Equine Model" Cells 8, no. 10: 1116. https://doi.org/10.3390/cells8101116
APA StyleMazzotti, E., Teti, G., Falconi, M., Chiarini, F., Barboni, B., Mazzotti, A., & Muttini, A. (2019). Age-Related Alterations Affecting the Chondrogenic Differentiation of Synovial Fluid Mesenchymal Stromal Cells in an Equine Model. Cells, 8(10), 1116. https://doi.org/10.3390/cells8101116