Nucleolar Structure and Function in Trypanosomatid Protozoa



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
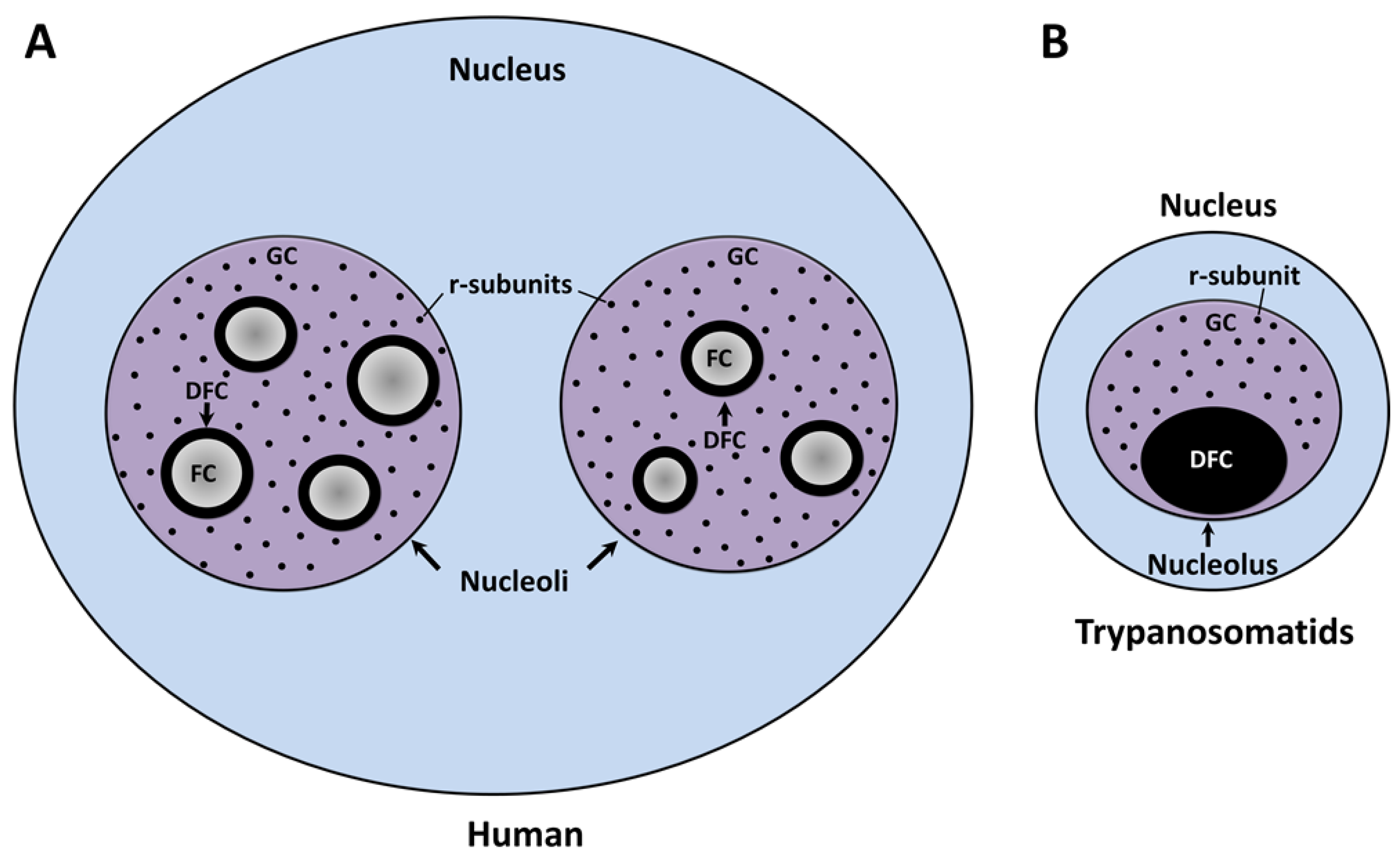
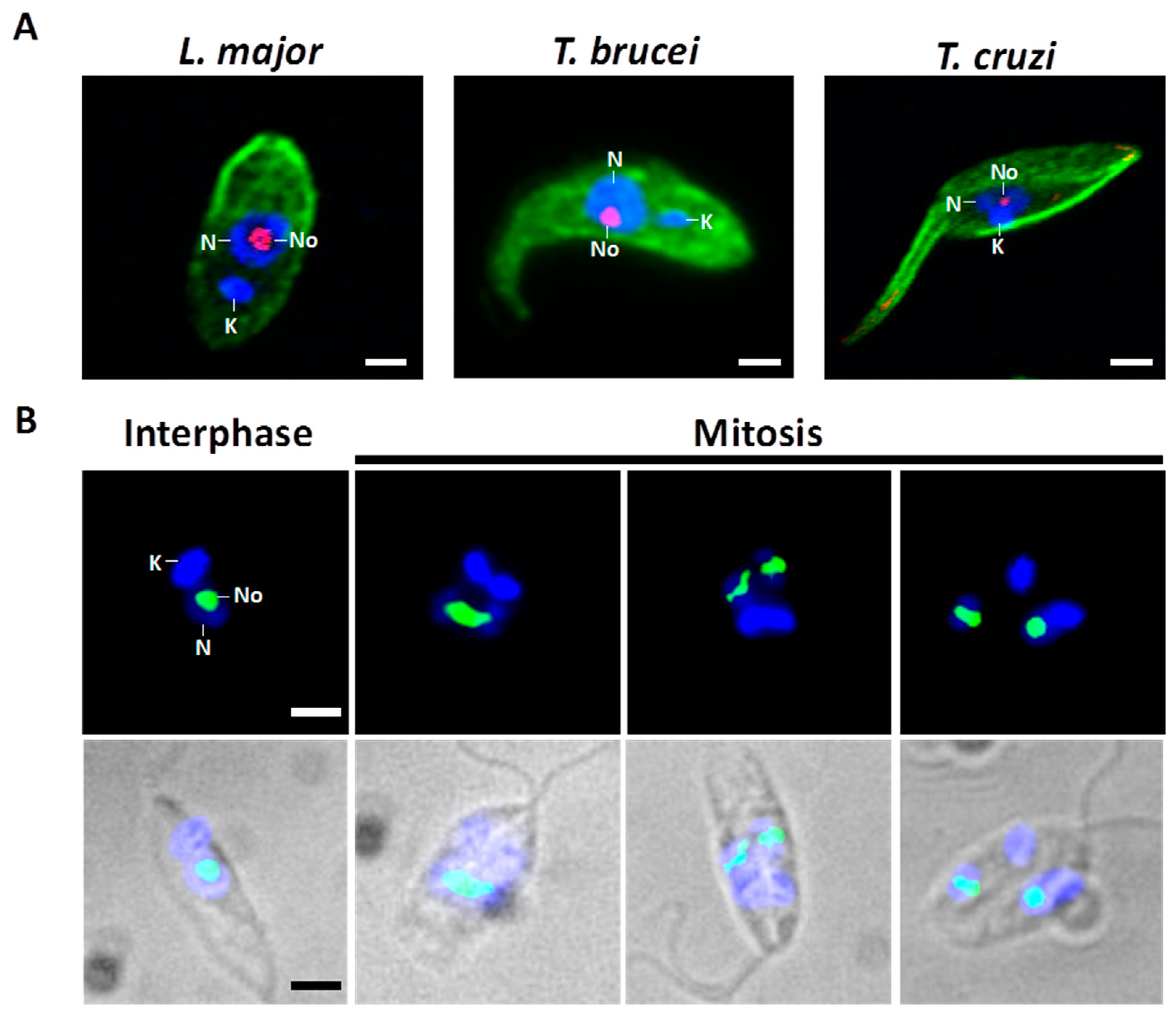
2. Nucleolar Structure
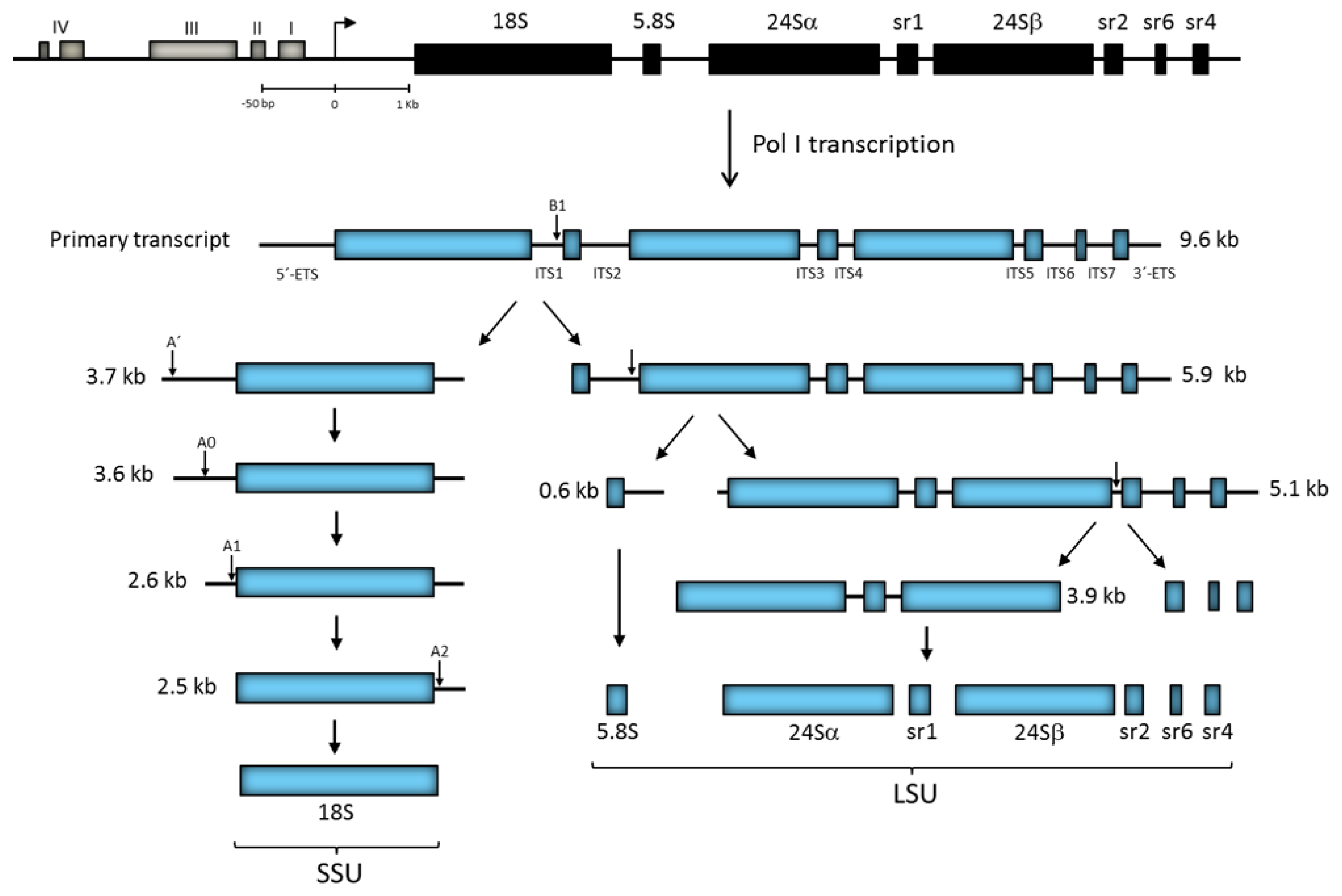
3. Ribosomal RNA Genes
3.1. rRNA Gene Repeats
3.2. Transcription of the rRNA Gene Repeat
3.2.1. Promoter Regions
3.2.2. Transcription Factors and Pol I Subunits
3.2.3. Epigenetic Regulation of rRNA Gene Repeats
3.3. Gene Organization and Transcription of 5S rRNA Genes
4. Processing and Nucleotide Modifications in rRNA
4.1. Roles of snoRNAs in rRNA Maturation
4.2. Primary Transcript Processing
4.3. Modifications in rRNA Molecules
5. Ribosome Biogenesis
6. Other Functions of the Nucleolus
6.1. Stress Response
6.2. Transcription and Processing of mRNAs Encoding Procyclins
7. Conclusions
8. Future Outlook
Author Contributions
Funding
Conflicts of Interest
References
- Smirnov, E.; Cmarko, D.; Mazel, T.; Hornacek, M.; Raska, I. Nucleolar DNA: The host and the guests. Histochem. Cell Biol. 2016, 145, 359–372. [Google Scholar] [CrossRef]
- McStay, B. Nucleolar organizer regions: Genomic ‘dark matter’ requiring illumination. Genes Dev. 2016, 30, 1598–1610. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.; Zomerdijk, J.C. RNA-polymerase-I-directed rDNA transcription, life and works. Trends Biochem. Sci. 2005, 30, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koberna, K.; Malinsky, J.; Pliss, A.; Masata, M.; Vecerova, J.; Fialova, M.; Bednar, J.; Raska, I. Ribosomal genes in focus: New transcripts label the dense fibrillar components and form clusters indicative of “Christmas trees” in situ. J. Cell Biol. 2002, 157, 743–748. [Google Scholar] [PubMed]
- Huang, S. Building an efficient factory: Where is pre-rRNA synthesized in the nucleolus? J. Cell Biol. 2002, 157, 739–741. [Google Scholar]
- Matera, A.G.; Terns, R.M.; Terns, M.P. Non-coding RNAs: Lessons from the small nuclear and small nucleolar RNAs. Nat. Rev. Mol. Cell Biol. 2007, 8, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Thiry, M.; Lamaye, F.; Lafontaine, D.L. The nucleolus: When 2 became 3. Nucleus 2011, 2, 289–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pena, C.; Hurt, E.; Panse, V.G. Eukaryotic ribosome assembly, transport and quality control. Nat. Struct. Mol. Biol. 2017, 24, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Leung, E.; Brown, J.D. Biogenesis of the signal recognition particle. Biochem. Soc. Trans. 2010, 38, 1093–1098. [Google Scholar] [CrossRef] [Green Version]
- Boulon, S.; Westman, B.J.; Hutten, S.; Boisvert, F.M.; Lamond, A.I. The nucleolus under stress. Mol. Cell 2010, 40, 216–227. [Google Scholar] [CrossRef]
- Field, M.C.; Horn, D.; Fairlamb, A.H.; Ferguson, M.A.J.; Gray, D.W.; Read, K.D.; De Rycker, M.; Torrie, L.S.; Wyatt, P.G.; Wyllie, S.; et al. Anti-trypanosomatid drug discovery: An ongoing challenge and a continuing need. Nat. Rev. Microbiol. 2017, 15, 447. [Google Scholar] [CrossRef]
- Clayton, C.; Shapira, M. Post-transcriptional regulation of gene expression in trypanosomes and leishmanias. Mol. Biochem. Parasitol. 2007, 156, 93–101. [Google Scholar] [CrossRef]
- Haile, S.; Papadopoulou, B. Developmental regulation of gene expression in trypanosomatid parasitic protozoa. Curr. Opin. Microbiol. 2007, 10, 569–577. [Google Scholar] [CrossRef]
- Bern, C. Chagas’ Disease. N. Engl. J. Med. 2015, 373, 456–466. [Google Scholar] [CrossRef]
- Perez-Molina, J.A.; Molina, I. Chagas disease. Lancet 2018, 391, 82–94. [Google Scholar] [CrossRef]
- Borghi, S.M.; Fattori, V.; Conchon-Costa, I.; Pinge-Filho, P.; Pavanelli, W.R.; Verri, W.A., Jr. Leishmania infection: Painful or painless? Parasitol. Res. 2017, 116, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Matthews, K.R.; Ellis, J.R.; Paterou, A. Molecular regulation of the life cycle of African trypanosomes. Trends Parasitol. 2004, 20, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Franco, J.R.; Simarro, P.P.; Diarra, A.; Jannin, J.G. Epidemiology of human African trypanosomiasis. Clin. Epidemiol. 2014, 6, 257–275. [Google Scholar] [Green Version]
- Stuart, K.; Panigrahi, A.K. RNA editing: Complexity and complications. Mol. Microbiol. 2002, 45, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Read, L.K.; Lukes, J.; Hashimi, H. Trypanosome RNA editing: The complexity of getting U in and taking U out. Wiley Interdiscip. Rev. RNA 2016, 7, 33–51. [Google Scholar] [CrossRef]
- Martinez-Calvillo, S.; Vizuet-de-Rueda, J.C.; Florencio-Martinez, L.E.; Manning-Cela, R.G.; Figueroa-Angulo, E.E. Gene expression in trypanosomatid parasites. J. Biomed. Biotechnol. 2010, 2010, 525241. [Google Scholar] [CrossRef] [PubMed]
- Clayton, C.E. Gene expression in Kinetoplastids. Curr. Opin. Microbiol. 2016, 32, 46–51. [Google Scholar] [CrossRef]
- Michaeli, S. Trans-splicing in trypanosomes: Machinery and its impact on the parasite transcriptome. Future Microbiol. 2011, 6, 459–474. [Google Scholar] [CrossRef]
- Gunzl, A.; Kirkham, J.K.; Nguyen, T.N.; Badjatia, N.; Park, S.H. Mono-allelic VSG expression by RNA polymerase I in Trypanosoma brucei: Expression site control from both ends? Gene 2015, 556, 68–73. [Google Scholar] [CrossRef]
- Schwarzacher, H.G.; Mosgoeller, W. Ribosome biogenesis in man: Current views on nucleolar structures and function. Cytogenet. Cell Genet. 2000, 91, 243–252. [Google Scholar] [CrossRef]
- Hernandez-Verdun, D.; Roussel, P.; Thiry, M.; Sirri, V.; Lafontaine, D.L. The nucleolus: Structure/function relationship in RNA metabolism. Wiley Interdiscip. Rev. RNA 2010, 1, 415–431. [Google Scholar] [CrossRef]
- Jimenez-Garcia, L.F.; Segura-Valdez, M.L.; Ochs, R.L.; Rothblum, L.I.; Hannan, R.; Spector, D.L. Nucleologenesis: U3 snRNA-containing prenucleolar bodies move to sites of active pre-rRNA transcription after mitosis. Mol. Biol. Cell 1994, 5, 955–966. [Google Scholar] [CrossRef]
- Dimario, P.J. Cell and molecular biology of nucleolar assembly and disassembly. Int. Rev. Cytol. 2004, 239, 99–178. [Google Scholar]
- Hernandez-Verdun, D. Assembly and disassembly of the nucleolus during the cell cycle. Nucleus 2011, 2, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Ogbadoyi, E.; Ersfeld, K.; Robinson, D.; Sherwin, T.; Gull, K. Architecture of the Trypanosoma brucei nucleus during interphase and mitosis. Chromosoma 2000, 108, 501–513. [Google Scholar] [CrossRef]
- Lopez-Velazquez, G.; Hernandez, R.; Lopez-Villasenor, I.; Reyes-Vivas, H.; Segura-Valdez, M.L.; Jimenez-Garcia, L.F. Electron microscopy analysis of the nucleolus of Trypanosoma cruzi. Microsc. Microanal. 2005, 11, 293–299. [Google Scholar] [CrossRef]
- Nepomuceno-Mejia, T.; Lara-Martinez, R.; Cevallos, A.M.; Lopez-Villasenor, I.; Jimenez-Garcia, L.F.; Hernandez, R. The Trypanosoma cruzi nucleolus: A morphometrical analysis of cultured epimastigotes in the exponential and stationary phases. FEMS Microbiol. Lett. 2010, 313, 41–46. [Google Scholar] [CrossRef]
- Knibiehler, B.; Mirre, C.; Navarro, A.; Rosset, R. Studies on chromatin organization in a nucleolus without fibrillar centres. Presence of a sub-nucleolar structure in KCo cells of Drosophila. Cell Tissue Res. 1984, 236, 279–288. [Google Scholar] [CrossRef]
- Thiry, M.; Lafontaine, D.L. Birth of a nucleolus: The evolution of nucleolar compartments. Trends Cell Biol. 2005, 15, 194–199. [Google Scholar] [CrossRef]
- Jimenez-Garcia, L.F.; Zavala, G.; Chavez-Munguia, B.; Ramos-Godinez, M.P.; Lopez-Velazquez, G.; Segura-Valdez, M.L.; Montanez, C.; Hehl, A.B.; Arguello-Garcia, R.; Ortega-Pierres, G. Identification of nucleoli in the early branching protist Giardia duodenalis. Int. J. Parasitol. 2008, 38, 1297–1304. [Google Scholar] [CrossRef]
- Kumar, G.; Kajuluri, L.P.; Gupta, C.M.; Sahasrabuddhe, A.A. A twinfilin-like protein coordinates karyokinesis by influencing mitotic spindle elongation and DNA replication in Leishmania. Mol. Microbiol. 2016, 100, 173–187. [Google Scholar] [CrossRef]
- Nepomuceno-Mejia, T.; Lara-Martinez, R.; Hernandez, R.; Segura-Valdez, M.L.; Jimenez-Garcia, L.F. Nucleologenesis in Trypanosoma cruzi. Microsc. Microanal. 2016, 22, 621–629. [Google Scholar] [CrossRef]
- Nepomuceno-Mejia, T.; Florencio-Martinez, L.E.; Martinez-Calvillo, S. Nucleolar Division in the Promastigote Stage of Leishmania major Parasite: A Nop56 Point of View. Biomed. Res. Int. 2018, 2018, 1641839. [Google Scholar] [CrossRef]
- Paule, M.R.; White, R.J. Survey and summary: Transcription by RNA polymerases I and III. Nucleic Acids Res. 2000, 28, 1283–1298. [Google Scholar] [CrossRef]
- Goodfellow, S.J.; Zomerdijk, J.C. Basic mechanisms in RNA polymerase I transcription of the ribosomal RNA genes. Subcell. Biochem. 2012, 61, 211–236. [Google Scholar]
- Torres-Machorro, A.L.; Hernandez, R.; Cevallos, A.M.; Lopez-Villasenor, I. Ribosomal RNA genes in eukaryotic microorganisms: Witnesses of phylogeny? FEMS Microbiol. Rev. 2010, 34, 59–86. [Google Scholar] [CrossRef]
- Hasan, G.; Turner, M.J.; Cordingley, J.S. Ribosomal RNA genes of Trypanosoma brucei: Mapping the regions specifying the six small ribosomal RNAs. Gene 1984, 27, 75–86. [Google Scholar] [CrossRef]
- Spencer, D.F.; Collings, J.C.; Schnare, M.N.; Gray, M.W. Multiple spacer sequences in the nuclear large subunit ribosomal RNA gene of Crithidia fasciculata. EMBO J. 1987, 6, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, R.; Diaz de Leon, F.; Castaneda, M. Molecular cloning and partial characterization of ribosomal RNA genes from Trypanosoma cruzi. Mol. Biochem. Parasitol. 1988, 27, 275–279. [Google Scholar] [CrossRef]
- Martinez-Calvillo, S.; Sunkin, S.M.; Yan, S.; Fox, M.; Stuart, K.; Myler, P.J. Genomic organization and functional characterization of the Leishmania major Friedlin ribosomal RNA gene locus. Mol. Biochem. Parasitol. 2001, 116, 147–157. [Google Scholar] [CrossRef]
- Hashem, Y.; des Georges, A.; Fu, J.; Buss, S.N.; Jossinet, F.; Jobe, A.; Zhang, Q.; Liao, H.Y.; Grassucci, R.A.; Bajaj, C.; et al. High-resolution cryo-electron microscopy structure of the Trypanosoma brucei ribosome. Nature 2013, 494, 385–389. [Google Scholar] [CrossRef]
- Liu, Z.; Gutierrez-Vargas, C.; Wei, J.; Grassucci, R.A.; Ramesh, M.; Espina, N.; Sun, M.; Tutuncuoglu, B.; Madison-Antenucci, S.; Woolford, J.L., Jr.; et al. Structure and assembly model for the Trypanosoma cruzi 60S ribosomal subunit. Proc. Natl. Acad. Sci. USA 2016, 113, 12174–12179. [Google Scholar] [CrossRef]
- Sollner-Webb, B.; Mougey, E.B. News from the nucleolus: rRNA gene expression. Trends Biochem. Sci. 1991, 16, 58–62. [Google Scholar] [CrossRef]
- Ivens, A.C.; Peacock, C.S.; Worthey, E.A.; Murphy, L.; Aggarwal, G.; Berriman, M.; Sisk, E.; Rajandream, M.A.; Adlem, E.; Aert, R.; et al. The genome of the kinetoplastid parasite, Leishmania major. Science 2005, 309, 436–442. [Google Scholar] [CrossRef]
- van Leeuwen, F.; Kieft, R.; Cross, M.; Borst, P. Tandemly repeated DNA is a target for the partial replacement of thymine by beta-D-glucosyl-hydroxymethyluracil in Trypanosoma brucei. Mol. Biochem. Parasitol. 2000, 109, 133–145. [Google Scholar] [CrossRef]
- Janz, L.; Clayton, C. The PARP and rRNA promoters of Trypanosoma brucei are composed of dissimilar sequence elements that are functionally interchangeable. Mol. Cell Biol. 1994, 14, 5804–5811. [Google Scholar] [CrossRef] [PubMed]
- Zomerdijk, J.C.; Kieft, R.; Shiels, P.G.; Borst, P. Alpha-amanitin-resistant transcription units in trypanosomes: A comparison of promoter sequences for a VSG gene expression site and for the ribosomal RNA genes. Nucleic Acids Res. 1991, 19, 5153–5158. [Google Scholar] [CrossRef]
- Laufer, G.; Schaaf, G.; Bollgonn, S.; Gunzl, A. In vitro analysis of alpha-amanitin-resistant transcription from the rRNA, procyclic acidic repetitive protein, and variant surface glycoprotein gene promoters in Trypanosoma brucei. Mol. Cell Biol. 1999, 19, 5466–5473. [Google Scholar] [CrossRef] [PubMed]
- Gunzl, A.; Vanhamme, L.; Myler, P.J. Transcription in trypanosomes: A different means to the end. In Trypanosomes: After the Genome; Barry, J.D., McCulloch, R., Mottram, J.C., Acosta-Serrano, A., Eds.; Horizon Bioscience: Wymonham, UK, 2007; pp. 177–208. [Google Scholar]
- Uliana, S.R.; Fischer, W.; Stempliuk, V.A.; Floeter-Winter, L.M. Structural and functional characterization of the Leishmania amazonensis ribosomal RNA promoter. Mol. Biochem. Parasitol. 1996, 76, 245–255. [Google Scholar] [CrossRef]
- Gay, L.S.; Wilson, M.E.; Donelson, J.E. The promoter for the ribosomal RNA genes of Leishmania chagasi. Mol. Biochem. Parasitol. 1996, 77, 193–200. [Google Scholar] [CrossRef]
- Yan, S.; Lodes, M.J.; Fox, M.; Myler, P.J.; Stuart, K. Characterization of the Leishmania donovani ribosomal RNA promoter. Mol. Biochem. Parasitol. 1999, 103, 197–210. [Google Scholar] [CrossRef]
- Martinez-Calvillo, S.; Hernandez, R. Trypanosoma cruzi ribosomal DNA: Mapping of a putative distal promoter. Gene 1994, 142, 243–247. [Google Scholar] [CrossRef]
- Figueroa-Angulo, E.; Martinez-Calvillo, S.; Lopez-Villasenor, I.; Hernandez, R. Evidence supporting a major promoter in the Trypanosoma cruzi rRNA gene. FEMS Microbiol. Lett. 2003, 225, 221–225. [Google Scholar] [CrossRef]
- Stolf, B.S.; Souto, R.P.; Pedroso, A.; Zingales, B. Two types of ribosomal RNA genes in hybrid Trypanosoma cruzi strains. Mol. Biochem. Parasitol. 2003, 126, 73–80. [Google Scholar] [CrossRef]
- de Andrade Stempliuk, V.; Floeter-Winter, L.M. Functional domains of the rDNA promoter display a differential recognition in Leishmania. Int. J. Parasitol. 2002, 32, 437–447. [Google Scholar] [CrossRef]
- Figueroa-Angulo, E.; Cevallos, A.M.; Zentella, A.; Lopez-Villasenor, I.; Hernandez, R. Potential regulatory elements in the Trypanosoma cruzi rRNA gene promoter. Biochim. Biophys. Acta 2006, 1759, 497–501. [Google Scholar] [CrossRef]
- Hernandez, R.; Cevallos, A.M. Ribosomal RNA gene transcription in trypanosomes. Parasitol. Res. 2014, 113, 2415–2424. [Google Scholar] [CrossRef] [PubMed]
- Requena, J.M.; Soto, M.; Quijada, L.; Carrillo, G.; Alonso, C. A region containing repeated elements is associated with transcriptional termination of Leishmania infantum ribosomal RNA genes. Mol. Biochem. Parasitol. 1997, 84, 101–110. [Google Scholar] [CrossRef]
- Abreu-Blanco, M.T.; Ramirez, J.L.; Pinto-Santini, D.M.; Papadopoulou, B.; Guevara, P. Analysis of ribosomal RNA transcription termination and 3’ end processing in Leishmania amazonensis. Gene 2010, 451, 15–22. [Google Scholar] [CrossRef]
- Thomas, S.; Green, A.; Sturm, N.R.; Campbell, D.A.; Myler, P.J. Histone acetylations mark origins of polycistronic transcription in Leishmania major. BMC. Genom. 2009, 10, 152. [Google Scholar] [CrossRef]
- Brandenburg, J.; Schimanski, B.; Nogoceke, E.; Nguyen, T.N.; Padovan, J.C.; Chait, B.T.; Cross, G.A.; Gunzl, A. Multifunctional class I transcription in Trypanosoma brucei depends on a novel protein complex. EMBO J. 2007, 26, 4856–4866. [Google Scholar] [CrossRef]
- Kirkham, J.K.; Park, S.H.; Nguyen, T.N.; Lee, J.H.; Gunzl, A. Dynein Light Chain LC8 Is Required for RNA Polymerase I-Mediated Transcription in Trypanosoma brucei, Facilitating Assembly and Promoter Binding of Class I Transcription Factor, A. Mol. Cell Biol. 2016, 36, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.N.; Nguyen, B.N.; Lee, J.H.; Panigrahi, A.K.; Gunzl, A. Characterization of a novel class I transcription factor A (CITFA) subunit that is indispensable for transcription by the multifunctional RNA polymerase I of Trypanosoma brucei. Eukaryot. Cell 2012, 11, 1573–1581. [Google Scholar] [CrossRef]
- Alsford, S.; Horn, D. Elongator Protein 3b Negatively Regulates Ribosomal DNA Transcription in African Trypanosomes. Mol. Cell Biol. 2011, 31, 1822–1832. [Google Scholar] [CrossRef]
- Moreno-Campos, R.; Florencio-Martinez, L.E.; Nepomuceno-Mejia, T.; Rojas-Sanchez, S.; Velez-Ramirez, D.E.; Padilla-Mejia, N.E.; Figueroa-Angulo, E.; Manning-Cela, R.; Martinez-Calvillo, S. Molecular characterization of 5S ribosomal RNA genes and transcripts in the protozoan parasite Leishmania major. Parasitology 2016, 143, 1917–1929. [Google Scholar] [CrossRef] [PubMed]
- Walgraffe, D.; Devaux, S.; Lecordier, L.; Dierick, J.F.; Dieu, M.; Van den Abbeele, J.; Pays, E.; Vanhamme, L. Characterization of subunits of the RNA polymerase I complex in Trypanosoma brucei. Mol. Biochem. Parasitol. 2005, 139, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.N.; Schimanski, B.; Zahn, A.; Klumpp, B.; Gunzl, A. Purification of an eight subunit RNA polymerase I complex in Trypanosoma brucei. Mol. Biochem. Parasitol. 2006, 149, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.N.; Schimanski, B.; Gunzl, A. Active RNA polymerase I of Trypanosoma brucei harbors a novel subunit essential for transcription. Mol. Cell Biol. 2007, 27, 6254–6263. [Google Scholar] [CrossRef]
- Kelly, S.; Wickstead, B.; Gull, K. An in silico analysis of trypanosomatid RNA polymerases: Insights into their unusual transcription. Biochem. Soc. Trans. 2005, 33, 1435–1437. [Google Scholar] [CrossRef] [PubMed]
- Canela-Perez, I.; Lopez-Villasenor, I.; Cevallos, A.M.; Hernandez, R. Nuclear distribution of the Trypanosoma cruzi RNA Pol I subunit RPA31 during growth and metacyclogenesis, and characterization of its nuclear localization signal. Parasitol. Res. 2018, 117, 911–918. [Google Scholar] [CrossRef]
- Grummt, I. Different epigenetic layers engage in complex crosstalk to define the epigenetic state of mammalian rRNA genes. Hum. Mol. Genet. 2007, 16, R21–R27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, R.J.; Pikaard, C.S. Chromatin turn ons and turn offs of ribosomal RNA genes. Cell Cycle 2004, 3, 880–883. [Google Scholar] [CrossRef]
- McStay, B.; Grummt, I. The epigenetics of rRNA genes: From molecular to chromosome biology. Annu. Rev. Cell Dev. Biol. 2008, 24, 131–157. [Google Scholar] [CrossRef]
- Hamperl, S.; Wittner, M.; Babl, V.; Perez-Fernandez, J.; Tschochner, H.; Griesenbeck, J. Chromatin states at ribosomal DNA loci. Biochim. Biophys. Acta 2013, 1829, 405–417. [Google Scholar] [CrossRef]
- Zentner, G.E.; Balow, S.A.; Scacheri, P.C. Genomic characterization of the mouse ribosomal DNA locus. G3 (Bethesda) 2014, 4, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Zentner, G.E.; Saiakhova, A.; Manaenkov, P.; Adams, M.D.; Scacheri, P.C. Integrative genomic analysis of human ribosomal DNA. Nucleic Acids Res. 2011, 39, 4949–4960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vizuet-de-Rueda, J.C.; Florencio-Martinez, L.E.; Padilla-Mejia, N.E.; Manning-Cela, R.; Hernandez-Rivas, R.; Martinez-Calvillo, S. Ribosomal RNA Genes in the Protozoan Parasite Leishmania major Possess a Nucleosomal Structure. Protist 2016, 167, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Stanne, T.M.; Rudenko, G. Active VSG expression sites in Trypanosoma brucei are depleted of nucleosomes. Eukaryot. Cell 2010, 9, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, L.M.; Cross, G.A. Nucleosomes are depleted at the VSG expression site transcribed by RNA polymerase I in African trypanosomes. Eukaryot. Cell 2010, 9, 148–154. [Google Scholar] [CrossRef]
- Jones, H.S.; Kawauchi, J.; Braglia, P.; Alen, C.M.; Kent, N.A.; Proudfoot, N.J. RNA polymerase I in yeast transcribes dynamic nucleosomal rDNA. Nat. Struct. Mol. Biol. 2007, 14, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Calvillo, S.; Romero-Meza, G.; Vizuet-de-Rueda, J.C.; Florencio-Martinez, L.E.; Manning-Cela, R.; Nepomuceno-Mejia, T. Epigenetic Regulation of Transcription in Trypanosomatid Protozoa. Curr. Genom. 2018, 19, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Stanne, T.M.; Narayanan, M.S.; Ridewood, S.; Ling, A.; Witmer, K.; Kushwaha, M.; Wiesler, S.; Wickstead, B.; Wood, J.; Rudenko, G. Identification of the ISWI Chromatin Remodeling Complex of the Early Branching Eukaryote Trypanosoma brucei. J. Biol. Chem. 2015, 290, 26954–26967. [Google Scholar] [CrossRef]
- Denninger, V.; Rudenko, G. FACT plays a major role in histone dynamics affecting VSG expression site control in Trypanosoma brucei. Mol. Microbiol. 2014, 94, 945–962. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, M.S.; Rudenko, G. TDP1 is an HMG chromatin protein facilitating RNA polymerase I transcription in African trypanosomes. Nucleic Acids Res. 2013, 41, 2981–2992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, I.M. RNA polymerase III. Genes, factors and transcriptional specificity. Eur. J. Biochem. 1993, 212, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Venema, J.; Tollervey, D. Ribosome synthesis in Saccharomyces cerevisiae. Annu. Rev. Genet. 1999, 33, 261–311. [Google Scholar] [CrossRef]
- Cordingley, J.S. Nucleotide sequence of the 5S ribosomal RNA gene repeat of Trypanosoma brucei. Mol. Biochem. Parasitol. 1985, 17, 321–330. [Google Scholar] [CrossRef]
- Hernandez-Rivas, R.; Martinez-Calvillo, S.; Romero, M.; Hernandez, R. Trypanosoma cruzi 5S rRNA genes: Molecular cloning, structure and chromosomal organization. FEMS Microbiol. Lett. 1992, 71, 63–67. [Google Scholar] [CrossRef]
- Haeusler, R.A.; Engelke, D.R. Spatial organization of transcription by RNA polymerase III. Nucleic Acids Res. 2006, 34, 4826–4836. [Google Scholar] [CrossRef] [Green Version]
- Highett, M.I.; Beven, A.F.; Shaw, P.J. Localization of 5S genes and transcripts in Pisum sativum nuclei. J. Cell Sci. 1993, 105, 1151–1158. [Google Scholar] [PubMed]
- Matera, A.G.; Frey, M.R.; Margelot, K.; Wolin, S.L. A perinucleolar compartment contains several RNA polymerase III transcripts as well as the polypyrimidine tract-binding protein, hnRNP I. J. Cell Biol. 1995, 129, 1181–1193. [Google Scholar] [CrossRef] [PubMed]
- Ersfeld, K.; Gull, K. Partitioning of large and minichromosomes in Trypanosoma brucei. Science 1997, 276, 611–614. [Google Scholar] [CrossRef]
- Watkins, N.J.; Bohnsack, M.T. The box C/D and H/ACA snoRNPs: Key players in the modification, processing and the dynamic folding of ribosomal RNA. Wiley Interdiscip. Rev. RNA 2012, 3, 397–414. [Google Scholar] [CrossRef]
- Geiduschek, E.P.; Kassavetis, G.A. The RNA polymerase III transcription apparatus. J. Mol. Biol. 2001, 310, 1–26. [Google Scholar] [CrossRef]
- Lenardo, M.J.; Dorfman, D.M.; Reddy, L.V.; Donelson, J.E. Characterization of the Trypanosoma brucei 5S ribosomal RNA gene and transcript: The 5S rRNA is a spliced-leader-independent species. Gene 1985, 35, 131–141. [Google Scholar] [CrossRef]
- Velez-Ramirez, D.E.; Florencio-Martinez, L.E.; Romero-Meza, G.; Rojas-Sanchez, S.; Moreno-Campos, R.; Arroyo, R.; Ortega-Lopez, J.; Manning-Cela, R.; Martinez-Calvillo, S. BRF1, a subunit of RNA polymerase III transcription factor TFIIIB, is essential for cell growth of Trypanosoma brucei. Parasitology 2015, 142, 1563–1573. [Google Scholar] [CrossRef]
- Romero-Meza, G.; Velez-Ramirez, D.E.; Florencio-Martinez, L.E.; Roman-Carraro, F.C.; Manning-Cela, R.; Hernandez-Rivas, R.; Martinez-Calvillo, S. Maf1 is a negative regulator of transcription in Trypanosoma brucei. Mol. Microbiol. 2017, 103, 452–468. [Google Scholar] [CrossRef]
- Lombrana, R.; Alvarez, A.; Fernandez-Justel, J.M.; Almeida, R.; Poza-Carrion, C.; Gomes, F.; Calzada, A.; Requena, J.M.; Gomez, M. Transcriptionally Driven DNA Replication Program of the Human Parasite Leishmania major. Cell Rep. 2016, 16, 1774–1786. [Google Scholar] [CrossRef]
- Tschochner, H.; Hurt, E. Pre-ribosomes on the road from the nucleolus to the cytoplasm. Trends Cell Biol. 2003, 13, 255–263. [Google Scholar] [CrossRef]
- Tomecki, R.; Sikorski, P.J.; Zakrzewska-Placzek, M. Comparison of preribosomal RNA processing pathways in yeast, plant and human cells—Focus on coordinated action of endo- and exoribonucleases. FEBS Lett. 2017, 591, 1801–1850. [Google Scholar] [CrossRef]
- Quinternet, M.; Chagot, M.E.; Rothe, B.; Tiotiu, D.; Charpentier, B.; Manival, X. Structural Features of the Box C/D snoRNP Pre-assembly Process Are Conserved through Species. Structure 2016, 24, 1693–1706. [Google Scholar] [CrossRef]
- Guerrero-Sanchez, E.; Cevallos, A.M.; Martinez-Calvillo, S.; Lopez-Villaseñor, I.; Hernandez, R. Relocation of nucleolar fibrillarin in Trypanosoma cruzi during stationary phase. Parasitol. Open 2015, 1, 1–8. [Google Scholar] [CrossRef]
- Barth, S.; Shalem, B.; Hury, A.; Tkacz, I.D.; Liang, X.H.; Uliel, S.; Myslyuk, I.; Doniger, T.; Salmon-Divon, M.; Unger, R.; et al. Elucidating the role of C/D snoRNA in rRNA processing and modification in Trypanosoma brucei. Eukaryot. Cell 2008, 7, 86–101. [Google Scholar] [CrossRef] [PubMed]
- Barth, S.; Hury, A.; Liang, X.H.; Michaeli, S. Elucidating the role of H/ACA-like RNAs in trans-splicing and rRNA processing via RNA interference silencing of the Trypanosoma brucei CBF5 pseudouridine synthase. J. Biol. Chem. 2005, 280, 34558–34568. [Google Scholar] [CrossRef]
- Ruan, J.P.; Ullu, E.; Tschudi, C. Characterization of the Trypanosoma brucei cap hypermethylase Tgs1. Mol. Biochem. Parasitol. 2007, 155, 66–69. [Google Scholar] [CrossRef]
- Preti, M.; Ribeyre, C.; Pascali, C.; Bosio, M.C.; Cortelazzi, B.; Rougemont, J.; Guarnera, E.; Naef, F.; Shore, D.; Dieci, G. The telomere-binding protein Tbf1 demarcates snoRNA gene promoters in Saccharomyces cerevisiae. Mol. Cell 2010, 38, 614–620. [Google Scholar] [CrossRef]
- Li, T.; Zhou, X.; Wang, X.; Zhu, D.; Zhang, Y. Identification and characterization of human snoRNA core promoters. Genomics 2010, 96, 50–56. [Google Scholar] [CrossRef]
- Liang, X.H.; Uliel, S.; Hury, A.; Barth, S.; Doniger, T.; Unger, R.; Michaeli, S. A genome-wide analysis of C/D and H/ACA-like small nucleolar RNAs in Trypanosoma brucei reveals a trypanosome-specific pattern of rRNA modification. RNA 2005, 11, 619–645. [Google Scholar] [CrossRef]
- Eliaz, D.; Doniger, T.; Tkacz, I.D.; Biswas, V.K.; Gupta, S.K.; Kolev, N.G.; Unger, R.; Ullu, E.; Tschudi, C.; Michaeli, S. Genome-wide analysis of small nucleolar RNAs of Leishmania major reveals a rich repertoire of RNAs involved in modification and processing of rRNA. RNA Biol. 2015, 12, 1222–1255. [Google Scholar] [CrossRef]
- Chikne, V.; Gupta, S.K.; Doniger, T.; SR, K.; Cohen-Chalamish, S.; Waldman Ben-Asher, H.; Kolet, L.; Yahia, N.H.; Unger, R.; Ullu, E.; et al. The Canonical Poly (A) Polymerase PAP1 Polyadenylates Non-Coding RNAs and Is Essential for snoRNA Biogenesis in Trypanosoma brucei. J. Mol. Biol. 2017, 429, 3301–3318. [Google Scholar] [CrossRef]
- Chikne, V.; Shanmugha Rajan, K.; Shalev-Benami, M.; Decker, K.; Cohen-Chalamish, S.; Madmoni, H.; Biswas, V.K.; Kumar, G.S.; Doniger, T.; Unger, R.; et al. Small nucleolar RNAs controlling rRNA processing in Trypanosoma brucei. Nucleic Acids Res. 2019, 47, 2609–2629. [Google Scholar] [CrossRef]
- Hartshorne, T.; Toyofuku, W. Two 5’-ETS regions implicated in interactions with U3 snoRNA are required for small subunit rRNA maturation in Trypanosoma brucei. Nucleic Acids Res. 1999, 27, 3300–3309. [Google Scholar] [CrossRef]
- Hartshorne, T.; Toyofuku, W.; Hollenbaugh, J. Trypanosoma brucei 5’ETS A’-cleavage is directed by 3’-adjacent sequences, but not two U3 snoRNA-binding elements, which are all required for subsequent pre-small subunit rRNA processing events. J. Mol. Biol. 2001, 313, 733–749. [Google Scholar] [CrossRef]
- Gupta, S.K.; Hury, A.; Ziporen, Y.; Shi, H.; Ullu, E.; Michaeli, S. Small nucleolar RNA interference in Trypanosoma brucei: Mechanism and utilization for elucidating the function of snoRNAs. Nucleic Acids Res. 2010, 38, 7236–7247. [Google Scholar] [CrossRef]
- Michaeli, S.; Doniger, T.; Gupta, S.K.; Wurtzel, O.; Romano, M.; Visnovezky, D.; Sorek, R.; Unger, R.; Ullu, E. RNA-seq analysis of small RNPs in Trypanosoma brucei reveals a rich repertoire of non-coding RNAs. Nucleic Acids Res. 2012, 40, 1282–1298. [Google Scholar] [CrossRef] [PubMed]
- White, T.C.; Rudenko, G.; Borst, P. Three small RNAs within the 10 kb trypanosome rRNA transcription unit are analogous to domain VII of other eukaryotic 28S rRNAs. Nucleic Acids Res. 1986, 14, 9471–9489. [Google Scholar] [CrossRef] [PubMed]
- Campbell, D.A.; Kubo, K.; Clark, C.G.; Boothroyd, J.C. Precise identification of cleavage sites involved in the unusual processing of trypanosome ribosomal RNA. J. Mol. Biol. 1987, 196, 113–124. [Google Scholar] [CrossRef]
- Jensen, B.C.; Wang, Q.; Kifer, C.T.; Parsons, M. The NOG1 GTP-binding protein is required for biogenesis of the 60 S ribosomal subunit. J. Biol. Chem. 2003, 278, 32204–32211. [Google Scholar] [CrossRef]
- Jensen, B.C.; Brekken, D.L.; Randall, A.C.; Kifer, C.T.; Parsons, M. Species specificity in ribosome biogenesis: A nonconserved phosphoprotein is required for formation of the large ribosomal subunit in Trypanosoma brucei. Eukaryot. Cell 2005, 4, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Droll, D.; Archer, S.; Fenn, K.; Delhi, P.; Matthews, K.; Clayton, C. The trypanosome Pumilio-domain protein PUF7 associates with a nuclear cyclophilin and is involved in ribosomal RNA maturation. FEBS Lett. 2010, 584, 1156–1162. [Google Scholar] [CrossRef] [Green Version]
- Shalev-Benami, M.; Zhang, Y.; Matzov, D.; Halfon, Y.; Zackay, A.; Rozenberg, H.; Zimmerman, E.; Bashan, A.; Jaffe, C.L.; Yonath, A.; et al. 2.8-A Cryo-EM Structure of the Large Ribosomal Subunit from the Eukaryotic Parasite Leishmania. Cell Rep. 2016, 16, 288–294. [Google Scholar] [CrossRef]
- Schumann Burkard, G.; Kaser, S.; de Araujo, P.R.; Schimanski, B.; Naguleswaran, A.; Knusel, S.; Heller, M.; Roditi, I. Nucleolar proteins regulate stage-specific gene expression and ribosomal RNA maturation in Trypanosoma brucei. Mol. Microbiol. 2013, 88, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Sakyiama, J.; Zimmer, S.L.; Ciganda, M.; Williams, N.; Read, L.K. Ribosome biogenesis requires a highly diverged XRN family 5’->3’ exoribonuclease for rRNA processing in Trypanosoma brucei. RNA 2013, 19, 1419–1431. [Google Scholar] [CrossRef] [PubMed]
- Umaer, K.; Ciganda, M.; Williams, N. Ribosome biogenesis in african trypanosomes requires conserved and trypanosome-specific factors. Eukaryot. Cell 2014, 13, 727–737. [Google Scholar] [CrossRef]
- Kala, S.; Mehta, V.; Yip, C.W.; Moshiri, H.; Najafabadi, H.S.; Ma, R.; Nikpour, N.; Zimmer, S.L.; Salavati, R. The interaction of a Trypanosoma brucei KH-domain protein with a ribonuclease is implicated in ribosome processing. Mol. Biochem. Parasitol. 2017, 211, 94–103. [Google Scholar] [CrossRef]
- Faktorova, D.; Bar, A.; Hashimi, H.; McKenney, K.; Horak, A.; Schnaufer, A.; Rubio, M.A.T.; Alfonzo, J.D.; Lukes, J. TbUTP10, a protein involved in early stages of pre-18S rRNA processing in Trypanosoma brucei. Mol. Biochem. Parasitol. 2018, 225, 84–93. [Google Scholar] [CrossRef]
- Charette, M.; Gray, M.W. Pseudouridine in RNA: What, where, how, and why. IUBMB Life 2000, 49, 341–351. [Google Scholar] [PubMed]
- Caton, E.A.; Kelly, E.K.; Kamalampeta, R.; Kothe, U. Efficient RNA pseudouridylation by eukaryotic H/ACA ribonucleoproteins requires high affinity binding and correct positioning of guide RNA. Nucleic Acids Res. 2018, 46, 905–916. [Google Scholar] [CrossRef]
- Chikne, V.; Doniger, T.; Rajan, K.S.; Bartok, O.; Eliaz, D.; Cohen-Chalamish, S.; Tschudi, C.; Unger, R.; Hashem, Y.; Kadener, S.; et al. A pseudouridylation switch in rRNA is implicated in ribosome function during the life cycle of Trypanosoma brucei. Sci. Rep. 2016, 6, 25296. [Google Scholar] [CrossRef] [PubMed]
- Erales, J.; Marchand, V.; Panthu, B.; Gillot, S.; Belin, S.; Ghayad, S.E.; Garcia, M.; Laforets, F.; Marcel, V.; Baudin-Baillieu, A.; et al. Evidence for rRNA 2’-O-methylation plasticity: Control of intrinsic translational capabilities of human ribosomes. Proc. Natl. Acad. Sci. USA 2017, 114, 12934–12939. [Google Scholar] [CrossRef]
- Pertschy, B.; Schneider, C.; Gnadig, M.; Schafer, T.; Tollervey, D.; Hurt, E. RNA helicase Prp43 and its co-factor Pfa1 promote 20 to 18S rRNA processing catalyzed by the endonuclease Nob1. J. Biol. Chem. 2009, 284, 35079–35091. [Google Scholar] [CrossRef]
- Woolford, J.L., Jr.; Baserga, S.J. Ribosome biogenesis in the yeast Saccharomyces cerevisiae. Genetics 2013, 195, 643–681. [Google Scholar] [CrossRef]
- Klinge, S.; Woolford, J.L., Jr. Ribosome assembly coming into focus. Nat. Rev. Mol. Cell Biol. 2019, 20, 116–131. [Google Scholar]
- Nerurkar, P.; Altvater, M.; Gerhardy, S.; Schutz, S.; Fischer, U.; Weirich, C.; Panse, V.G. Eukaryotic Ribosome Assembly and Nuclear Export. Int. Rev. Cell Mol. Biol. 2015, 319, 107–140. [Google Scholar]
- Rabl, J.; Leibundgut, M.; Ataide, S.F.; Haag, A.; Ban, N. Crystal structure of the eukaryotic 40S ribosomal subunit in complex with initiation factor 1. Science 2011, 331, 730–736. [Google Scholar] [CrossRef]
- Klinge, S.; Voigts-Hoffmann, F.; Leibundgut, M.; Arpagaus, S.; Ban, N. Crystal structure of the eukaryotic 60S ribosomal subunit in complex with initiation factor 6. Science 2011, 334, 941–948. [Google Scholar] [CrossRef]
- Ben-Shem, A.; Garreau de Loubresse, N.; Melnikov, S.; Jenner, L.; Yusupova, G.; Yusupov, M. The structure of the eukaryotic ribosome at 3.0 A resolution. Science 2011, 334, 1524–1529. [Google Scholar] [CrossRef]
- Khatter, H.; Myasnikov, A.G.; Natchiar, S.K.; Klaholz, B.P. Structure of the human 80S ribosome. Nature 2015, 520, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Ayub, M.J.; Levin, M.J.; Frank, J. The structure of the 80S ribosome from Trypanosoma cruzi reveals unique rRNA components. Proc. Natl. Acad. Sci. USA 2005, 102, 10206–10211. [Google Scholar] [CrossRef]
- Zhang, X.; Lai, M.; Chang, W.; Yu, I.; Ding, K.; Mrazek, J.; Ng, H.L.; Yang, O.O.; Maslov, D.A.; Zhou, Z.H. Structures and stabilization of kinetoplastid-specific split rRNAs revealed by comparing leishmanial and human ribosomes. Nat. Commun. 2016, 7, 13223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prohaska, K.; Williams, N. Assembly of the Trypanosoma brucei 60S ribosomal subunit nuclear export complex requires trypanosome-specific proteins P34 and P37. Eukaryot. Cell 2009, 8, 77–87. [Google Scholar] [CrossRef]
- Umaer, K.; Williams, N. Kinetoplastid Specific RNA-Protein Interactions in Trypanosoma cruzi Ribosome Biogenesis. PLoS ONE 2015, 10, e0131323. [Google Scholar] [CrossRef] [PubMed]
- Kamina, A.D.; Jaremko, D.; Christen, L.; Williams, N. Essential Assembly Factor Rpf2 Forms Novel Interactions within the 5S RNP in Trypanosoma brucei. mSphere 2017, 2, e00394-17. [Google Scholar] [CrossRef]
- Atwood, J.A., III; Weatherly, D.B.; Minning, T.A.; Bundy, B.; Cavola, C.; Opperdoes, F.R.; Orlando, R.; Tarleton, R.L. The Trypanosoma cruzi proteome. Science 2005, 309, 473–476. [Google Scholar] [CrossRef]
- Ayub, M.J.; Atwood, J.; Nuccio, A.; Tarleton, R.; Levin, M.J. Proteomic analysis of the Trypanosoma cruzi ribosomal proteins. Biochem. Biophys. Res. Commun. 2009, 382, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Cerca, S.; Poll, G.; Kuhn, H.; Neueder, A.; Jakob, S.; Tschochner, H.; Milkereit, P. Analysis of the in vivo assembly pathway of eukaryotic 40S ribosomal proteins. Mol. Cell 2007, 28, 446–457. [Google Scholar] [CrossRef]
- Ahmad, Y.; Boisvert, F.M.; Gregor, P.; Cobley, A.; Lamond, A.I. NOPdb: Nucleolar Proteome Database—2008 update. Nucleic Acids Res. 2009, 37, D181–D184. [Google Scholar] [CrossRef]
- Coute, Y.; Burgess, J.A.; Diaz, J.J.; Chichester, C.; Lisacek, F.; Greco, A.; Sanchez, J.C. Deciphering the human nucleolar proteome. Mass Spectrom. Rev. 2006, 25, 215–234. [Google Scholar] [CrossRef]
- Tsai, R.Y.; Pederson, T. Connecting the nucleolus to the cell cycle and human disease. FASEB J. 2014, 28, 3290–3296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertrand, E.; Houser-Scott, F.; Kendall, A.; Singer, R.H.; Engelke, D.R. Nucleolar localization of early tRNA processing. Genes Dev. 1998, 12, 2463–2468. [Google Scholar] [CrossRef] [Green Version]
- Thompson, M.; Haeusler, R.A.; Good, P.D.; Engelke, D.R. Nucleolar clustering of dispersed tRNA genes. Science 2003, 302, 1399–1401. [Google Scholar] [CrossRef]
- Nuñez Villacis, L.; Wong, M.S.; Ferguson, L.L.; Hein, N.; George, A.J.; Hannan, K.M. New Roles for the Nucleolus in Health and Disease. Bioessays 2018, 40, e1700233. [Google Scholar] [CrossRef]
- Rubbi, C.P.; Milner, J. Disruption of the nucleolus mediates stabilization of p53 in response to DNA damage and other stresses. EMBO J. 2003, 22, 6068–6077. [Google Scholar] [CrossRef] [Green Version]
- Olson, M.O. Sensing cellular stress: Another new function for the nucleolus? Sci. STKE 2004, 2004, e10. [Google Scholar] [CrossRef] [PubMed]
- Pestov, D.G.; Strezoska, Z.; Lau, L.F. Evidence of p53-dependent cross-talk between ribosome biogenesis and the cell cycle: Effects of nucleolar protein Bop1 on G(1)/S transition. Mol. Cell Biol. 2001, 21, 4246–4255. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.; Grummt, I. Cellular stress and nucleolar function. Cell Cycle 2005, 4, 1036–1038. [Google Scholar] [CrossRef] [PubMed]
- Grummt, I. The nucleolus-guardian of cellular homeostasis and genome integrity. Chromosoma 2013, 122, 487–497. [Google Scholar] [CrossRef]
- James, A.; Wang, Y.; Raje, H.; Rosby, R.; DiMario, P. Nucleolar stress with and without p53. Nucleus 2014, 5, 402–426. [Google Scholar] [CrossRef] [Green Version]
- Elias, M.C.; Marques-Porto, R.; Freymuller, E.; Schenkman, S. Transcription rate modulation through the Trypanosoma cruzi life cycle occurs in parallel with changes in nuclear organisation. Mol. Biochem. Parasitol. 2001, 112, 79–90. [Google Scholar] [CrossRef]
- Gluenz, E.; Taylor, M.C.; Kelly, J.M. The Trypanosoma cruzi metacyclic-specific protein Met-III associates with the nucleolus and contains independent amino and carboxyl terminal targeting elements. Int. J. Parasitol. 2007, 37, 617–625. [Google Scholar] [CrossRef]
- Nazer, E.; Sanchez, D.O. Nucleolar accumulation of RNA binding proteins induced by Actinomycin D is functional in Trypanosoma cruzi and Leishmania mexicana but not in T. brucei. PLoS ONE 2011, 6, e24184. [Google Scholar]
- Nazer, E.; Verdun, R.E.; Sanchez, D.O. Nucleolar localization of RNA binding proteins induced by actinomycin D and heat shock in Trypanosoma cruzi. PLoS ONE 2011, 6, e19920. [Google Scholar] [CrossRef] [PubMed]
- Nazer, E.; Verdun, R.E.; Sanchez, D.O. Severe heat shock induces nucleolar accumulation of mRNAs in Trypanosoma cruzi. PLoS ONE 2012, 7, e43715. [Google Scholar] [CrossRef] [PubMed]
- Gunzl, A.; Bruderer, T.; Laufer, G.; Schimanski, B.; Tu, L.C.; Chung, H.M.; Lee, P.T.; Lee, M.G. RNA polymerase I transcribes procyclin genes and variant surface glycoprotein gene expression sites in Trypanosoma brucei. Eukaryot. Cell 2003, 2, 542–551. [Google Scholar] [CrossRef]
- Borst, P. Antigenic variation and allelic exclusion. Cell 2002, 109, 5–8. [Google Scholar] [CrossRef]
- Pays, E.; Vanhamme, L.; Perez-Morga, D. Antigenic variation in Trypanosoma brucei: Facts, challenges and mysteries. Curr. Opin. Microbiol. 2004, 7, 369–374. [Google Scholar] [CrossRef]
- Berriman, M.; Hall, N.; Sheader, K.; Bringaud, F.; Tiwari, B.; Isobe, T.; Bowman, S.; Corton, C.; Clark, L.; Cross, G.A.; et al. The architecture of variant surface glycoprotein gene expression sites in Trypanosoma brucei. Mol. Biochem. Parasitol. 2002, 122, 131–140. [Google Scholar] [CrossRef]
- Vanhamme, L.; Pays, E. Control of gene expression in trypanosomes. Microbiol. Rev. 1995, 59, 223–240. [Google Scholar]
- Hoek, M.; Zanders, T.; Cross, G.A. Trypanosoma brucei expression-site-associated-gene-8 protein interacts with a Pumilio family protein. Mol. Biochem. Parasitol. 2002, 120, 269–283. [Google Scholar] [CrossRef]
- Navarro, M.; Gull, K. A pol I transcriptional body associated with VSG mono-allelic expression in Trypanosoma brucei. Nature 2001, 414, 759–763. [Google Scholar] [CrossRef]
- Konig, E.; Delius, H.; Carrington, M.; Williams, R.O.; Roditi, I. Duplication and transcription of procyclin genes in Trypanosoma brucei. Nucleic Acids Res. 1989, 17, 8727–8739. [Google Scholar] [CrossRef]
- Landeira, D.; Navarro, M. Nuclear repositioning of the VSG promoter during developmental silencing in Trypanosoma brucei. J. Cell Biol. 2007, 176, 133–139. [Google Scholar] [CrossRef]
- Sherman, D.R.; Janz, L.; Hug, M.; Clayton, C. Anatomy of the parp gene promoter of Trypanosoma brucei. EMBO J. 1991, 10, 3379–3386. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.D.; Huang, J.; Van der Ploeg, L.H. The promoter for the procyclic acidic repetitive protein (PARP) genes of Trypanosoma brucei shares features with RNA polymerase I promoters. Mol. Cell Biol. 1992, 12, 2644–2652. [Google Scholar] [CrossRef]
- Kim, H.S.; Park, S.H.; Gunzl, A.; Cross, G.A. MCM-BP is required for repression of life-cycle specific genes transcribed by RNA polymerase I in the mammalian infectious form of Trypanosoma brucei. PLoS ONE 2013, 8, e57001. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, M.S.; Kushwaha, M.; Ersfeld, K.; Fullbrook, A.; Stanne, T.M.; Rudenko, G. NLP is a novel transcription regulator involved in VSG expression site control in Trypanosoma brucei. Nucleic Acids Res. 2011, 39, 2018–2031. [Google Scholar] [CrossRef]
- Stanne, T.M.; Kushwaha, M.; Wand, M.; Taylor, J.E.; Rudenko, G. TbISWI regulates multiple polymerase I (Pol I)-transcribed loci and is present at Pol II transcription boundaries in Trypanosoma brucei. Eukaryot. Cell 2011, 10, 964–976. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Calvillo, S.; Florencio-Martínez, L.E.; Nepomuceno-Mejía, T. Nucleolar Structure and Function in Trypanosomatid Protozoa. Cells 2019, 8, 421. https://doi.org/10.3390/cells8050421
Martínez-Calvillo S, Florencio-Martínez LE, Nepomuceno-Mejía T. Nucleolar Structure and Function in Trypanosomatid Protozoa. Cells. 2019; 8(5):421. https://doi.org/10.3390/cells8050421
Chicago/Turabian StyleMartínez-Calvillo, Santiago, Luis E. Florencio-Martínez, and Tomás Nepomuceno-Mejía. 2019. "Nucleolar Structure and Function in Trypanosomatid Protozoa" Cells 8, no. 5: 421. https://doi.org/10.3390/cells8050421
APA StyleMartínez-Calvillo, S., Florencio-Martínez, L. E., & Nepomuceno-Mejía, T. (2019). Nucleolar Structure and Function in Trypanosomatid Protozoa. Cells, 8(5), 421. https://doi.org/10.3390/cells8050421