In Silico Genome-Wide Analysis of Respiratory Burst Oxidase Homolog (RBOH) Family Genes in Five Fruit-Producing Trees, and Potential Functional Analysis on Lignification of Stone Cells in Chinese White Pear

Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Identification of Respiratory Burst Oxidase Homologs (RBOHs) in Five Fruit-Producing Trees
2.2. Conserved Motif, Gene Structure and Evolution Analysis
2.3. RBOH Family Genes’ Physical Localization and Gene Duplications
2.4. Microsynteny and Cis-Acting Elements Analysis of RBOH Family Genes
2.5. Plant Materials and Treatments
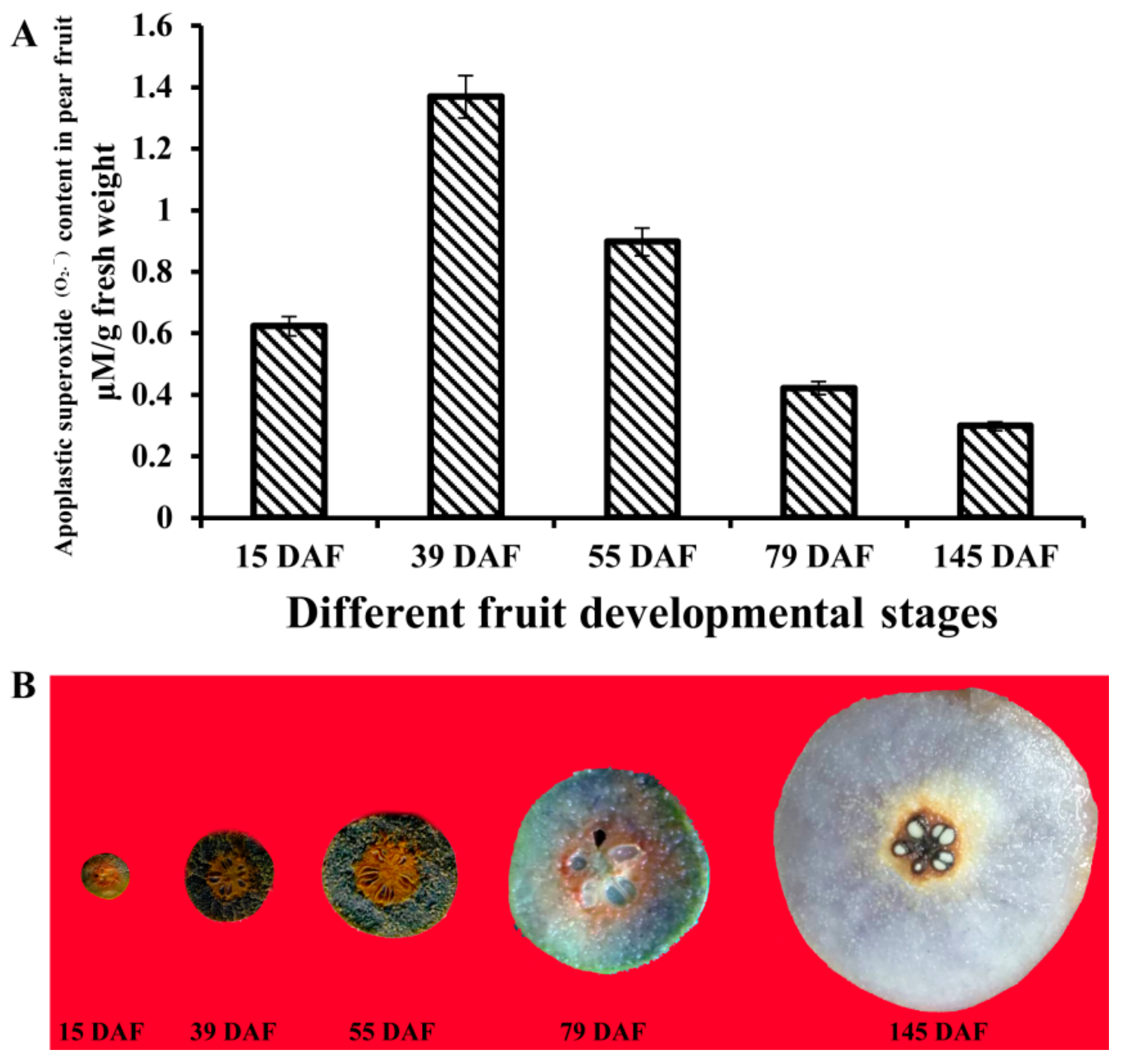
2.6. Determination of Apoplastic Superoxide (O2.−) Content in Pear Fruit
2.7. RNA Extraction and Reverse Transcription-Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR) Analysis
2.8. Subcellular Localization of PbRBOHs
3. Results
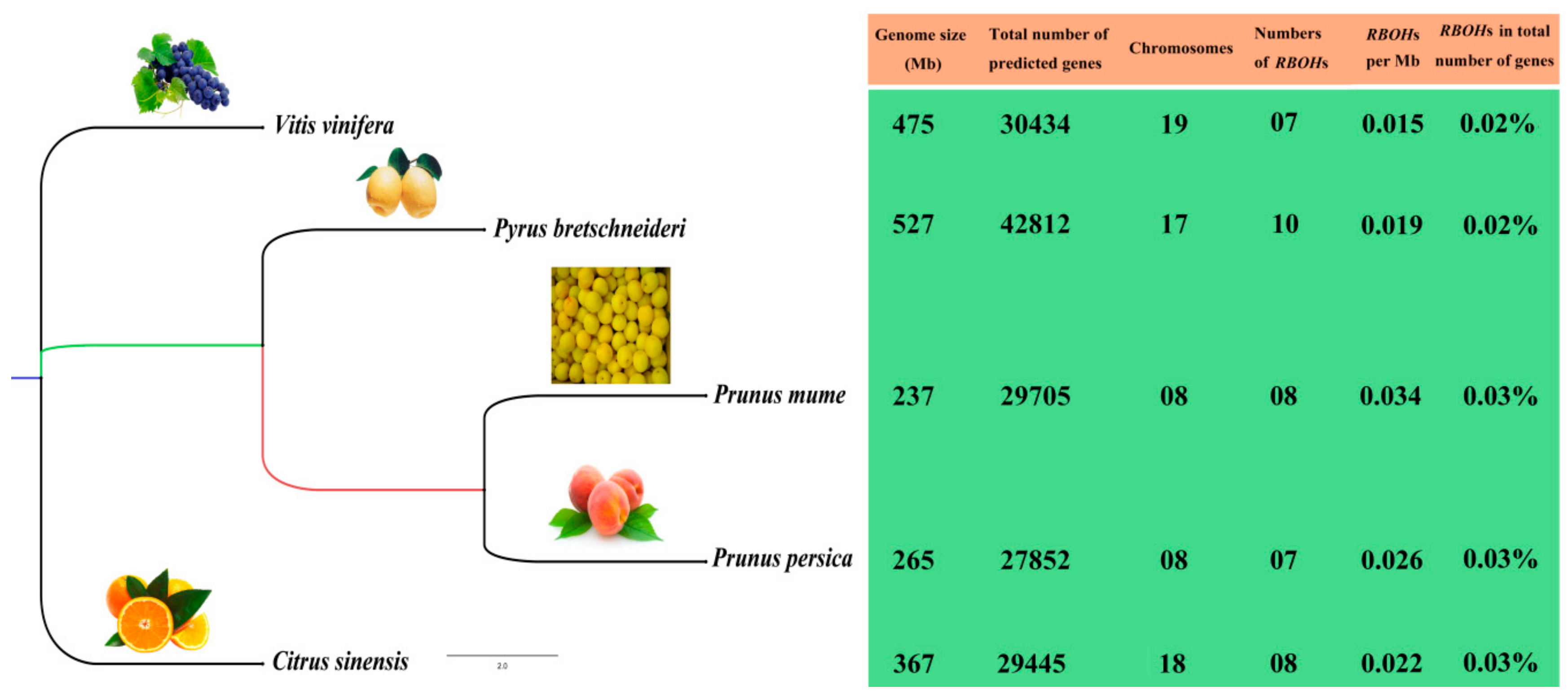
3.1. Identification, Characterization and Genomic Distribution of RBOH Family Genes in Five Fruit-Producing Trees
3.2. Chromosome Distribution and Duplication Events of RBOHs in Five Fruit-Producing Trees
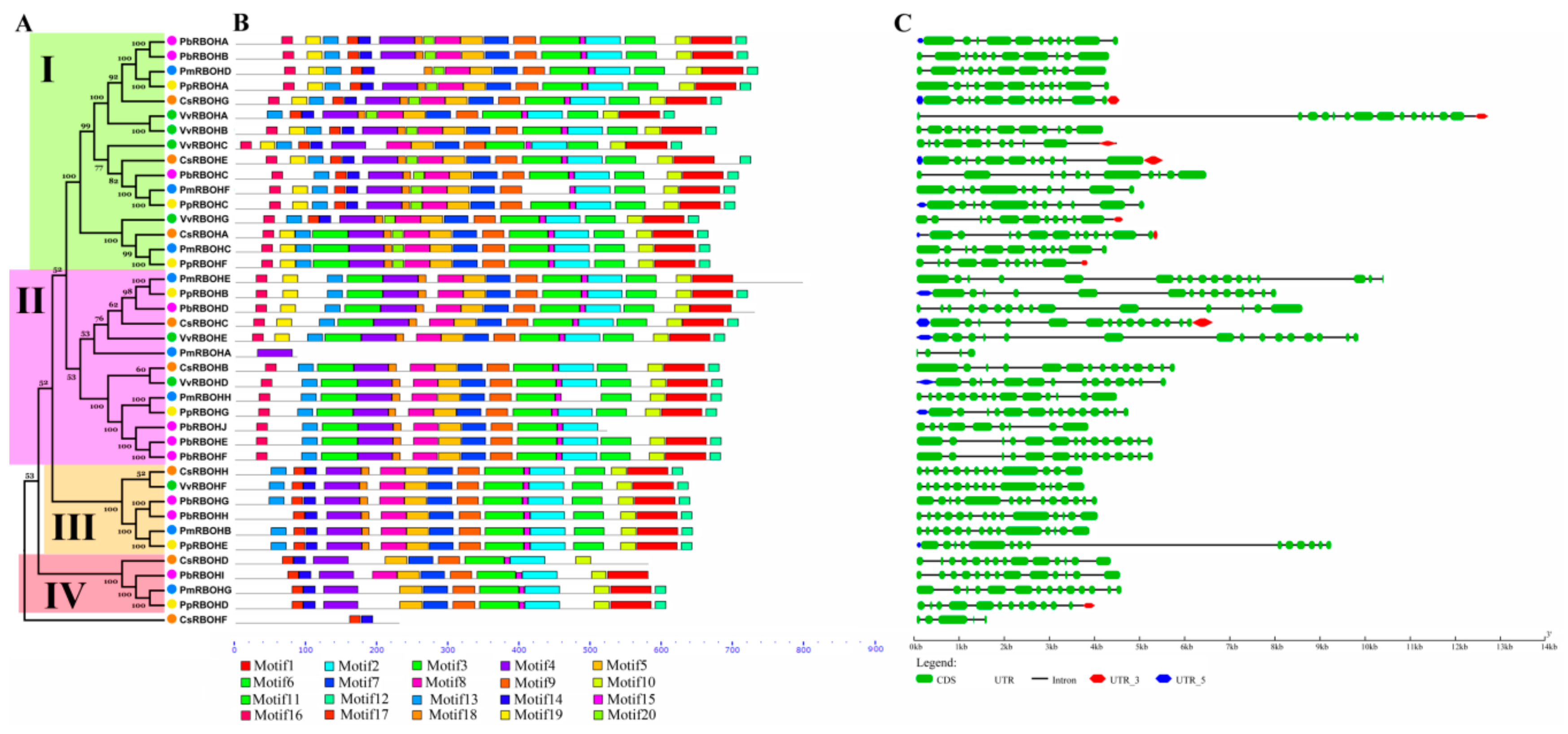
3.3. Phylogenetic Relationship, Conserved Motifs and Intron/Exon Structures of RBOHs in Five Fruit-Producing Trees
3.4. Amino Acid Sequence and Characteristic Domain Analysis of PbRBOHs
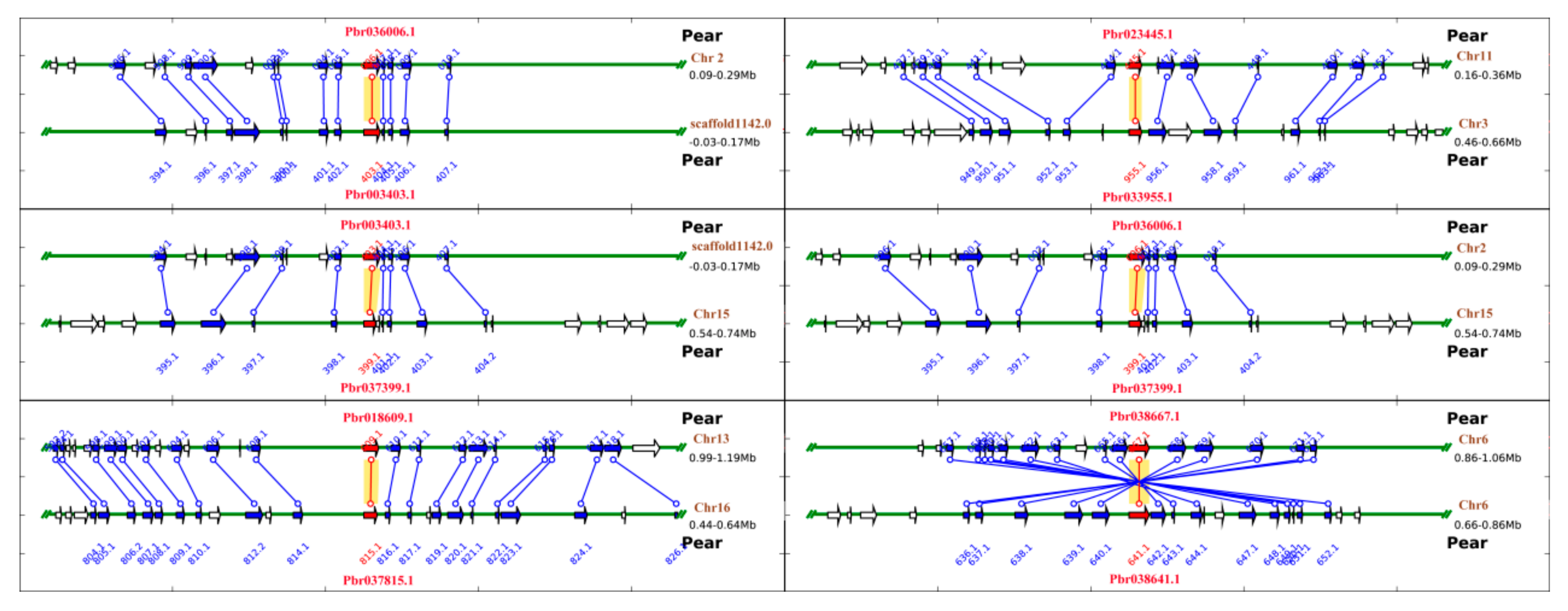
3.5. Microsynteny Analysis of PbRBOHs
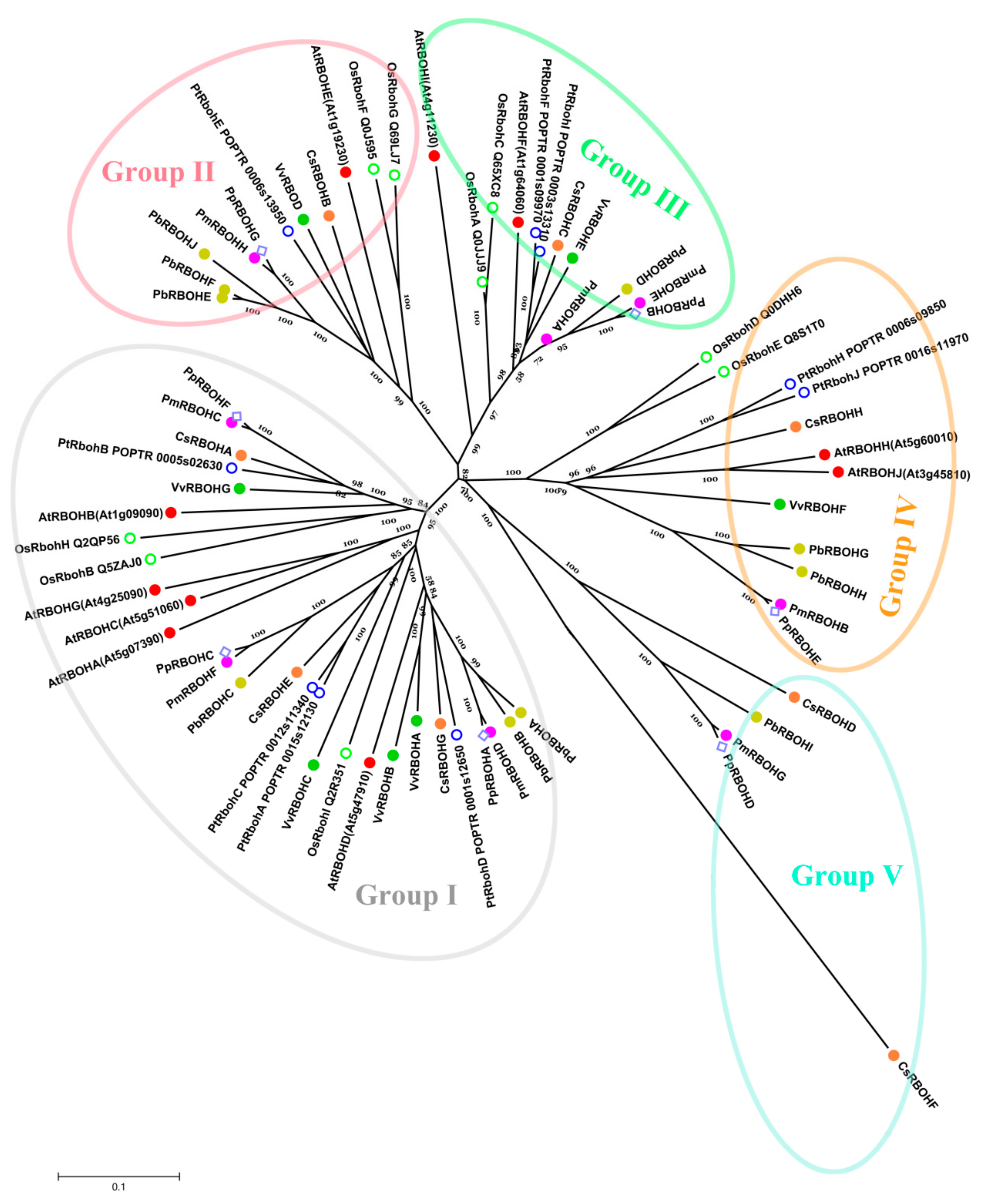
3.6. Comparative Phylogenetic Analysis and Functional Prediction of PbRBOHs
3.7. Analysis of Cis-Acting Elements in Putative PbRBOH Promoters
3.8. Differentially Expressed PbRBOHs under Hormonal Treatments
3.9. Histochemical Staining and Determination of Apoplastic Superoxide (O2.−) in Pear Fruit
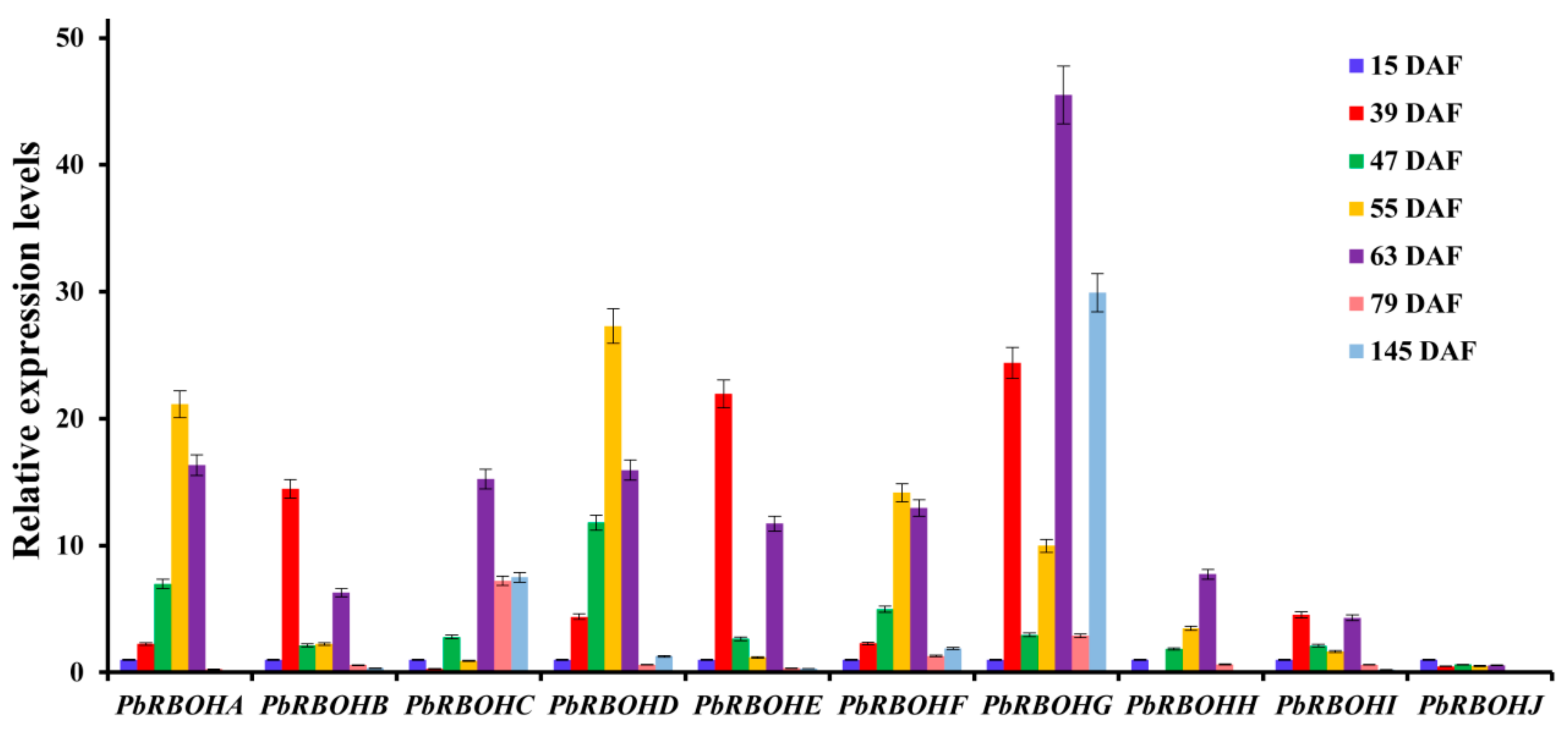
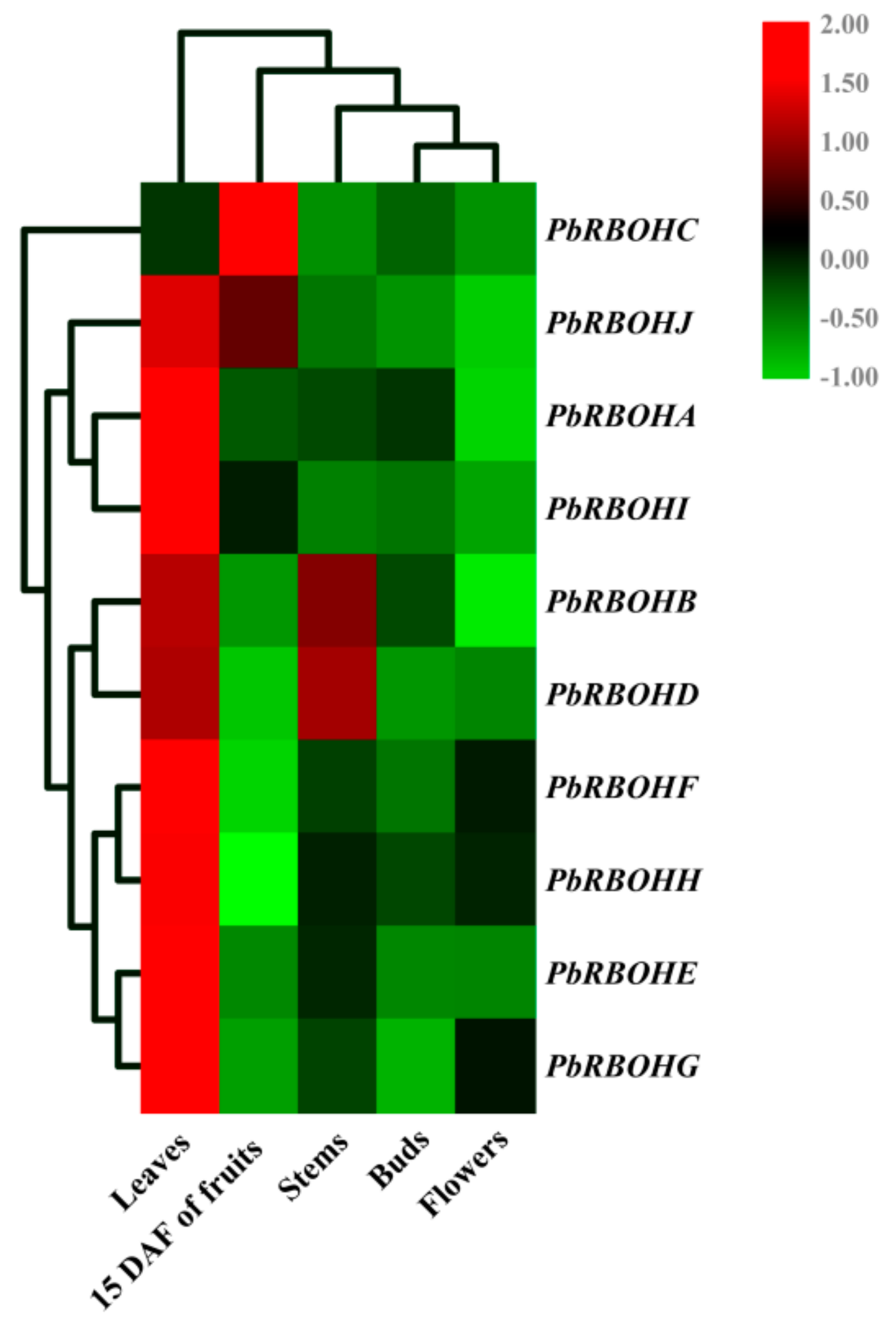
3.10. Tissue Specificity and Temporal Expression Patterns in Pear Fruit during Development
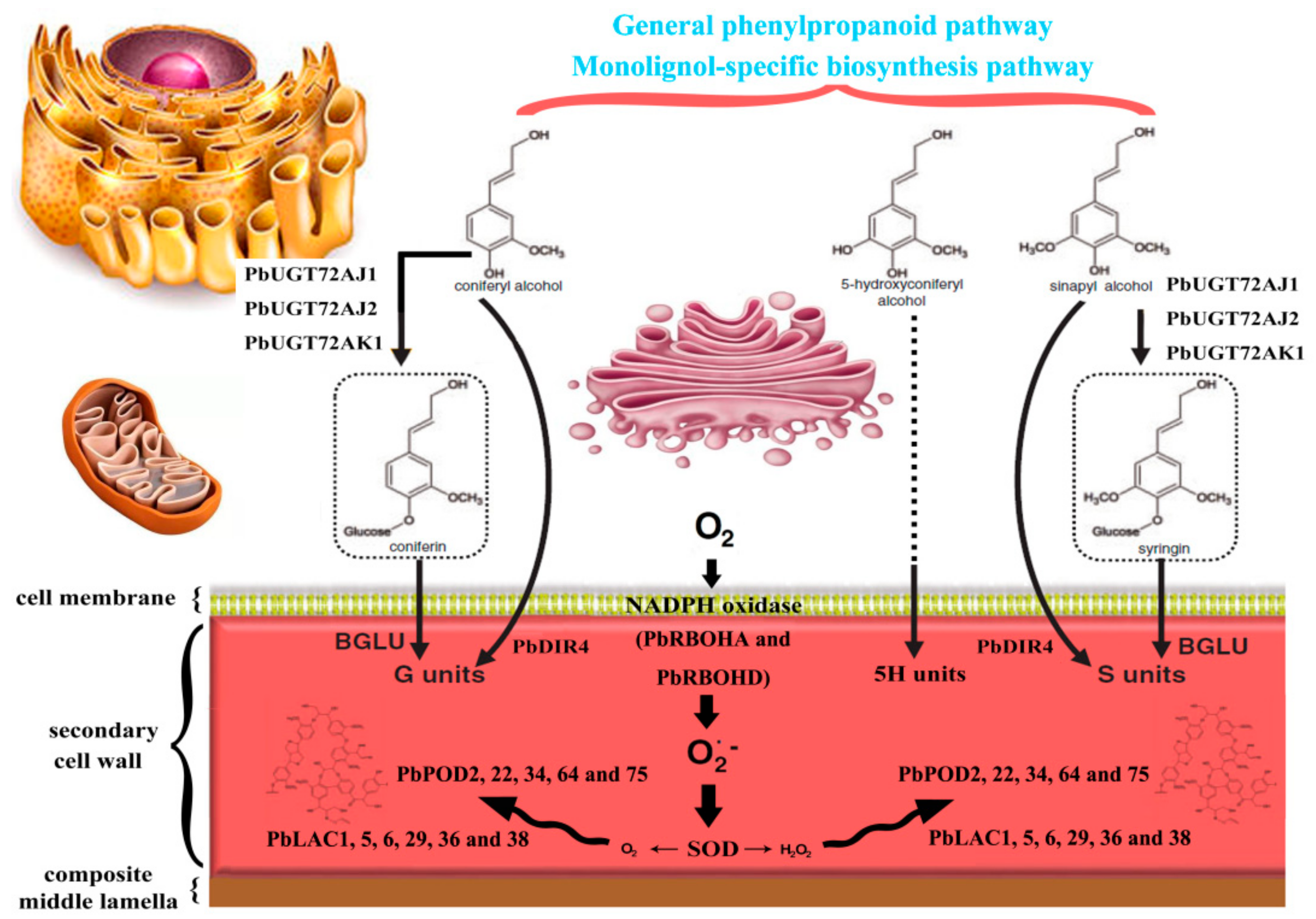
3.11. Subcellular Localization Analysis of Candidate PbRBOHs for Pear Stone Cell Lignification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wu, J.; Wang, Y.; Xu, J.; Korban, S.S.; Fei, Z.; Tao, S.; Ming, R.; Tai, S.; Khan, A.M.; Postman, J.D.; et al. Diversification and independent domestication of Asian and European pears. Genome Biol. 2018, 19, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Cao, Y.; Gao, Y.; Dong, X. Effect of stone cells size and flesh texture in pear cultivars. Acta Hortic. Sin. 2011, 38, 1225–1234. [Google Scholar]
- Bao, L.; Chen, K.; Zhang, D.; Cao, Y.; Yamamoto, T.; Teng, Y. Genetic diversity and similarity of pear (Pyrus L.) cultivars native to East Asia revealed by SSR (simple sequence repeat) markers. Genet. Resour. Crop Evol. 2007, 54, 959–971. [Google Scholar] [CrossRef]
- Cheng, X.; Su, X.; Muhammad, A.; Li, M.; Zhang, J.; Sun, Y.; Li, G.; Jin, Q.; Cai, Y.; Lin, Y. Molecular characterization, evolution, and expression profiling of the dirigent (DIR) family genes in Chinese white pear (Pyrus bretschneideri). Front. Genet. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Cheng, X.; Li, G.; Muhammad, A.; Zhang, J.; Jiang, T.; Jin, Q.; Zhao, H.; Cai, Y.; Lin, Y. Molecular identification, phylogenomic characterization and expression patterns analysis of the LIM (LIN-11, Isl1 and MEC-3 domains) gene family in pear (Pyrus bretschneideri) reveal its potential role in lignin metabolism. Gene 2019, 686, 237–249. [Google Scholar] [CrossRef]
- Zhang, J.; Cheng, X.; Jin, Q.; Su, X.; Li, M.; Yan, C.; Jiao, X.; Li, D.; Lin, Y.; Cai, Y. Comparison of the transcriptomic analysis between two Chinese white pear (Pyrus bretschneideri Rehd.) genotypes of different stone cells contents. PLoS ONE 2017, 12, 1–22. [Google Scholar] [CrossRef]
- Zhao, M.; Zhu, H. Development and morphology of stone cells in phloem of Toxicodendron vernicifluum. Trees Struct. Funct. 2014, 28, 1553–1558. [Google Scholar] [CrossRef]
- Barros, J.; Serk, H.; Granlund, I.; Pesquet, E. The cell biology of lignification in higher plants. Ann. Bot. 2015, 115, 1053–1074. [Google Scholar] [CrossRef] [Green Version]
- Xue, C.; Yao, J.L.; Qin, M.F.; Zhang, M.Y.; Allan, A.C.; Wang, D.F.; Wu, J. PbrmiR397a regulates lignification during stone cell development in pear fruit. Plant Biotechnol. J. 2018, 1–15. [Google Scholar] [CrossRef]
- Cai, Y.; Li, G.; Nie, J.; Lin, Y.; Nie, F.; Zhang, J.; Xu, Y. Study of the structure and biosynthetic pathway of lignin in stone cells of pear. Sci. Hortic. 2010, 125, 374–379. [Google Scholar] [CrossRef]
- Cheng, X.; Yan, C.; Zhang, J.; Ma, C.; Li, S.; Jin, Q.; Zhang, N.; Cao, Y.; Lin, Y.; Cai, Y. The effect of different pollination on the expression of Dangshan Su pear microRNA. Biomed. Res. Int. 2017, 2017, 2794040. [Google Scholar] [CrossRef]
- Yan, C.; Yin, M.; Zhang, N.; Jin, Q.; Fang, Z.; Lin, Y.; Cai, Y. Stone cell distribution and lignin structure in various pear varieties. Sci. Hortic. 2014, 174, 142–150. [Google Scholar] [CrossRef]
- Choi, J.; Choi, J.; Hong, K.; Kim, W. Cultivar differences of stone cells in pear flesh and their effects on fruit quality. Hortic. Environ. Biotechnol. 2007, 48, 17–31. [Google Scholar]
- Choi, J.; Lee, S. Distribution of stone cell in Asian, Chinese, and European pear fruit and its morphological changes. J. Appl. Bot. Food Qual. 2013, 189, 185–189. [Google Scholar] [CrossRef]
- Tian, L.; Dong, X.; Cao, Y.; Zhang, Y.; Qi, D. Correlation of flesh in Pyrus fruit with its stone cells lignin. Southwest China J. Agric. Sci. 2017, 30, 2091–2096. [Google Scholar] [CrossRef]
- Cheng, X.; Li, M.; Li, D.; Zhang, J.; Jin, Q.; Sheng, L.; Cai, Y.; Lin, Y. Characterization and analysis of CCR and CAD gene families at the whole-genome level for lignin synthesis of stone cells in pear (Pyrus bretschneideri) fruit. Biol. Open 2017, 6, 1602–1613. [Google Scholar] [CrossRef]
- Tao, S.; Khanizadeh, S.; Zhang, H.; Zhang, S. Anatomy, ultrastructure and lignin distribution of stone cells in two Pyrus species. Plant Sci. 2009, 176, 413–419. [Google Scholar] [CrossRef]
- Jin, Q.; Yan, C.; Qiu, J.; Zhang, N.; Lin, Y.; Cai, Y. Structural characterization and deposition of stone cell lignin in Dangshan Su pear. Sci. Hortic. 2013, 155, 123–130. [Google Scholar] [CrossRef]
- Cheng, X.; Muhammad, A.; Li, G.; Zhang, J.; Cheng, J.; Qiu, J.; Jiang, T. Family-1 UDP glycosyltransferases in pear (Pyrus bretschneideri): Molecular identification, phylogenomic characterization and expression profiling during stone cell formation. Mol. Biol. Rep. 2019, 46, 2153–2175. [Google Scholar] [CrossRef]
- Brahem, M.; Renard, C.M.G.C.; Gouble, B.; Bureau, S.; Le Bourvellec, C. Characterization of tissue specific differences in cell wall polysaccharides of ripe and overripe pear fruit. Carbohydr. Polym. 2017, 156, 152–164. [Google Scholar] [CrossRef]
- Li, S.; Su, X.; Abdullah, M.; Sun, Y.; Li, G.; Cheng, X.; Lin, Y.; Cai, Y.; Jin, Q. Effects of different pollens on primary metabolism and lignin biosynthesis in pear. Int. J. Mol. Sci. 2018, 19, 2273. [Google Scholar] [CrossRef]
- Zarei, A.; Zamani, Z.; Fatahi, R.; Mousavi, A.; Salami, S.A.; Avila, C.; Cánovas, F.M. Differential expression of cell wall related genes in the seeds of soft- and hard-seeded pomegranate genotypes. Sci. Hortic. 2016, 205, 7–16. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Yang, S.; Yuan, Y. Metabolite and Transcriptome analyses indicate the involvement of lignin in programmed changes in peach fruit texture. J. Agric. Food Chem. 2018, 66, acs.jafc.8b04284. [Google Scholar] [CrossRef]
- Jia, N.; Liu, J.; Sun, Y.; Tan, P.; Cao, H.; Xie, Y.; Wen, B.; Gu, T.; Liu, J.; Li, M.; et al. Citrus sinensis MYB transcription factors CsMYB330 and CsMYB308 regulate fruit juice sac lignification through fine-tuning expression of the Cs4CL1 gene. Plant Sci. 2018, 277, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Yin, X.R.; Zeng, J.K.; Ge, H.; Song, M.; Xu, C.J.; Li, X.; Ferguson, I.B.; Chen, K.S. Activator-and repressor-type MYB transcription factors are involved in chilling injury induced flesh lignification in loquat via their interactions with the phenylpropanoid pathway. J. Exp. Bot. 2014, 65, 4349–4359. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Z.; Shi, Z.; Zhang, S.; Ming, R.; Zhu, S.; Khan, M.A.; Tao, S.; Korban, S.S.; Wang, H.; et al. The genome of the pear (Pyrus bretschneideri Rehd.). Genome Res. 2013, 23, 396–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, C.; Xie, Z. Function analysis of VvmiR397a and its target genes VvLACs in grape berry development. Acta Hortic. Sin. 2018, 45, 1441–1455. [Google Scholar] [CrossRef]
- Lee, Y.; Rubio, M.C.; Alassimone, J.; Geldner, N. A mechanism for localized lignin deposition in the endodermis. Cell 2013, 153, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Koutaniemi, S. Lignin biosynthesis studies in plant tissue cultures. J. Integr. Plant Biol. 2010, 52, 176–185. [Google Scholar] [CrossRef]
- Shafi, A.; Gill, T.; Zahoor, I.; Singh, P.; Yelam, A.; Sanjay, S. Ectopic expression of SOD and APX genes in Arabidopsis alters metabolic pools and genes related to secondary cell wall cellulose biosynthesis and improve salt tolerance. Mol. Biol. Rep. 2019, 46, 1985–2002. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Yoon, T.H.; Lee, J.; Cho, H.K.; Mang, H.; Kwak, J.M.; Lee, Y.; Yoon, T.H.; Lee, J.; Jeon, S.Y.; et al. A lignin molecular brace controls precision processing of cell walls critical for surface integrity in Arabidopsis. Cell 2018, 173, 1468–1480. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; He, Y.; Hu, W.; Zhang, Y.; Wang, X.; Tang, H. Identification of NADPH oxidase family members associated with cold stress in strawberry. FEBS Open Bio 2018, 8, 593–605. [Google Scholar] [CrossRef]
- Meents, M.J.; Watanabe, Y.; Samuels, A.L. The cell biology of secondary cell wall biosynthesis. Ann. Bot. 2018, 1107–1125. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, L.; Zhang, Z.; Ma, M.; Wang, R.; Qian, M.; Zhang, S. Genome-wide identification and comparative analysis of the superoxide dismutase gene family in pear and their functions during fruit ripening. Postharvest Biol. Technol. 2018, 143, 68–77. [Google Scholar] [CrossRef]
- Xi, C.; Li, G.; Ma, C.; Abdullah, M.; Zhang, J.; Zhao, H.; Jin, Q.; Cai, Y.; Lin, Y. Comprehensive genome-wide analysis of the pear (Pyrus bretschneideri) laccase gene (PbLAC) family and functional identification of PbLAC1 involved in lignin biosynthesis. PLoS ONE 2019, 14, e0210892. [Google Scholar]
- Cao, Y.; Han, Y.; Meng, D.; Li, D.; Jin, Q.; Lin, Y. Structural, evolutionary, and functional analysis of the Class III peroxidase gene family in Chinese pear (Pyrus bretschneideri). Front. Plant Sci. 2016, 7, 1874. [Google Scholar] [CrossRef] [PubMed]
- Sagi, M. Production of reactive oxygen species by plant NADPH oxidases. Plant Physiol. 2006, 141, 336–340. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2018, 47, 427–432. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2018, 46, D493–D496. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Xu, Y.; Jiang, H.; Jiang, C.; Du, Y.; Gong, C.; Wang, W.; Zhu, S.; Han, G.; Cheng, B. Systematic identification, evolution and expression analysis of the Zea mays PHT1 gene family reveals several new members involved in root colonization by arbuscular mycorrhizal fungi. Int. J. Mol. Sci. 2016, 17, 1. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.X.; Jiang, H.Y.; Chu, Z.X.; Tang, X.L.; Zhu, S.W.; Cheng, B.J. Genome-wide identification, classification and analysis of heat shock transcription factor family in maize. BMC Genom. 2011, 12, 76. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.; Zhang, H.; Luo, J. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Xiong, Y.; Li, D.H.; Cheng, J.; Cao, Y.P.; Yan, C.C.; Jin, Q.; Sun, N.; Cai, Y.P.; Lin, Y. Bioinformatic and expression analysis of the OMT gene family in Pyrus bretschneideri cv. Dangshan Su. Genet. Mol. Res. 2016, 15, 1–17. [Google Scholar] [CrossRef]
- Abdullah, M.; Cao, Y.; Cheng, X.; Shakoor, A.; Su, X.; Gao, J.; Cai, Y. Genome-wide analysis characterization and evolution of SBP Genes in Fragaria vesca, Pyrus bretschneideri, Prunus persica and Prunus mume. Front. Genet. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Luo, T.; Fu, X.; Fan, Q.; Liu, J. Cloning and molecular characterization of a mitogen-activated protein kinase gene from Poncirus trifoliata whose ectopic expression confers dehydration/drought tolerance in transgenic tobacco. J. Exp. Bot. 2011, 62, 5191–5206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kaur, G.; Pati, P.K. In silico insights on diverse interacting partners and phosphorylation sites of respiratory burst oxidase homolog (Rbohs) gene families from Arabidopsis and rice. BMC Plant Biol. 2018, 18, 161. [Google Scholar] [CrossRef]
- Kaur, G.; Guruprasad, K.; Temple BR, S.; Shirvanyants, D.G.; Dokholyan, N.V.; Pati, P.K. Structural complexity and functional diversity of plant NADPH oxidases. Amino Acids 2018, 50, 79–94. [Google Scholar] [CrossRef]
- Lightfoot, D.; Boettcher, A.; Little, A.; Shirley, N.; Able, A.J. Identification and characterisation of barley (Hordeum vulgare) respiratory burst oxidase homologue family members. Funct. Plant Biol. 2008, 35, 347–359. [Google Scholar] [CrossRef]
- Zhu, Y.; Du, B.; Qian, J.; Zou, B.; Hua, J. Disease Resistance gene-induced growth inhibition is enhanced by rcd1 independent of defense activation in Arabidopsis. Plant Physiol. 2013, 161, 2005–2013. [Google Scholar] [CrossRef]
- Kwak, J.M.; Mori, I.C.; Pei, Z.; Leonhardt, N.; Torres, M.A.; Dangl, J.L.; Bloom, R.E.; Bodde, S.; Jones JD, G.; Schroeder, J.I. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J. 2003, 22, 2623–2633. [Google Scholar] [CrossRef]
- Cepauskas, D.; Miliute, I.; Staniene, G.; Gelvonauskiene, D.; Stanys, V.; Jesaitis, A.J.; Baniulis, D. Characterization of apple NADPH oxidase genes and their expression associated with oxidative stress in shoot culture in vitro. Plant Cell. Tissue Organ Cult. 2016, 124, 621–633. [Google Scholar] [CrossRef]
- Kaur, G.; Pati, P.K. Analysis of cis-acting regulatory elements of Respiratory burst oxidase homolog (Rboh) gene families in Arabidopsis and rice provides clues for their diverse functions. Comput. Biol. Chem. 2016, 62, 104–118. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Ho, T.D. Functional dissection of an abscisic acid (ABA)-inducible gene reveals two independent ABA-responsive complexes each containing a G-Box and a nove1 cis-acting element. Plant Cell 1995, 7, 295–307. [Google Scholar] [CrossRef]
- Li, X.; Singh, J.; Qin, M.; Li, S.; Zhang, X.; Zhang, M.; Khan, A.; Zhang, S.; Wu, J. Development of an integrated 200K SNP genotyping array and application for genetic mapping, genome assembly improvement and genome wide association studies in pear (Pyrus). Plant Biotechnol. J. 2019. [Google Scholar] [CrossRef] [PubMed]
- Berens, M.L.; Berry, H.M.; Mine, A.; Argueso, C.T.; Tsuda, K. Evolution of hormone signaling networks in plant defense. Annu. Rev. Phytopathol. 2017, 55, 401–425. [Google Scholar] [CrossRef]
- Tao, S.; Wang, D.; Jin, C.; Sun, W.; Liu, X.; Zhang, S.; Gao, F.; Khanizadeh, S. Cinnamate-4-hydroxylase gene is involved in the step of lignin biosynthesis in Chinese white pear. J. Am. Soc. Hortic. Sci. 2015, 140, 573–579. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Wang, R.; Bai, Y.; Liu, C.; Yuan, Y.; Yang, Y.; Yang, S. Differential gene expression analysis of “Chili” (Pyrus bretschneideri) fruit pericarp with two types of bagging treatments. Hortic. Res. 2017, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, J.M.; Huang, X.S.; Li, L.T.; Zheng, D.M.; Xue, C.; Zhang, S.L.; Wu, J. Proteome analysis of pear reveals key genes associated with fruit development and quality. Planta 2015, 241, 1363–1379. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sequence ID | Name | Chr | Protein | |||
|---|---|---|---|---|---|---|---|
| Length (aa) | MW (kDa) | pI | Subcellular Localization Predicted | ||||
| Pear | Pbr018609.1 | PbRBOHA | 13 | 962 | 108.07 | 9.02 | Plas |
| Pbr037815.1 | PbRBOHB | 16 | 964 | 108.42 | 9.11 | Plas | |
| Pbr038667.1 | PbRBOHC | 6 | 947 | 106.18 | 8.07 | Plas | |
| Pbr006277.1 | PbRBOHD | 6 | 978 | 111.34 | 9.37 | Cyto | |
| Pbr036006.1 | PbRBOHE | 2 | 915 | 103.56 | 8.59 | Cyto | |
| Pbr003403.1 | PbRBOHF | / | 914 | 103.11 | 8.59 | Cyto | |
| Pbr023445.1 | PbRBOHG | 11 | 856 | 97.65 | 8.66 | Cyto | |
| Pbr033955.1 | PbRBOHH | 3 | 860 | 97.56 | 8.79 | Cyto | |
| Pbr007212.1 | PbRBOHI | 14 | 779 | 89.07 | 9.06 | Plas | |
| Pbr037399.1 | PbRBOHJ | 15 | 701 | 79.09 | 9.29 | Chlo | |
| Peach | Prupe.1G211000 | PpRBOHA | 1 | 971 | 108.87 | 9.18 | Chlo |
| Prupe.5G107400 | PpRBOHB | 5 | 964 | 109.47 | 9.32 | Chlo | |
| Prupe.5G138300 | PpRBOHC | 5 | 941 | 105.22 | 8.97 | Plas | |
| Prupe.5G204900 | PpRBOHD | 5 | 810 | 92.48 | 8.85 | Plas | |
| Prupe.6G088800 | PpRBOHE | 6 | 859 | 98.48 | 8.88 | Nucl | |
| Prupe.6G321500 | PpRBOHF | 6 | 893 | 101.94 | 8.87 | Nucl | |
| Prupe.7G193000 | PpRBOHG | 7 | 906 | 102.84 | 8.93 | Cyto | |
| Mei | Pm000665 | PmRBOHA | 1 | 116 | 13.74 | 4.58 | Cyto |
| Pm000754 | PmRBOHB | 1 | 860 | 98.62 | 8.83 | Cyto | |
| Pm003305 | PmRBOHC | 1 | 893 | 102.01 | 9.00 | Nucl | |
| Pm007134 | PmRBOHD | 2 | 982 | 110.11 | 9.02 | Nucl | |
| Pm023955 | PmRBOHE | 7 | 1067 | 121.09 | 9.29 | Chlo | |
| Pm024297 | PmRBOHF | 7 | 940 | 105.50 | 8.73 | Chlo | |
| Pm024959 | PmRBOHG | 7 | 810 | 92.32 | 8.81 | Plas | |
| Pm027130 | PmRBOHH | 8 | 914 | 103.69 | 8.74 | Plas | |
| Grape | GSVIVT01001122001 | VvRBOHA | 1 | 827 | 94.13 | 9.16 | Chlo |
| GSVIVT01001123001 | VvRBOHB | 1 | 906 | 102.80 | 9.13 | Nucl | |
| GSVIVT01014350001 | VvRBOHC | 19 | 840 | 95.40 | 6.47 | Plas | |
| GSVIVT01015025001 | VvRBOHD | 11 | 917 | 103.81 | 9.26 | Chlo | |
| GSVIVT01019429001 | VvRBOHE | 2 | 922 | 104.75 | 9.26 | Cyto | |
| GSVIVT01025074001 | VvRBOHF | 6 | 852 | 97.14 | 9.12 | Cyto | |
| GSVIVT01031128001 | VvRBOHG | 14 | 873 | 99.44 | 9.34 | Cyto | |
| Sweet orange | Cs3g14240.1 | CsRBOHA | 3 | 889 | 101.04 | 9.14 | Cyto |
| Cs4g06920.1 | CsRBOHB | 4 | 910 | 103.34 | 9.19 | Chlo | |
| Cs5g02940.1 | CsRBOHC | 5 | 946 | 107.50 | 9.37 | Chlo | |
| Cs5g11890.1 | CsRBOHD | 5 | 777 | 88.70 | 8.57 | Plas | |
| Cs7g19320.2 | CsRBOHE | 7 | 970 | 109.69 | 8.78 | Nucl | |
| Cs7g19380.1 | CsRBOHF | 7 | 308 | 35.23 | 6.72 | Plas | |
| Cs8g12000.1 | CsRBOHG | 8 | 915 | 103.52 | 9.11 | Plas | |
| Cs8g17640.1 | CsRBOHH | 8 | 842 | 96.61 | 9.11 | Plas | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, X.; Li, G.; Manzoor, M.A.; Wang, H.; Abdullah, M.; Su, X.; Zhang, J.; Jiang, T.; Jin, Q.; Cai, Y.; et al. In Silico Genome-Wide Analysis of Respiratory Burst Oxidase Homolog (RBOH) Family Genes in Five Fruit-Producing Trees, and Potential Functional Analysis on Lignification of Stone Cells in Chinese White Pear. Cells 2019, 8, 520. https://doi.org/10.3390/cells8060520
Cheng X, Li G, Manzoor MA, Wang H, Abdullah M, Su X, Zhang J, Jiang T, Jin Q, Cai Y, et al. In Silico Genome-Wide Analysis of Respiratory Burst Oxidase Homolog (RBOH) Family Genes in Five Fruit-Producing Trees, and Potential Functional Analysis on Lignification of Stone Cells in Chinese White Pear. Cells. 2019; 8(6):520. https://doi.org/10.3390/cells8060520
Chicago/Turabian StyleCheng, Xi, Guohui Li, Muhammad Aamir Manzoor, Han Wang, Muhammad Abdullah, Xueqiang Su, Jingyun Zhang, Taoshan Jiang, Qing Jin, Yongping Cai, and et al. 2019. "In Silico Genome-Wide Analysis of Respiratory Burst Oxidase Homolog (RBOH) Family Genes in Five Fruit-Producing Trees, and Potential Functional Analysis on Lignification of Stone Cells in Chinese White Pear" Cells 8, no. 6: 520. https://doi.org/10.3390/cells8060520
APA StyleCheng, X., Li, G., Manzoor, M. A., Wang, H., Abdullah, M., Su, X., Zhang, J., Jiang, T., Jin, Q., Cai, Y., & Lin, Y. (2019). In Silico Genome-Wide Analysis of Respiratory Burst Oxidase Homolog (RBOH) Family Genes in Five Fruit-Producing Trees, and Potential Functional Analysis on Lignification of Stone Cells in Chinese White Pear. Cells, 8(6), 520. https://doi.org/10.3390/cells8060520