C89 Induces Autophagy of Female Germline Stem Cells via Inhibition of the PI3K-Akt Pathway In Vitro

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Chemical Compounds
2.3. Culture of FGSCs In Vitro
2.4. Cell Counting Kit 8 and 5-Ethynyl-2’-Deoxyuridine Labeling Assay
2.5. RNA Isolation and Reverse Transcription-Polymerase Chain Reaction
2.6. Quantitative Real-Time Polymerase Chain Reaction
2.7. Cell Apoptosis Assay
2.8. Western Blotting
2.9. Immunofluorescence
2.10. RNA Isolation and RNA Sequencing
2.11. Gene Ontology and Kyoto Encyclopedia of Genes and Genomes Pathway Analysis
2.12. Statistical Analyses
3. Results
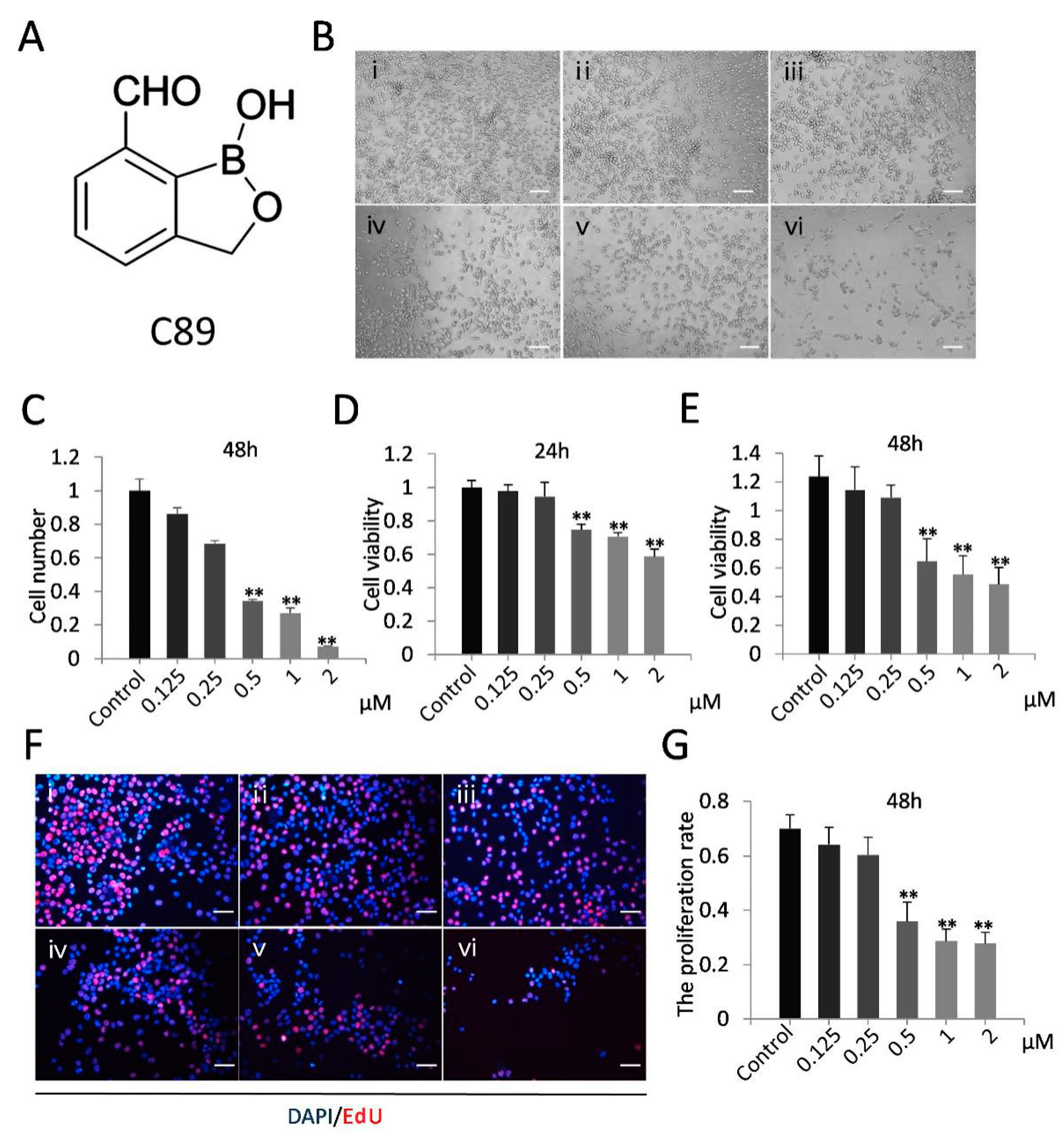
3.1. C89 Reduced Cell Number and Inhibited Cell Viability and Proliferation of FGSCs In Vitro
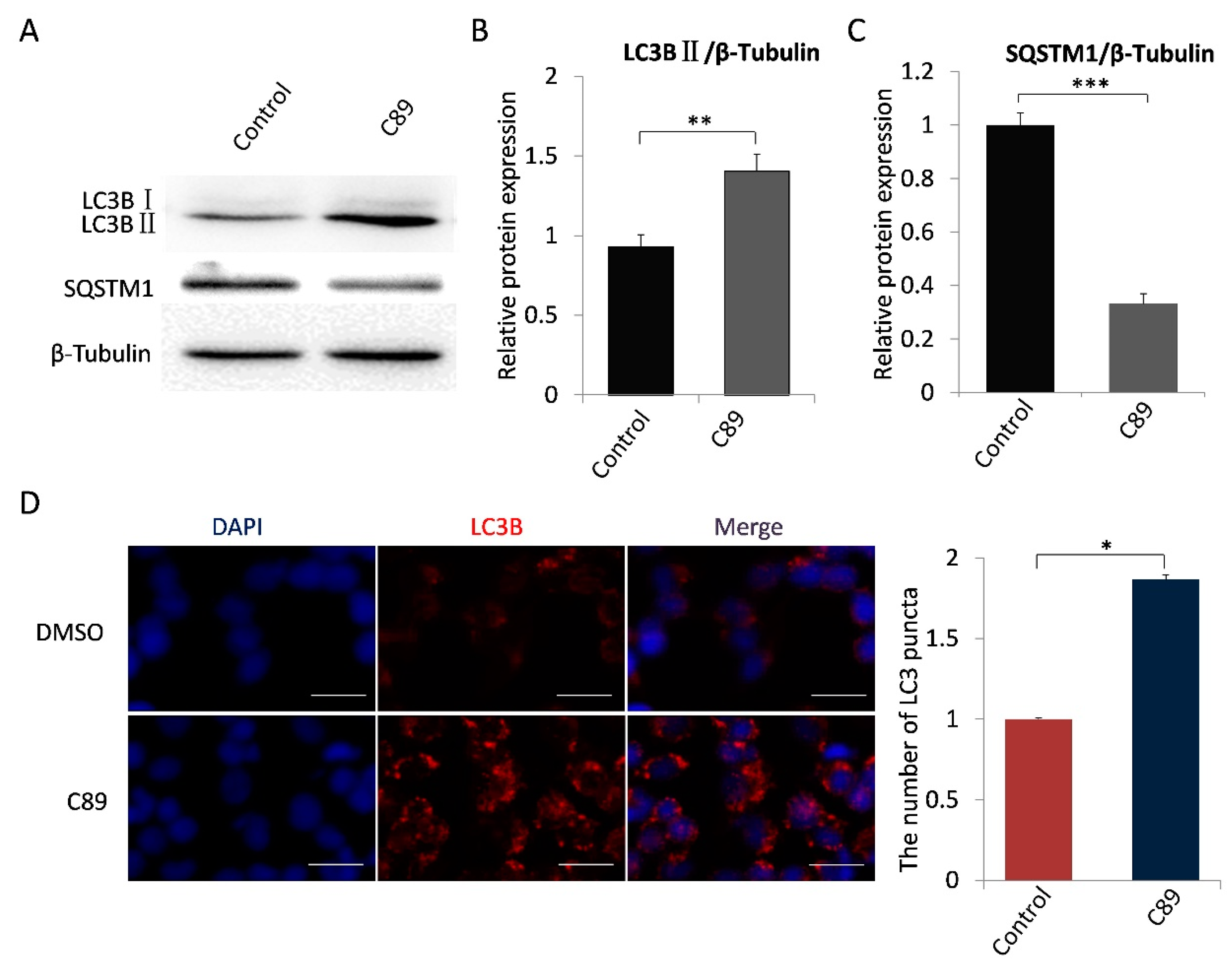
3.2. C89 Induced Autophagy but Not Apoptosis or Differentiation of FGSCs In Vitro
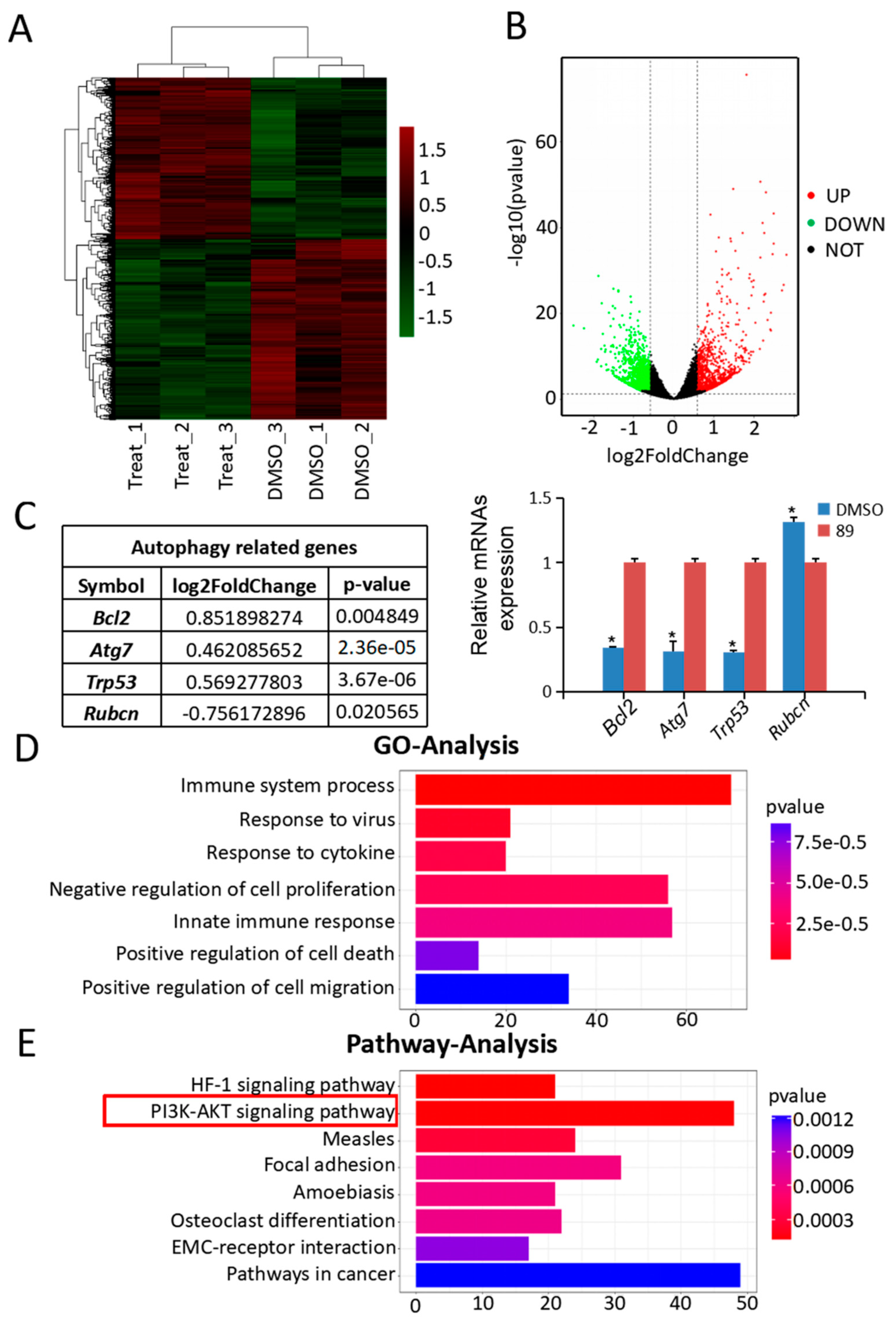
3.3. Autophagy-Associated Genes and Pathways in C89-Treated FGSCs
3.4. C89 Induced FGSC Autophagy by Inhibiting the Phosphorylation of PI3K and Akt
3.5. Cooperative Functions of C89 and LY294002 in Inducing Autophagy via Suppressing the PI3K-Akt Pathway
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- White, Y.A.; Woods, D.C.; Takai, Y.; Ishihara, O.; Seki, H.; Tilly, J.L. Oocyte formation by mitotically active germ cells purified from ovaries of reproductive-age women. Nat. Med. 2012, 18, 413–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Yang, Z.; Yang, Y.; Wang, S.; Shi, L.; Xie, W.; Sun, K.; Zou, K.; Wang, L.; Xiong, J.; et al. Production of transgenic mice by random recombination of targeted genes in female germline stem cells. J. Mol. Cell Biol. 2011, 3, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Bai, Y.; Chu, Z.; Wang, J.; Wang, L.; Yu, M.; Lian, Z.; Hua, J. Gsk3 inhibitor-bioregulates proliferation of female germline stem cells from the postnatal mouse ovary. Cell Prolif. 2012, 45, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Zou, K.; Yuan, Z.; Yang, Z.; Luo, H.; Sun, K.; Zhou, L.; Xiang, J.; Shi, L.; Yu, Q.; Zhang, Y.; et al. Production of offspring from a germline stem cell line derived from neonatal ovaries. Nat. Cell Biol. 2009, 11, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Liu, G.; Xu, B.; Wu, C.; Hui, N.; Ni, X.; Wang, J.; Du, M.; Teng, X.; Wu, J. Human GV oocytes generated by mitotically active germ cells obtained from follicular aspirates. Sci. Rep. 2016, 6, 28218. [Google Scholar] [CrossRef]
- Li, X.; Ao, J.; Wu, J. Systematic identification and comparison of expressed profiles of lncrnas and circrnas with associated co-expression and cerna networks in mouse germline stem cells. Oncotarget 2017, 8, 26573–26590. [Google Scholar] [CrossRef]
- Wu, C.; Xu, B.; Li, X.; Ma, W.; Zhang, P.; Chen, X.; Wu, J. Tracing and characterizing the development of transplanted female germline stem cells in vivo. Mol. J. Am. Soc. Gene 2017, 25, 1408–1419. [Google Scholar] [CrossRef]
- Zhu, X.; Tian, G.G.; Yu, B.; Yang, Y.; Wu, J. Effects of bisphenol a on ovarian follicular development and female germline stem cells. Arch. Toxicol. 2018, 92, 1581–1591. [Google Scholar] [CrossRef]
- Zou, K.; Wang, J.; Bi, H.; Zhang, Y.; Tian, X.; Tian, N.; Ma, W.; Wu, J. Comparison of different in vitro differentiation conditions for murine female germline stem cells. Cell Prolif. 2019, 52, e12530. [Google Scholar] [CrossRef]
- Wu, J.; Ding, X.; Wang, J. Stem cells in mammalian gonads. Results Probl. Cell Differ. 2016, 58, 289–307. [Google Scholar]
- Nakatogawa, H.; Suzuki, K.; Kamada, Y.; Ohsumi, Y. Dynamics and diversity in autophagy mechanisms: Lessons from yeast. Nat. Rev. Mol. Cell Biol. 2009, 10, 458–467. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Klionsky, D.J. Regulation mechanisms and signaling pathways of autophagy. Annu. Rev. Genet. 2009, 43, 67–93. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ng, S.; Wang, J.; Zhou, J.; Tan, S.H.; Yang, N.; Lin, Q.; Xia, D.; Shen, H.M. Histone deacetylase inhibitors induce autophagy through foxo1-dependent pathways. Autophagy 2015, 11, 629–642. [Google Scholar] [CrossRef] [PubMed]
- Ravanan, P.; Srikumar, I.F.; Talwar, P. Autophagy: The spotlight for cellular stress responses. Life Sci. 2017, 188, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Choi, A.M.; Ryter, S.W.; Levine, B. Autophagy in human health and disease. N. Engl. J. Med. 2013, 368, 651–662. [Google Scholar] [CrossRef]
- Levine, B.; Kroemer, G. Autophagy in the pathogenesis of disease. Cell 2008, 132, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Maiuri, M.C.; Kroemer, G. Autophagy in stress and disease. Cell Death Differ. 2015, 22, 365–366. [Google Scholar] [CrossRef] [Green Version]
- White, E.; Mehnert, J.M.; Chan, C.S. Autophagy, metabolism, and cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2015, 21, 5037–5046. [Google Scholar] [CrossRef]
- Netea-Maier, R.T.; Plantinga, T.S.; Van de Veerdonk, F.L.; Smit, J.W.; Netea, M.G. Modulation of inflammation by autophagy: Consequences for human disease. Autophagy 2016, 12, 245–260. [Google Scholar] [CrossRef] [Green Version]
- Revuelta, M.; Matheu, A. Autophagy in stem cell aging. Aging Cell 2017, 16, 912–915. [Google Scholar] [CrossRef]
- Ho, T.T.; Warr, M.R.; Adelman, E.R.; Lansinger, O.M.; Flach, J.; Verovskaya, E.V.; Figueroa, M.E.; Passegue, E. Autophagy maintains the metabolism and function of young and old stem cells. Nature 2017, 543, 205–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jungverdorben, J.; Till, A.; Brustle, O. Induced pluripotent stem cell-based modeling of neurodegenerative diseases: A focus on autophagy. J. Mol. Med. 2017, 95, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Fleming, A.; Ricketts, T.; Pavel, M.; Virgin, H.; Menzies, F.M.; Rubinsztein, D.C. Autophagy regulates notch degradation and modulates stem cell development and neurogenesis. Nat. Commun. 2016, 7, 10533. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Prat, L.; Martinez-Vicente, M.; Perdiguero, E.; Ortet, L.; Rodriguez-Ubreva, J.; Rebollo, E.; Ruiz-Bonilla, V.; Gutarra, S.; Ballestar, E.; Serrano, A.L.; et al. Autophagy maintains stemness by preventing senescence. Nature 2016, 529, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Heras-Sandoval, D.; Perez-Rojas, J.M.; Hernandez-Damian, J.; Pedraza-Chaverri, J. The role of pi3k/akt/mtor pathway in the modulation of autophagy and the clearance of protein aggregates in neurodegeneration. Cell Signal. 2014, 26, 2694–2701. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Luo, F.; Ling, M.; Lu, L.; Shi, L.; Lu, X.; Xu, H.; Chen, C.; Yang, Q.; Xue, J.; et al. Micro rna-21 activation of erk signaling via pten is involved in arsenite-induced autophagy in human hepatic l-02 cells. Toxicol. Lett. 2016, 252, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Jiang, K.; Ding, S. Concise review: A chemical approach to control cell fate and function. Stem Cells 2012, 30, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Schugar, R.C.; Robbins, P.D.; Deasy, B.M. Small molecules in stem cell self-renewal and differentiation. Gene Ther. 2008, 15, 126–135. [Google Scholar] [CrossRef]
- Lin, S.R.; Fu, Y.S.; Tsai, M.J.; Cheng, H.; Weng, C.F. Natural compounds from herbs that can potentially execute as autophagy inducers for cancer therapy. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef]
- Deng, Y.; Zhu, L.; Cai, H.; Wang, G.; Liu, B. Autophagic compound database: A resource connecting autophagy-modulating compounds, their potential targets and relevant diseases. Cell Prolif. 2018, 51, e12403. [Google Scholar] [CrossRef]
- Li, W.; Ding, S. Small molecules that modulate embryonic stem cell fate and somatic cell reprogramming. Trends Pharm. Sci. 2010, 31, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yang, Y.J.; Wang, H.; Dong, Q.T.; Wang, T.J.; Qian, H.Y.; Xu, H. Autophagy activation: A novel mechanism of atorvastatin to protect mesenchymal stem cells from hypoxia and serum deprivation via amp-activated protein kinase/mammalian target of rapamycin pathway. Stem Cells Dev. 2012, 21, 1321–1332. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Ding, D.; Feng, Y.; Xie, D.; Wu, P.; Guo, H.; Meng, Q.; Zhou, H. Convenient and versatile synthesis of formyl-substituted benzoxaboroles. Tetrahedron 2009, 65, 8738–8744. [Google Scholar] [CrossRef]
- Ding, D.; Meng, Q.; Gao, G.; Zhao, Y.; Wang, Q.; Nare, B.; Jacobs, R.; Rock, F.; Alley, M.R.K.; Plattner, J.J.; et al. Design, synthesis, and structure−activity relationship of Trypanosoma brucei leucyl-trna synthetase inhibitors as antitrypanosomal agents. J. Med. Chem. 2011, 54, 1276–1287. [Google Scholar] [CrossRef] [PubMed]
- Manhas, R.; Tandon, S.; Sen, S.S.; Tiwari, N.; Munde, M.; Madhubala, R. Leishmania donovani parasites are inhibited by the benzoxaborolean 2690 targeting leucyl-trnasynthetase. Antimicrob. Agents Chemother. 2018, 62, e00079–e00118. [Google Scholar] [PubMed]
- Hu, L.; Zaloudek, C.; Mills, G.B.; Gray, J.; Jaffe, R.B. In vivo and in vitro ovarian carcinoma growth inhibition by a phosphatidyl inositol 3-kinase inhibitor (ly294002). Clin. Cancer Res. 2000, 6, 880–886. [Google Scholar] [PubMed]
- Zhang, C.; Wu, J. Production of offspring from a germline stem cell line derived from prepubertal ovaries of germline reporter mice. Mol. Hum. Reprod. 2016, 22, 457–464. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, L.; Kang, J.X.; Xie, W.; Li, X.; Wu, C.; Xu, B.; Wu, J. Production of fat-1 transgenic rats using a post-natal female germline stem cellline. Mol. Hum. Reprod. 2014, 20, 271–281. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, J.; Wu, H.; Pei, X.; Chang, Q.; Ma, W.; Ma, H.; Hei, C.; Zheng, X.; Cai, Y.; et al. The increased expression of connexin and vegfin mouse ovarian tissue vitrification by follicle stimulating hormone. Biomed. Res. Int. 2015, 2015, 397264. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, K.H.; Yoon, H.; Lee, O.H.; Kim, E.; Park, M.; Jang, H.; Hong, K.; Song, H.; Ko, J.J.; et al. Rasd1 knock down results in failure of oocyte maturation. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharm. 2016, 40, 1289–1302. [Google Scholar] [CrossRef]
- Xie, F.; Liu, W.; Feng, F.; Li, X.; He, L.; Lv, D.; Qin, X.; Li, L.; Li, L.; Chen, L. Apelin-13 promotes cardiomyocyte hypertrophy via pi3k-akt-erk1/2-p70s6k and pi3k-induced autophagy. Acta Biochim. Biophys. Sin. 2015, 47, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Hu, Z.; Xu, H.; Xu, J.; Zhang, S.; Zhong, Y.; He, X.; Wang, N. Advanced glycation end products trigger autophagy in cardiomyocyte via rage/pi3k/akt/mtor pathway. Cardiovasc. Diabetol. 2014, 13, 78. [Google Scholar] [CrossRef] [PubMed]
- Seranova, E.; Connolly, K.J.; Zatyka, M.; Rosenstock, T.R.; Barrett, T.; Tuxworth, R.I.; Sarkar, S. Dysregulation of autophagy as a common mechanism in lysosomal storage diseases. Essays Biochem. 2017, 61, 733–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, P.; Kriel, J.; Shubha Priya, B.; Shivananju, N.S.; Loos, B. Modulating autophagy in cancer therapy: Advancements and challenges for cancer cell death sensitization. Biochem. Pharm. 2018, 147, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Kuang, W.; Wu, B.; Xie, C.; Liu, C.; Tu, Z. Il-12 induces autophagy in human breast cancer cells through ampk and the pi3k/akt pathway. Mol. Med. Rep. 2017, 16, 4113–4118. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Fang, L.; Wu, H.; Mei, X.; He, F.; Ding, P.; Liu, R. Tlr2 regulates allergic airway inflammation and autophagy through pi3k/akt signaling pathway. Inflammation 2017, 40, 1382–1392. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, J.X.; Zhang, Q.L. Pi3k/akt/mtor-mediated autophagy in the development of autism spectrum disorder. Brain Res. Bull. 2016, 125, 152–158. [Google Scholar] [CrossRef]
- Fruman, D.A.; Chiu, H.; Hopkins, B.D.; Bagrodia, S.; Cantley, L.C.; Abraham, R.T. The pi3k pathway in human disease. Cell 2017, 170, 605–635. [Google Scholar] [CrossRef]
- Shu, L.; Zhang, W.; Huang, C.; Huang, G.; Su, G. Troxerutin protects against myocardial ischemia/reperfusion injury via pi3k/akt pathway in rats. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharm. 2017, 44, 1939–1948. [Google Scholar] [CrossRef]
- Shao, Y.; Wolf, P.G.; Guo, S.; Guo, Y.; Gaskins, H.R.; Zhang, B. Zinc enhances intestinal epithelial barrier function through the pi3k/akt/mtor signaling pathway in caco-2cells. J. Nutr. Biochem. 2017, 43, 18–26. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Hu, X.; Tian, G.G.; Cheng, P.; Li, Z.; Zhu, M.; Zhou, H.; Wu, J. C89 Induces Autophagy of Female Germline Stem Cells via Inhibition of the PI3K-Akt Pathway In Vitro. Cells 2019, 8, 606. https://doi.org/10.3390/cells8060606
Li X, Hu X, Tian GG, Cheng P, Li Z, Zhu M, Zhou H, Wu J. C89 Induces Autophagy of Female Germline Stem Cells via Inhibition of the PI3K-Akt Pathway In Vitro. Cells. 2019; 8(6):606. https://doi.org/10.3390/cells8060606
Chicago/Turabian StyleLi, Xinyue, Xiaopeng Hu, Geng G. Tian, Ping Cheng, Zezhong Li, Mingyan Zhu, Huchen Zhou, and Ji Wu. 2019. "C89 Induces Autophagy of Female Germline Stem Cells via Inhibition of the PI3K-Akt Pathway In Vitro" Cells 8, no. 6: 606. https://doi.org/10.3390/cells8060606
APA StyleLi, X., Hu, X., Tian, G. G., Cheng, P., Li, Z., Zhu, M., Zhou, H., & Wu, J. (2019). C89 Induces Autophagy of Female Germline Stem Cells via Inhibition of the PI3K-Akt Pathway In Vitro. Cells, 8(6), 606. https://doi.org/10.3390/cells8060606