Phosphorylation of TIP3 Aquaporins during Phaseolus vulgaris Embryo Development

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Peptide Synthesis and Purification
2.3. Membrane Protein Isolation
2.4. Phosphoprotein Detection
2.5. Phosphoamino Acid Detection
2.6. PvTIP3;1 and PvTIP3;2 Immunodetection
2.7. Overexpression of PvTIP3;2 in P. pastoris
2.8. Phosphotyrosine Immunoprecipitation
2.9. Mass Spectrometry
2.10. Circular Dichroism (CD) Spectrometry
3. Results
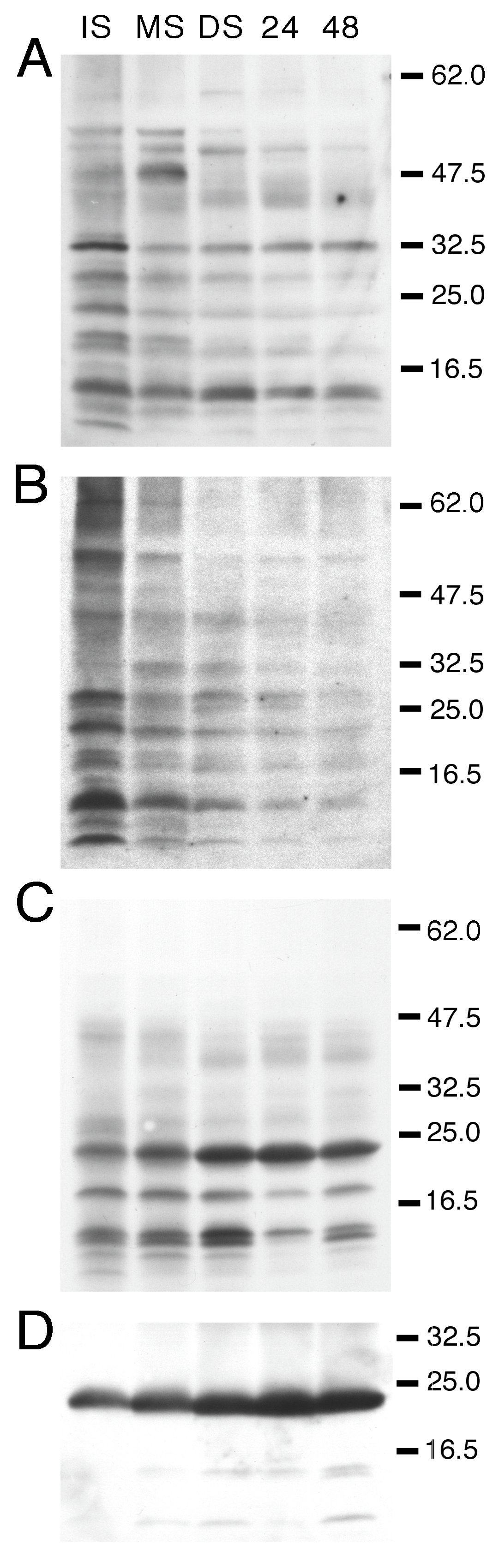
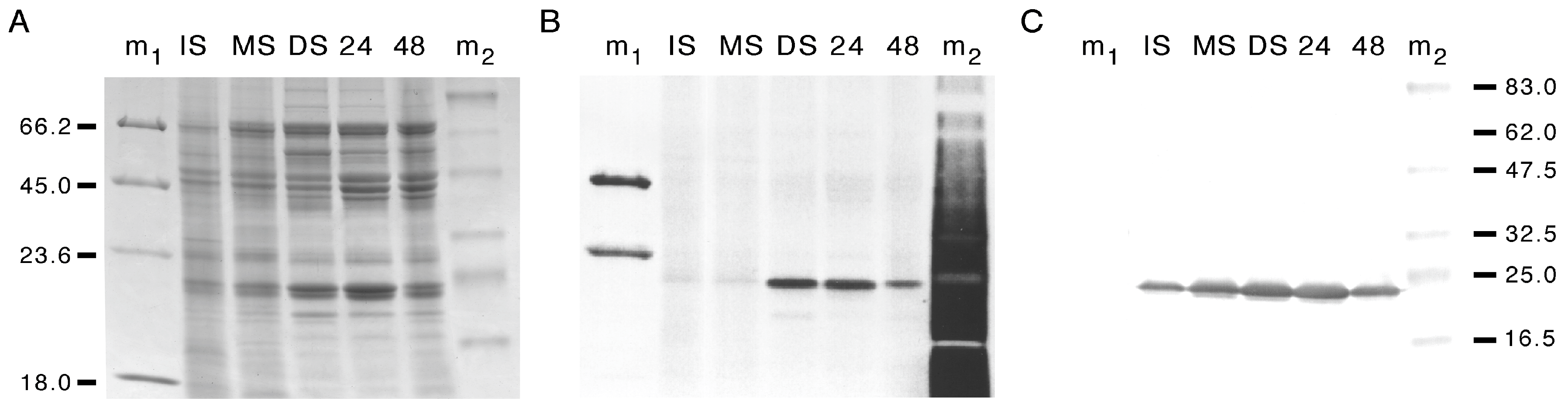
3.1. Developmental Changes in the Abundance of Seed-Membrane Phosphoproteins
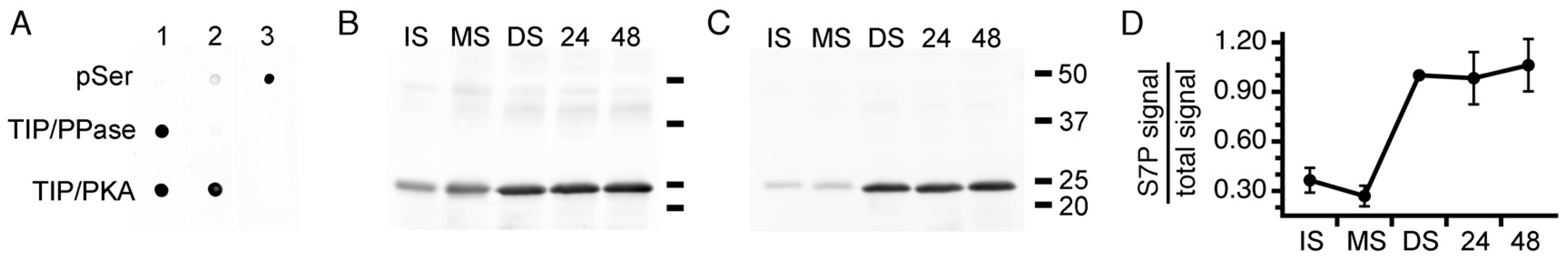
3.2. PvTIP3 Proteins Are the Primary Seed-Membrane Phosphoproteins
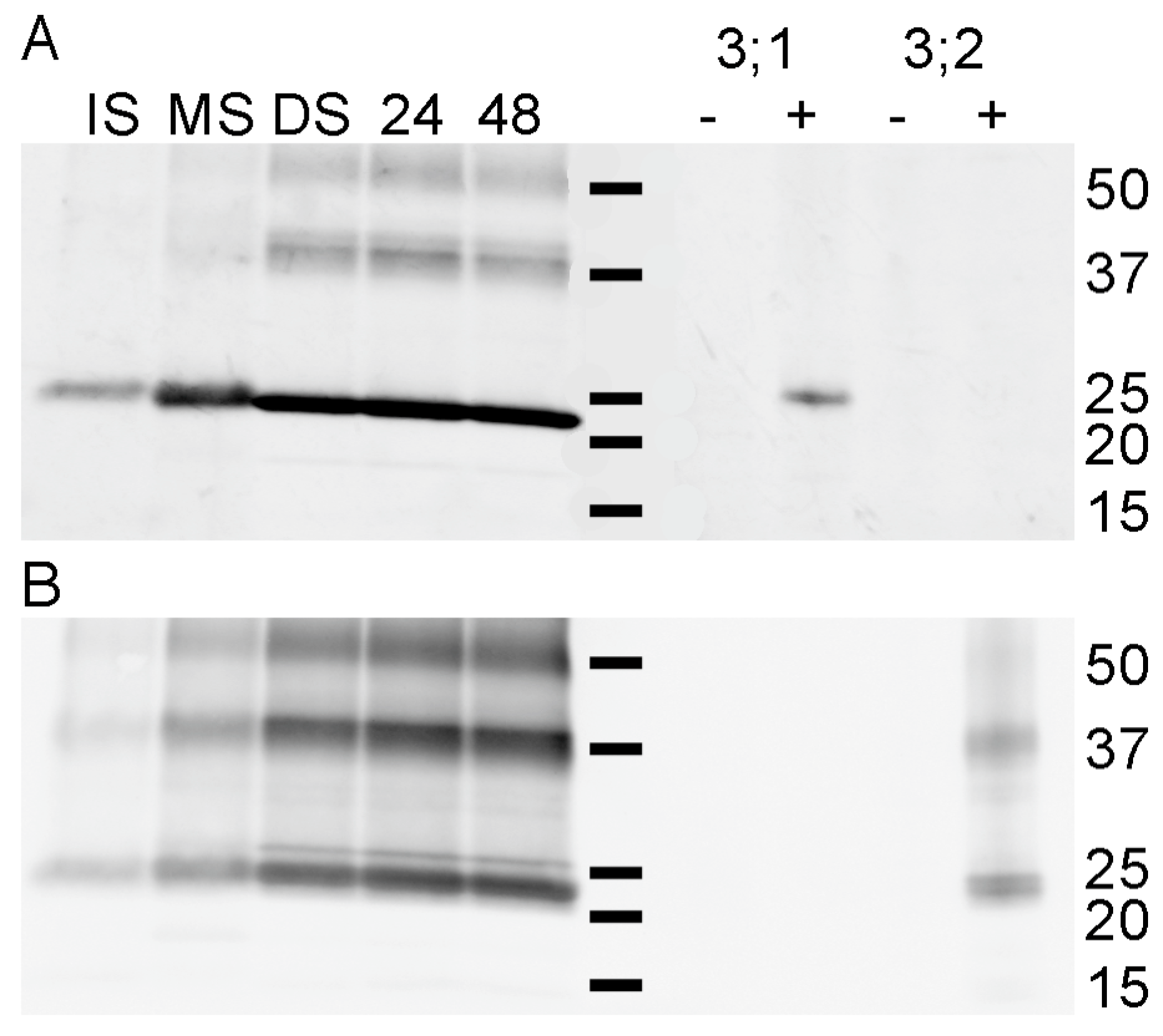
3.3. Expression of PvTIP3;1 and PvTIP3;2 in Seed Development
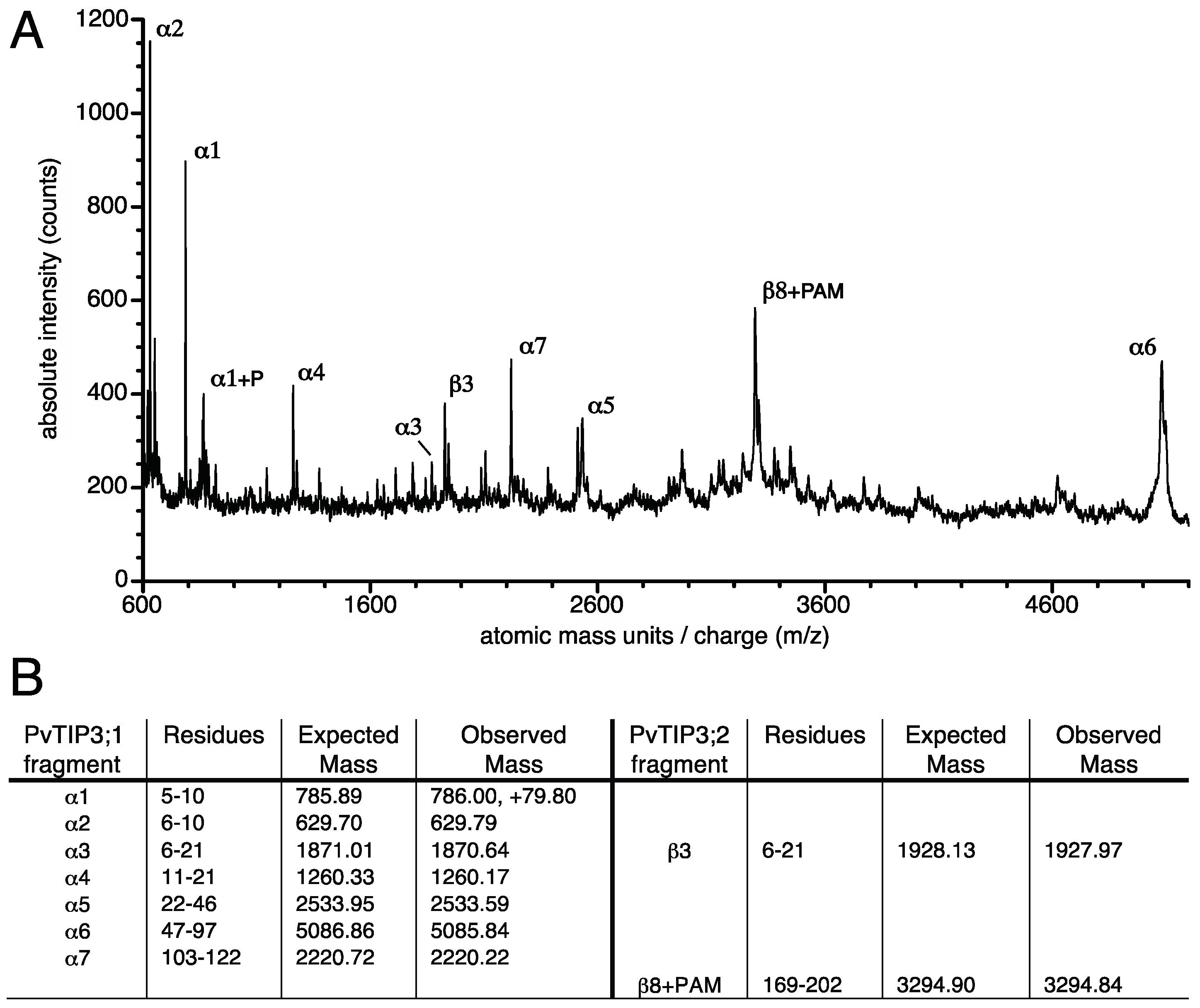
3.4. Ser7 Phosphorylation in PvTIP3;1 Increases Following Seed Maturation
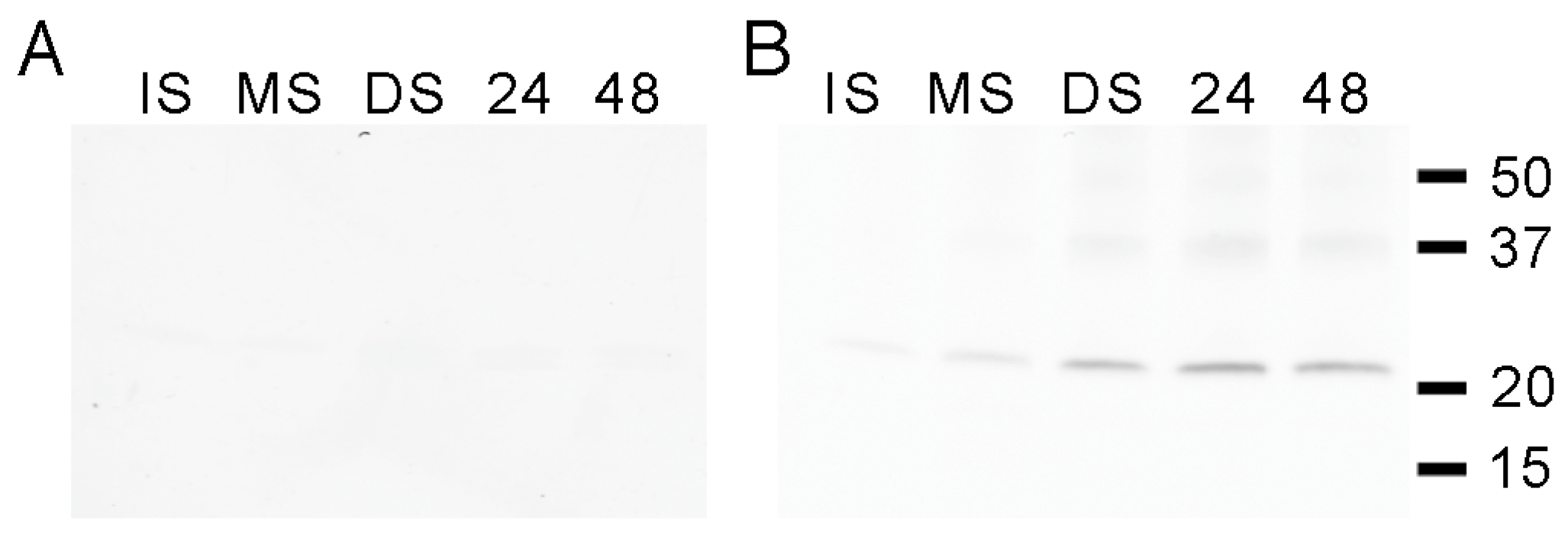
3.5. Tyrosine Phosphorylation of the PvTIP3;2 Amino-Terminal Peptide
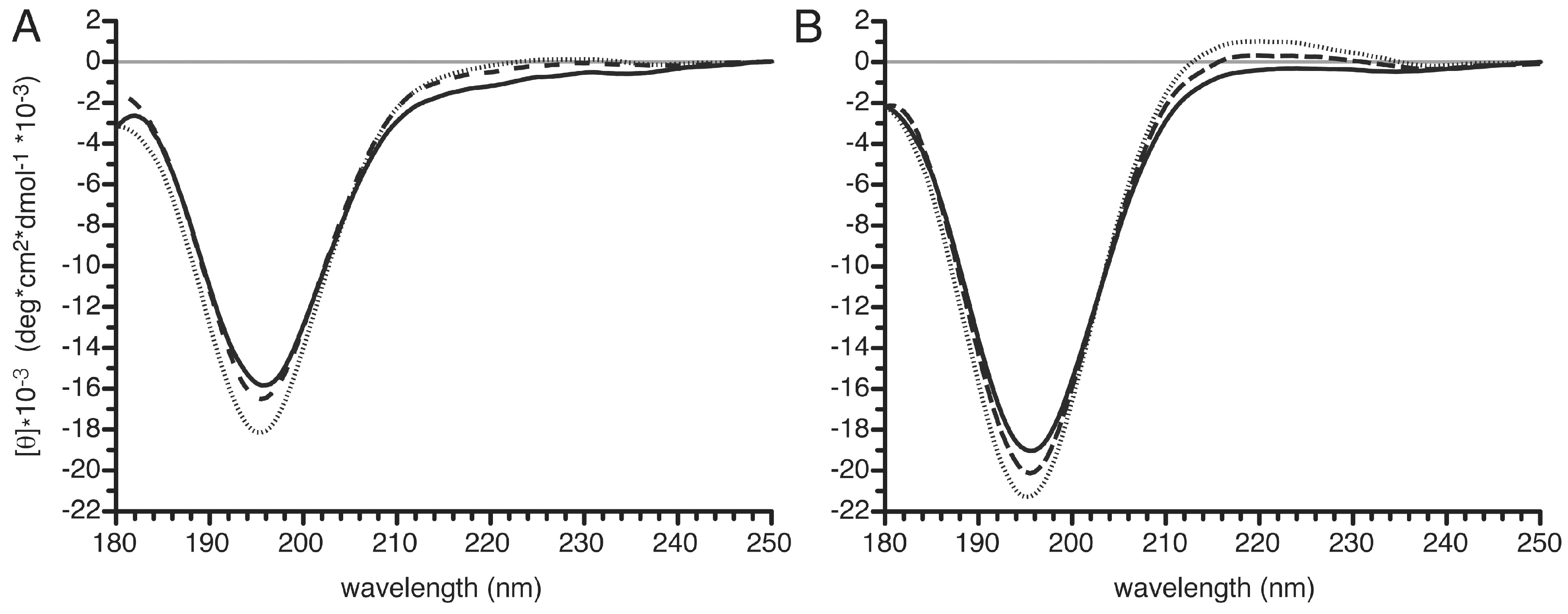
3.6. Structural Changes in the PvTIP3;1 Amino Terminal Peptide upon Phosphorylation
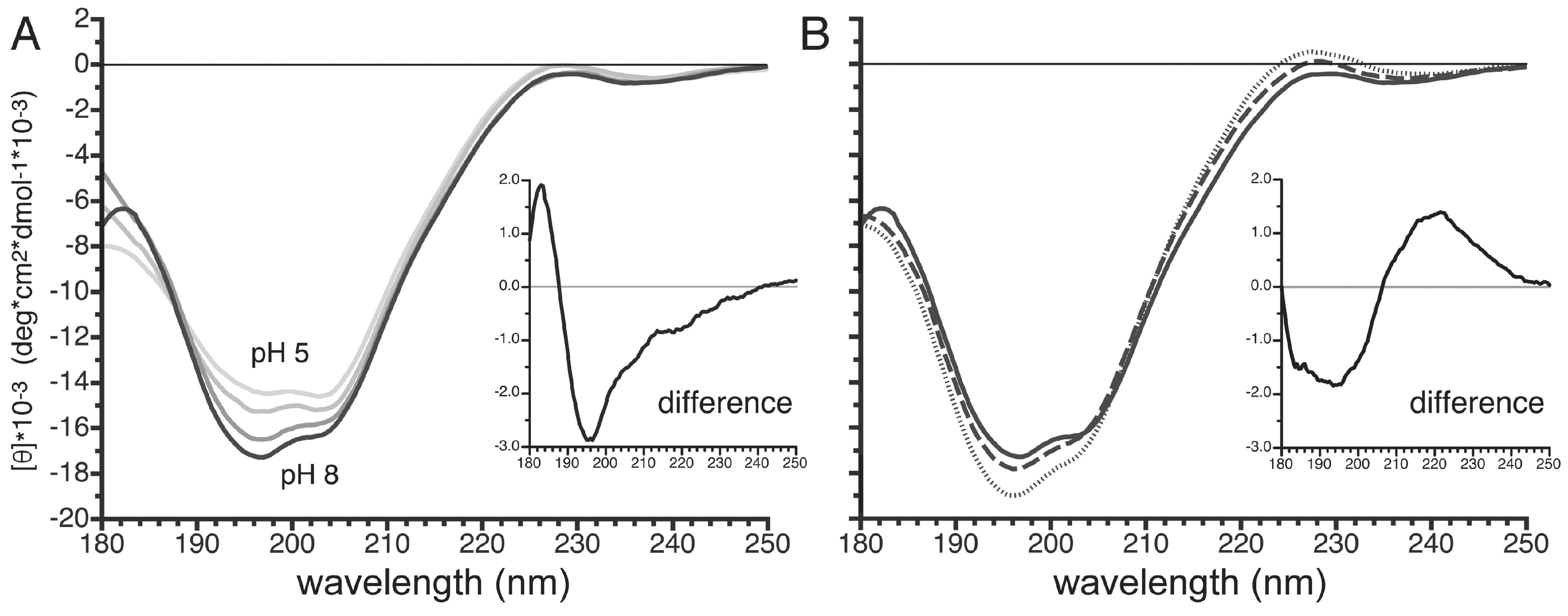
3.7. pH- and Temperature-Dependent Structural Changes in the Carboxy Terminus of PvTIP3;1
4. Discussion
4.1. The Membrane Phosphoproteome Changes Dynamically during Seed Maturation, Dormancy and Germination
4.2. Phosphorylation-Dependent Appearance of Polyproline II Secondary Structure
4.3. Serine Phosphorylation of PvTIP3;1 during Seed Maturation
4.4. Role of PvTIP3;2 in Seed Development
4.5. Possible Roles of PvTIP3;2 Tyrosine Phosphorylation
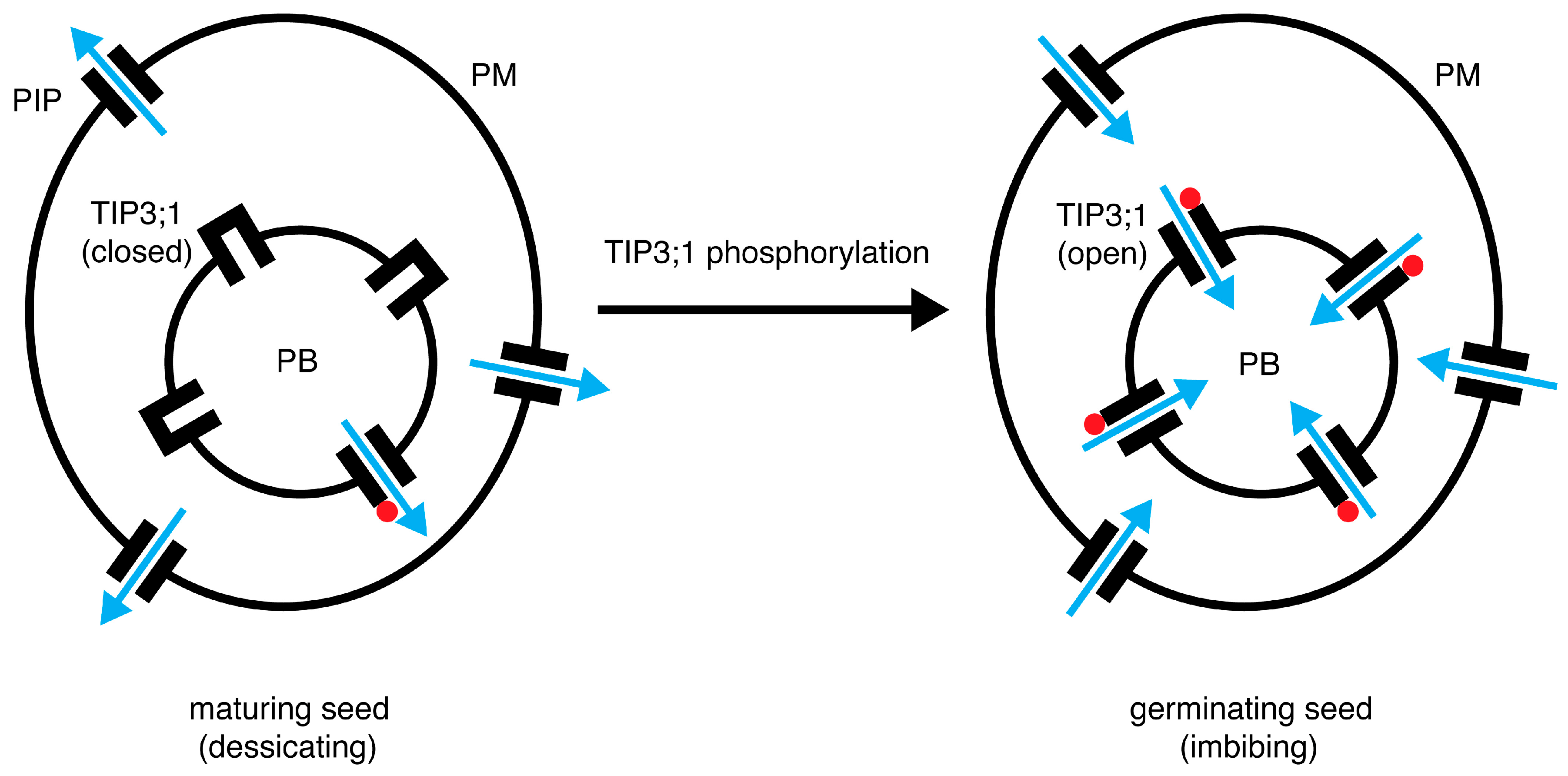
4.6. A Model for the Role of PvTIP3;1 Ser7 Phosphorylation in Seed Development
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gomes, D.; Agasse, A.; Thiébaud, P.; Delrot, S.; Gerós, H.; Chaumont, F. Aquaporins are multifunctional water and solute transporters highly divergent in living organisms. Biochim. Biophys. Acta 2009, 1788, 1213–1228. [Google Scholar] [CrossRef] [Green Version]
- Maurel, C.; Kado, R.T.; Guern, J.; Chrispeels, M.J. Phosphorylation regulates the water channel activity of the seed-specific aquaporin α-TIP. EMBO J. 1995, 14, 3028–3035. [Google Scholar] [CrossRef]
- Maurel, C.; Boursiac, Y.; Luu, D.T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef] [PubMed]
- Fox, A.R.; Maistriaux, L.C.; Chaumont, F. Toward understanding of the high number of plant aquaporin isoforms and multiple regulation mechanisms. Plant. Sci. 2017, 264, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.D.; Herman, E.M.; Chrispeels, M.J. An abundant, highly conserved tonoplast protein in seeds. Plant. Physiol. 1989, 91, 1006–1013. [Google Scholar] [CrossRef] [PubMed]
- Gattolin, S.; Sorieul, M.; Frigerio, L. Mapping of tonoplast intrinsic proteins in maturing and germinating Arabidopsis seeds reveals dual localization of embryonic TIPs to the tonoplast and plasma membrane. Mol. Plant. 2011, 4, 180–189. [Google Scholar] [CrossRef]
- Johnson, K.D.; Chrispeels, M.J. Tonoplast-bound protein kinase phosphorylates tonoplast intrinsic protein. Plant. Physiol. 1992, 100, 1787–1795. [Google Scholar] [CrossRef]
- Maurel, C.; Chrispeels, M.; Lurin, C.; Tacnet, F.; Geelen, D.; Ripoche, P.; Guern, J. Function and regulation of seed aquaporins. J. Exp. Bot. 1997, 48, 421–430. [Google Scholar] [CrossRef]
- Daniels, M.J.; Yeager, M. Phosphorylation of aquaporin PvTIP3;1 defined by mass spectrometry and molecular modeling. Biochemistry 2005, 44, 14443–14454. [Google Scholar] [CrossRef]
- Johnson, K.D.; Höfte, H.; Chrispeels, M.J. An intrinsic tonoplast protein of protein storage vacuoles in seeds is structurally related to a bacterial solute transporter (GlpF). Plant Cell 1990, 2, 525–532. [Google Scholar] [CrossRef]
- Ariani, A.; Gepts, P. Genome-wide identification and characterization of aquaporin gene family in common bean (Phaseolus vulgaris L.). Mol. Genet. Genomics 2015, 290, 1771–1785. [Google Scholar] [CrossRef] [PubMed]
- Vander Willigen, C.; Postaire, O.; Tournaire-Roux, C.; Boursiac, Y.; Maurel, C. Expression and inhibition of aquaporins in germinating Arabidopsis seeds. Plant Cell Physiol. 2006, 47, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Staehelin, L.A. Protein storage vacuoles are transformed into lytic vacuoles in root meristematic cells of germinating seedlings by multiple, cell type-specific mechanisms. Plant Physiol. 2011, 155, 2023–2035. [Google Scholar] [CrossRef] [PubMed]
- Weber, H.; Borisjuk, L.; Wobus, U. Molecular physiology of legume seed development. Ann. Rev. Plant Biol. 2005, 56, 253–279. [Google Scholar] [CrossRef]
- Daniels, M.J.; Chrispeels, M.J.; Yeager, M. Projection structure of a plant vacuole membrane aquaporin by electron cryo-crystallography. J. Mol. Biol. 1999, 294, 1337–1349. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.F.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Steinberg, T.H.; Agnew, B.J.; Gee, K.R.; Leung, W.-Y.; Goodman, T.; Schulenberg, B.; Hendrickson, J.; Beechem, J.M.; Haugland, R.P.; Patton, W.F. Global quantitative phosphoprotein analysis using multiplexed proteomics technology. Proteomics 2003, 3, 1128–1144. [Google Scholar] [CrossRef]
- Makrantoni, V.; Antrobus, R.; Botting, C.H.; Coote, P.J. Rapid enrichment and analysis of yeast phosphoproteins using affinity chromatography, 2D-PAGE and peptide mass fingerprinting. Yeast 2005, 22, 401–414. [Google Scholar] [CrossRef]
- Zhang, G.; Neubert, T.A. Use of detergents to increase selectivity of immunoprecipitation of tyrosine phosphorylated peptides prior to identification by MALDI quadrupole-TOF MS. Proteomics 2006, 6, 571–578. [Google Scholar] [CrossRef]
- Kelly, S.M.; Jess, T.J.; Price, N.C. How to study proteins by circular dichroism. Biochim. Biophys. Acta 2005, 1751, 119–139. [Google Scholar] [CrossRef]
- Pace, C.N.; Vajdos, F.; Fee, L.; Grimsley, G.; Gray, T. How to measure and predict the molar absorption coefficient of a protein. Protein Sci. 1995, 4, 2411–2423. [Google Scholar] [CrossRef] [Green Version]
- Greenfield, N.J. Using circular dichroism spectra to estimate protein secondary structure. Nat. Protoc. 2006, 1, 2876–2890. [Google Scholar] [CrossRef] [PubMed]
- Lees, J.G.; Smith, B.R.; Wien, F.; Miles, A.J.; Wallace, B.A. CDtool- an integrated software package for circular dichroism spectroscopic data processing, analysis, and archiving. Anal. Biochem. 2004, 332, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Krittanai, C.; Johnson, W.C., Jr. Correcting the circular dichroism spectra of peptides for contributions of absorbing side chains. Anal. Biochem. 1997, 253, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Sreerama, N.; Woody, R.W. Computation and analysis of protein circular dichroism spectra. Methods Enzymol. 2004, 383, 318–351. [Google Scholar]
- Whitmore, L.; Wallace, B.A. Protein secondary structure analyses from circular dichroism spectroscopy: Methods and reference databases. Biopolymers 2008, 89, 392–400. [Google Scholar] [CrossRef]
- Abdul-Gader, A.; Miles, A.J.; Wallace, B.A. A reference dataset for the analyses of membrane protein secondary structures and transmembrane residues using circular dichroism spectroscopy. Bioinformatics 2011, 27, 1630–1636. [Google Scholar] [CrossRef]
- Stankiewicz, P.J.; Tracey, A.S.; Crans, D.C. Inhibition of phosphate-metabolizing enzymes by oxovanadium(V) complexes. In Metal Ions in Biological Systems; Sigel, H., Sigel, A., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1995; Volume 31, pp. 287–324. [Google Scholar]
- Oufattole, M.; Park, J.H.; Poxleitner, M.; Jiang, L.; Rogers, J.C. Selective membrane protein internalization accompanies movement from the endoplasmic reticulum to the protein storage vacuole pathway in Arabidopsis. Plant Cell 2005, 17, 3066–3080. [Google Scholar] [CrossRef]
- Daniels, M.J.; Chaumont, F.; Mirkov, T.E.; Chrispeels, M.J. Characterization of a new vacuolar membrane aquaporin sensitive to mercury at a unique site. Plant Cell 1996, 8, 587–599. [Google Scholar]
- Hunter, P.R.; Craddock, C.P.; Di Benedetto, S.; Roberts, L.M.; Frigerio, L. Fluorescent reporter proteins for the tonoplast and the vacuolar lumen identify a single vacuolar compartment in Arabidopsis cells. Plant Physiol. 2007, 145, 1371–1382. [Google Scholar] [CrossRef]
- Utsugi, S.; Shibasaka, M.; Maekawa, M.; Katsuhara, M. Control of the water transport activity of barley HvTIP3;1 specifically expressed in seeds. Plant Cell Physiol. 2015, 56, 1831–1840. [Google Scholar] [CrossRef]
- Aebersold, R.; Patterson, S.D. Current Problems and Technical Solutions in Protein Biochemistry. In Proteins: Analysis and Design; Angeletti, R.H., Ed.; Academic Press: San Diego, CA, USA, 1998; pp. 4–120. [Google Scholar]
- Höfte, H.; Chrispeels, M.J. Protein sorting to the vacuolar membrane. Plant Cell 1992, 4, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C. Plant aquaporins: Novel functions and regulation properties. FEBS Lett. 2007, 581, 2227–2236. [Google Scholar] [CrossRef] [PubMed]
- Hjerrild, M.; Stensballe, A.; Rasmussen, T.E.; Kofoed, C.B.; Blom, N.; Sicheritz-Ponten, T.; Larsen, M.R.; Brunak, S.; Jensen, O.N.; Gammeltoft, S. Identification of phosphorylation sites in protein kinase A substrates using artificial neural networks and mass spectrometry. J. Proteome Res. 2004, 3, 426–433. [Google Scholar] [CrossRef]
- Bah, A.; Forman-Kay, J.D. Modulation of intrinsically disordered protein function by post-translational modifications. J. Biol. Chem. 2016, 291, 6696–6705. [Google Scholar] [CrossRef]
- Sreerama, N.; Woody, R.W. Poly(Pro)II helices in globular proteins: Identification and circular dichroic analysis. Biochemistry 1994, 33, 10022–10025. [Google Scholar] [CrossRef]
- Tiffany, M.L.; Krimm, S. Extended conformations of polypeptides and proteins in urea and guanidine hydrochloride. Biopolymers 1973, 12, 575–587. [Google Scholar] [CrossRef] [Green Version]
- Venyaminov, S.Y.; Baikalov, I.A.; Shen, Z.M.; Wu, C.-S.C.; Yang, J.T. Circular dichroic analysis of denatured proteins: Inclusion of denatured proteins in the reference set. Anal. Biochem. 1993, 214, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Lopes, J.L.S.; Miles, A.J.; Whitmore, L.; Wallace, B.A. Distinct circular dichroism spectroscopic signatures of polyproline II and unordered secondary structures: Applications in secondary structure analyses. Protein Sci. 2014, 23, 1765–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, E.L.; da Silva, A.C.; da Silva, M.J.; Leite, A.; Ottoboni, L.M.M. Endogenous protein phosphorylation and casein kinase activity during seed development in Araucaria angustifolia. Phytochemistry 2002, 61, 835–842. [Google Scholar] [CrossRef]
- Agrawal, G.K.; Thelen, J.J. Large scale identification and quantitative profiling of phosphoproteins expressed during seed filling in oilseed rape. Mol. Cell Proteomics 2006, 5, 2044–2059. [Google Scholar] [CrossRef]
- Mukhopadhyay, K.; Singh, M. Endogenous protein phosphorylation and protein kinase activity in winged bean. Phytochemistry 1997, 46, 455–459. [Google Scholar] [CrossRef]
- Kovaleva, V.; Cramer, R.; Krynytskyy, H.; Gout, I.; Gout, R. Analysis of tyrosine phosphorylation and phosphotyrosine-binding proteins in germinating seeds from Scots pine. Plant Physiol. Biochem. 2013, 67, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Herman, E.M.; Larkins, B.A. Protein storage bodies and vacuoles. Plant Cell 1999, 11, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Iakoucheva, L.M.; Radivojac, P.; Brown, C.J.; O’Connor, T.R.; Sikes, J.G.; Obradovic, Z.; Dunker, A.K. The importance of intrinsic disorder for protein phosphorylation. Nucleic Acids Res. 2004, 32, 1037–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manning, M.C.; Woody, R.W. Theoretical CD studies of polypeptide helices: Examination of important electronic and geometric factors. Biopolymers 1991, 31, 569–586. [Google Scholar] [CrossRef] [PubMed]
- Elam, W.A.; Schrank, T.P.; Campagnolo, A.J.; Hilser, V.J. Evolutionary conservation of the polyproline II conformation surrounding intrinsically disordered phosphorylation sites. Protein Sci. 2013, 22, 405–417. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-Y.; Jung, Y.; Hwang, G.-S.; Han, H.; Cho, M. Phosphorylation alters backbone conformational preferences of serine and threonine peptides. Proteins 2011, 79, 3155–3165. [Google Scholar] [CrossRef]
- Chaumont, F.; Moshelion, M.; Daniels, M.J. Regulation of plant aquaporin activity. Biol. Cell 2005, 97, 749–764. [Google Scholar] [CrossRef] [Green Version]
- Jolivet, P.; Roux, E.; d’Andrea, S.; Davanture, M.; Negroni, L.; Zivy, M.; Chardot, T. Protein composition of oil bodies in Arabidopsis thaliana ecotype WS. Plant Physiol. Biochem. 2004, 42, 501–509. [Google Scholar] [CrossRef]
- Li, G.-W.; Peng, Y.-H.; Yu, X.; Zhang, M.-H.; Cai, W.-M.; Sun, W.-N.; Su, W.-A. Transport functions and expression analysis of vacuolar membrane aquaporins in response to various stresses in rice. J. Plant Physiol. 2008, 165, 1879–1888. [Google Scholar] [CrossRef]
- Mao, Z.; Sun, W. Arabidopsis seed-specific vacuolar aquaporins are involved in maintaining seed longevity under the control of ABSCISIC ACID INSENSITIVE 3. J. Exp. Bot. 2015, 66, 4781–4794. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.M.; Birdsell, D.N.; Yool, A.J. The activity of human aquaporin 1 as a cGMP-gated cation channel is regulated by tyrosine phosphorylation in the carboxyl-terminal domain. Mol. Pharmacol. 2012, 81, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Yanochko, G.M.; Yool, A.J. Regulated cationic channel function in Xenopus oocytes expressing Drosophila Big Brain. J. Neurosci. 2002, 22, 2530–2540. [Google Scholar] [CrossRef] [PubMed]
- O’Rourke, J.A.; Iniguez, L.P.; Fu, F.; Bucciarelli, B.; Miller, S.S.; Jackson, S.A.; McClean, P.E.; Li, J.; Dai, X.; Zhao, P.X.; et al. An RNA-Seq based gene expression atlas of the common bean. BMC Genomics 2014, 15, 866–881. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.X.; Moon, S.; Jung, K.-H. Genome-wide expression analysis of rice aquaporin genes and development of a functional gene network mediated by aquaporin expression in roots. Planta 2013, 238, 669–681. [Google Scholar] [CrossRef]
- Melroy, D.L.; Herman, E.M. TIP, an integral membrane protein of the protein-storage vacuoles of the soybean cotyledon undergoes developmentally regulated membrane accumulation and removal. Planta 1991, 184, 113–122. [Google Scholar] [CrossRef]
- Robinson, D.G.; Hinz, G. Vacuole biogenesis and protein transport to the plant vacuole: A comparison with the yeast vacuole and the mammalian lysosome. Protoplasma 1997, 197, 1–25. [Google Scholar] [CrossRef]
- Strzalka, K.; Hara-Nishimura, I.; Nishimura, M. Changes in physical properties of vacuolar membrane during transformation of protein bodies into vacuoles in germinating pumpkin seeds. Biochim. Biophys. Acta 1995, 1239, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Capelle, V.; Remoué, C.; Moreau, L.; Reyss, A.; Mahé, A.; Massonneau, A.; Falque, M.; Charcosset, A.; Thévenot, C.; Rogowsky, P.; et al. QTLs and candidate genes for desiccation and abscisic acid content in maize kernels. BMC Plant. Biol. 2010, 10, 2–23. [Google Scholar] [CrossRef]
- Veselova, T.V.; Veselovskii, V.A.; Usmanov, P.D.; Usmanova, O.V.; Kozar, V.I. Hypoxia and imbibition injuries to aging seeds. Russ. J. Plant. Physiol 2003, 50, 835–842. [Google Scholar] [CrossRef]
- Zhou, Y.; Setz, N.; Niemietz, C.; Qu, H.; Offler, C.E.; Tyerman, S.D.; Patrick, J.W. Aquaporins and unloading of phloem-imported water in coats of developing bean seeds. Plant. Cell Environ. 2007, 30, 1566–1577. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| kD | Immature Seed | Mature Seed | Dry Seed | 24 h Imbibed | 48 h Imbibed |
|---|---|---|---|---|---|
| 70 | + | ++ | ++ | ++ | +++ |
| 67 | +++ | ++++ | ++ | ++ | +++ |
| 60 | - | + | ++ | + | ++ |
| 52 | ++++ | +++ | ++ | +++ | ++++ |
| 48 | ++++ | +++ | ++ | +++ | ++++ |
| 45 | + | + | ++ | ++ | +++ |
| 36 | + | - | - | - | - |
| 33 | + | + | - | + | + |
| 32 | + | ++ | + | + | + |
| 29 | ++ | ++ | + | + | ++ |
| 24 | ++++ | ++++ | ++++ | ++++ | ++++ |
| 22 | ++++ | ++++ | +++ | ++++ | ++++ |
| 19 | - | + | ++ | + | + |
| 15 | - | + | + | + | + |
| 12 | + | + | - | - | - |
| kD | Immature Seed | Mature Seed | Dry Seed | 24 h Imbibed | 48 h Imbibed |
|---|---|---|---|---|---|
| 24 | + | + | ++++ | ++++ | +++ |
| 19 | - | - | + | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daniels, M.J.; Yeager, M. Phosphorylation of TIP3 Aquaporins during Phaseolus vulgaris Embryo Development. Cells 2019, 8, 1362. https://doi.org/10.3390/cells8111362
Daniels MJ, Yeager M. Phosphorylation of TIP3 Aquaporins during Phaseolus vulgaris Embryo Development. Cells. 2019; 8(11):1362. https://doi.org/10.3390/cells8111362
Chicago/Turabian StyleDaniels, Mark J., and Mark Yeager. 2019. "Phosphorylation of TIP3 Aquaporins during Phaseolus vulgaris Embryo Development" Cells 8, no. 11: 1362. https://doi.org/10.3390/cells8111362
APA StyleDaniels, M. J., & Yeager, M. (2019). Phosphorylation of TIP3 Aquaporins during Phaseolus vulgaris Embryo Development. Cells, 8(11), 1362. https://doi.org/10.3390/cells8111362