NLRX1 Enhances Glutamate Uptake and Inhibits Glutamate Release by Astrocytes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Primary Mouse Astrocyte Cultures
2.3. Glutamate Uptake and Release Assay
2.4. Quantitative Real-Time PCR (qPCR)
2.5. Flow Cytometry Staining
2.6. Measurement of Intracellular ATP
2.7. Measurement of Mitochondrial DNA (mtDNA)
2.8. Statistical Analysis
3. Results
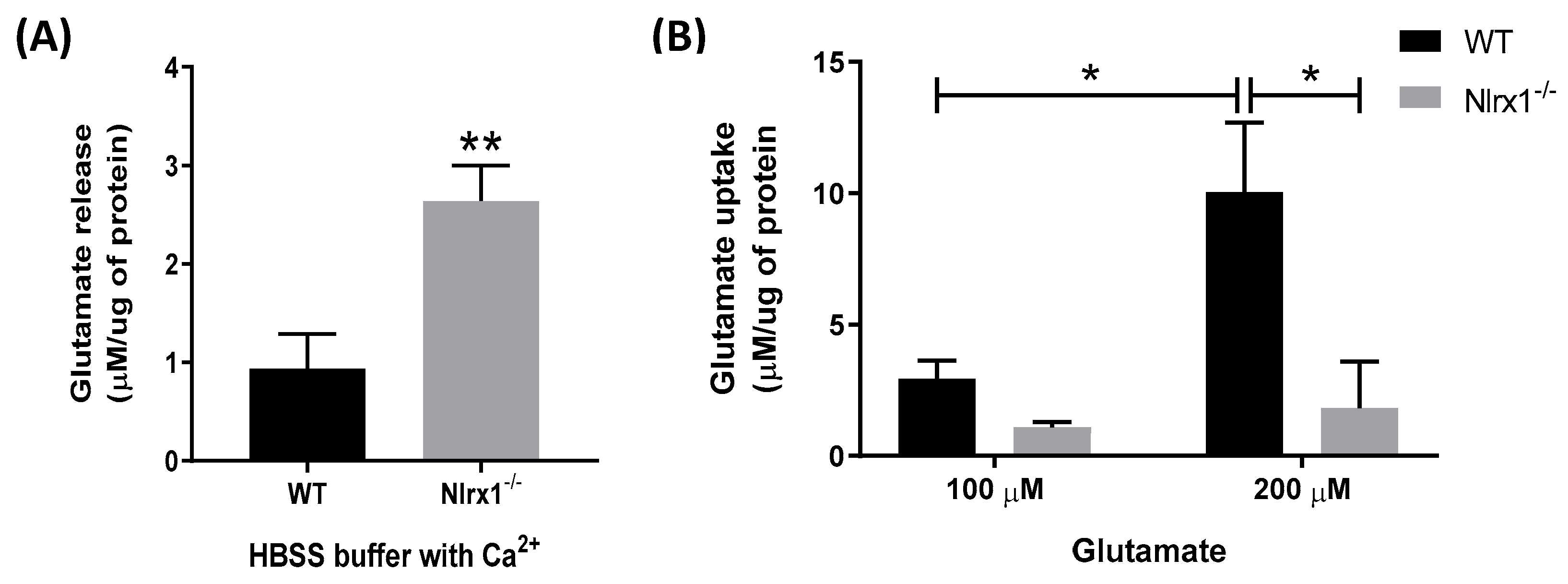
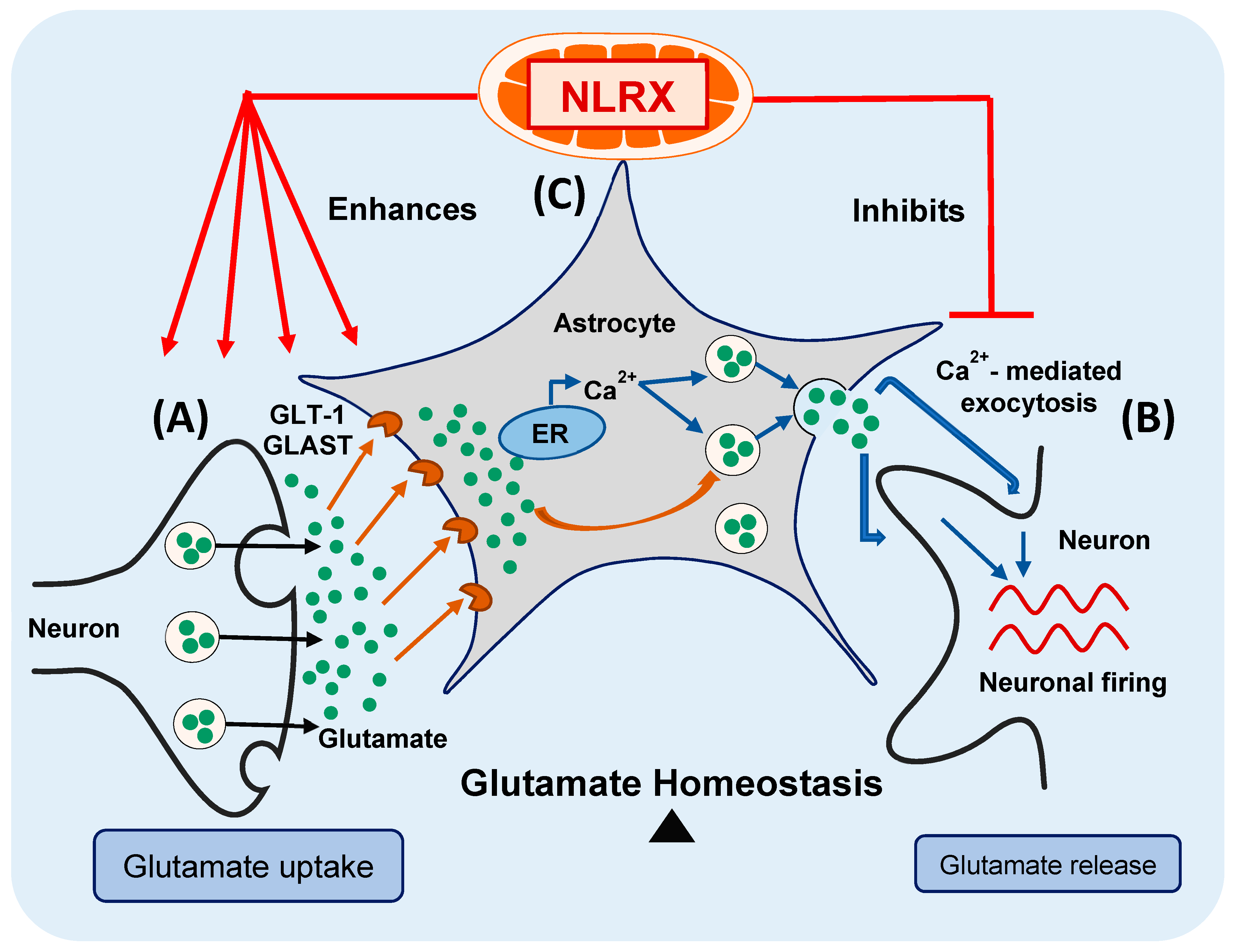
3.1. NLRX1 Inhibits Excess Glutamate Release and Enhances Glutamate Uptake by Astrocytes
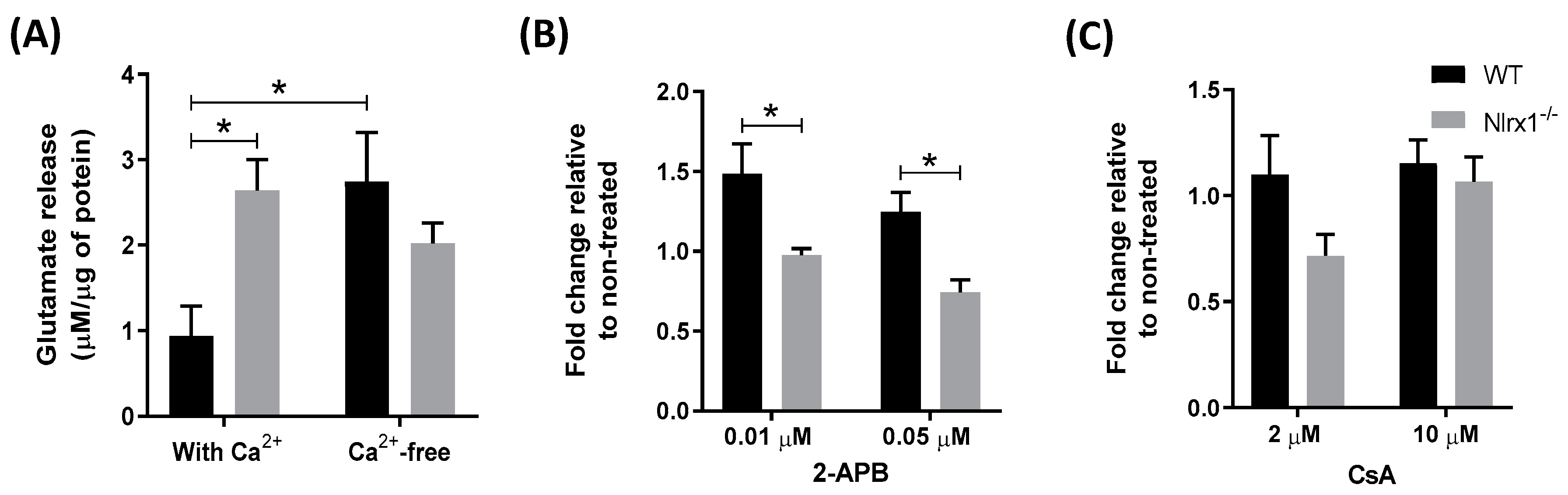
3.2. Nlrx1-/- Astrocytes’ Excess Glutamate Release Is Ca2+-Dependent
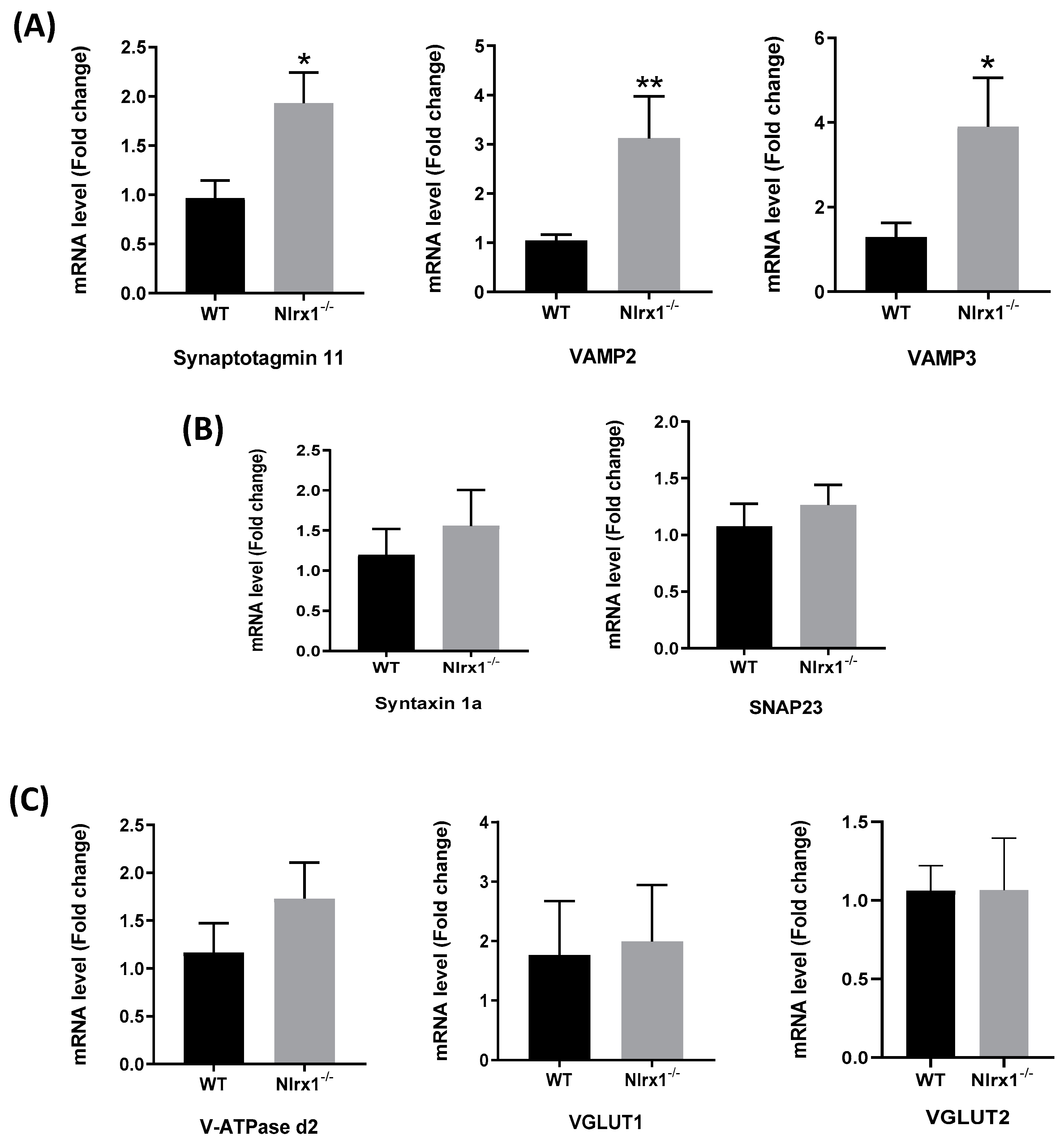
3.3. Glutamate Release by Nlrx1-/- Astrocytes Is Mediated By Exocytosis
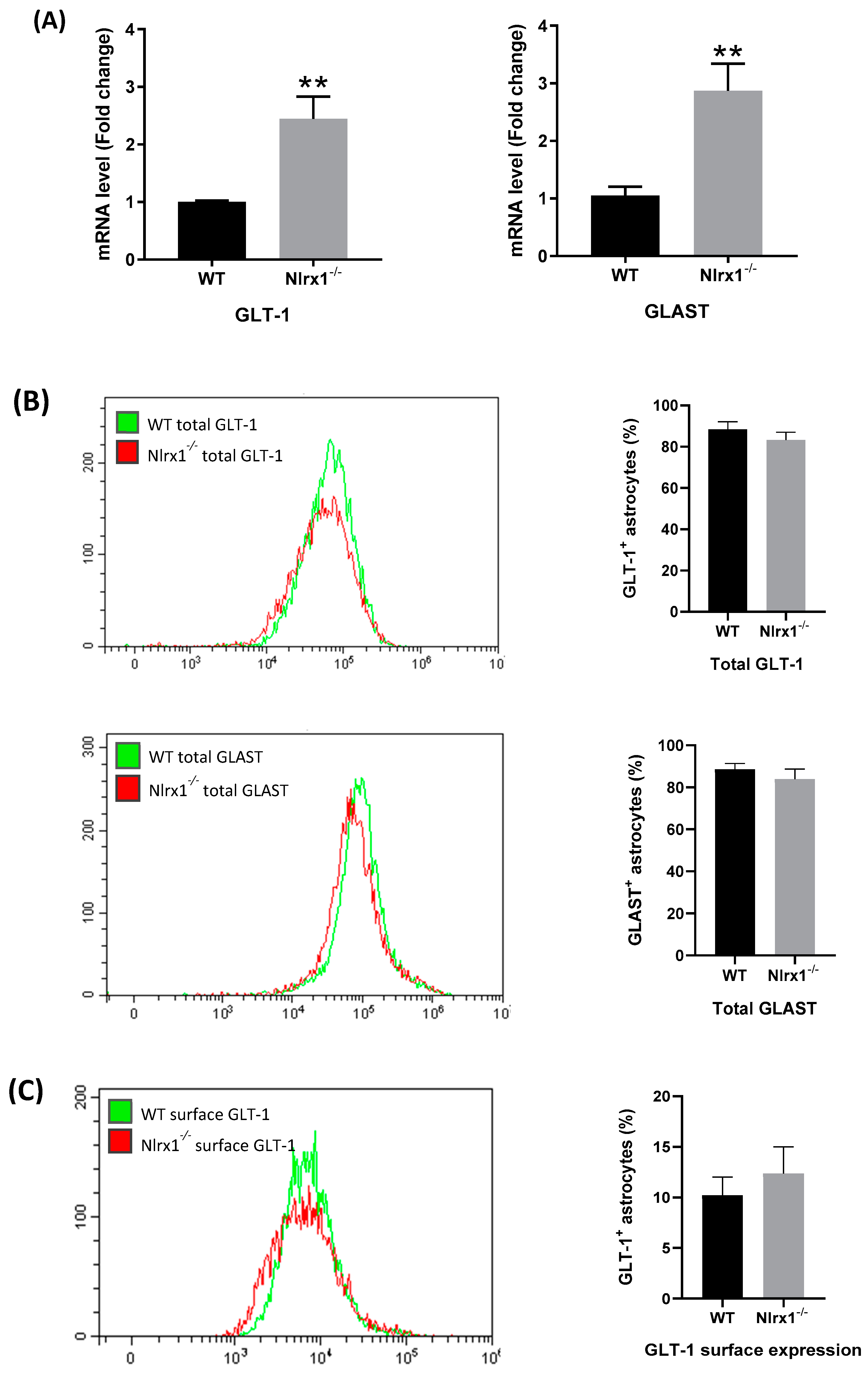
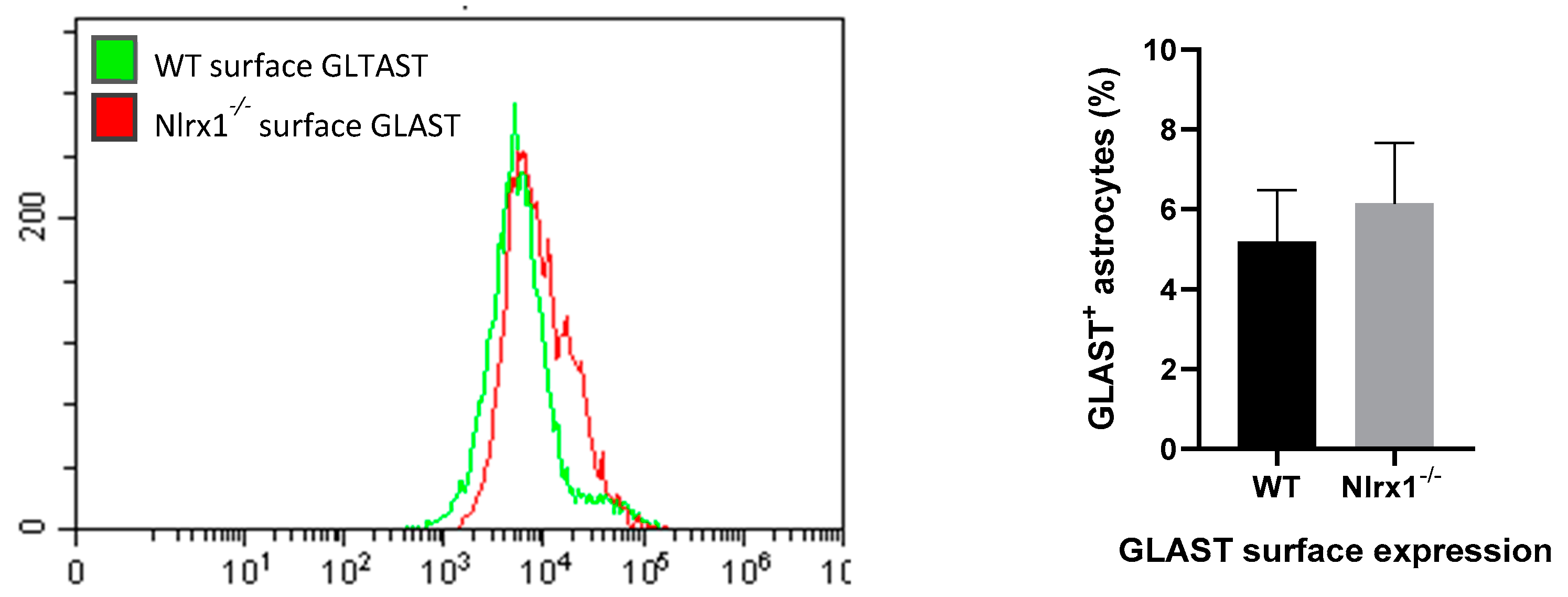
3.4. mRNA and Protein Expression of Glutamate Transporters in Astrocytes
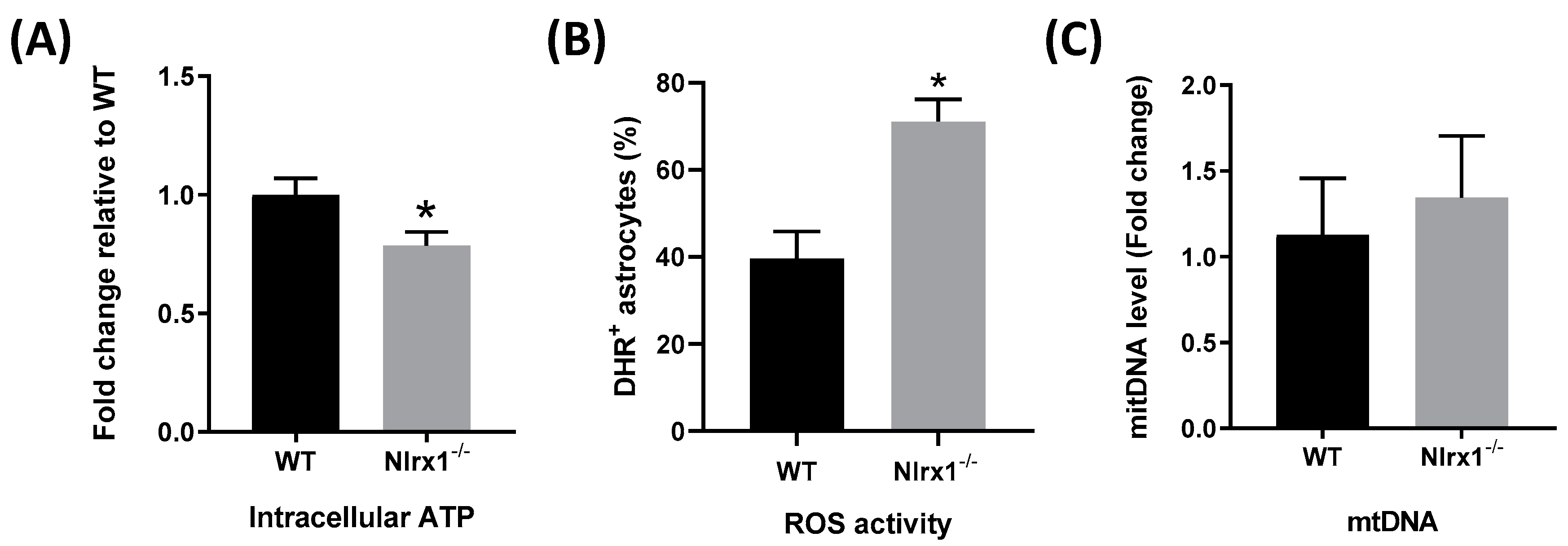
3.5. NLRX1 Enhances Mitochondrial Functions in Astrocytes
4. Discussion
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CNS | Central nervous system |
| EAAT1 and EAAT2 | Excitatory amino acid transporters 1 and 2 |
| GLAST | Glutamate–aspartate transporter |
| GLT-1 | Glutamate transporter-1 |
| ATP | Adenosine triphosphate |
| VGLUT1 and VGLUT2 | Vesicular glutamate transporters 1 and 2 |
| V-ATPase | Vacuolar (H+) ATPase |
| VAMP2 and VAMP3 | Vesicle-associated membrane protein 2 and 3 |
| ER | Endoplasmic reticulum |
| SNAP23 | Soluble N-ethylmaleimide-sensitive factor attachment protein 23 |
| HIV | Human immunodeficiency virus |
| MS | Multiple sclerosis |
| ALS | Amyotrophic lateral sclerosis |
| AD | Alzheimer’s disease |
| PD | Parkinson’s disease |
| NLRs | NOD-like receptors |
| NF-κB | Nuclear factor-κB |
| NLRX1 | NOD-like receptor X1 |
| IBD | Inflammatory bowel disease |
| EAE | Experimental autoimmune encephalomyelitis |
| WT | Wild-type |
| Nlrx1-/- | Nlrx1 knockout |
| dFBS | Deactivated fetal bovine serum |
| DMSO | Dimethyl sulfoxide |
| HBSS | Hank’s Balanced Salt Solution |
| qPCR | Quantitative real-time PCR |
| PBS | Phosphate-buffered saline |
| ROS | Reactive oxygen species |
| DHR | Dihydrorhodamine 123 |
| mtDNA | Mitochondrial DNA |
| SEM | Standard error of the mean |
| 2-APB | 2-Aminoethyl diphenylborinate |
| IP3 | Inositol-1,4,5-trisphosphate |
| CsA | Cyclosporin A |
| PKC | Protein kinase C |
| AA | Arachidonic acid |
| SH group | Sulfhydryl group |
| TCA | Tricarboxylic acid |
References
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [PubMed]
- Curtis, D.; Johnston, G. Amino acid transmitters in the mammalian central nervous system. In Ergebnisse der Physiologie Reviews of Physiology; Springer: Berlin/Heidelberg, Germany, 1974; Volume 69, pp. 97–188. [Google Scholar] [CrossRef]
- Fonnum, F. Glutamate: A neurotransmitter in mammalian brain. J. Neurochem. 1984, 42, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, S.; Gharagozloo, M.; Simard, C.; Gris, D.; Mahmoud, S.; Gharagozloo, M.; Simard, C.; Gris, D. Astrocytes Maintain Glutamate Homeostasis in the CNS by Controlling the Balance between Glutamate Uptake and Release. Cells 2019, 8, 184. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.M.; Swanson, R.A. Astrocyte glutamate transport: Review of properties, regulation, and physiological functions. Glia 2000, 32, 1–14. [Google Scholar] [CrossRef]
- Shashidharan, P.; Plaitakis, A. Cloning and characterization of a glutamate transporter cDNA from human cerebellum. Biochim. Biophys. Acta 1993, 1216, 161–164. [Google Scholar] [CrossRef]
- Shashidharan, P.; Wittenberg, I.; Plaitakis, A. Molecular cloning of human brain glutamate/aspartate transporter II. Biochim. Biophys. Acta-Biomembr. 1994, 1191, 393–396. [Google Scholar] [CrossRef]
- Storck, T.; Schulte, S.; Hofmann, K.; Stoffel, W. Structure, expression, and functional analysis of a Na(+)-dependent glutamate/aspartate transporter from rat brain. Proc. Natl. Acad. Sci. USA 1992, 89, 10955–10959. [Google Scholar] [CrossRef]
- Pines, G.; Danbolt, N.C.; Bjørås, M.; Zhang, Y.; Bendahan, A.; Eide, L.; Koepsell, H.; Storm-Mathisen, J.; Seeberg, E.; Kanner, B.I. Cloning and expression of a rat brain l-glutamate transporter. Nature 1992, 360, 464–467. [Google Scholar] [CrossRef]
- Kojima, S.; Nakamura, T.; Nidaira, T.; Nakamura, K.; Ooashi, N.; Ito, E.; Watase, K.; Tanaka, K.; Wada, K.; Kudo, Y.; et al. Optical detection of synaptically induced glutamate transport in hippocampal slices. J. Neurosci. 1999, 19, 2580–2588. [Google Scholar] [CrossRef] [PubMed]
- Bergles, D.E.; Jahr, C.E. Glial contribution to glutamate uptake at Schaffer collateral-commissural synapses in the hippocampus. J. Neurosci. 1998, 18, 7709–7716. [Google Scholar] [CrossRef]
- Sibson, N.R.; Dhankhar, A.; Mason, G.F.; Rothman, D.L.; Behar, K.L.; Shulman, R.G. Stoichiometric coupling of brain glucose metabolism and glutamatergic neuronal activity. Proc. Natl. Acad. Sci. USA 1998, 95, 316–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellerin, L.; Magistretti, P.J. Glutamate uptake stimulates Na+,K+-ATPase activity in astrocytes via activation of a distinct subunit highly sensitive to ouabain. J. Neurochem. 1997, 69, 2132–2137. [Google Scholar] [CrossRef] [PubMed]
- Gegelashvili, G.; Schousboe, A. High affinity glutamate transporters: Regulation of expression and activity. Mol. Pharmacol. 1997, 52, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Chi-Castañeda, D.; Suárez-Pozos, E.; Ortega, A. Regulation of Glutamate Transporter Expression in Glial Cells. Adv. Neurobiol. 2017, 16, 199–224. [Google Scholar]
- Sattler, R.; Rothstein, J.D. Regulation and dysregulation of glutamate transporters. Handb. Exp. Pharmacol. 2006, 277–303. [Google Scholar]
- Hamilton, N.B.; Attwell, D. Do astrocytes really exocytose neurotransmitters? Nat. Rev. Neurosci. 2010, 11, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Parpura, V.; Haydon, P.G. Physiological astrocytic calcium levels stimulate glutamate release to modulate adjacent neurons. Proc. Natl. Acad. Sci. USA 2000, 97, 8629–8634. [Google Scholar] [CrossRef] [Green Version]
- Malarkey, E.B.; Parpura, V. Mechanisms of glutamate release from astrocytes. Neurochem. Int. 2008, 52, 142–154. [Google Scholar] [CrossRef] [Green Version]
- Bezzi, P.; Gundersen, V.; Galbete, J.L.; Seifert, G.; Steinhäuser, C.; Pilati, E.; Volterra, A. Astrocytes contain a vesicular compartment that is competent for regulated exocytosis of glutamate. Nat. Neurosci. 2004, 7, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, L.; Zhou, Y.; Zheng, L.-H.; Zhou, Z. “Kiss-and-Run” Glutamate Secretion in Cultured and Freshly Isolated Rat Hippocampal Astrocytes. J. Neurosci. 2005, 25, 9236–9243. [Google Scholar] [CrossRef]
- Araque, A.; Li, N.; Doyle, R.T.; Haydon, P.G. SNARE protein-dependent glutamate release from astrocytes. J. Neurosci. 2000, 20, 666–673. [Google Scholar] [CrossRef]
- Parpura, V.; Zorec, R. Gliotransmission: Exocytotic release from astrocytes. Brain Res. Rev. 2010, 63, 83–92. [Google Scholar] [CrossRef]
- Pasti, L.; Zonta, M.; Pozzan, T.; Vicini, S.; Carmignoto, G. Cytosolic calcium oscillations in astrocytes may regulate exocytotic release of glutamate. J. Neurosci. 2001, 21, 477–484. [Google Scholar] [CrossRef]
- Zhang, Q.; Fukuda, M.; Van Bockstaele, E.; Pascual, O.; Haydon, P.G. Synaptotagmin IV regulates glial glutamate release. Proc. Natl. Acad. Sci. USA 2004, 101, 9441–9446. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-T.; Lu, J.-C.; Bai, J.; Chang, P.Y.; Martin, T.F.J.; Chapman, E.R.; Jackson, M.B. Different domains of synaptotagmin control the choice between kiss-and-run and full fusion. Nature 2003, 424, 943–947. [Google Scholar] [CrossRef]
- Sugita, S.; Han, W.; Butz, S.; Liu, X.; Fernández-Chacón, R.; Lao, Y.; Südhof, T.C. Synaptotagmin VII as a plasma membrane Ca(2+) sensor in exocytosis. Neuron 2001, 30, 459–473. [Google Scholar] [CrossRef]
- Montana, V.; Ni, Y.; Sunjara, V.; Hua, X.; Parpura, V. Vesicular Glutamate Transporter-Dependent Glutamate Release from Astrocytes. J. Neurosci. 2004, 24, 2633–2642. [Google Scholar] [CrossRef] [Green Version]
- Parpura, V.; Liu, F.; Brethorst, S.; Jeftinija, K.; Jeftinija, S.; Haydon, P.G. Alpha-latrotoxin stimulates glutamate release from cortical astrocytes in cell culture. FEBS Lett. 1995, 360, 266–270. [Google Scholar]
- Hepp, R.; Perraut, M.; Chasserot-Golaz, S.; Galli, T.; Aunis, D.; Langley, K.; Grant, N.J. Cultured glial cells express the SNAP-25 analogue SNAP-23. Glia 1999, 27, 181–187. [Google Scholar] [CrossRef]
- Jahn, R.; Scheller, R.H. SNAREs—Engines for membrane fusion. Nat. Rev. Mol. Cell Biol. 2006, 7, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Van Landeghem, F.K.H.; Weiss, T.; Oehmichen, M.; Deimling, A. Von Decreased Expression of Glutamate Transporters in Astrocytes after Human Traumatic Brain Injury. J. Neurotrauma 2006, 23, 1518–1528. [Google Scholar] [CrossRef]
- Vesce, S.; Bezzi, P.; Rossi, D.; Meldolesi, J.; Volterra, A. HIV-1 gp120 glycoprotein affects the astrocyte control of extracellular glutamate by both inhibiting the uptake and stimulating the release of the amino acid. FEBS Lett. 1997, 411, 107–109. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, A.L.; Robinson, M.B. The role of glutamate transporters in neurodegenerative diseases and potential opportunities for intervention. Neurochem. Int. 2007, 51, 333–355. [Google Scholar] [CrossRef] [Green Version]
- Pitt, D.; Werner, P.; Raine, C.S. Glutamate excitotoxicity in a model of multiple sclerosis. Nat. Med. 2000, 6, 67–70. [Google Scholar] [CrossRef]
- Bezzi, P.; Domercq, M.; Brambilla, L.; Galli, R.; Schols, D.; De Clercq, E.; Vescovi, A.; Bagetta, G.; Kollias, G.; Meldolesi, J.; et al. CXCR4-activated astrocyte glutamate release via TNFα: Amplification by microglia triggers neurotoxicity. Nat. Neurosci. 2001, 4, 702–710. [Google Scholar] [CrossRef]
- Bezzi, P.; Carmignoto, G.; Pasti, L.; Vesce, S.; Rossi, D.; Rizzini, B.L.; Pozzan, T.; Volterra, A. Prostaglandins stimulate calcium-dependent glutamate release in astrocytes. Nature 1998, 391, 281–285. [Google Scholar] [CrossRef]
- Wang, Y.; Qin, Z. Molecular and cellular mechanisms of excitotoxic neuronal death. Apoptosis 2010, 15, 1382–1402. [Google Scholar] [CrossRef]
- Gharagozloo, M.; Gris, K.V.; Mahvelati, T.; Amrani, A.; Lukens, J.R.; Gris, D. NLR-Dependent Regulation of Inflammation in Multiple Sclerosis. Front. Immunol. 2018, 8, 2012. [Google Scholar] [CrossRef]
- Moore, C.B.; Bergstralh, D.T.; Duncan, J.A.; Lei, Y.; Morrison, T.E.; Zimmermann, A.G.; Accavitti-Loper, M.A.; Madden, V.J.; Sun, L.; Ye, Z.; et al. NLRX1 is a regulator of mitochondrial antiviral immunity. Nature 2008, 451, 573–577. [Google Scholar] [CrossRef] [Green Version]
- Allen, I.C.; Moore, C.B.; Schneider, M.; Lei, Y.; Davis, B.K.; Scull, M.A.; Gris, D.; Roney, K.E.; Zimmermann, A.G.; Bowzard, J.B.; et al. NLRX1 protein attenuates inflammatory responses to infection by interfering with the RIG-I-MAVS and TRAF6-NF-κB signaling pathways. Immunity 2011, 34, 854–865. [Google Scholar] [CrossRef]
- Huang, J.-H.; Liu, C.-Y.; Wu, S.-Y.; Chen, W.-Y.; Chang, T.-H.; Kan, H.-W.; Hsieh, S.-T.; Ting, J.P.-Y.; Wu-Hsieh, B.A. NLRX1 Facilitates Histoplasma capsulatum-Induced LC3-Associated Phagocytosis for Cytokine Production in Macrophages. Front. Immunol. 2018, 9, 2761. [Google Scholar] [CrossRef]
- Aikawa, C.; Nakajima, S.; Karimine, M.; Nozawa, T.; Minowa-Nozawa, A.; Toh, H.; Yamada, S.; Nakagawa, I. NLRX1 Negatively Regulates Group A Streptococcus Invasion and Autophagy Induction by Interacting With the Beclin 1–UVRAG Complex. Front. Cell. Infect. Microbiol. 2018, 8, 403. [Google Scholar] [CrossRef]
- Ma, D.; Zhao, Y.; She, J.; Zhu, Y.; Zhao, Y.; Liu, L.; Zhang, Y. NLRX1 alleviates lipopolysaccharide-induced apoptosis and inflammation in chondrocytes by suppressing the activation of NF-κB signaling. Int. Immunopharmacol. 2019, 71, 7–13. [Google Scholar] [CrossRef]
- Tattoli, I.; Killackey, S.A.; Foerster, E.G.; Molinaro, R.; Maisonneuve, C.; Rahman, M.A.; Winer, S.; Winer, D.A.; Streutker, C.J.; Philpott, D.J.; et al. NLRX1 Acts as an Epithelial-Intrinsic Tumor Suppressor through the Modulation of TNF-Mediated Proliferation. Cell Rep. 2016, 14, 2576–2586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leber, A.; Hontecillas, R.; Tubau-Juni, N.; Zoccoli-Rodriguez, V.; Abedi, V.; Bassaganya-Riera, J. NLRX1 Modulates Immunometabolic Mechanisms Controlling the Host-Gut Microbiota Interactions during Inflammatory Bowel Disease. Front. Immunol. 2018, 9, 363. [Google Scholar] [CrossRef] [PubMed]
- Theus, M.H.; Brickler, T.; Meza, A.L.; Coutermarsh-Ott, S.; Hazy, A.; Gris, D.; Allen, I.C. Loss of NLRX1 Exacerbates Neural Tissue Damage and NF-κB Signaling following Brain Injury. J. Immunol. 2017, 199, 3547–3558. [Google Scholar] [CrossRef]
- Eitas, T.K.; Chou, W.-C.; Wen, H.; Gris, D.; Robbins, G.R.; Brickey, J.; Oyama, Y.; Ting, J.P.-Y. The Nucleotide-binding Leucine-rich Repeat (NLR) Family Member NLRX1 Mediates Protection against Experimental Autoimmune Encephalomyelitis and Represses Macrophage/Microglia-induced Inflammation. J. Biol. Chem. 2014, 289, 4173–4179. [Google Scholar] [CrossRef]
- Imbeault, E.; Mahvelati, T.M.; Braun, R.; Gris, P.; Gris, D. Nlrx1 regulates neuronal cell death. Mol. Brain 2014, 7, 90. [Google Scholar] [CrossRef]
- Stokman, G.; Kors, L.; Bakker, P.J.; Rampanelli, E.; Claessen, N.; Teske, G.J.D.; Butter, L.; van Andel, H.; van den Bergh Weerman, M.A.; Larsen, P.W.B.; et al. NLRX1 dampens oxidative stress and apoptosis in tissue injury via control of mitochondrial activity. J. Exp. Med. 2017, 214, 2405–2420. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Roy, M.; Prajapati, P.; Lipatova, A.; Sripada, L.; Gohel, D.; Singh, A.; Mane, M.; Godbole, M.M.; Chumakov, P.M.; et al. NLRX1 regulates TNF-α-induced mitochondria-lysosomal crosstalk to maintain the invasive and metastatic potential of breast cancer cells. Biochim. Biophys. Acta-Mol. Basis Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Killackey, S.A.; Rahman, M.A.; Soares, F.; Zhang, A.B.; Abdel-Nour, M.; Philpott, D.J.; Girardin, S.E. The mitochondrial Nod-like receptor NLRX1 modifies apoptosis through SARM1. Mol. Cell. Biochem. 2019, 453, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Soares, F.; Tattoli, I.; Rahman, M.A.; Robertson, S.J.; Belcheva, A.; Liu, D.; Streutker, C.; Winer, S.; Winer, D.A.; Martin, A.; et al. The mitochondrial protein NLRX1 controls the balance between extrinsic and intrinsic apoptosis. J. Biol. Chem. 2014, 289, 19317–19330. [Google Scholar] [CrossRef] [PubMed]
- Jaworska, J.; Coulombe, F.; Downey, J.; Tzelepis, F.; Shalaby, K.; Tattoli, I.; Berube, J.; Rousseau, S.; Martin, J.G.; Girardin, S.E.; et al. NLRX1 prevents mitochondrial induced apoptosis and enhances macrophage antiviral immunity by interacting with influenza virus PB1-F2 protein. Proc. Natl. Acad. Sci. USA 2014, 111, E2110–E2119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gharagozloo, M.; Mahvelati, T.M.; Imbeault, E.; Gris, P.; Zerif, E.; Bobbala, D.; Ilangumaran, S.; Amrani, A.; Gris, D. The nod-like receptor, Nlrp12, plays an anti-inflammatory role in experimental autoimmune encephalomyelitis. J. Neuroinflammation 2015, 12, 198. [Google Scholar] [CrossRef] [PubMed]
- Piao, C.; Ranaivo, H.R.; Rusie, A.; Wadhwani, N.; Koh, S.; Wainwright, M.S. Thrombin decreases expression of the glutamate transporter GLAST and inhibits glutamate uptake in primary cortical astrocytes via the Rho kinase pathway. Exp. Neurol. 2015, 273, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, J.M.; Smith, S.H.; Bilbo, S.D. FACS analysis of neuronal-glial interactions in the nucleus accumbens following morphine administration. Psychopharmacology 2013, 230, 525–535. [Google Scholar] [CrossRef]
- Gharagozloo, M.; Mahmoud, S.; Simard, C.; Mahvelati, T.M.; Amrani, A.; Gris, D. The Dual Immunoregulatory function of Nlrp12 in T Cell-Mediated Immune Response: Lessons from Experimental Autoimmune Encephalomyelitis. Cells 2018, 7, 119. [Google Scholar] [CrossRef]
- Gris, D.; Hamilton, E.F.; Weaver, L.C. The systemic inflammatory response after spinal cord injury damages lungs and kidneys. Exp. Neurol. 2008, 211, 259–270. [Google Scholar] [CrossRef]
- Farrell, S.M.J.; Groeger, G.; Bhatt, L.; Finnegan, S.; O’Brien, C.J.; Cotter, T.G. bFGF-mediated redox activation of the PI3K/Akt pathway in retinal photoreceptor cells. Eur. J. Neurosci. 2011, 33, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Marcaida, G.; Miñana, M.D.; Grisolía, S.; Felipo, V. Determination of intracellular ATP in primary cultures of neurons. Brain Res. Brain Res. Protoc. 1997, 1, 75–78. [Google Scholar] [CrossRef]
- Fuke, S.; Kubota-Sakashita, M.; Kasahara, T.; Shigeyoshi, Y.; Kato, T. Regional variation in mitochondrial DNA copy number in mouse brain. Biochim. Biophys. Acta-Bioenerg. 2011, 1807, 270–274. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, T.; Kanaji, T.; Nakade, S.; Kanno, T.; Mikoshiba, K. 2APB, 2-aminoethoxydiphenyl borate, a membrane-penetrable modulator of Ins(1,4,5)P3-induced Ca2+ release. J. Biochem. 1997, 122, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Reyes, R.C.; Parpura, V. Mitochondria modulate Ca2+-dependent glutamate release from rat cortical astrocytes. J. Neurosci. 2008, 28, 9682–9691. [Google Scholar] [CrossRef]
- Volterra, A.; Trotti, D.; Floridi, S.; Racagni, G. Reactive oxygen species inhibit high-affinity glutamate uptake: Molecular mechanism and neuropathological implications. Ann. N. Y. Acad. Sci. 1994, 738, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Sorg, O.; Horn, T.F.; Yu, N.; Gruol, D.L.; Bloom, F.E. Inhibition of astrocyte glutamate uptake by reactive oxygen species: Role of antioxidant enzymes. Mol. Med. 1997, 3, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Lehre, K.P.; Danbolt, N.C. The number of glutamate transporter subtype molecules at glutamatergic synapses: Chemical and stereological quantification in young adult rat brain. J. Neurosci. 1998, 18, 8751–8757. [Google Scholar] [CrossRef] [PubMed]
- Eulenburg, V.; Gomeza, J. Neurotransmitter transporters expressed in glial cells as regulators of synapse function. Brain Res. Rev. 2010, 63, 103–112. [Google Scholar] [CrossRef]
- Conradt, M.; Stoffel, W. Inhibition of the high-affinity brain glutamate transporter GLAST-1 via direct phosphorylation. J. Neurochem. 1997, 68, 1244–1251. [Google Scholar] [CrossRef]
- Zerangue, N.; Arriza, J.L.; Amara, S.G.; Kavanaugh, M.P. Differential modulation of human glutamate transporter subtypes by arachidonic acid. J. Biol. Chem. 1995, 270, 6433–6435. [Google Scholar] [CrossRef]
- Trotti, D.; Rizzini, B.L.; Rossi, D.; Haugeto, O.; Racagni, G.; Danbolt, N.C.; Volterra, A. Neuronal and glial glutamate transporters possess an SH-based redox regulatory mechanism. Eur. J. Neurosci. 1997, 9, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Grewer, C.; Gameiro, A.; Zhang, Z.; Tao, Z.; Braams, S.; Rauen, T. Glutamate forward and reverse transport: From molecular mechanism to transporter-mediated release after ischemia. IUBMB Life 2008, 60, 609–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, D.J.; Oshima, T.; Attwell, D. Glutamate release in severe brain ischaemia is mainly by reversed uptake. Nature 2000, 403, 316–321. [Google Scholar] [CrossRef]
- McKenna, M.C. Glutamate Pays Its Own Way in Astrocytes. Front. Endocrinol. 2013, 4, 191. [Google Scholar] [CrossRef]
- Kostic, M.; Zivkovic, N.; Cvetanovic, A.; Stojanovic, I.; Colic, M. IL-17 signalling in astrocytes promotes glutamate excitotoxicity: Indications for the link between inflammatory and neurodegenerative events in multiple sclerosis. Mult. Scler. Relat. Disord. 2017, 11, 12–17. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 18S | F: 5′ CGG CTA CCA CAT CCA AGG AA ′3 R: 5′ GCT GGA ATT ACC GCG GCT ′3 |
| Exocytosis | |
| V-ATPase d2 | F: 5′ TTC AGT TGC TAT CCA GGA CTC GGA ′3 R: 5′ GCA TGT CAT GTA GGT GAG AAA TGT GCT CA ′3 |
| VGLUT1 | F: 5′ GGT GGA GGG GGT CAC ATA C ′3 R: 5′ AGA TCC CGA AGC TGC CAT AGA ′3 |
| VGLUT2 | F: 5′ CCC TGG AGG TGC CTG AGA A ′3 R: 5′ GCG GTG GAT AGT GCT GTT GTT ′3 |
| Synaptotagmin 11 | F: 5′ GAC ACT TGC CGA AGA TGG ATA TC ′3 R: 5′ TGC GTT TTC TGC CGT AGT AGA ′3 |
| VAMP2 | F: 5′ CAC AAT CTG GTT CTT TGA GGA G ′3 R: 5′ AGA GAC TTC AGG CAG GAA TTA G ′3 |
| VAMP3 | F: 5′ CTC ACC AAG GCA TCA GTC TG ′3 R: 5′ ATT CTA AGA GCA CCA GGC ATC ′3 |
| Syntaxin 1a | F: 5′ TCC AAG CTA AAG AGC ATT GAG C ′3 R: 5′ GGC GTT GTA CTC GGA CAT GA ′3 |
| SNAP23 | F: 5′ AAT CCT GGG TTT AGC CAT TGA GTC ′3 R: 5′ TTG GTC CAT GCC TTC TTC TAT GC ′3 |
| Glutamate transporters | |
| GLT-1 | F: 5′ CGA TGA GCC AAA GCA CCG AA ′3 R: 5′ CTG GAG ATG ATA AGA GGG AGG ATG ′3 |
| GLAST | F: 5′ TCA AGT TCT GCC ACC CTA CC ′3 R: 5′ TCT GTC CAA AGT TCA GGT CAA ′3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmoud, S.; Gharagozloo, M.; Simard, C.; Amrani, A.; Gris, D. NLRX1 Enhances Glutamate Uptake and Inhibits Glutamate Release by Astrocytes. Cells 2019, 8, 400. https://doi.org/10.3390/cells8050400
Mahmoud S, Gharagozloo M, Simard C, Amrani A, Gris D. NLRX1 Enhances Glutamate Uptake and Inhibits Glutamate Release by Astrocytes. Cells. 2019; 8(5):400. https://doi.org/10.3390/cells8050400
Chicago/Turabian StyleMahmoud, Shaimaa, Marjan Gharagozloo, Camille Simard, Abdelaziz Amrani, and Denis Gris. 2019. "NLRX1 Enhances Glutamate Uptake and Inhibits Glutamate Release by Astrocytes" Cells 8, no. 5: 400. https://doi.org/10.3390/cells8050400
APA StyleMahmoud, S., Gharagozloo, M., Simard, C., Amrani, A., & Gris, D. (2019). NLRX1 Enhances Glutamate Uptake and Inhibits Glutamate Release by Astrocytes. Cells, 8(5), 400. https://doi.org/10.3390/cells8050400