Dyskerin Mutations Present in Dyskeratosis Congenita Patients Increase Oxidative Stress and DNA Damage Signalling in Dictyostelium Discoideum


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell culture and Transformation
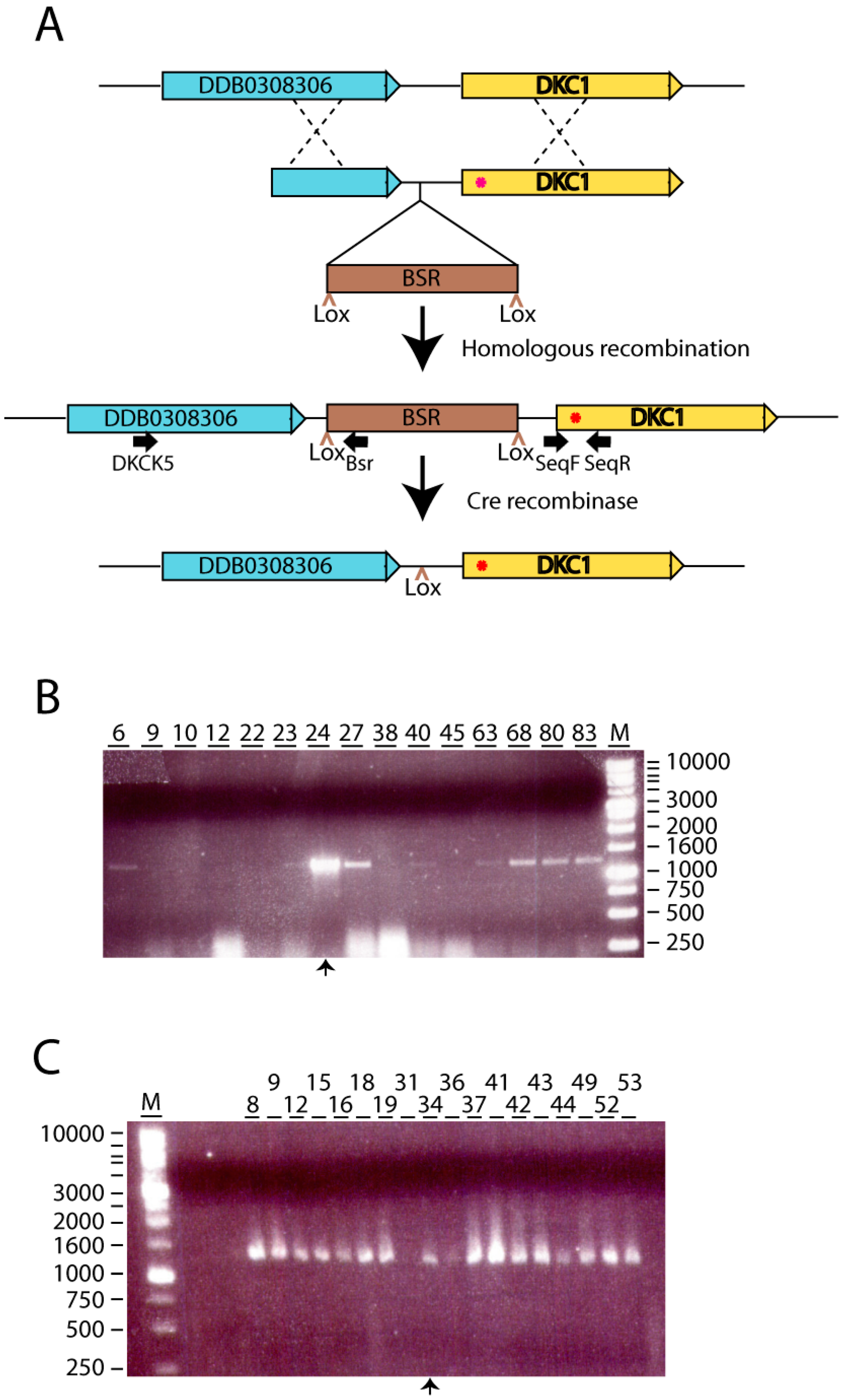
2.2. Generation of dkc1 Knockin Mutant Strains.
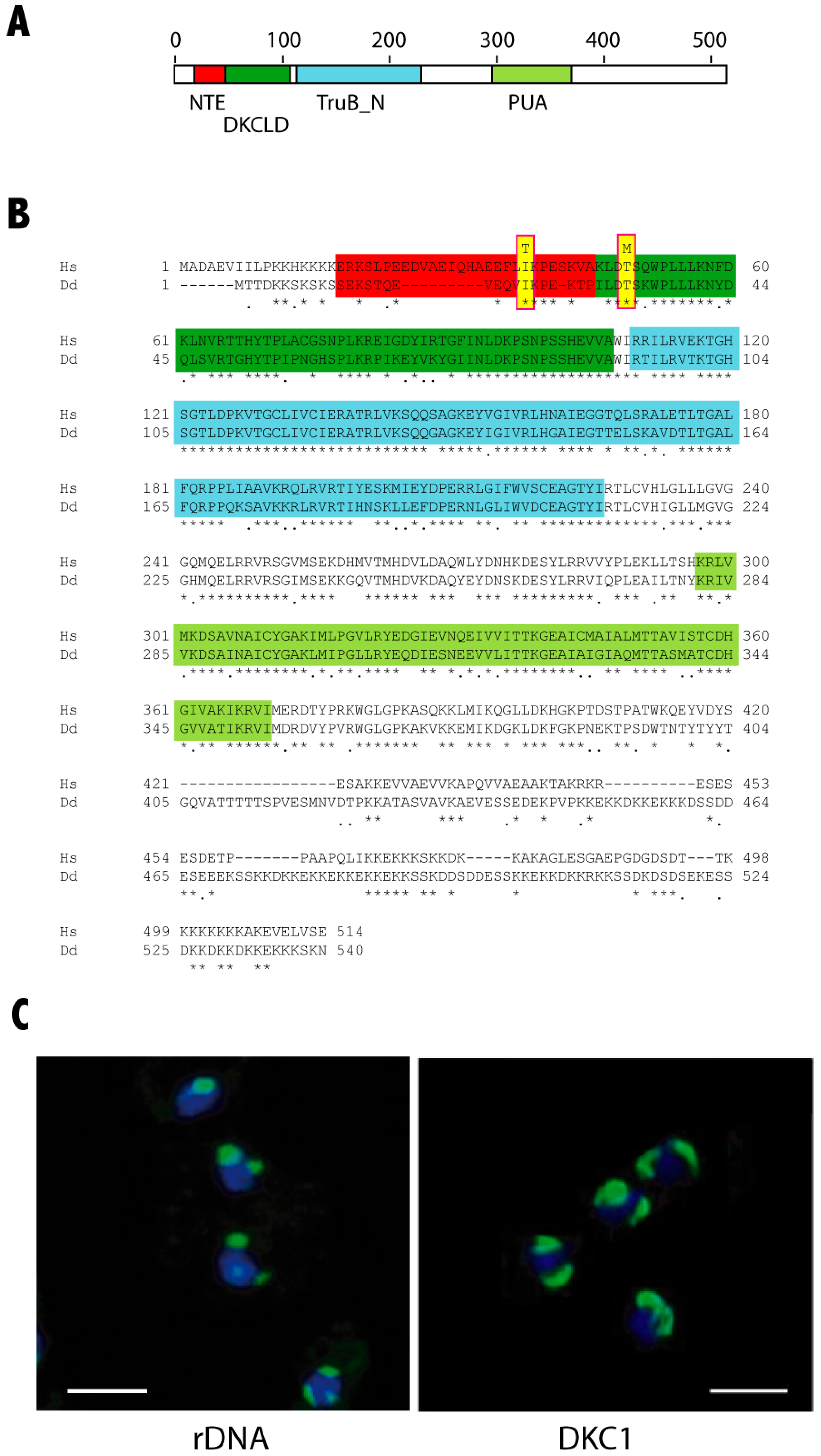
2.3. DKC1 and Nucleoli Localization
2.4. Pseudouridylation Assay
2.5. Quantitative PCR
2.6. Southern Blot
2.7. DNA Damage
2.8. ROS Assay
3. Results
3.1. Generation of D. Discoideum dkc1 Mutant Strains
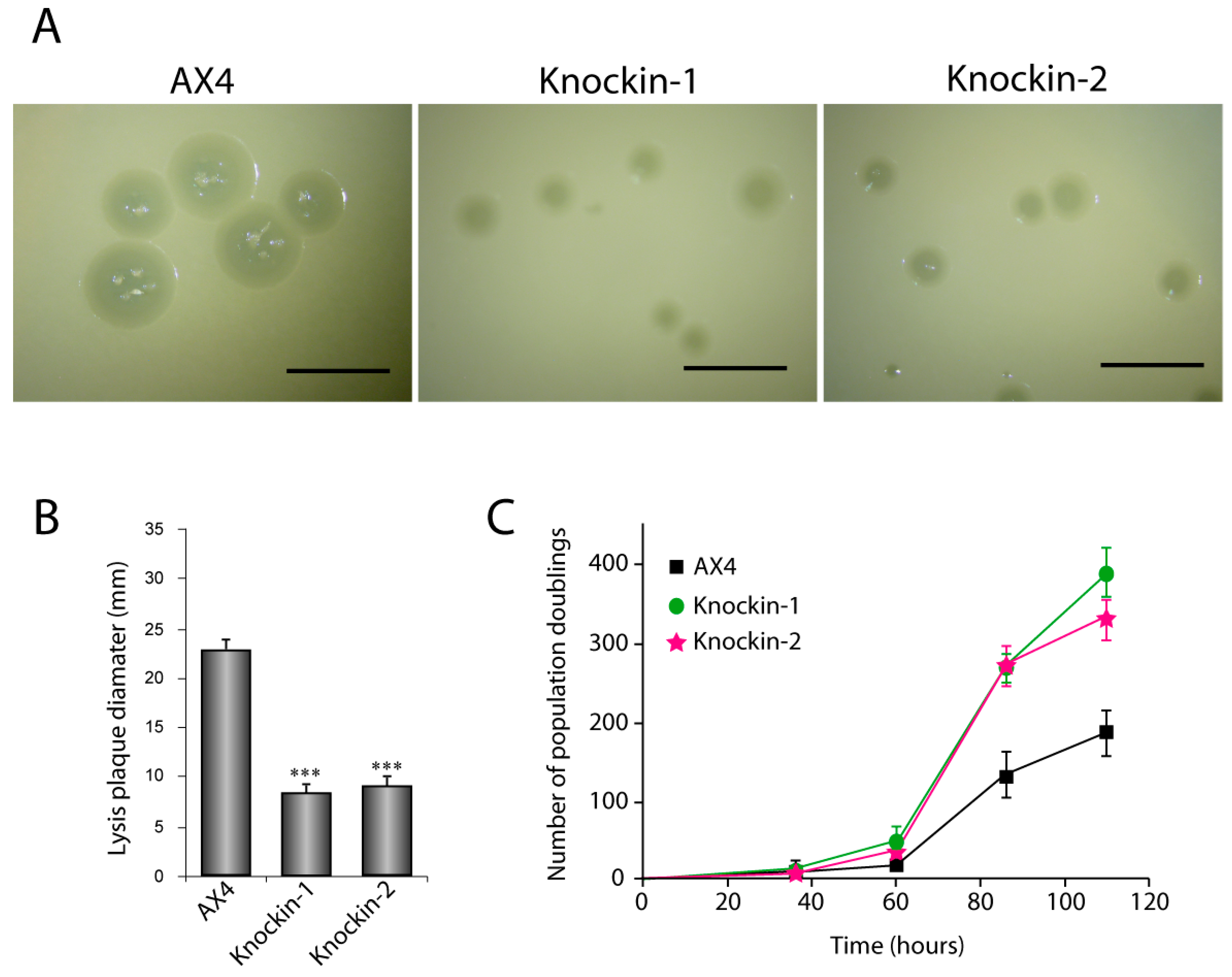
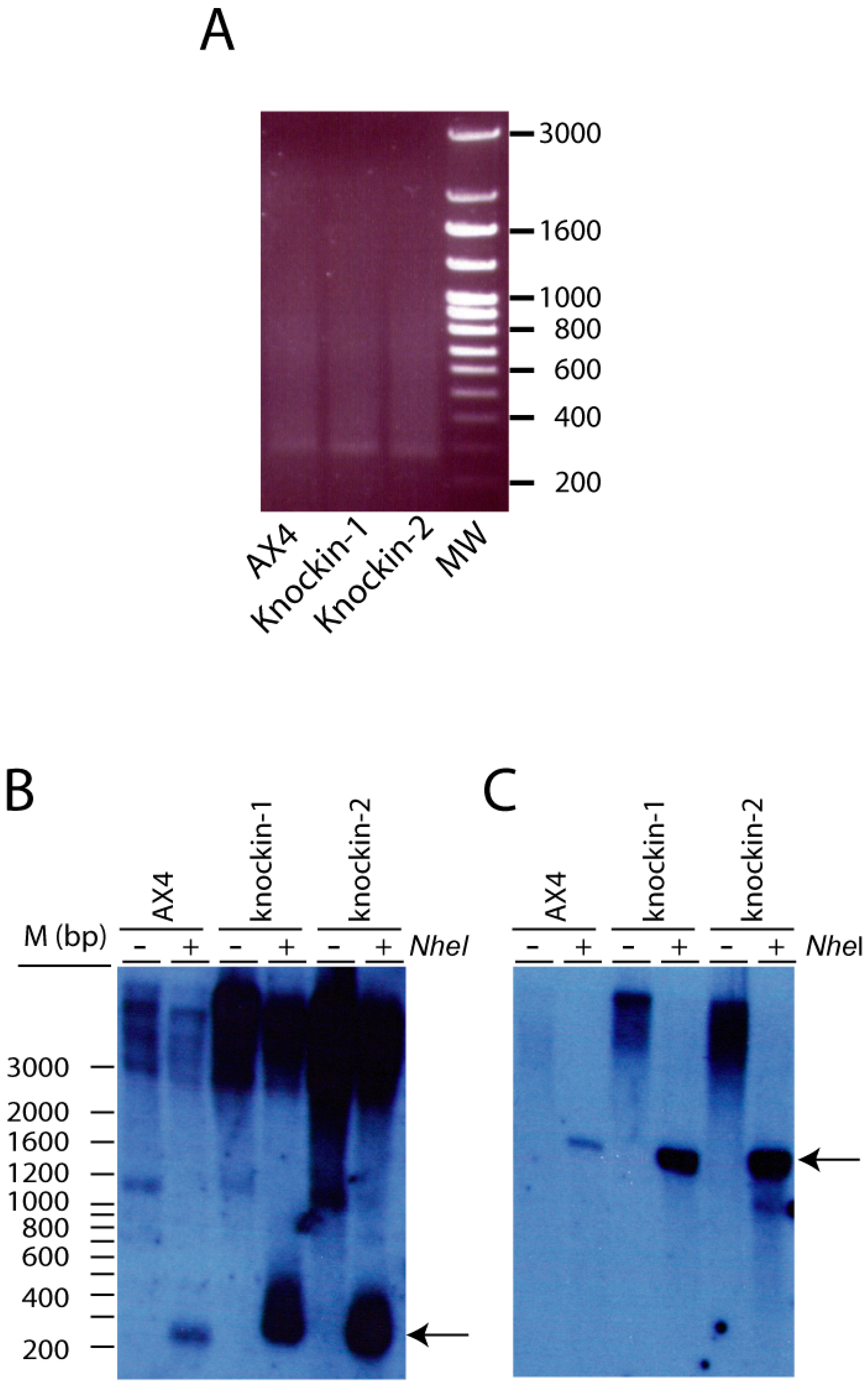
3.2. Proliferation and Telomere Structure of dkc1 Mutant Strains
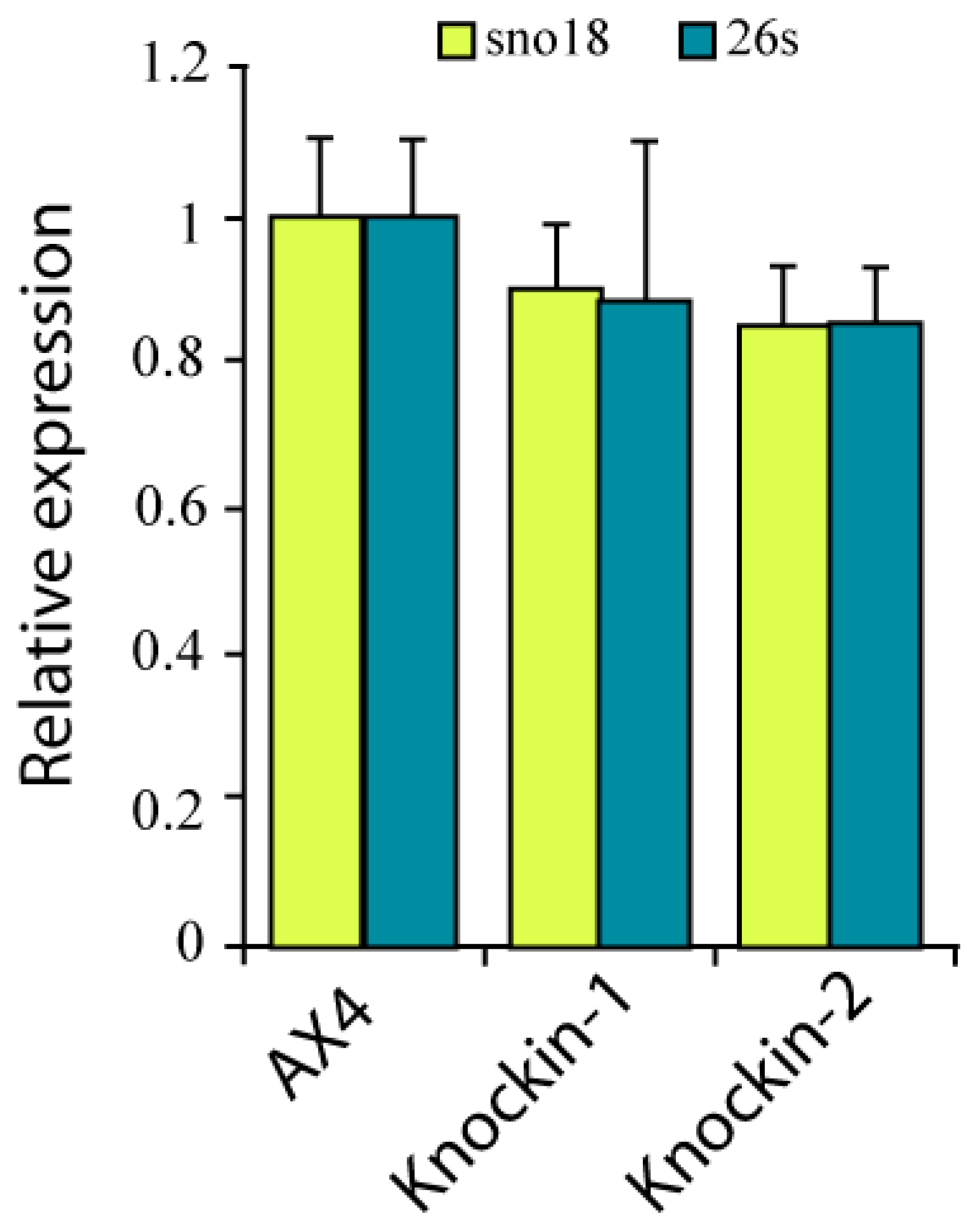
3.3. RNA Pseudouridylation in dkc1 Mutant Strains
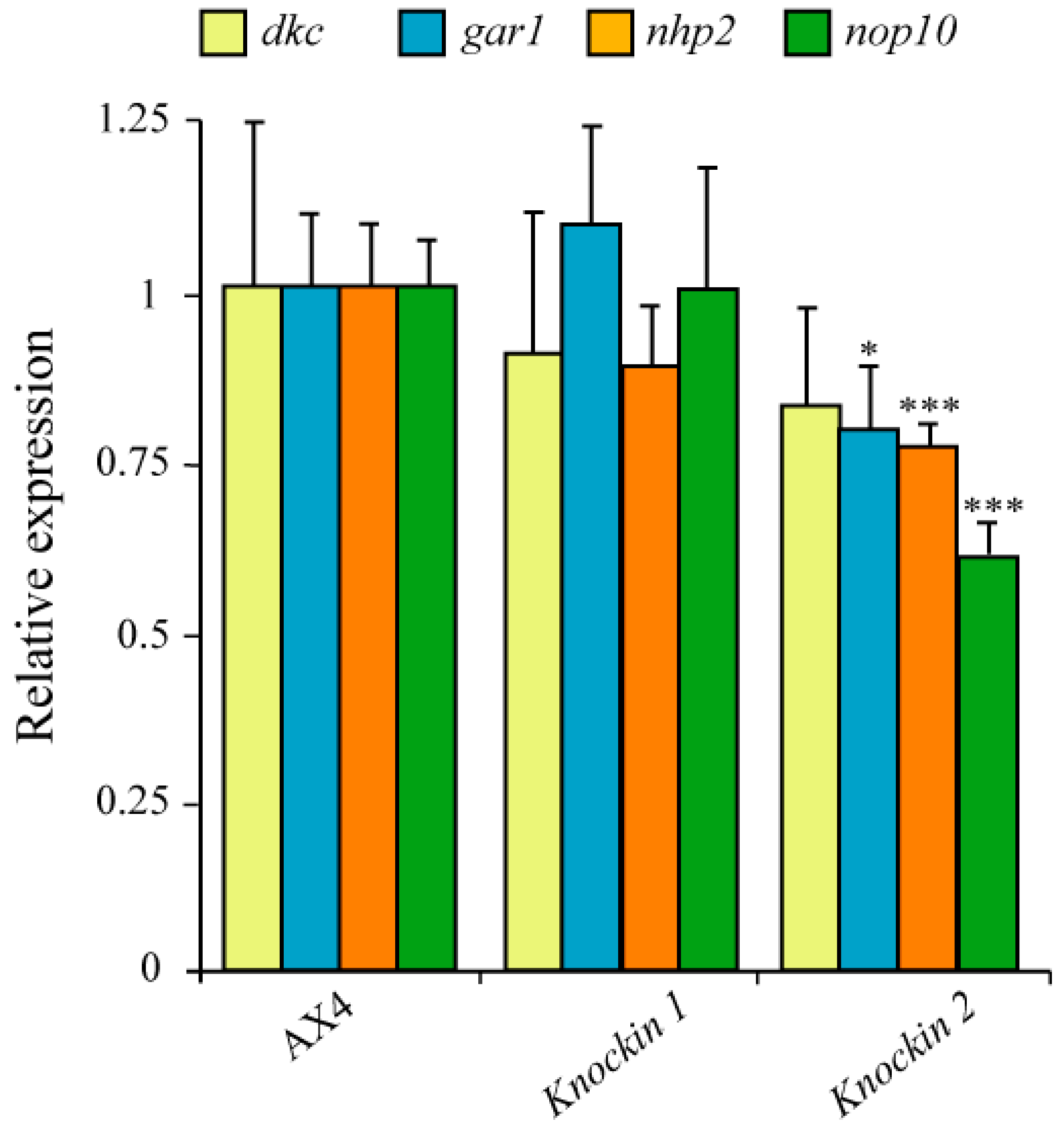
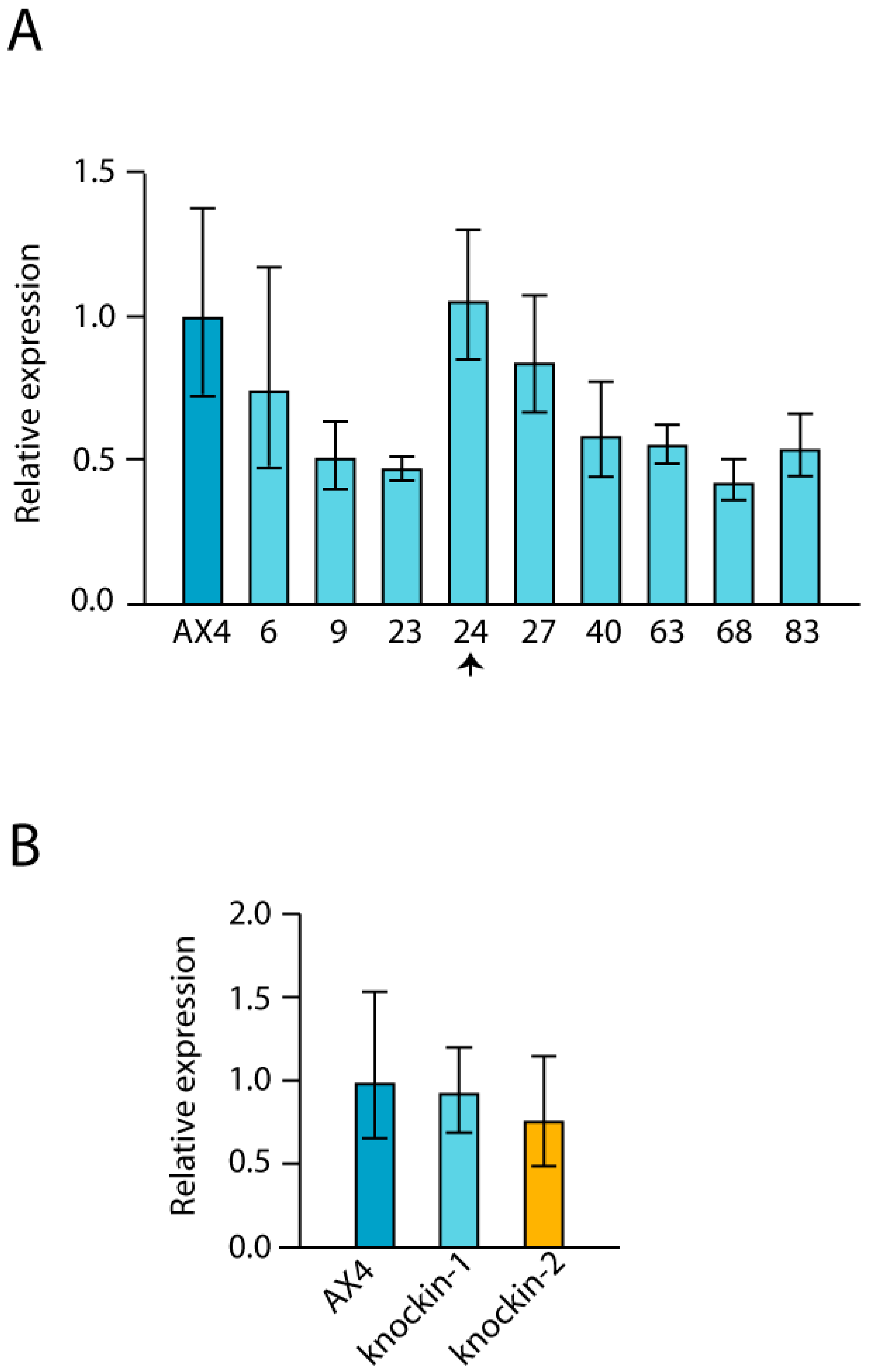
3.4. Expression Levels of GAR1, NHP2 and NOP10 in dkc1 Mutant Strains
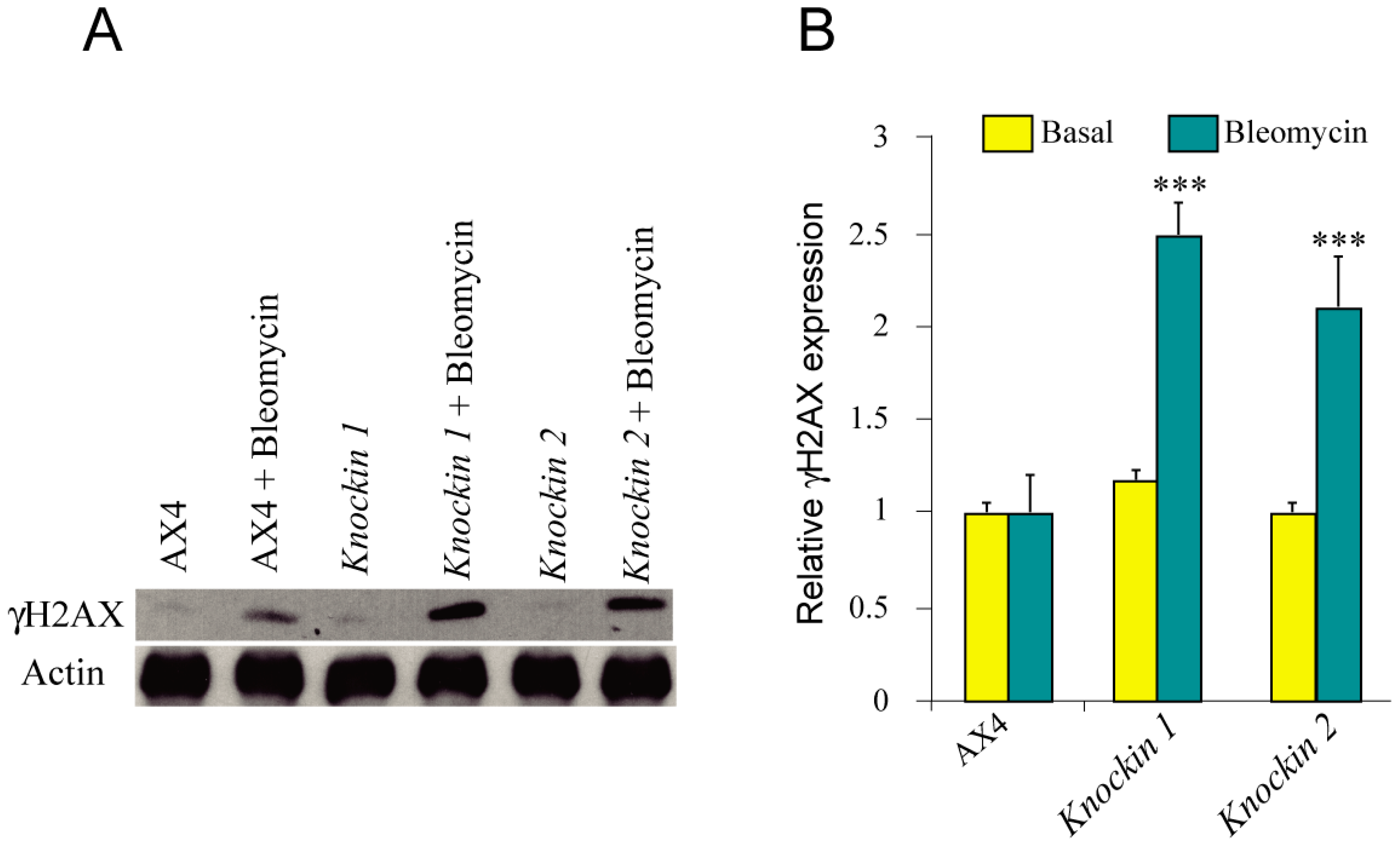
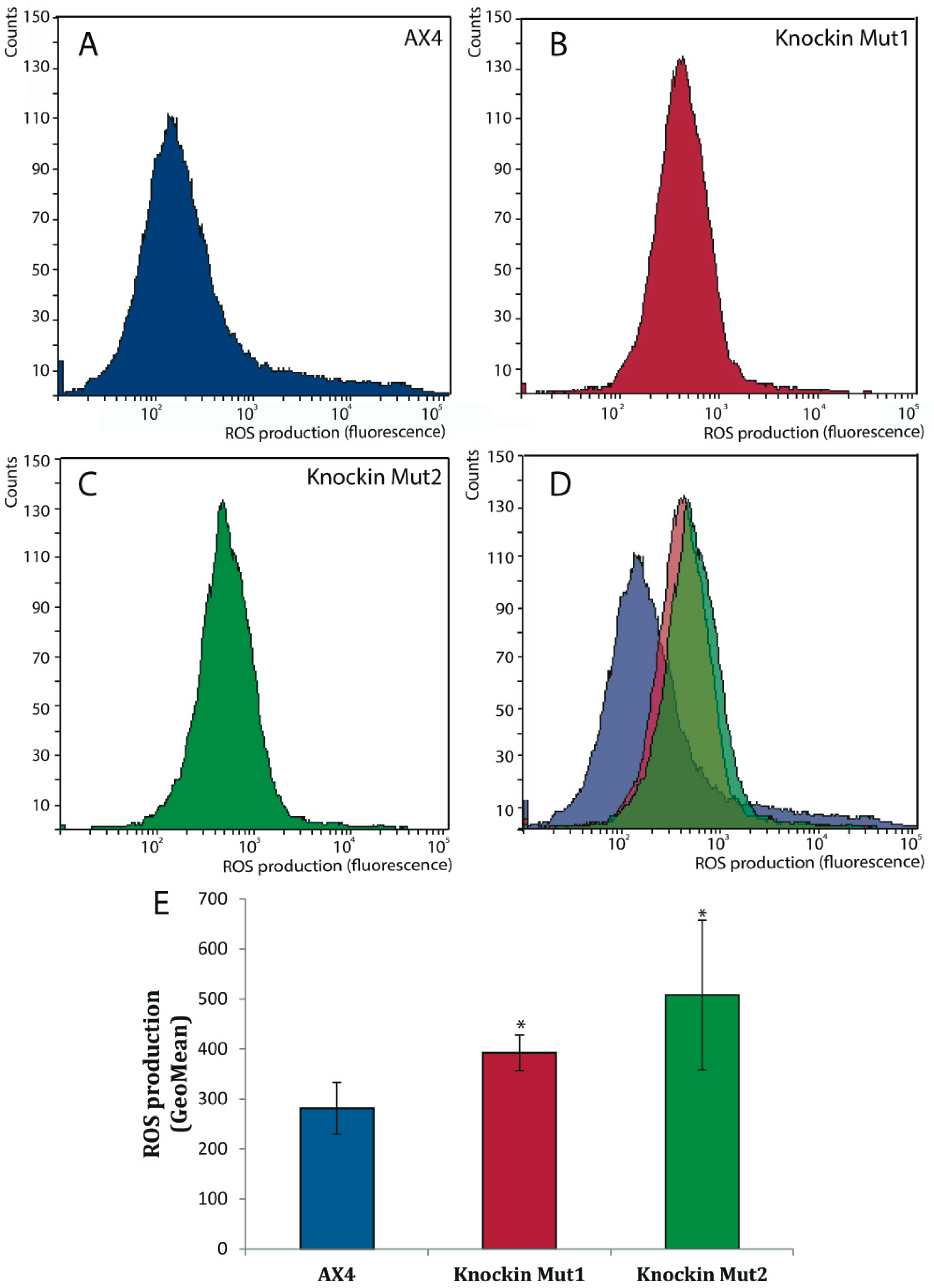
3.5. DNA-Damage Response and Oxidative Stress of dkc1 Mutant Strains
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

References
- Kiss, T.; Fayet, E.; Jady, B.E.; Richard, P.; Weber, M. Biogenesis and intranuclear trafficking of human box C/D and H/ACA RNPs. Cold Spring Harb Symp. Quant. Biol. 2006, 71, 407–417. [Google Scholar] [CrossRef]
- Schwartz, S.; Bernstein, D.A.; Mumbach, M.R.; Jovanovic, M.; Herbst, R.H.; Leon-Ricardo, B.X.; Engreitz, J.M.; Guttman, M.; Satija, R.; Lander, E.S.; et al. Transcriptome-wide mapping reveals widespread dynamic-regulated pseudouridylation of ncRNA and mRNA. Cell 2014, 159, 148–162. [Google Scholar] [CrossRef]
- Lafontaine, D.L.; Tollervey, D. Birth of the snoRNPs: The evolution of the modification-guide snoRNAs. Trends Biochem. Sci. 1998, 23, 383–388. [Google Scholar] [CrossRef]
- Lestrade, L.; Weber, M.J. snoRNA-LBME-db, a comprehensive database of human H/ACA and C/D box snoRNAs. Nucleic Acids Res. 2006, 34, D158–D162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, P.; Darzacq, X.; Bertrand, E.; Jady, B.E.; Verheggen, C.; Kiss, T. A common sequence motif determines the Cajal body-specific localization of box H/ACA scaRNAs. EMBO J. 2003, 22, 4283–4293. [Google Scholar] [CrossRef] [PubMed]
- Khanna, A.; Stamm, S. Regulation of alternative splicing by short non-coding nuclear RNAs. RNA Biol. 2010, 7, 480–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taft, R.J.; Glazov, E.A.; Lassmann, T.; Hayashizaki, Y.; Carninci, P.; Mattick, J.S. Small RNAs derived from snoRNAs. RNA 2009, 15, 1233–1240. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, J.R.; Cheng, J.; Collins, K. A box H/ACA small nucleolar RNA-like domain at the human telomerase RNA 3’ end. Mol. Cell. Biol. 1999, 19, 567–576. [Google Scholar] [CrossRef]
- Cohen, S.B.; Graham, M.E.; Lovrecz, G.O.; Bache, N.; Robinson, P.J.; Reddel, R.R. Protein composition of catalytically active human telomerase from immortal cells. Science 2007, 315, 1850–1853. [Google Scholar] [CrossRef]
- Nguyen, T.H.D.; Tam, J.; Wu, R.A.; Greber, B.J.; Toso, D.; Nogales, E.; Collins, K. Cryo-EM structure of substrate-bound human telomerase holoenzyme. Nature 2018, 557, 190–195. [Google Scholar] [CrossRef]
- Blackburn, E.H.; Greider, C.W.; Szostak, J.W. Telomeres and telomerase: The path from maize, Tetrahymena and yeast to human cancer and aging. Nat. Med. 2006, 12, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Harley, C.B.; Futcher, A.B.; Greider, C.W. Telomeres shorten during ageing of human fibroblasts. Nature 1990, 345, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Martinez, P.; Blasco, M.A. Replicating through telomeres: A means to an end. Trends Biochem. Sci. 2015, 40, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Angrisani, A.; Vicidomini, R.; Turano, M.; Furia, M. Human dyskerin: Beyond telomeres. Biol. Chem. 2014, 395, 593–610. [Google Scholar] [CrossRef]
- MacNeil, D.E.; Lambert-Lanteigne, P.; Autexier, C. N-terminal residues of human dyskerin are required for interactions with telomerase RNA that prevent RNA degradation. Nucleic Acids Res. 2019, 47, 5368–5380. [Google Scholar] [CrossRef] [Green Version]
- Perona, R.; Iarriccio, L.; Pintado-Berninches, L.; Rodriguez-Centeno, J.; Manguan-Garcia, C.; Garcia, E.; Lopez-Ayllon, B.D.; Sastre, L. Molecular diagnosis and precission therapeutic approaches for telomere biology disorders. In Telomeres; Larramendy, M., Soloneski, S., Eds.; INTECH: Rijeka, Croatia, 2016; pp. 77–117. [Google Scholar]
- Savage, S.A. Beginning at the ends: Telomeres and human disease. F1000Res 2018, 7. [Google Scholar] [CrossRef]
- Jiang, W.; Middleton, K.; Yoon, H.J.; Fouquet, C.; Carbon, J. An essential yeast protein, CBF5p, binds in vitro to centromeres and microtubules. Mol. Cell. Biol. 1993, 13, 4884–4893. [Google Scholar] [CrossRef]
- Giordano, E.; Peluso, I.; Senger, S.; Furia, M. minifly, a Drosophila gene required for ribosome biogenesis. J. Cell Biol. 1999, 144, 1123–1133. [Google Scholar] [CrossRef]
- Li, L.; Ye, K. Crystal structure of an H/ACA box ribonucleoprotein particle. Nature 2006, 443, 302–307. [Google Scholar] [CrossRef]
- Mitchell, J.R.; Wood, E.; Collins, K. A telomerase component is defective in the human disease dyskeratosis congenita. Nature 1999, 402, 551–555. [Google Scholar] [CrossRef]
- Bertuch, A.A. The Molecular Genetics of the Telomere Biology Disorders. RNA Biol. 2016, 13, 696. [Google Scholar] [CrossRef] [PubMed]
- Eichinger, L.; Pachebat, J.A.; Glockner, G.; Rajandream, M.; Sucgang, R.; Berriman, M.; Song, J.; Olsen, R.; Szafranski, K.; Xu, Q. The genome of the social amoeba Dictyostelium discoideum. Nature 2005, 435, 43–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loomis, W.F. Cell signaling during development of Dictyostelium. Dev. Biol. 2014, 391, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carnell, M.J.; Insall, R.H. Actin on disease—Studying the pathobiology of cell motility using Dictyostelium discoideum. Semin. Cell Dev. Biol. 2011. [Google Scholar] [CrossRef]
- Rodriguez-Centeno, J.; Manguan-Garcia, C.; Perona, R.; Sastre, L. Structure of Dictyostelium discoideum telomeres. Analysis of possible replication mechanisms. PLoS ONE 2019, 14, e0222909. [Google Scholar] [CrossRef] [PubMed]
- Pang, K.M.; Lynes, M.A.; Knecht, D.A. Variables controlling the expression level of exogenous genes in Dictyostelium. Plasmid 1999, 41, 187–197. [Google Scholar] [CrossRef]
- Faix, J.; Kreppel, L.; Shaulsky, G.; Schleicher, M.; Kimmel, A.R. A rapid and efficient method to generate multiple gene disruptions in Dictyostelium discoideum using a single selectable marker and the Cre-loxP system. Nucleic Acids Res. 2004, 32, e143. [Google Scholar] [CrossRef]
- Meima, M.E.; Weening, K.E.; Schaap, P. Vectors for expression of proteins with single or combinatorial fluorescent protein and tandem affinity purification tags in Dictyostelium. Protein Expr. Purif. 2007, 53, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Bakin, A.; Ofengand, J. Four newly located pseudouridylate residues in Escherichia coli 23S ribosomal RNA are all at the peptidyltransferase center: Analysis by the application of a new sequencing technique. Biochemistry 1993, 32, 9754–9762. [Google Scholar] [CrossRef]
- Galardi-Castilla, M.; Pergolizzi, B.; Bloomfield, G.; Skelton, J.; Ivens, A.; Kay, R.R.; Bozzaro, S.; Sastre, L. SrfB, a member of the Serum Response Factor family of transcription factors, regulates starvation response and early development in Dictyostelium. Dev. Biol. 2008, 316, 260–274. [Google Scholar] [CrossRef] [Green Version]
- Galardi-Castilla, M.; Fernandez-Aguado, I.; Suarez, T.; Sastre, L. Mef2A, a homologue of animal Mef2 transcription factors, regulates cell differentiation in Dictyostelium discoideum. BMC Dev. Biol. 2013, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.E.; Welker, D.L. A mini-screen technique for analyzing nuclear DNA from a single Dictyostelium colony. Nucl. Acids Res. 1988, 16, 2338. [Google Scholar] [CrossRef] [PubMed]
- Cossu, F.; Vulliamy, T.J.; Marrone, A.; Badiali, M.; Cao, A.; Dokal, I. A novel DKC1 mutation, severe combined immunodeficiency (T+B-NK-SCID) and bone marrow transplantation in an infant with Hoyeraal-Hreidarsson syndrome. Br. J. Haematol. 2002, 119, 765–768. [Google Scholar] [CrossRef] [PubMed]
- Knight, S.W.; Heiss, N.S.; Vulliamy, T.J.; Greschner, S.; Stavrides, G.; Pai, G.S.; Lestringant, G.; Varma, N.; Mason, P.J.; Dokal, I.; et al. X-linked dyskeratosis congenita is predominantly caused by missense mutations in the DKC1 gene. Am. J. Hum. Genet. 1999, 65, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Martinez, P.; Flores, J.M.; Blasco, M.A. 53BP1 deficiency combined with telomere dysfunction activates ATR-dependent DNA damage response. J. Cell Biol. 2012, 197, 283–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Capetillo, O.; Chen, H.T.; Celeste, A.; Ward, I.; Romanienko, P.J.; Morales, J.C.; Naka, K.; Xia, Z.; Camerini-Otero, R.D.; Motoyama, N.; et al. DNA damage-induced G2-M checkpoint activation by histone H2AX and 53BP1. Nat. Cell Biol. 2002, 4, 993–997. [Google Scholar] [CrossRef]
- Gu, B.W.; Fan, J.M.; Bessler, M.; Mason, P.J. Accelerated hematopoietic stem cell aging in a mouse model of dyskeratosis congenita responds to antioxidant treatment. Aging Cell 2011, 10, 338–348. [Google Scholar] [CrossRef] [Green Version]
- Manguan-Garcia, C.; Pintado-Berninches, L.; Carrillo, J.; Machado-Pinilla, R.; Sastre, L.; Perez-Quilis, C.; Esmoris, I.; Gimeno, A.; Garcia-Gimenez, J.L.; Pallardo, F.V.; et al. Expression of the genetic suppressor element 24.2 (GSE24.2) decreases DNA damage and oxidative stress in X-linked dyskeratosis congenita cells. PLoS ONE 2014, 9, e101424. [Google Scholar] [CrossRef]
- Ahmed, W.; Lingner, J. Impact of oxidative stress on telomere biology. Differentiation 2018, 99, 21–27. [Google Scholar] [CrossRef]
- Marrone, A.; Walne, A.; Dokal, I. Dyskeratosis congenita: Telomerase, telomeres and anticipation. Curr. Opin. Genet. Dev. 2005, 15, 249–257. [Google Scholar] [CrossRef]
- Grozdanov, P.N.; Fernandez-Fuentes, N.; Fiser, A.; Meier, U.T. Pathogenic NAP57 mutations decrease ribonucleoprotein assembly in dyskeratosis congenita. Hum. Mol. Genet. 2009, 18, 4546–4551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vulliamy, T.J.; Dokal, I. Dyskeratosis congenita: The diverse clinical presentation of mutations in the telomerase complex. Biochimie 2008, 90, 122–130. [Google Scholar] [CrossRef]
- Gu, B.W.; Bessler, M.; Mason, P.J. A pathogenic dyskerin mutation impairs proliferation and activates a DNA damage response independent of telomere length in mice. Proc. Natl. Acad. Sci. USA 2008, 105, 10173–10178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Morimoto, K.; Danilova, N.; Zhang, B.; Lin, S. Zebrafish models for dyskeratosis congenita reveal critical roles of p53 activation contributing to hematopoietic defects through RNA processing. PLoS ONE 2012, 7, e30188. [Google Scholar] [CrossRef] [PubMed]
- Di Maio, N.; Vicidomini, R.; Angrisani, A.; Belli, V.; Furia, M.; Turano, M. A new role for human dyskerin in vesicular trafficking. FEBS Open Bio. 2017, 7, 1453–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angrisani, A.; Matrone, N.; Belli, V.; Vicidomini, R.; Di Maio, N.; Turano, M.; Scialo, F.; Netti, P.A.; Porcellini, A.; Furia, M. A functional connection between dyskerin and energy metabolism. Redox Biol. 2017, 14, 557–565. [Google Scholar] [CrossRef]
- Fong, Y.W.; Ho, J.J.; Inouye, C.; Tjian, R. The dyskerin ribonucleoprotein complex as an OCT4/SOX2 coactivator in embryonic stem cells. Elife 2014, 3, e03573. [Google Scholar] [CrossRef]
- Vicidomini, R.; Petrizzo, A.; di Giovanni, A.; Cassese, L.; Lombardi, A.A.; Pragliola, C.; Furia, M. Drosophila dyskerin is required for somatic stem cell homeostasis. Sci. Rep. 2017, 7, 347. [Google Scholar] [CrossRef]
- Fey, P.; Kowal, A.S.; Gaudet, P.; Pilcher, K.E.; Chisholm, R.L. Protocols for growth and development of Dictyostelium discoideum. Nat. Protoc. 2007, 2, 1307–1316. [Google Scholar] [CrossRef]
- Bozzaro, S.; Bucci, C.; Steinert, M. Phagocytosis and host-pathogen interactions in Dictyostelium with a look at macrophages. Int. Rev. Cell Mol. Biol. 2008, 271, 253–300. [Google Scholar] [CrossRef]
- Batista, L.F.; Pech, M.F.; Zhong, F.L.; Nguyen, H.N.; Xie, K.T.; Zaug, A.J.; Crary, S.M.; Choi, J.; Sebastiano, V.; Cherry, A.; et al. Telomere shortening and loss of self-renewal in dyskeratosis congenita induced pluripotent stem cells. Nature 2011, 474, 399–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tortoriello, G.; de Celis, J.F.; Furia, M. Linking pseudouridine synthases to growth, development and cell competition. FEBS J. 2010, 277, 3249–3263. [Google Scholar] [CrossRef] [PubMed]
- Iarriccio, L.; Manguan-Garcia, C.; Pintado-Berninches, L.; Mancheno, J.M.; Molina, A.; Perona, R.; Sastre, L. GSE4, a Small Dyskerin- and GSE24.2-Related Peptide, Induces Telomerase Activity, Cell Proliferation and Reduces DNA Damage, Oxidative Stress and Cell Senescence in Dyskerin Mutant Cells. PLoS ONE 2015, 10, e0142980. [Google Scholar] [CrossRef] [PubMed]
- von Zglinicki, T. Oxidative stress shortens telomeres. Trends Biochem. Sci. 2002, 27, 339–344. [Google Scholar] [CrossRef]
- Tan, R.; Lan, L. Guarding chromosomes from oxidative DNA damage to the very end. Acta Biochim. Biophys. Sin. (Shanghai) 2016, 48, 617–622. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, J.; Gonzalez, A.; Manguan-Garcia, C.; Pintado-Berninches, L.; Perona, R. p53 pathway activation by telomere attrition in X-DC primary fibroblasts occurs in the absence of ribosome biogenesis failure and as a consequence of DNA damage. Clin. Transl. Oncol. 2013, 16, 529–538. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′-3′) |
|---|---|---|
| dkc1 | DKC1F | GGAAGCTTATGACAACTGGTATGTGTGAAA |
| DKC1R | GGTCTAGAATTCTTTGATTTCTTTTTTTCTT | |
| Mut1dkc1F | CAAGTTACTAAACCAGAAAAAACCC | |
| Mut1dkc1R | CTGGTTTAGTAACTTGTTCAACTTC | |
| Mut2dkc1F | CAATTTTAGATATGAGTAAATGGCCATTATTA | |
| Mut2dkc1R | GCCATTTACTCATATCTAAAATTGGGGTTTTT | |
| Arm1F | GGGGGCCCGGGTGTTGTAGAGATGCAAGATGG | |
| Arm1R | GGGTCGACGGATTTCTTGATGAAATACTAGGAG | |
| Arm2F | GGCTGCAGCTCCTAGTATTTCATCAAGAAATCC | |
| Arm2R | GGAAGCTTTGTTGAAATTTCTTATTTTAAATAA | |
| DKCK5 | CATCACCATTGAAGAGGGT | |
| BsR | CCAACCAAGTTTTTTTAAACC | |
| SeqF | GGAAGCTTATGACAACTGGTATGTGTGAAA | |
| SeqR | GGAATTGGTGTATAATGACCAG | |
| PCRDKC1F | GATGACTCTGATGATGAATCATCC | |
| PCRDKC1R | CCCACAAATTTATAAAGTTTATTTTTATTAATTAG | |
| sno18 | Sno18F | CAGCCTGCCTCCGTTTTGTGTTTG |
| Sno18R | GATTACAGATTGATTAAAGGCACATGTATG | |
| 26S rRNA | 26srRNAF | CCGCTGAACTTAAGCATATCAGTAAGC |
| 26srRNAR | GCAGTCACAACAGCGGGCTCC | |
| gar1 | Gar1F | CAGTTGATGAAATTTTCGGACC |
| Gar1R | GGTACTTTAGCAATTGGTTTTGG | |
| nhp2 | Nhp2F | GGCGATGTCAGCCCAATCG |
| Nhp2R | GTTGATGAAGCAGTACCTAAAG | |
| nop10 | Nop10F | GTACTATAACGACAAAGATGGCC |
| Nop10R | CCAAATCTCTTCTTTAATGCAATTC | |
| lmtrRNA | lmtrRNAF | GGGTAGTTTGACTGGGGCGG |
| lmtrRNAR | CACTTTAATGGGTGAACACC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez-Centeno, J.; Perona, R.; Sastre, L. Dyskerin Mutations Present in Dyskeratosis Congenita Patients Increase Oxidative Stress and DNA Damage Signalling in Dictyostelium Discoideum. Cells 2019, 8, 1406. https://doi.org/10.3390/cells8111406
Rodriguez-Centeno J, Perona R, Sastre L. Dyskerin Mutations Present in Dyskeratosis Congenita Patients Increase Oxidative Stress and DNA Damage Signalling in Dictyostelium Discoideum. Cells. 2019; 8(11):1406. https://doi.org/10.3390/cells8111406
Chicago/Turabian StyleRodriguez-Centeno, Javier, Rosario Perona, and Leandro Sastre. 2019. "Dyskerin Mutations Present in Dyskeratosis Congenita Patients Increase Oxidative Stress and DNA Damage Signalling in Dictyostelium Discoideum" Cells 8, no. 11: 1406. https://doi.org/10.3390/cells8111406
APA StyleRodriguez-Centeno, J., Perona, R., & Sastre, L. (2019). Dyskerin Mutations Present in Dyskeratosis Congenita Patients Increase Oxidative Stress and DNA Damage Signalling in Dictyostelium Discoideum. Cells, 8(11), 1406. https://doi.org/10.3390/cells8111406