AQP1-Containing Exosomes in Peritoneal Dialysis Effluent As Biomarker of Dialysis Efficiency

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples Collection
2.2. Measurements
2.3. Calculations
2.4. Antibodies
2.5. Immunofluorescence
2.6. Purification of Exosomes from Peritoneal Dialysate
2.7. Immunoblotting
2.8. Immunoelectron Microscopy
2.9. ELISA Test for AQP1 Quantitation in Peritoneal Dialysate Samples
3. Results
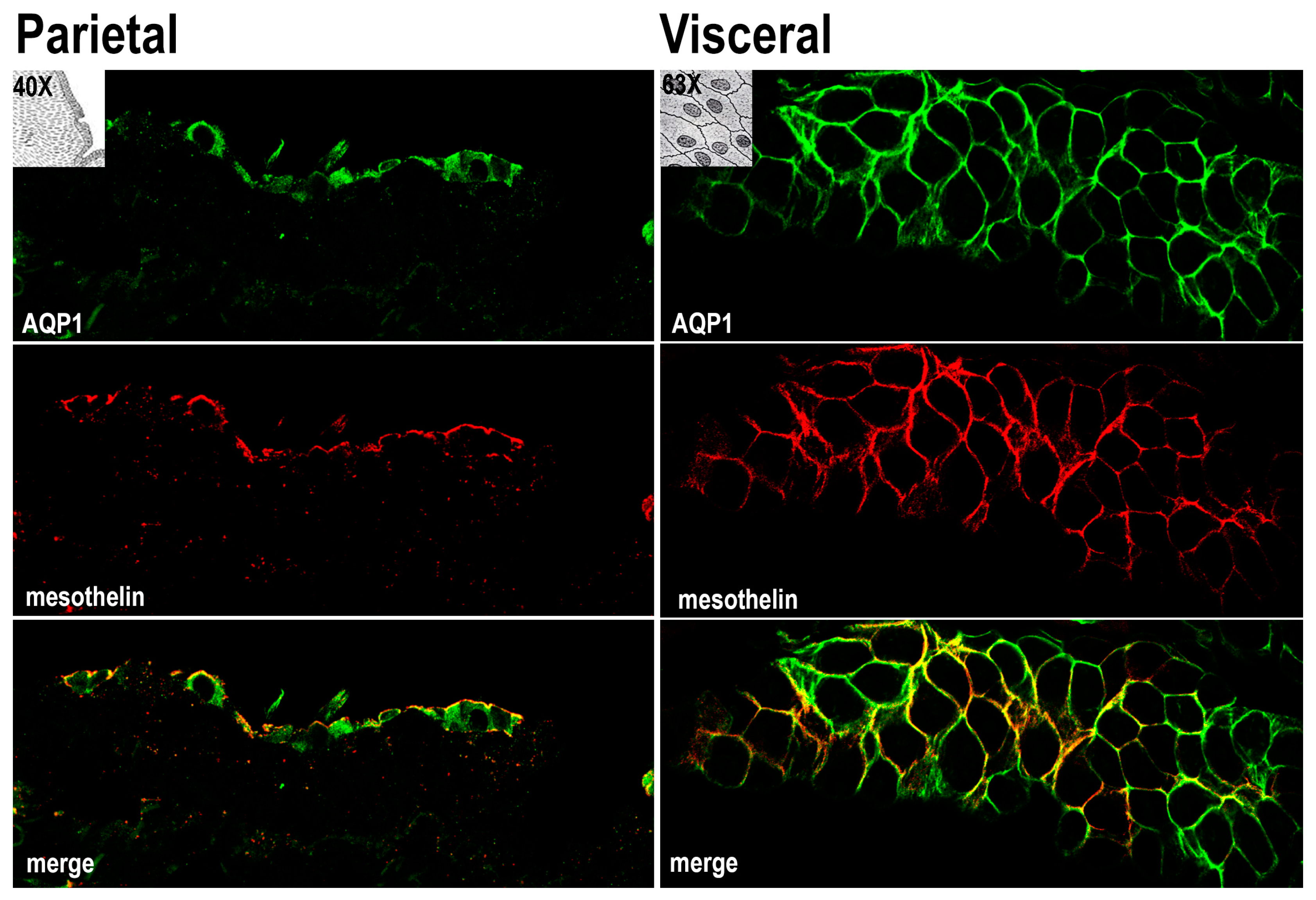
3.1. AQP1 Is Expressed in Human Mesothelial Cells of both Parietal and Visceral Mesothelium In Vivo
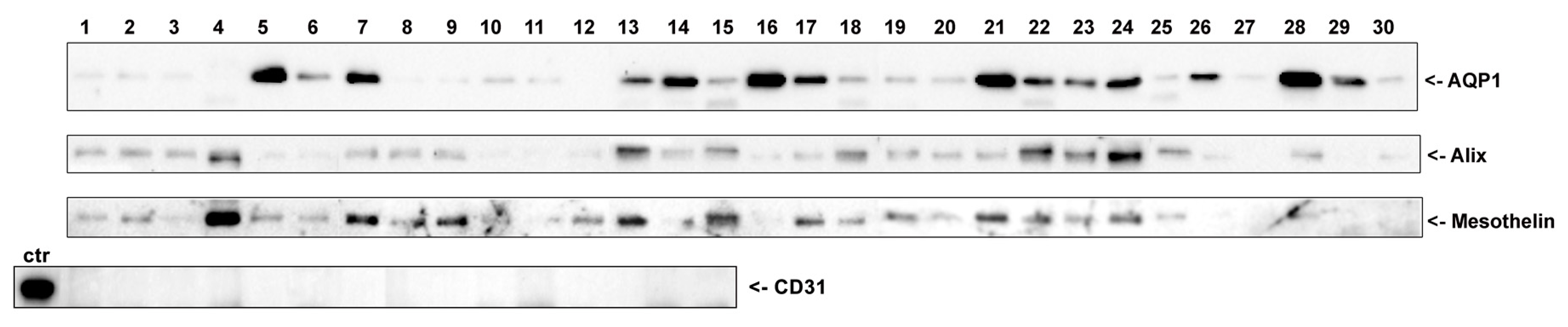
3.2. AQP1 Is Released by Mesothelial Cells in the PD Effluent through Exosomes (Qualitative Analysis)
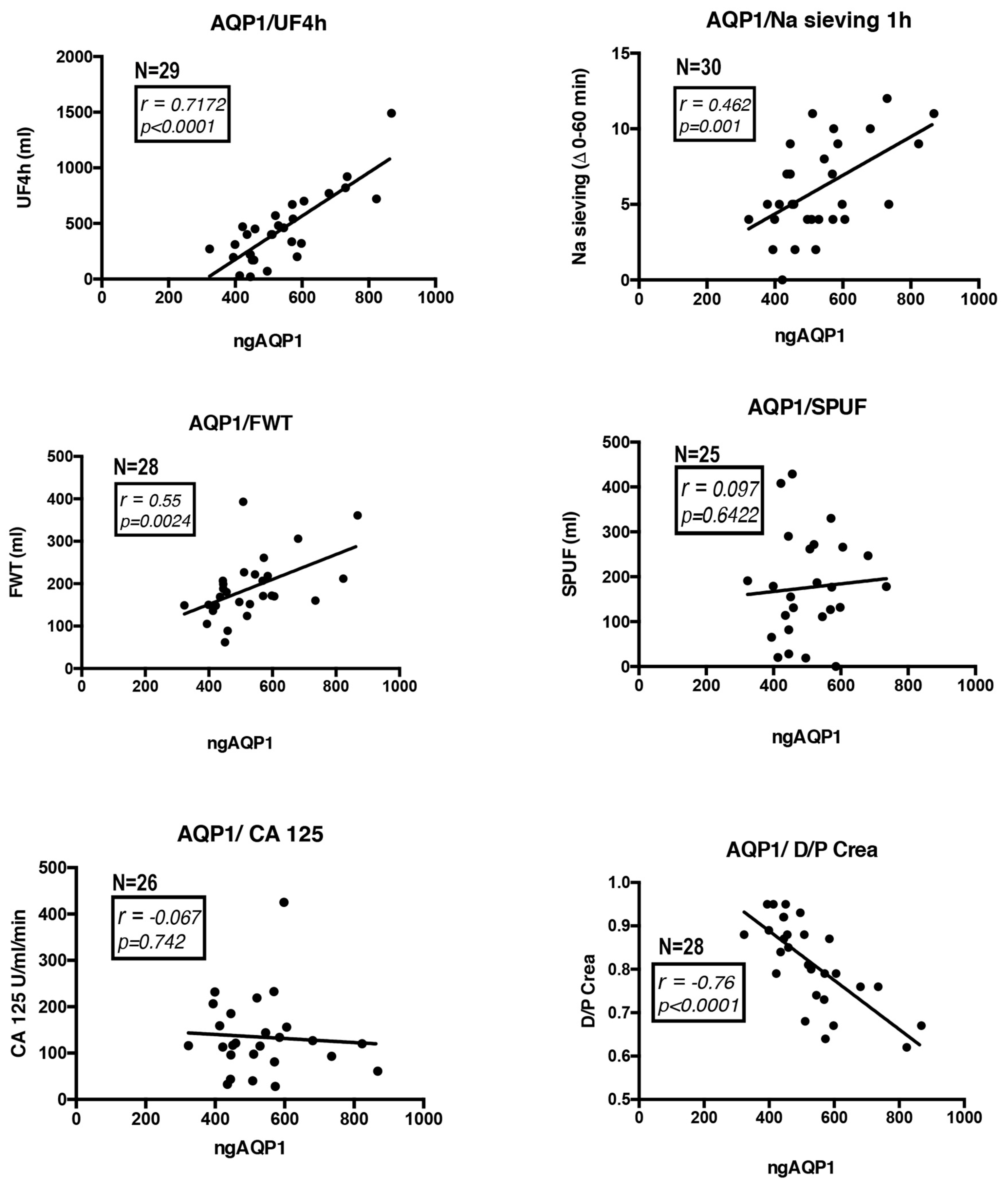
3.3. AQP1 Abundance in the PD Effluent Positively Correlates with the Efficiency of PD (Quantitative Analysis)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Abraham, G.; Varughese, S.; Mathew, M.; Vijayan, M. A review of acute and chronic peritoneal dialysis in developing countries. Clin. Kidney J. 2015, 8, 310–317. [Google Scholar] [CrossRef]
- Nakamoto, H.; Hamada, C.; Shimaoka, T.; Sekiguchi, Y.; Io, H.; Kaneko, K.; Horikoshi, S.; Tomino, Y. Accumulation of advanced glycation end products and beta 2-microglobulin in fibrotic thickening of the peritoneum in long-term peritoneal dialysis patients. J. Artif. Organs 2014, 17, 60–68. [Google Scholar] [CrossRef]
- Van Diepen, A.T.; Jassal, S.V. A qualitative systematic review of the literature supporting a causal relationship between exit-site infection and subsequent peritonitis in patients with end-stage renal disease treated with peritoneal dialysis. Perit. Dial. Int. 2013, 33, 604–610. [Google Scholar] [CrossRef]
- Twardowski, Z.J.N.; Nolph, K.D.; Prowant, B.; Ryan, L.; Moore, H.; Nielsen, M.P. Peritoneal equilibration test. Perit. Dial. Bull. 1987, 7, 10. [Google Scholar]
- La Milia, V.; Di Filippo, S.; Crepaldi, M.; Del Vecchio, L.; Dell’Oro, C.; Andrulli, S.; Locatelli, F. Mini-peritoneal equilibration test: A simple and fast method to assess free water and small solute transport across the peritoneal membrane. Kidney Int. 2005, 68, 840–846. [Google Scholar] [CrossRef] [Green Version]
- Cnossen, T.T.; Smit, W.; Konings, C.J.; Kooman, J.P.; Leunissen, K.M.; Krediet, R.T. Quantification of free water transport during the peritoneal equilibration test. Perit. Dial. Int. 2009, 29, 523–527. [Google Scholar]
- Rippe, B.; Stelin, G. Simulations of peritoneal solute transport during capd. Application of two-pore formalism. Kidney Int. 1989, 35, 1234–1244. [Google Scholar] [CrossRef]
- Stelin, G.; Rippe, B. A phenomenological interpretation of the variation in dialysate volume with dwell time in capd. Kidney Int. 1990, 38, 465–472. [Google Scholar] [CrossRef]
- Preston, G.M.; Carroll, T.P.; Guggino, W.B.; Agre, P. Appearance of water channels in xenopus oocytes expressing red cell chip28 protein. Science 1992, 256, 385–387. [Google Scholar] [CrossRef]
- Rippe, B. A three-pore model of peritoneal transport. Perit. Dial. Int. 1993, 13 (Suppl. 2), S35–S38. [Google Scholar]
- Carlsson, O.; Nielsen, S.; Zakaria el, R.; Rippe, B. In vivo inhibition of transcellular water channels (aquaporin-1) during acute peritoneal dialysis in rats. Am. J. Physiol. 1996, 271, H2254–H2262. [Google Scholar] [CrossRef]
- Devuyst, O.; Nielsen, S.; Cosyns, J.P.; Smith, B.L.; Agre, P.; Squifflet, J.P.; Pouthier, D.; Goffin, E. Aquaporin-1 and endothelial nitric oxide synthase expression in capillary endothelia of human peritoneum. Am. J. Physiol. 1998, 275, H234–H242. [Google Scholar] [CrossRef]
- Pannekeet, M.M.; Mulder, J.B.; Weening, J.J.; Struijk, D.G.; Zweers, M.M.; Krediet, R.T. Demonstration of aquaporin-chip in peritoneal tissue of uremic and capd patients. Perit. Dial. Int. 1996, 16 (Suppl. 1), S54–S57. [Google Scholar]
- Lai, K.N.; Li, F.K.; Lan, H.Y.; Tang, S.; Tsang, A.W.; Chan, D.T.; Leung, J.C. Expression of aquaporin-1 in human peritoneal mesothelial cells and its upregulation by glucose in vitro. J. Am. Soc. Nephrol. 2001, 12, 1036–1045. [Google Scholar]
- Zhai, Y.; Bloch, J.; Homme, M.; Schaefer, J.; Hackert, T.; Philippin, B.; Schwenger, V.; Schaefer, F.; Schmitt, C.P. Buffer-dependent regulation of aquaporin-1 expression and function in human peritoneal mesothelial cells. Pediatr. Nephrol. 2012, 27, 1165–1177. [Google Scholar] [CrossRef]
- Aufricht, C.; Beelen, R.; Eberl, M.; Fischbach, M.; Fraser, D.; Jorres, A.; Kratochwill, K.; LopezCabrera, M.; Rutherford, P.; Schmitt, C.P.; et al. Biomarker research to improve clinical outcomes of peritoneal dialysis: Consensus of the european training and research in peritoneal dialysis (eutripd) network. Kidney Int. 2017, 92, 824–835. [Google Scholar] [CrossRef]
- Lopes Barreto, D.; Krediet, R.T. Current status and practical use of effluent biomarkers in peritoneal dialysis patients. Am. J. Kidney Dis. 2013, 62, 823–833. [Google Scholar] [CrossRef]
- Bernardo, A.P.; Bajo, M.A.; Santos, O.; del Peso, G.; Carvalho, M.J.; Cabrita, A.; Selgas, R.; Rodrigues, A. Two-in-one protocol: Simultaneous small-pore and ultrasmall-pore peritoneal transport quantification. Perit. Dial. Int. 2012, 32, 537–544. [Google Scholar] [CrossRef]
- Chang, K.; Pastan, I. Molecular cloning of mesothelin, a differentiation antigen present on mesothelium, mesotheliomas, and ovarian cancers. Proc. Natl. Acad. Sci. USA 1996, 93, 136–140. [Google Scholar] [CrossRef]
- Verkman, A.S. Aquaporins at a glance. J. Cell Sci. 2011, 124, 2107–2112. [Google Scholar] [CrossRef] [Green Version]
- Pisitkun, T.; Shen, R.F.; Knepper, M.A. Identification and proteomic profiling of exosomes in human urine. Proc. Natl. Acad. Sci. USA 2004, 101, 13368–13373. [Google Scholar] [CrossRef] [Green Version]
- Henne, W.M.; Buchkovich, N.J.; Emr, S.D. The escrt pathway. Dev. Cell 2011, 21, 77–91. [Google Scholar] [CrossRef]
- Pusztaszeri, M.P.; Seelentag, W.; Bosman, F.T. Immunohistochemical expression of endothelial markers cd31, cd34, von willebrand factor, and fli-1 in normal human tissues. J. Histochem. Cytochem. 2006, 54, 385–395. [Google Scholar] [CrossRef]
- Rippe, B.; Stelin, G.; Haraldsson, B. Computer simulations of peritoneal fluid transport in capd. Kidney Int. 1991, 40, 315–325. [Google Scholar] [CrossRef]
- Rippe, B.; Haraldsson, B. Transport of macromolecules across microvascular walls: The two-pore theory. Physiol. Rev. 1994, 74, 163–219. [Google Scholar] [CrossRef]
- Kabanda, A.; Goffin, E.; Bernard, A.; Lauwerys, R.; van Ypersele de Strihou, C. Factors influencing serum levels and peritoneal clearances of low molecular weight proteins in continuous ambulatory peritoneal dialysis. Kidney Int. 1995, 48, 1946–1952. [Google Scholar] [CrossRef] [Green Version]
- King, L.S.; Yasui, M.; Agre, P. Aquaporins in health and disease. Mol. Med. Today 2000, 6, 60–65. [Google Scholar] [CrossRef]
- Kozono, D.; Yasui, M.; King, L.S.; Agre, P. Aquaporin water channels: Atomic structure molecular dynamics meet clinical medicine. J. Clin. Investig. 2002, 109, 1395–1399. [Google Scholar] [CrossRef]
- Yang, B.; Folkesson, H.G.; Yang, J.; Matthay, M.A.; Ma, T.; Verkman, A.S. Reduced osmotic water permeability of the peritoneal barrier in aquaporin-1 knockout mice. Am. J. Physiol. 1999, 276, C76–C81. [Google Scholar] [CrossRef]
- Zhang, W.; Freichel, M.; van der Hoeven, F.; Nawroth, P.P.; Katus, H.; Kalble, F.; Zitron, E.; Schwenger, V. Novel endothelial cell-specific aqp1 knockout mice confirm the crucial role of endothelial aqp1 in ultrafiltration during peritoneal dialysis. PLoS ONE 2016, 11, e0145513. [Google Scholar] [CrossRef]
- Fevrier, B.; Raposo, G. Exosomes: Endosomal-derived vesicles shipping extracellular messages. Curr. Opin. Cell Biol. 2004, 16, 415–421. [Google Scholar] [CrossRef]
- Baietti, M.F.; Zhang, Z.; Mortier, E.; Melchior, A.; Degeest, G.; Geeraerts, A.; Ivarsson, Y.; Depoortere, F.; Coomans, C.; Vermeiren, E.; et al. Syndecan-syntenin-alix regulates the biogenesis of exosomes. Nat. Cell Biol. 2012, 14, 677–685. [Google Scholar] [CrossRef]
- Hassan, R.; Ho, M. Mesothelin targeted cancer immunotherapy. Eur. J. Cancer 2008, 44, 46–53. [Google Scholar] [CrossRef]
- Thery, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef]
- Murer, L.; Addabbo, F.; Carmosino, M.; Procino, G.; Tamma, G.; Montini, G.; Rigamonti, W.; Zucchetta, P.; Della Vella, M.; Venturini, A.; et al. Selective decrease in urinary aquaporin 2 and increase in prostaglandin e2 excretion is associated with postobstructive polyuria in human congenital hydronephrosis. J. Am. Soc. Nephrol. 2004, 15, 2705–2712. [Google Scholar] [CrossRef]
- Procino, G.; Portincasa, P.; Mastrofrancesco, L.; Castorani, L.; Bonfrate, L.; Addabbo, F.; Carmosino, M.; Di Ciaula, A.; Svelto, M. Simvastatin increases aqp2 urinary excretion in hypercholesterolemic patients: A pleiotropic effect of interest for patients with impaired aqp2 trafficking. Clin. Pharmacol. Ther. 2016, 99, 528–537. [Google Scholar] [CrossRef]
- Rossi, L.; Nicoletti, M.C.; Carmosino, M.; Mastrofrancesco, L.; Di Franco, A.; Indrio, F.; Lella, R.; Laviola, L.; Giorgino, F.; Svelto, M.; et al. Urinary excretion of kidney aquaporins as possible diagnostic biomarker of diabetic nephropathy. J. Diabetes Res. 2017, 2017, 4360357. [Google Scholar] [CrossRef]
- Valenti, G.; Laera, A.; Gouraud, S.; Pace, G.; Aceto, G.; Penza, R.; Selvaggi, F.P.; Svelto, M. Low-calcium diet in hypercalciuric enuretic children restores aqp2 excretion and improves clinical symptoms. Am. J. Physiol. Renal Physiol. 2002, 283, F895–F903. [Google Scholar] [CrossRef]
- Valenti, G.; Laera, A.; Pace, G.; Aceto, G.; Lospalluti, M.L.; Penza, R.; Selvaggi, F.P.; Chiozza, M.L.; Svelto, M. Urinary aquaporin 2 and calciuria correlate with the severity of enuresis in children. J. Am. Soc. Nephrol. 2000, 11, 1873–1881. [Google Scholar]
- Ni, J.; Verbavatz, J.M.; Rippe, A.; Boisde, I.; Moulin, P.; Rippe, B.; Verkman, A.S.; Devuyst, O. Aquaporin-1 plays an essential role in water permeability and ultrafiltration during peritoneal dialysis. Kidney Int. 2006, 69, 1518–1525. [Google Scholar] [CrossRef] [Green Version]
- Krediet, R.T. Dialysate cancer antigen 125 concentration as marker of peritoneal membrane status in patients treated with chronic peritoneal dialysis. Perit. Dial. Int. 2001, 21, 560–567. [Google Scholar]
- Devuyst, O.; Rippe, B. Water transport across the peritoneal membrane. Kidney Int. 2014, 85, 750–758. [Google Scholar] [CrossRef] [Green Version]
- Mutsaers, S.E. Mesothelial cells: Their structure, function and role in serosal repair. Respirology 2002, 7, 171–191. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Yorioka, N.; Kyuden, Y.; Asakimori, Y.; Kiribayashi, K.; Ogawa, T.; Kohno, N. Effect of glucose polymer on the intercellular junctions of cultured human peritoneal mesothelial cells. Nephron Clin. Pract. 2003, 93, c97–c105. [Google Scholar] [CrossRef]
- Simionescu, M.; Simionescu, N. Organization of cell junctions in the peritoneal mesothelium. J. Cell Biol. 1977, 74, 98–110. [Google Scholar] [CrossRef] [Green Version]
- Guo, P.; Weinstein, A.M.; Weinbaum, S. A dual-pathway ultrastructural model for the tight junction of rat proximal tubule epithelium. Am. J. Physiol. Renal Physiol. 2003, 285, F241–F257. [Google Scholar] [CrossRef] [Green Version]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corciulo, S.; Nicoletti, M.C.; Mastrofrancesco, L.; Milano, S.; Mastrodonato, M.; Carmosino, M.; Gerbino, A.; Corciulo, R.; Russo, R.; Svelto, M.; et al. AQP1-Containing Exosomes in Peritoneal Dialysis Effluent As Biomarker of Dialysis Efficiency. Cells 2019, 8, 330. https://doi.org/10.3390/cells8040330
Corciulo S, Nicoletti MC, Mastrofrancesco L, Milano S, Mastrodonato M, Carmosino M, Gerbino A, Corciulo R, Russo R, Svelto M, et al. AQP1-Containing Exosomes in Peritoneal Dialysis Effluent As Biomarker of Dialysis Efficiency. Cells. 2019; 8(4):330. https://doi.org/10.3390/cells8040330
Chicago/Turabian StyleCorciulo, Simone, Maria Celeste Nicoletti, Lisa Mastrofrancesco, Serena Milano, Maria Mastrodonato, Monica Carmosino, Andrea Gerbino, Roberto Corciulo, Roberto Russo, Maria Svelto, and et al. 2019. "AQP1-Containing Exosomes in Peritoneal Dialysis Effluent As Biomarker of Dialysis Efficiency" Cells 8, no. 4: 330. https://doi.org/10.3390/cells8040330
APA StyleCorciulo, S., Nicoletti, M. C., Mastrofrancesco, L., Milano, S., Mastrodonato, M., Carmosino, M., Gerbino, A., Corciulo, R., Russo, R., Svelto, M., Gesualdo, L., & Procino, G. (2019). AQP1-Containing Exosomes in Peritoneal Dialysis Effluent As Biomarker of Dialysis Efficiency. Cells, 8(4), 330. https://doi.org/10.3390/cells8040330