Differential Expression and Localization of Branchial AQP1 and AQP3 in Japanese Medaka (Oryzias latipes)

 , and
, and 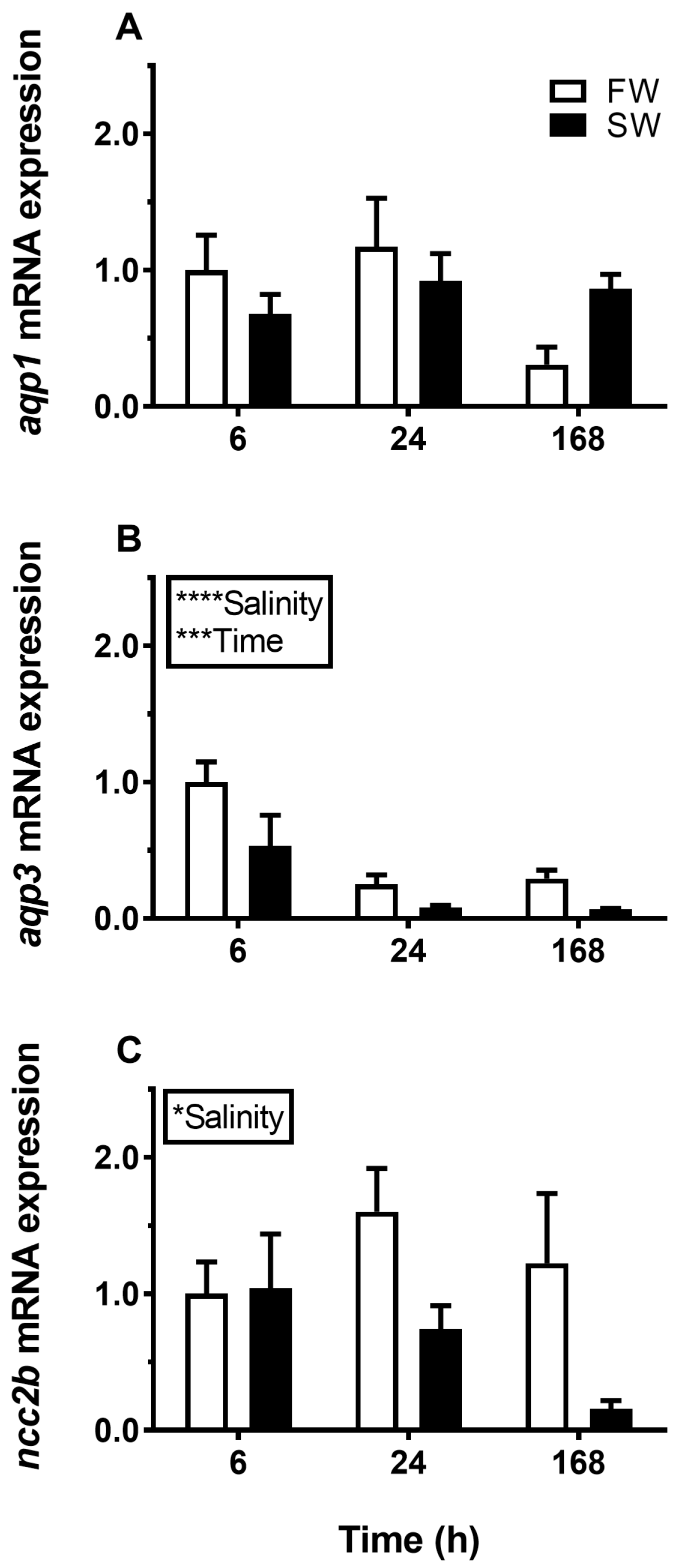{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish and Maintenance
2.2. Salinity Transfer Experiments: FW–SW, FW–Ion-Poor Water (IPW).
2.3. Ex vivo Hormone Incubation Experiments
2.4. RNA Isolation, cDNA Synthesis, and Real-Time qPCR
2.5. Western Blot Analysis
2.6. Immunohistochemistry
2.7. Statistical Analysis
3. Results
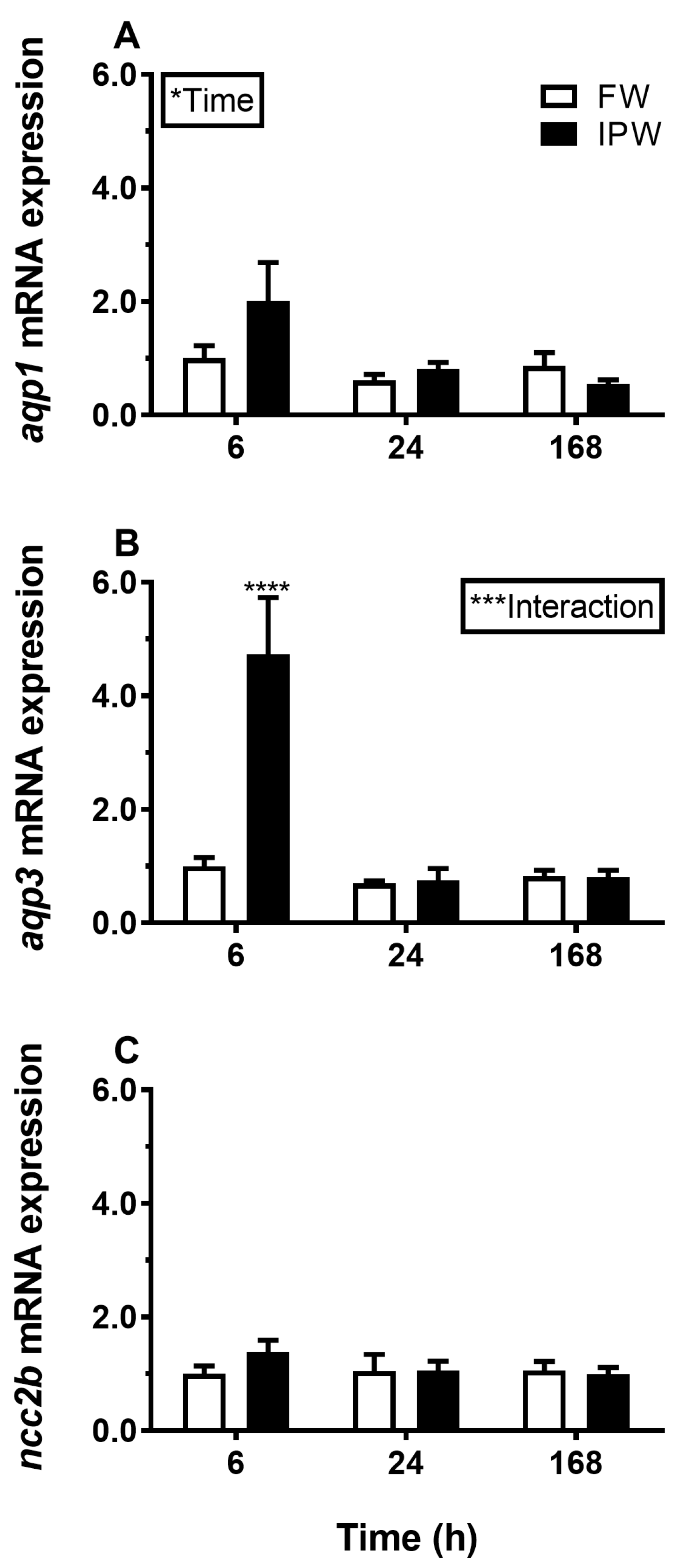
3.1. Aquaporin mRNA Response to Osmotic Challenges
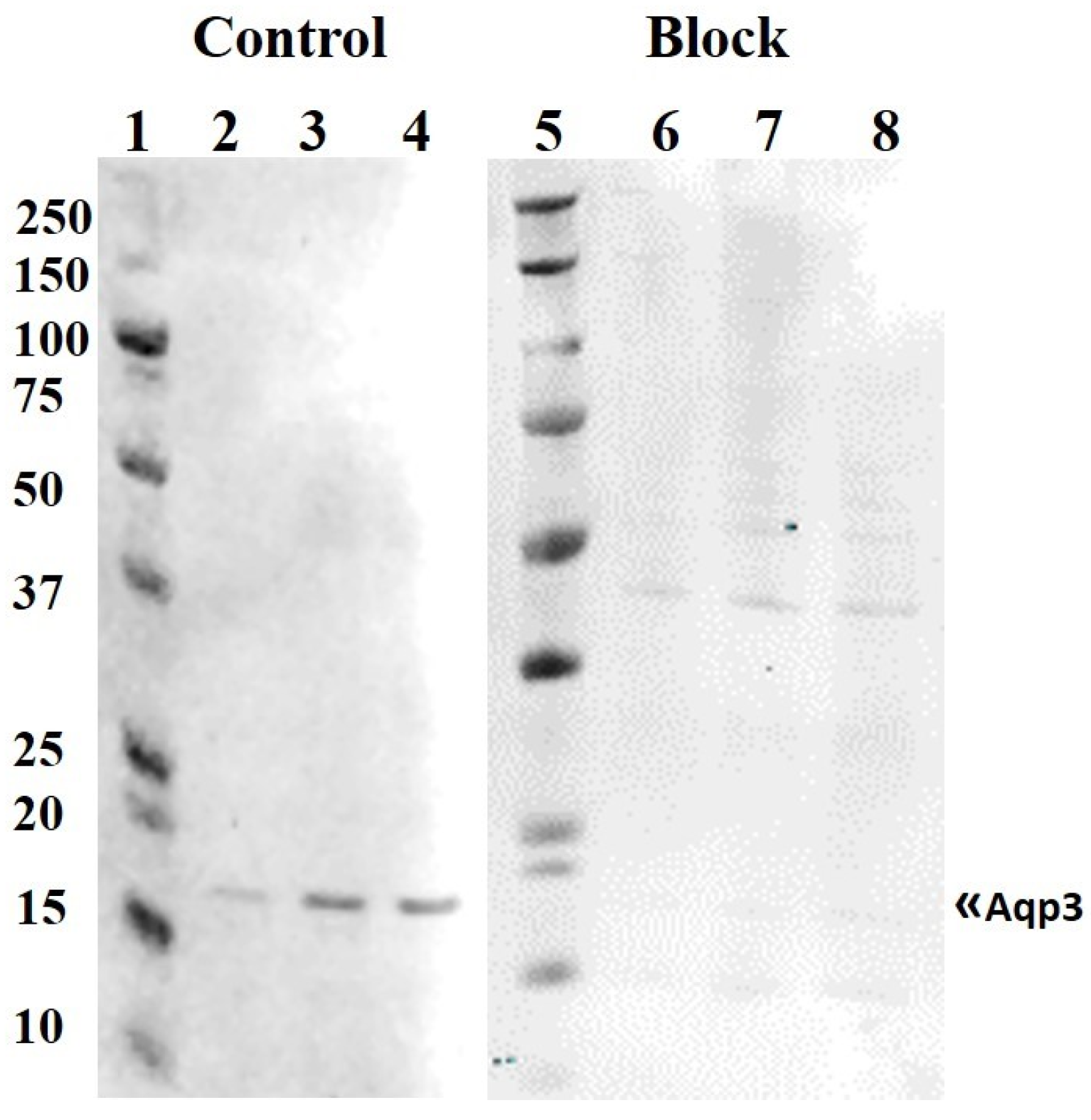
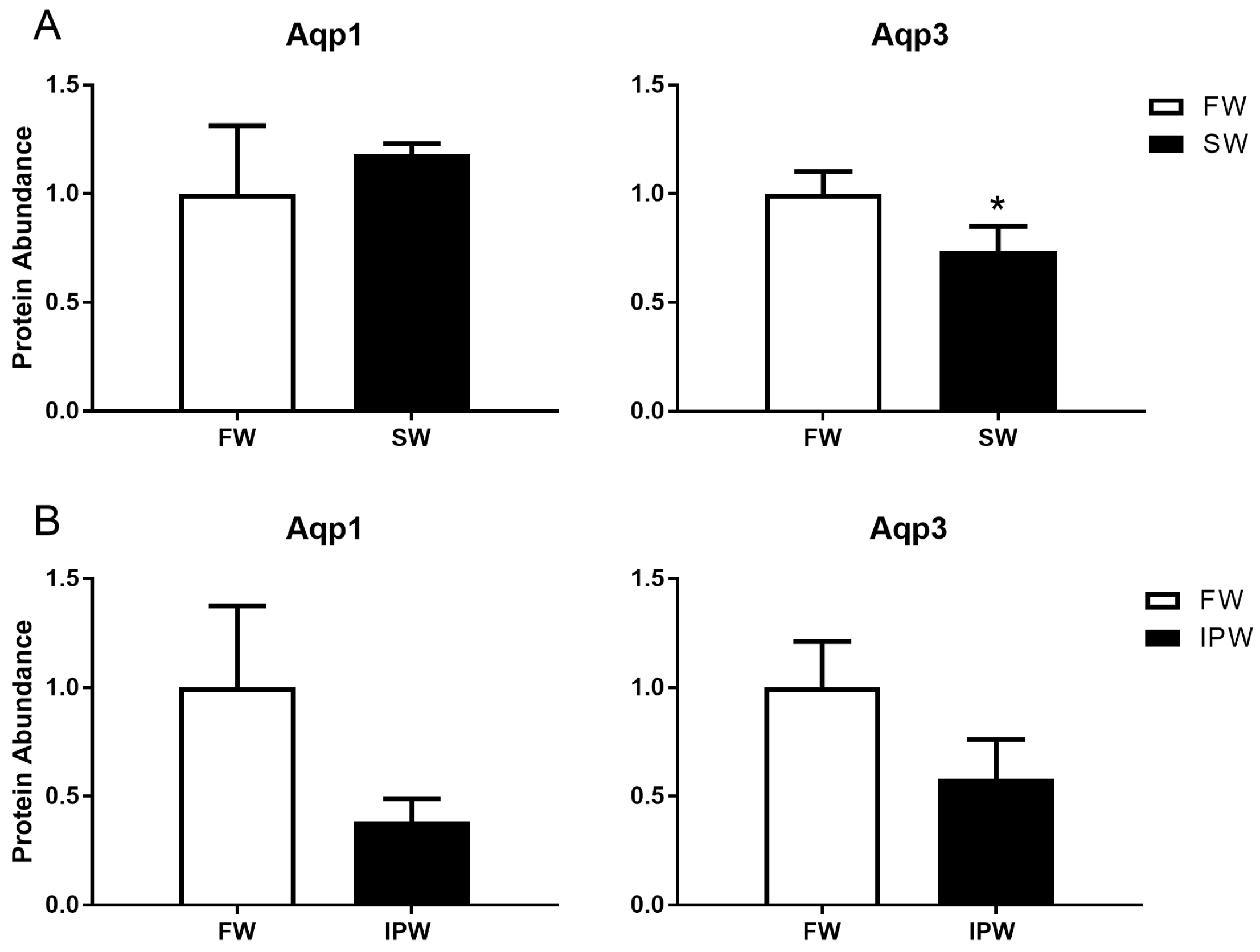
3.2. Western Blotting and Aquaporin Protein Levels
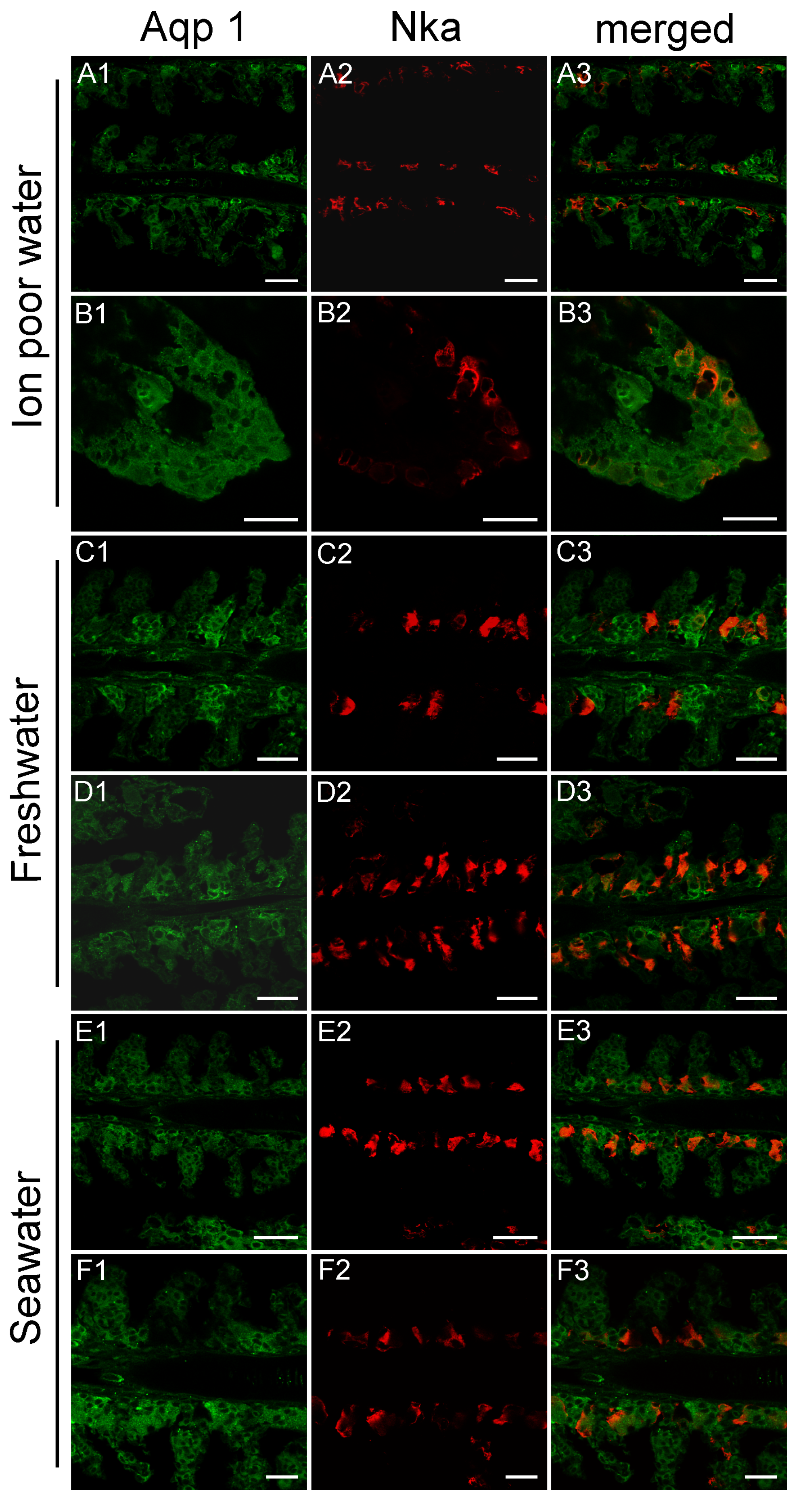
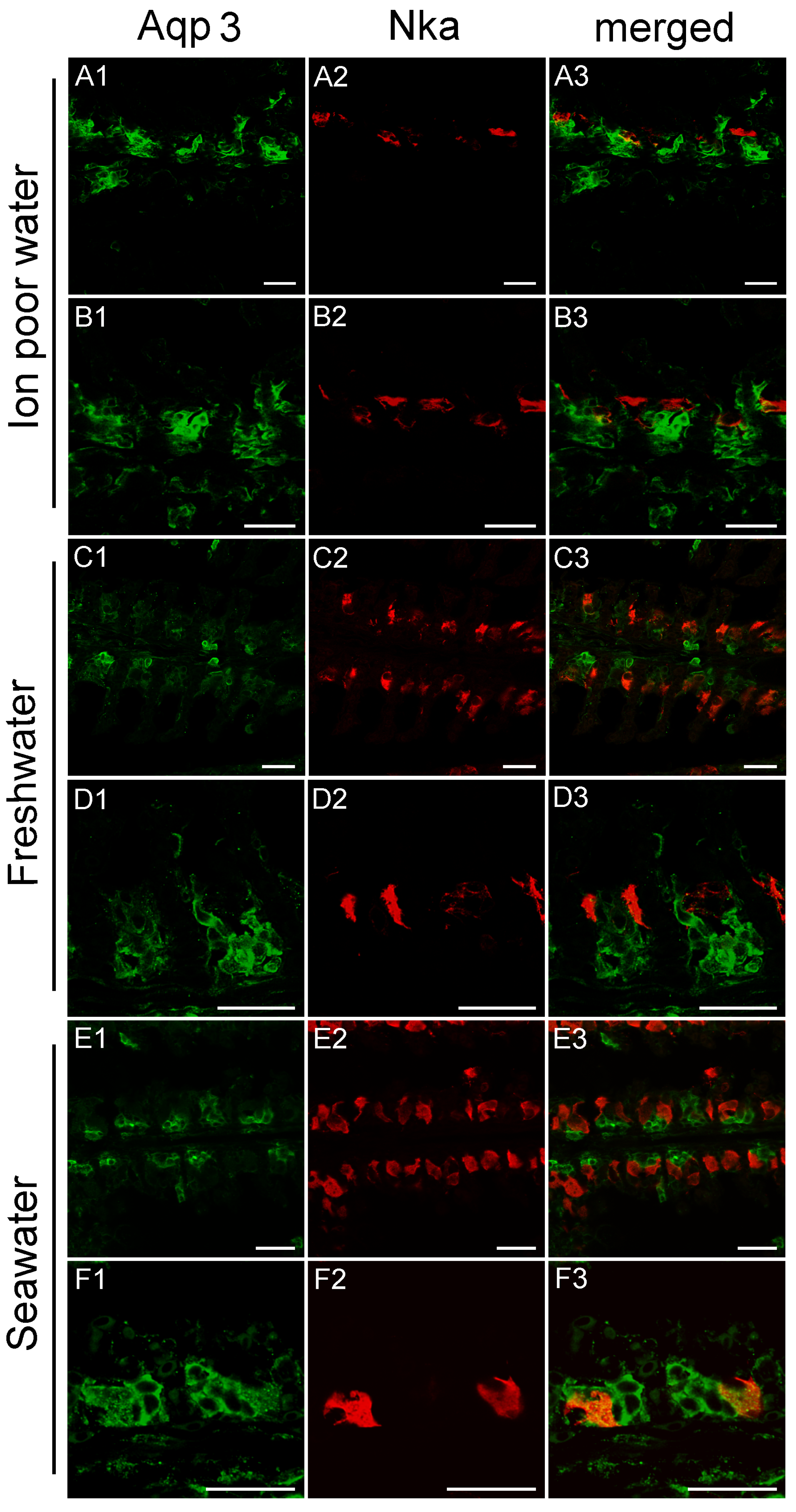
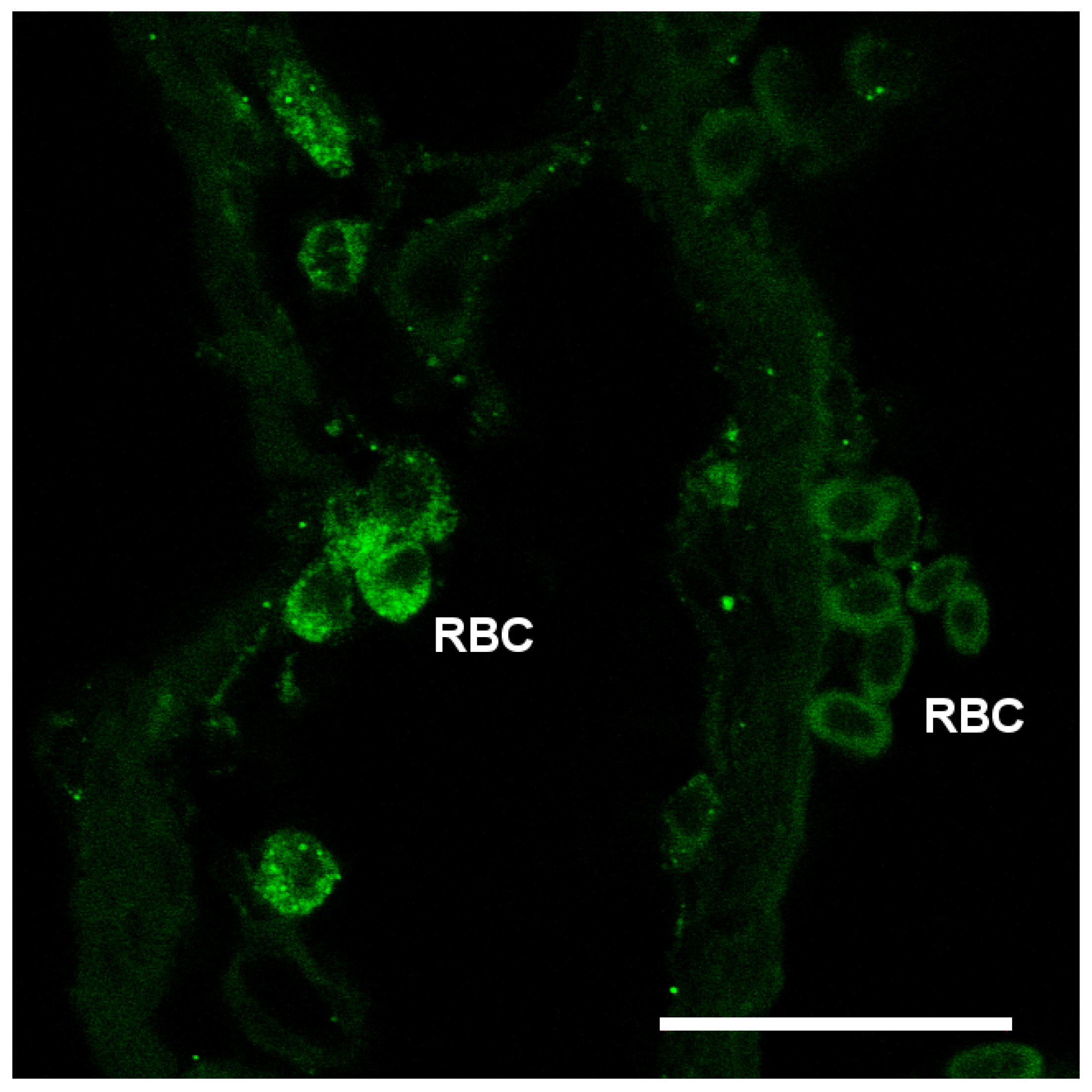
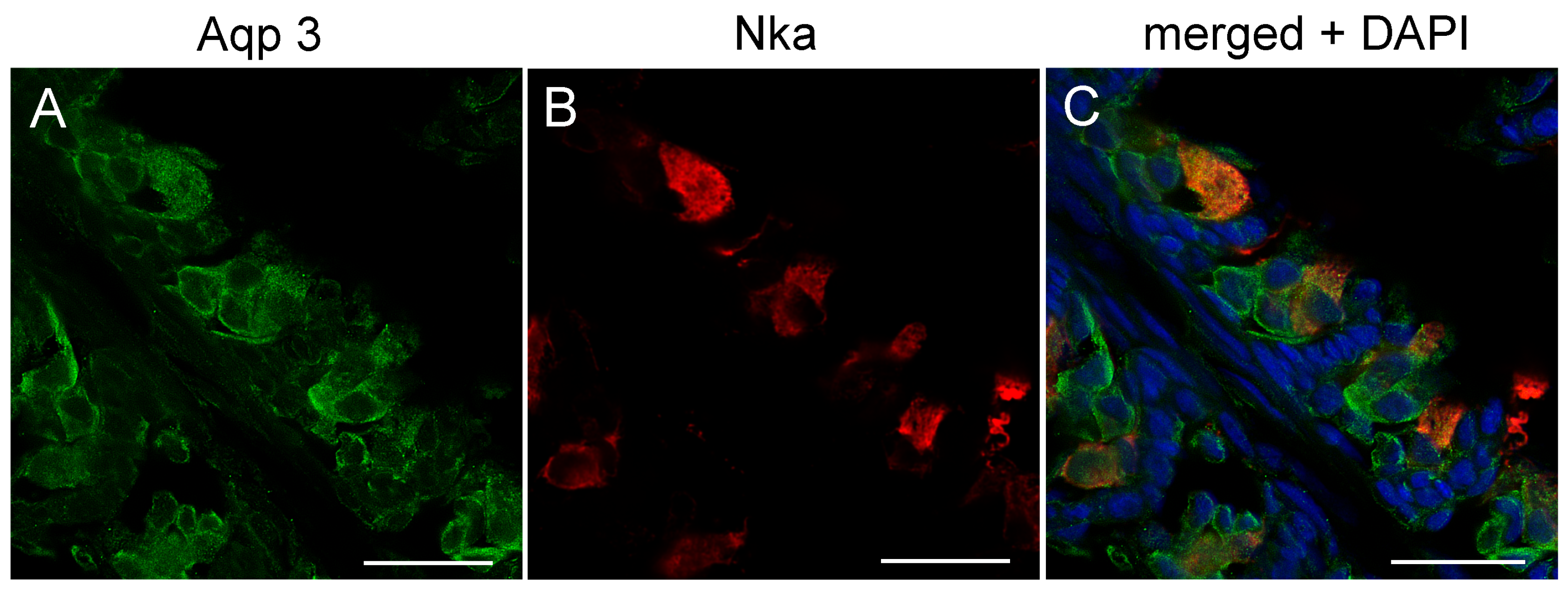
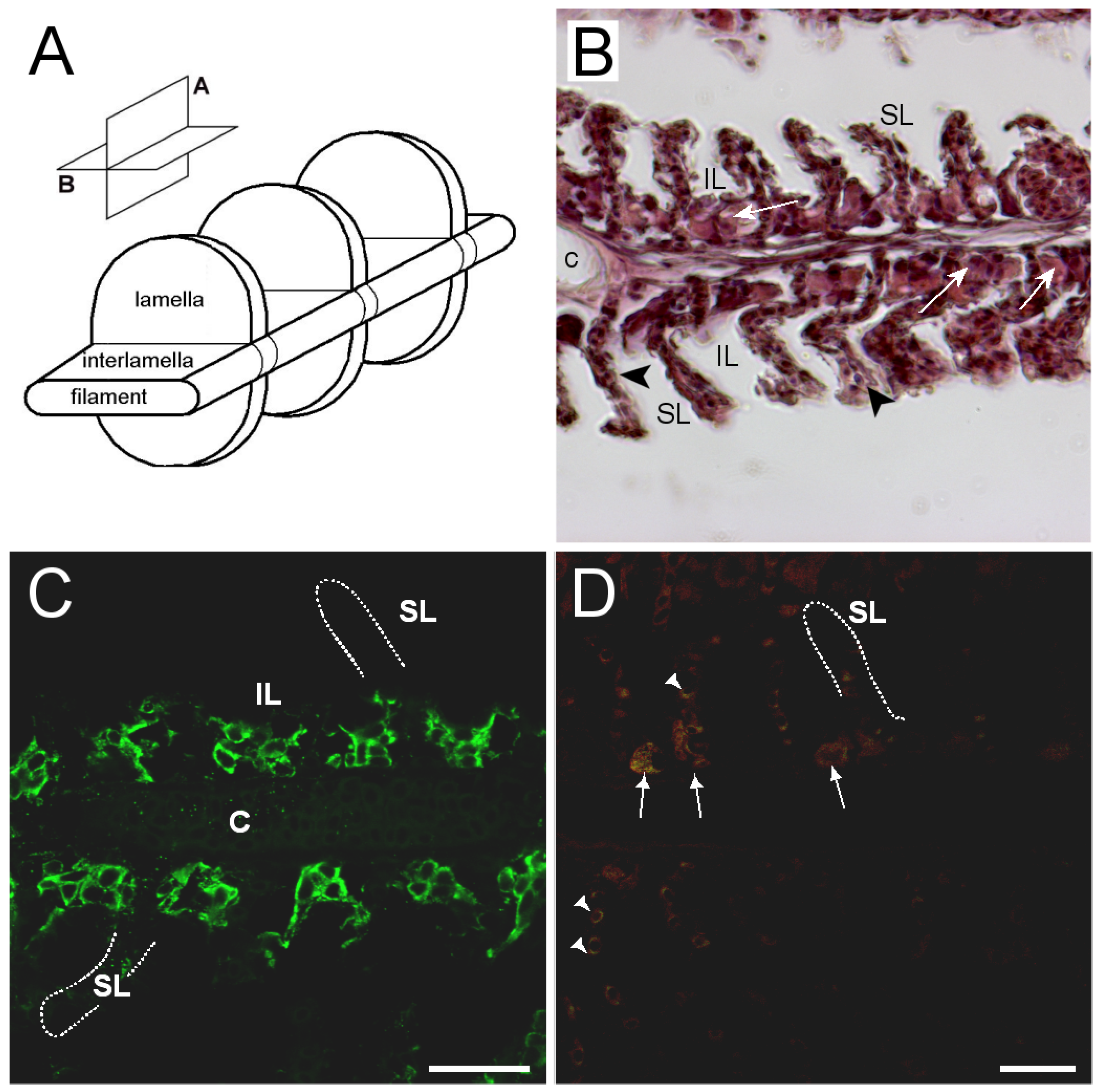
3.3. Branchial Aquaporin Localization
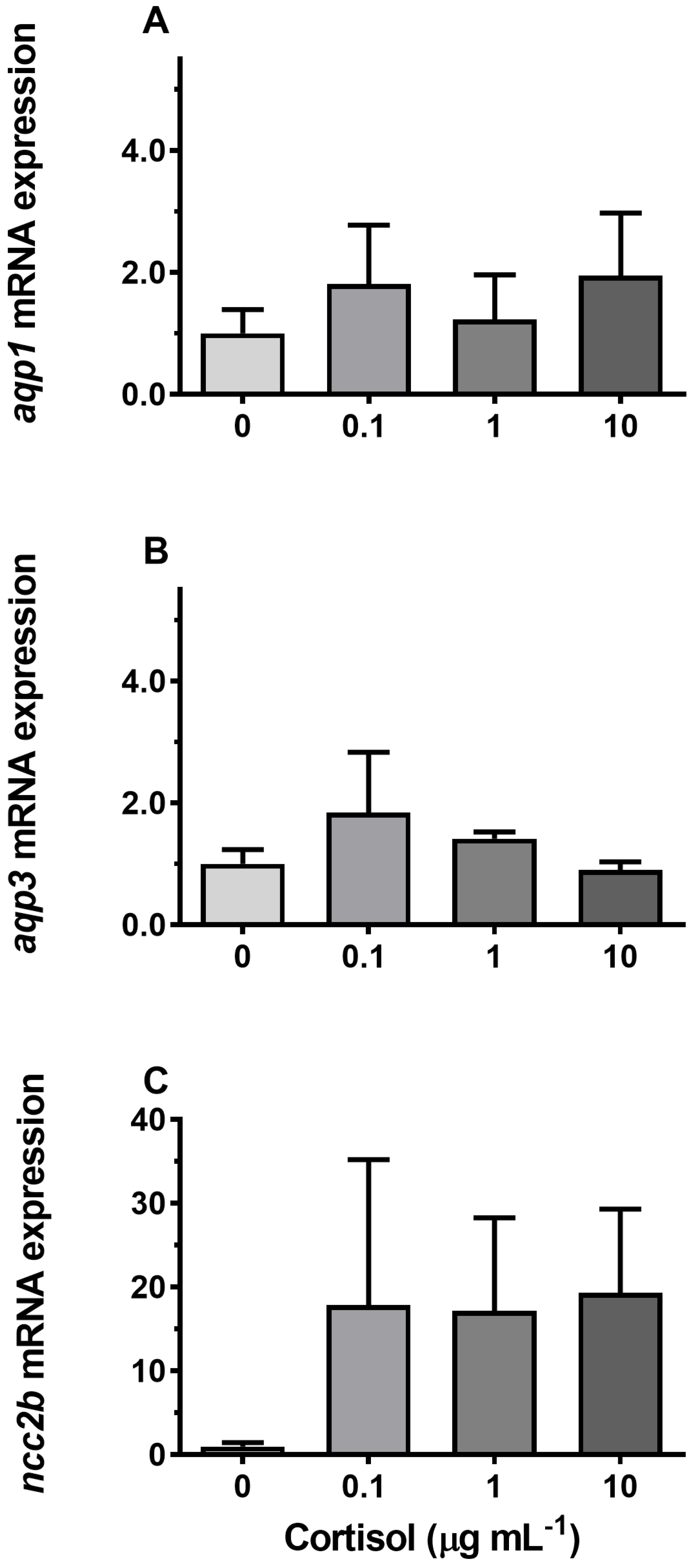
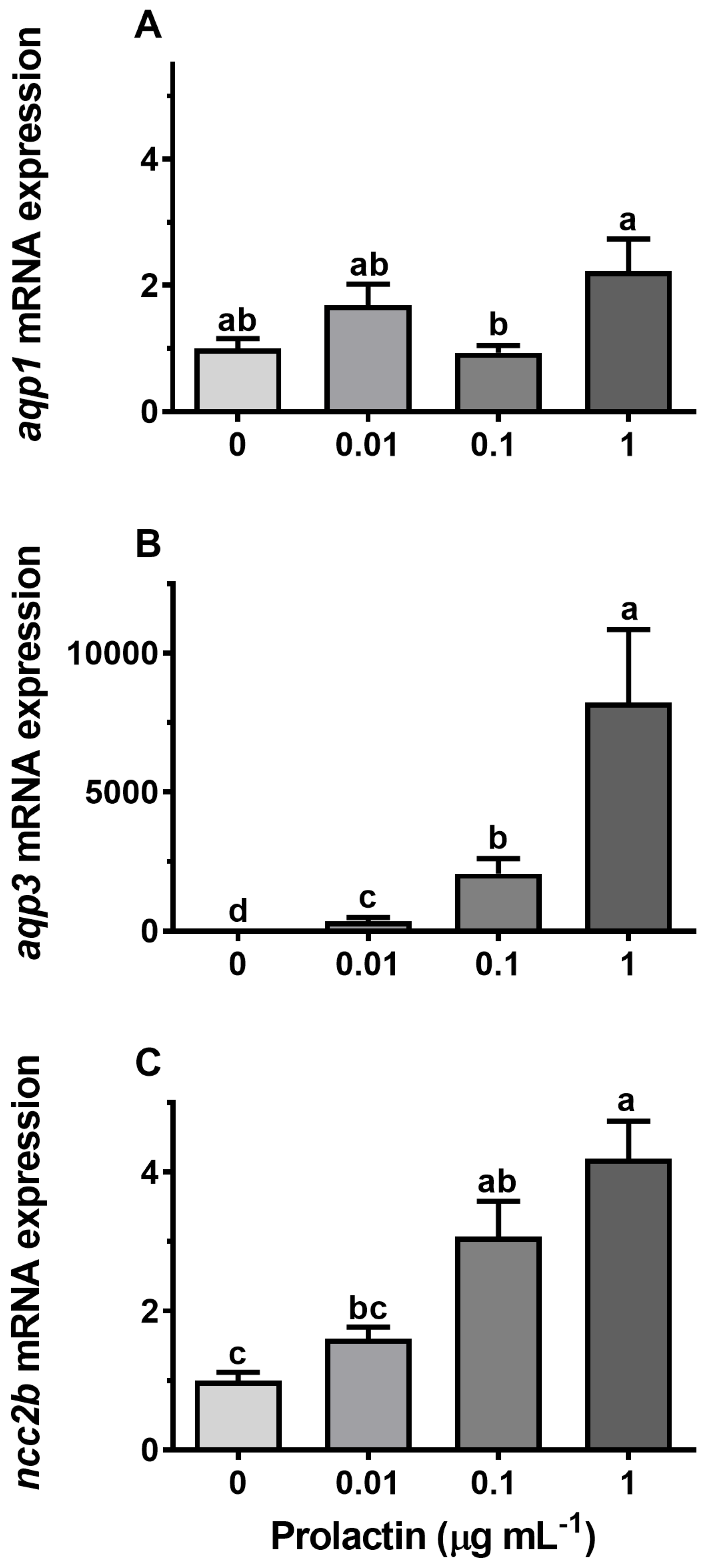
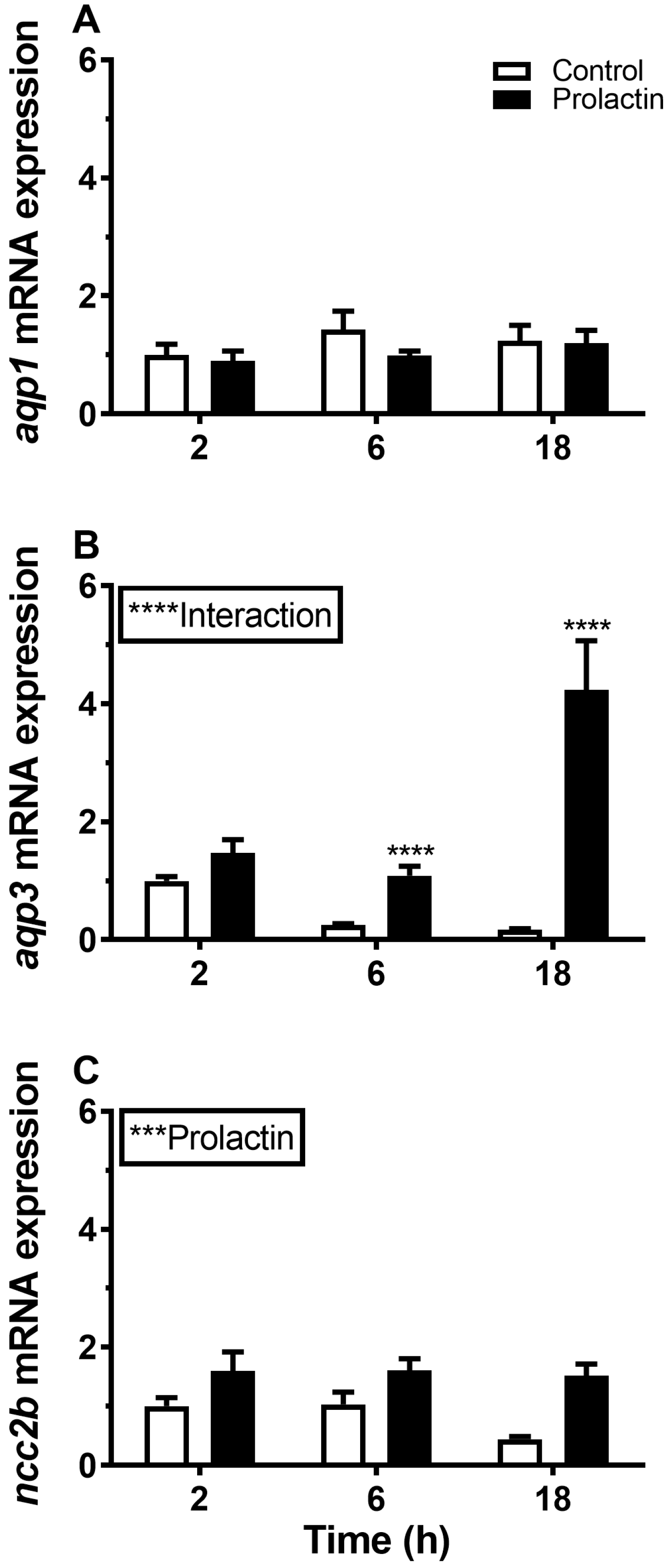
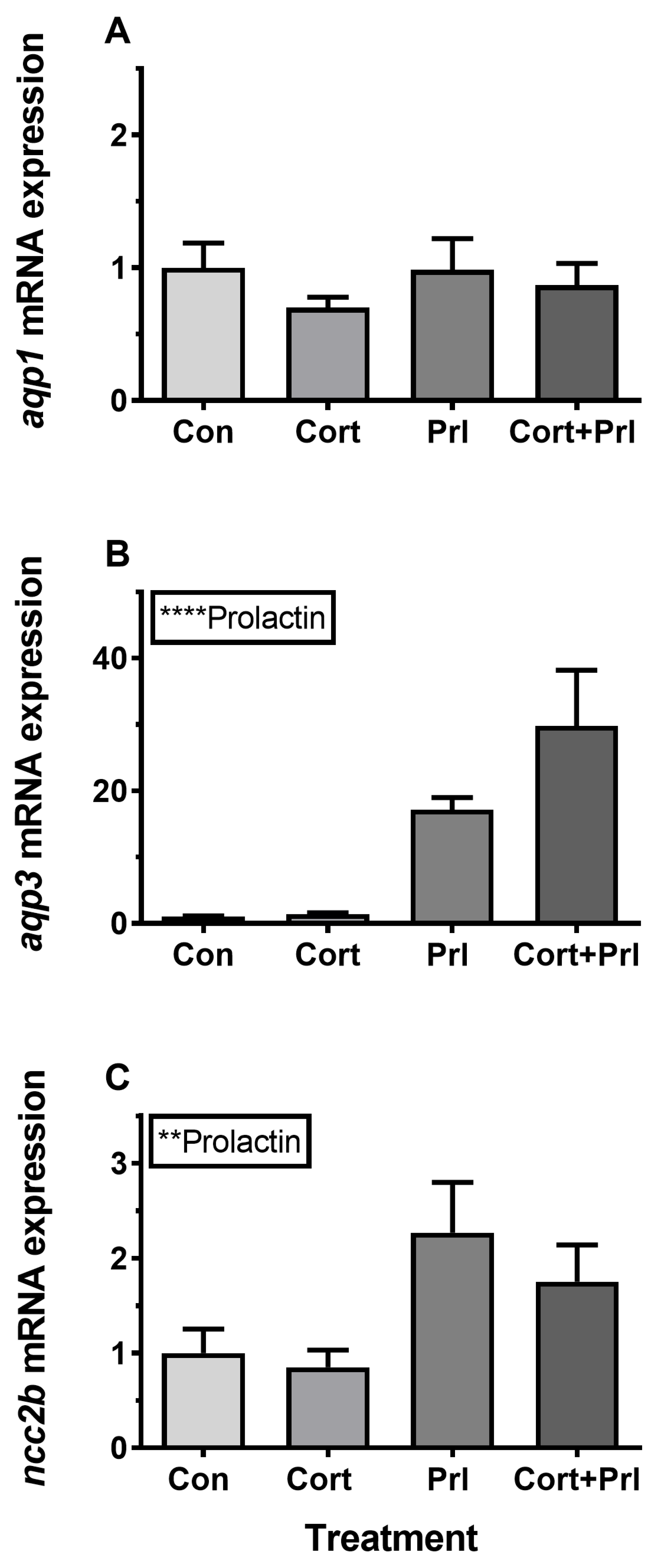
3.4. Hormonal Effects on AQP Expression Ex Vivo
4. Discussion
4.1. Salinity Response of Branchial Aquaporins
4.2. Immunolocalization and Physiological Function of Branchial Aquaporins
4.3. Endocrine Regulation of Branchial Aquaporins
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Edwards, S.L.; Marshall, W.S. Principles and patterns of osmoregulation and euryhalinity in fishes. In Fish Physiology; McCormick, S., Farrel, A., Brauner, C., Eds.; Elsevier: Cambrdige, MA, USA, 2013; Volume 32, pp. 1–44. ISBN 9780123969514. [Google Scholar]
- Evans, D.H.; Piermarini, P.M.; Choe, K.P. The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Phys. Rev. 2005, 85, 97–177. [Google Scholar] [CrossRef]
- Hsu, H.H.; Lin, L.Y.; Tseng, Y.C.; Horng, J.L.; Hwang, P.P. A new model for fish ion regulation: Identification of ionocytes in freshwater-and seawater-acclimated medaka (Oryzias latipes). Cell Tissue Res. 2014, 357, 225–243. [Google Scholar] [CrossRef]
- Marshall, W.S.; Lynch, E.A.; Cozzi, R.R.F. Redistribution of immunofluorescence of CFTR anion channel and NKCC cotransporter in chloride cells during adaptation of the killifish Fundulus heteroclitus to sea water. J. Exp. Biol. 2002, 205, 1265–1273. [Google Scholar]
- Marshall, W.S.; Grosell, M. Ion transport, osmoregulation, and acid-base regulation. In The Physiology of Fishes; Evans, D.H., Clairborne, J.B., Eds.; Taylor and Francis Group: Boca Raton, FL, USA, 2006; pp. 177–210. ISBN 978-0849380426. [Google Scholar]
- Wilson, J.M.; Laurent, P. Fish gill morphology: Inside out. J. Exp. Zool. 2002, 293, 192–213. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, S. Control of gill blood flow. In Fish Physiology: Recent Advances; Nilsson, S., Ed.; Springer: New York, NY, USA, 1986; pp. 86–101. ISBN 978-94-011-6560-0. [Google Scholar]
- Motais, R.; Isaia, J.; Rankin, J.; Maetz, J. Adaptive changes of the water permeability of the teleostean gill epithelium in relation to external salinity. J. Exp. Biol. 1969, 51, 529–546. [Google Scholar]
- Isaia, J. Water and nonelectrolyte permeation. In Fish Physiology; Hoar, W.S., Randall, D.J., Eds.; Academic Press: London, UK, 1984; Volume 10, pp. 1–38. ISBN 1546-5098. [Google Scholar]
- Hoffmann, E.K.; Lambert, I.H.; Pedersen, S.F. Physiology of cell volume regulation in vertebrates. Phys. Rev. 2009, 89, 193–277. [Google Scholar] [CrossRef] [PubMed]
- Agre, P.E.; Preston, G.M.; Smith, B.L.; Jung, J.S.; Raina, S.U.; Moon, C.H.; Guggino, W.B.; Nielsen, S.B. Aquaporin CHIP: The archetypal molecular water channel. Am. J. Phys.-Renal 1993, 265, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Ecelbarger, C.A.; Terris, J.A.; Frindt, G.U.; Echevarria, M.I.; Marples, D.; Nielsen, S.; Knepper, M.A. Aquaporin-3 water channel localization and regulation in rat kidney. Am. J. Phys.-Renal 1995, 269, 663–672. [Google Scholar] [CrossRef]
- Ishibashi, K.; Hara, S.; Kondo, S. Aquaporin water channels in mammals. Clin. Exp. Nephrol. 2009, 13, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Tingaud-Sequeira, A.; Chauvigné, F.; Fabra, M.; Lozano, J.; Raldúa, D.; Cerdà, J.A. Structural and functional divergence of two fish aquaporin-1 water channels following teleost-specific gene duplication. BMC Evol. Biol. 2008, 8, 259. [Google Scholar] [CrossRef]
- Sugiyama, Y.; Ota, Y.; Hara, M.; Inoue, S. Osmotic stress up-regulates aquaporin-3 gene expression in cultured human keratinocytes. BBA-Gene Struct. Expr. 2001, 1522, 82–88. [Google Scholar] [CrossRef]
- Boury-Jamot, M.; Sougrat, R.; Tailhardat, M.; Le Varlet, B.; Bonte, F.; Dumas, M.; Verbavatz, J.M. Expression and function of aquaporins in human skin: Is aquaporin-3 just a glycerol transporter? BBA-Biomembranes 2006, 1758, 1034–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutler, C.P.; Martinez, A.S.; Cramb, G. The role of aquaporin 3 in teleost fish. Comp. Biochem. Phys. A 2007, 148, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Nam, Y.K.; Kim, Y.K. Characterization and expression profiles of aquaporins (AQPs) 1a and 3a in mud loach Misgurnus mizolepis after experimental challenges. Fish. Aquat. Sci. 2017, 20, 23. [Google Scholar] [CrossRef]
- Finn, R.N.; Chauvigné, F.; Hlidberg, J.B.; Cutler, C.P.; Cerdà, J. The lineage-specific evolution of aquaporin gene clusters facilitated tetrapod terrestrial adaptation. PLoS ONE 2014, 9, e113686. [Google Scholar] [CrossRef]
- Madsen, S.S.; Engelund, M.B.; Cutler, C.P. Water transport and functional dynamics of aquaporins in osmoregulatory organs of fishes. Biol. Bull. 2015, 229, 70–92. [Google Scholar] [CrossRef]
- McCormick, S.D. Endocrine control of osmoregulation in teleost fish. Am. Zool. 2001, 41, 781–794. [Google Scholar] [CrossRef]
- Breves, J.P.; McCormick, S.D.; Karlstrom, R.O. Prolactin and teleost ionocytes: New insights into cellular and molecular targets of prolactin in vertebrate epithelia. Gen. Comp. Endocrinol. 2014, 203, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Flik, G.; Rentier-Delrue, F.; Wendelaar Bonga, S. Calcitropic effects of recombinant prolactins in Oreochromis mossambicus. Am. J. Phys.-Reg. I 1994, 266, 1302–1308. [Google Scholar] [CrossRef]
- Takei, Y.; McCormick, S.D. Euryhaline fishes: Hormonal control of fish euryhalinity. In Fish Physiology; McCormick, S., Farrel, A., Brauner, C., Eds.; Elsevier: Cambrdige, MA, USA, 2013; Volume 32, pp. 69–123. ISBN 9780123969514. [Google Scholar]
- Manzon, L.A. The role of prolactin in fish osmoregulation: A review. Gen. Comp. Endocrinol. 2002, 125, 291–310. [Google Scholar] [CrossRef]
- Tipsmark, C.K.; Madsen, S.S. Distinct hormonal regulation of Na+,K+-Atpase genes in the gill of Atlantic salmon (Salmo salar L.). J. Endocrinol. 2009, 203, 301–310. [Google Scholar] [CrossRef]
- Breves, J.; Inokuchi, M.; Yamaguchi, Y.; Seale, A.P.; Hunt, B.; Watanabe, S.; Lerner, D.; Kaneko, T.; Grau, G. Hormonal regulation of aquaporin 3: Opposing actions of prolactin and cortisol in tilapia gill. J. Endocrinol. 2016. [Google Scholar] [CrossRef]
- Breves, J.P.; Serizier, S.B.; Goffin, V.; McCormick, S.D.; Karlstrom, R.O. Prolactin regulates transcription of the ion uptake Na+/Cl− cotransporter (ncc) gene in zebrafish gill. Mol. Cell. Endocrinol. 2013, 369, 98–106. [Google Scholar] [CrossRef]
- Bossus, M.C.; Bollinger, R.J.; Reed, P.J.; Tipsmark, C.K. Prolactin and cortisol regulate branchial claudin expression in Japanese medaka. Gen. Comp. Endocrinol. 2017, 240, 77–83. [Google Scholar] [CrossRef]
- Tipsmark, C.K.; Luckenbach, J.A.; Madsen, S.S.; Borski, R.J. IGF-I and branchial IGF receptor expression and localization during salinity acclimation in striped bass. Am. J. Phys.-Reg. I 2007, 292, 535–543. [Google Scholar] [CrossRef]
- Breves, J.P.; Hirano, T.; Grau, E.G. Ionoregulatory and endocrine responses to disturbed salt and water balance in Mozambique tilapia exposed to confinement and handling stress. Comp. Biochem. Phys. A 2010, 155, 294–300. [Google Scholar] [CrossRef]
- Sakamoto, T.; Kozaka, T.; Takahashi, A.; Kawauchi, H.; Ando, M. Medaka (Oryzias latipes) as a model for hypoosmoregulation of euryhaline fishes. Aquaculture 2001, 193, 347–354. [Google Scholar] [CrossRef]
- Madsen, S.S.; Bujak, J.; Tipsmark, C.K. Aquaporin expression in the Japanese medaka (Oryzias latipes) in freshwater and seawater: Challenging the paradigm of intestinal water transport? J. Exp. Biol. 2014, 217, 3108–3121. [Google Scholar] [CrossRef]
- Bossus, M.C.; Madsen, S.S.; Tipsmark, C.K. Functional dynamics of claudin expression in Japanese medaka (Oryzias latipes): Response to environmental salinity. Comp. Biochem. Phys. A 2015, 187, 74–85. [Google Scholar] [CrossRef]
- Bollinger, R.J.; Madsen, S.S.; Bossus, M.C.; Tipsmark, C.K. Does Japanese medaka (Oryzias latipes) exhibit a gill Na+/K+-ATPase isoform switch during salinity change? J. Comp. Phys. B 2016, 186, 485–501. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29. [Google Scholar] [CrossRef]
- Bollinger, R.J.; Ellis, L.V.; Bossus, M.C.; Tipsmark, C.K. Prolactin controls Na+, Cl− cotransporter via Stat5 pathway in the teleost gill. Mol. Cell. Endocrinol. 2018, 477, 163–171. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Hiroi, J.; Yasumasu, S.; McCormick, S.D.; Hwang, P.P.; Kaneko, T. Evidence for an apical Na-Cl cotransporter involved in ion uptake in a teleost fish. J. Exp. Biol. 2008, 211, 2584–2599. [Google Scholar] [CrossRef] [Green Version]
- Inokuchi, M.; Hiroi, J.; Watanabe, S.; Lee, K.M.; Kaneko, T. Gene expression and morphological localization of NHE3, NCC and NKCC1a in branchial mitochondria-rich cells of Mozambique tilapia (Oreochromis mossambicus) acclimated to a wide range of salinities. Comp. Biochem. Phys. A 2008, 151, 151–158. [Google Scholar] [CrossRef]
- Wang, Y.F.; Tseng, Y.C.; Yan, J.J.; Hiroi, J.; Hwang, P.P. Role of SLC12A10.2, a Na-Cl cotransporter-like protein, in a Cl uptake mechanism in zebrafish (Danio rerio). Am. J. Phys.-Reg. I 2009, 296, 1650–1660. [Google Scholar] [CrossRef]
- Kwong, A.K.Y.; Ng, A.H.Y.; Leung, L.Y.; Man, A.K.Y.; Woo, N.Y.S. Effect of extracellular osmolality and ionic levels on pituitary prolactin release in euryhaline silver sea bream (Sparus sarba). Gen. Comp. Endocrinol. 2009, 160, 67–75. [Google Scholar] [CrossRef]
- Kwong, R.W.M.; Kumai, Y.; Perry, S.F. Neuroendocrine control of ionic balance in zebrafish. Gen. Comp. Endocrinol. 2016, 234, 40–46. [Google Scholar] [CrossRef]
- Cutler, C.P.; Cramb, G. Branchial expression of an aquaporin 3 (AQP-3) homologue is downregulated in the European eel Anguilla anguilla following seawater acclimation. J. Exp. Biol. 2002, 205, 2643–2651. [Google Scholar]
- Lignot, J.-H.; Cutler, C.P.; Hazon, N.; Cramb, G. Immunolocalisation of aquaporin 3 in the gill and the gastrointestinal tract of the European eel Anguilla anguilla (L.). J. Exp. Biol. 2002, 205, 2653–2663. [Google Scholar]
- Tse, W.K.F.; Au, D.W.T.; Wong, C.K.C. Characterization of ion channel and transporter mRNA expressions in isolated gill chloride and pavement cells of seawater acclimating eels. Biochem. Biophys. Res. Commun. 2006, 346, 1181–1190. [Google Scholar] [CrossRef] [PubMed]
- Giffard-Mena, I.; Boulo, V.; Aujoulat, F.; Fowden, H.; Castille, R.; Charmantier, G.; Cramb, G. Aquaporin molecular characterization in the sea-bass (Dicentrarchus labrax): The effect of salinity on AQP1 and AQP3 expression. Comp. Biochem. Phys. A 2007, 148, 430–444. [Google Scholar] [CrossRef]
- Giffard-Mena, I.; Lorin-Nebel, C.; Charmantier, G.; Castille, R.; Boulo, V. Adaptation of the sea-bass (Dicentrarchus labrax) to fresh water: Role of aquaporins and Na+/K+-ATPases. Comp. Biochem. Phys. A 2008, 150, 332–338. [Google Scholar] [CrossRef]
- Jeong, S.-Y.; Kim, J.-H.; Lee, W.-O.; Dahms, H.-U.; Han, K.-N. Salinity changes in the anadromous river pufferfish, Takifugu obscurus, mediate gene regulation. Fish Phys. Biochem. 2014, 40, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Deane, E.E.; Woo, N.Y. Tissue distribution, effects of salinity acclimation, and ontogeny of aquaporin 3 in the marine teleost, silver sea bream (Sparus sarba). Mar. Biotechnol. 2006, 8, 663–671. [Google Scholar] [CrossRef]
- Tipsmark, C.K.; Sørensen, K.J.; Madsen, S.S. Aquaporin expression dynamics in osmoregulatory tissues of Atlantic salmon during smoltification and seawater acclimation. J. Exp. Biol. 2010, 213, 368–379. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.K.; Lee, S.Y.; Kim, B.S.; Kim, D.S.; Nam, Y.K. Isolation and mRNA expression analysis of aquaporin isoforms in marine medaka Oryzias dancena, a euryhaline teleost. Comp. Biochem. Phys. A 2014, 171, 1–8. [Google Scholar] [CrossRef]
- Jung, D.; Sato, J.D.; Shaw, J.R.; Stanton, B.A. Expression of aquaporin 3 in gills of the Atlantic killifish (Fundulus heteroclitus): Effects of seawater acclimation. Comp. Biochem. Phys. A 2012, 161, 320–326. [Google Scholar] [CrossRef]
- Choi, Y.J.; Shin, H.S.; Kim, N.N.; Cho, S.H.; Yamamoto, Y.; Ueda, H.; Lee, J.; Choi, C.Y. Expression of aquaporin-3 and-8 mRNAs in the parr and smolt stages of sockeye salmon, Oncorhynchus nerka: Effects of cortisol treatment and seawater acclimation. Comp. Biochem. Phys. A 2013, 165, 228–236. [Google Scholar] [CrossRef]
- An, K.W.; Kim, N.N.; Choi, C.Y. Cloning and expression of aquaporin 1 and arginine vasotocin receptor mRNA from the black porgy, Acanthopagrus schlegeli: Effect of freshwater acclimation. Fish Phys. Biochem. 2008, 34, 185–194. [Google Scholar] [CrossRef]
- Ip, Y.K.; Soh, M.M.L.; Chen, X.L.; Ong, J.L.Y.; Chng, Y.R.; Ching, B.Y.; Wong, W.P.; Lam, S.H.; Chew, S.F. Molecular characterization of branchial aquaporin 1aa and effects of seawater acclimation, emersion or ammonia exposure on its mRNA expression in the gills, gut, kidney and skin of the freshwater climbing perch, Anabas testudineus. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Tipsmark, C.K.; Sørensen, K.J.; Hulgard, K.; Madsen, S.S. Claudin-15 and-25b expression in the intestinal tract of Atlantic salmon in response to seawater acclimation, smoltification and hormone treatment. Comp. Biochem. Phys. A 2010, 155, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Kaneko, T.; Aida, K. Aquaporin-3 expressed in the basolateral membrane of gill chloride cells in Mozambique tilapia Oreochromis mossambicus adapted to freshwater and seawater. J. Exp. Biol. 2005, 208, 2673–2682. [Google Scholar] [CrossRef] [PubMed]
- Brunelli, E.; Mauceri, A.; Salvatore, F.; Giannetto, A.; Maisano, M.; Tripepi, S. Localization of aquaporin 1 and 3 in the gills of the rainbow wrasse. Coris julis. Acta Histochem. 2010, 112, 251–258. [Google Scholar] [CrossRef]
- Hara-Chikuma, M.; Verkman, A. Physiological roles of glycerol-transporting aquaporins: The aquaglyceroporins. Cell. Mol. Life Sci. 2006, 63, 1386–1392. [Google Scholar] [CrossRef] [PubMed]
- King, L.S.; Kozono, D.; Agre, P. From structure to disease: The evolving tale of aquaporin biology. Nat. Rev. Mol. Cell. Biol. 2004, 5, 687. [Google Scholar] [CrossRef]
- Cerdà, J.; Finn, R.N. Piscine aquaporins: An overview of recent advances. J. Exp. Zool. A 2010, 313, 623–650. [Google Scholar] [CrossRef]
- Kwong, R.W.M.; Kumai, Y.; Perry, S.F. The role of aquaporin and tight junction proteins in the regulation of water movement in larval zebrafish (Danio rerio). PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Chen, L.-M.; Zhao, J.; Musa-Aziz, R.; Pelletier, M.F.; Drummond, I.A.; Boron, W.F. Cloning and characterization of a zebrafish homologue of human AQP1: A bifunctional water and gas channel. Am. J. Phys.-Reg. I 2010, 299, 1163–1174. [Google Scholar] [CrossRef]
- Horng, J.-L.; Chao, P.-L.; Chen, P.-Y.; Shih, T.-H.; Lin, L.-Y. Aquaporin 1 is involved in acid secretion by ionocytes of zebrafish embryos through facilitating CO2 transport. PLoS ONE 2015, 10, e0136440. [Google Scholar] [CrossRef]
- Jung, H.J.; Kwon, T.-H. Molecular mechanisms regulating aquaporin-2 in kidney collecting duct. Am. J. Phys.-Renal 2016, 311, 1318–1328. [Google Scholar] [CrossRef]
- Cutler, C.P.; Phillips, C.; Hazon, N.; Cramb, G. Cortisol regulates eel (Anguilla anguilla) aquaporin 3 (AQP3) mRNA expression levels in gill. Gen. Comp. Endocrinol. 2007, 152, 310–313. [Google Scholar] [CrossRef]
- Breves, J.P.; Watanabe, S.; Kaneko, T.; Hirano, T.; Grau, E.G. Prolactin restores branchial mitochondrion-rich cells expressing Na+/Cl− cotransporter in hypophysectomized Mozambique tilapia. Am. J. Phys.-Reg. I 2010, 299, 702–710. [Google Scholar] [CrossRef]
- Watanabe, S.; Hirano, T.; Grau, E.G.; Kaneko, T. Osmosensitivity of prolactin cells is enhanced by the water channel aquaporin-3 in a euryhaline Mozambique tilapia (Oreochromis mossambicus). Am. J. Phys.-Reg. I 2009, 296, 446–453. [Google Scholar] [CrossRef]













© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ellis, L.V.; Bollinger, R.J.; Weber, H.M.; Madsen, S.S.; Tipsmark, C.K. Differential Expression and Localization of Branchial AQP1 and AQP3 in Japanese Medaka (Oryzias latipes). Cells 2019, 8, 422. https://doi.org/10.3390/cells8050422
Ellis LV, Bollinger RJ, Weber HM, Madsen SS, Tipsmark CK. Differential Expression and Localization of Branchial AQP1 and AQP3 in Japanese Medaka (Oryzias latipes). Cells. 2019; 8(5):422. https://doi.org/10.3390/cells8050422
Chicago/Turabian StyleEllis, Laura V., Rebecca J. Bollinger, Hannah M. Weber, Steffen S. Madsen, and Christian K. Tipsmark. 2019. "Differential Expression and Localization of Branchial AQP1 and AQP3 in Japanese Medaka (Oryzias latipes)" Cells 8, no. 5: 422. https://doi.org/10.3390/cells8050422
APA StyleEllis, L. V., Bollinger, R. J., Weber, H. M., Madsen, S. S., & Tipsmark, C. K. (2019). Differential Expression and Localization of Branchial AQP1 and AQP3 in Japanese Medaka (Oryzias latipes). Cells, 8(5), 422. https://doi.org/10.3390/cells8050422