Regeneration of Dermis: Scarring and Cells Involved


Abstract
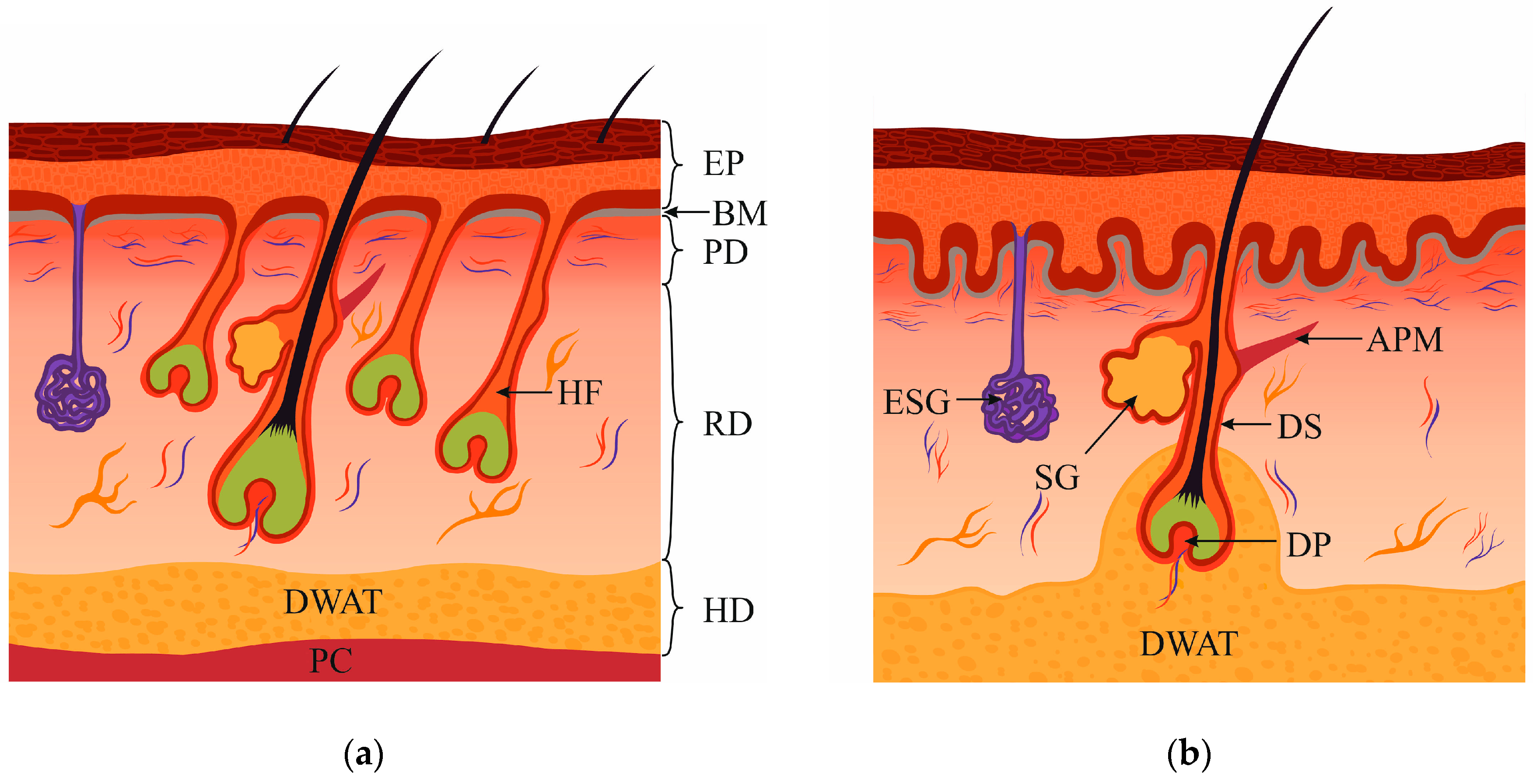
:1. Dermis Structure and Composition
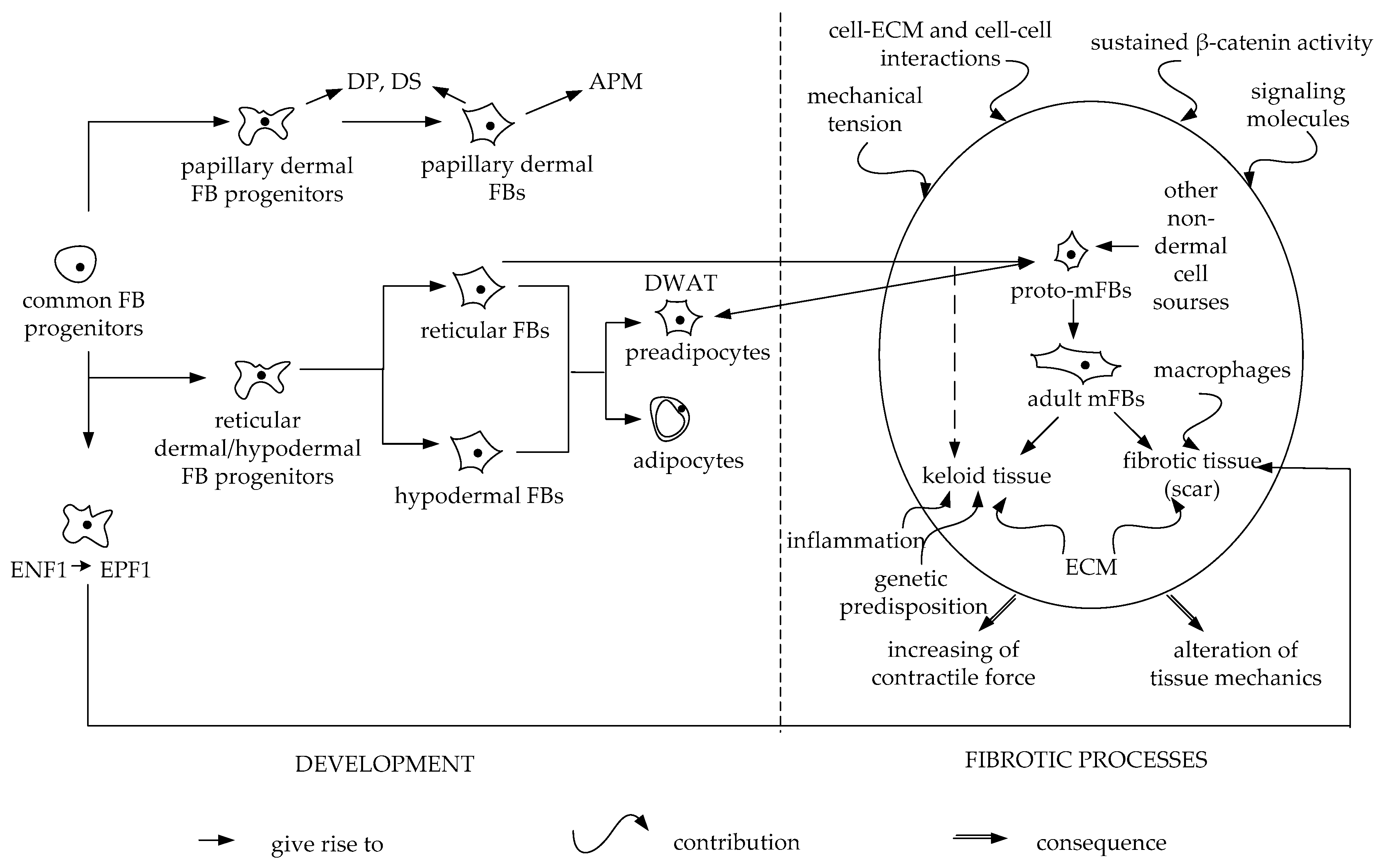
2. Papillary Fibroblasts
3. Reticular Fibroblasts
4. Fibroblasts Associated with Hair Follicles
5. Dermal Cells Participating in Different Modes of Wound Healing
6. Keloid Scars and Keloid Cells
7. Dermal White Adipose Tissue Involvement in Wound Healing
8. Myofibroblasts and Fibrosis
9. Prevention of Contraction Facilitates Scarless Regeneration
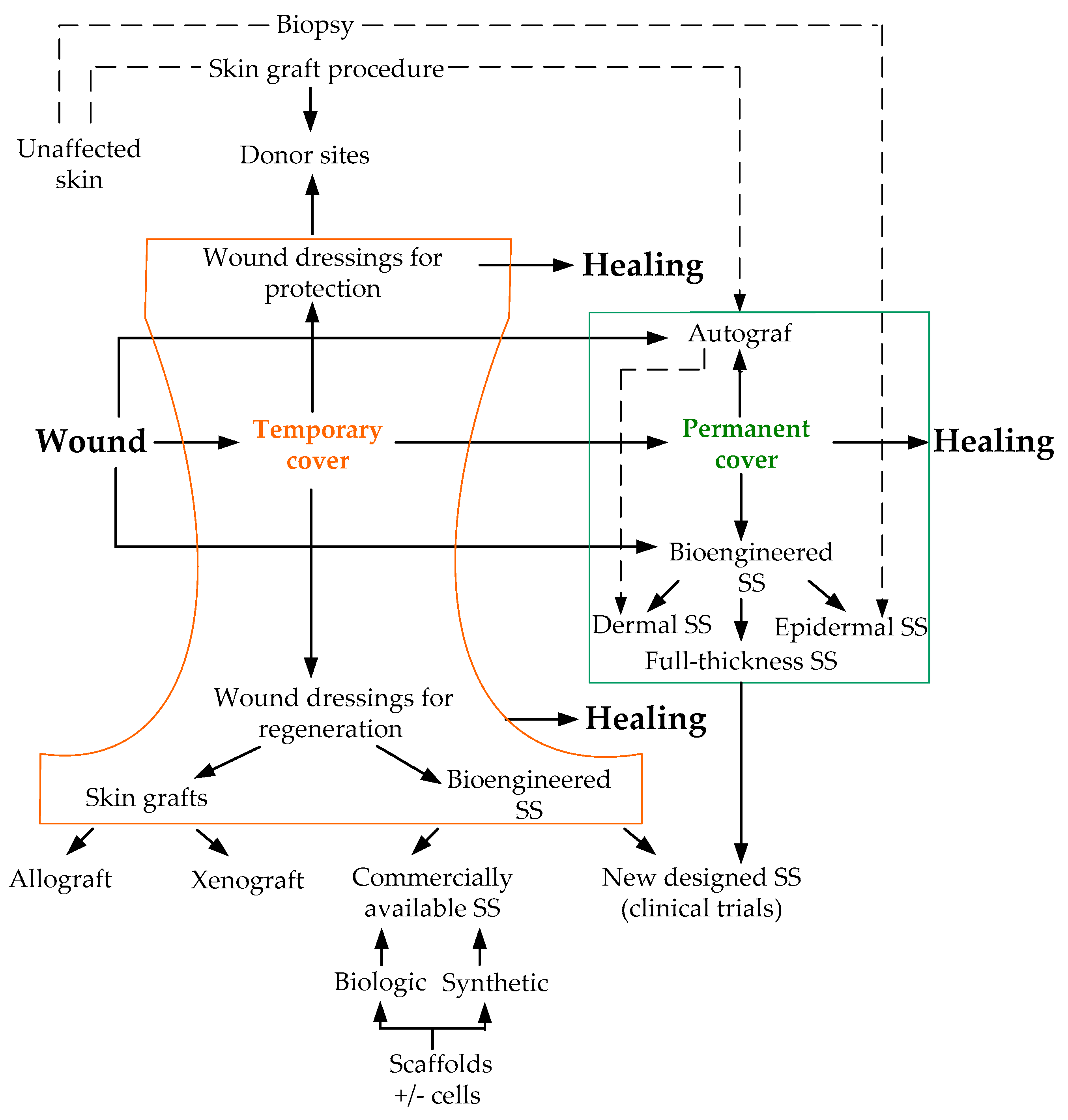
10. Skin Restoration Using Skin Substitutes
11. Conclusions
Funding
Conflicts of Interest
References
- Schulze, E.; Witt, M.; Fink, T.; Hofer, A.; Funk, R.H. Immunohistochemical detection of human skin nerve fibers. Acta Histochem. 1997, 99, 301–309. [Google Scholar] [CrossRef]
- Stücker, M.; Struk, A.; Altmeyer, P.; Herde, M.; Baumgärtl, H.; Lübbers, D.W. The cutaneous uptake of atmospheric oxygen contributes significantly to the oxygen supply of human dermis and epidermis. J. Physiol. 2002, 538, 985–994. [Google Scholar] [CrossRef] [PubMed]
- Mine, S.; Fortunel, N.O.; Pageon, H.; Asselineau, D. Aging alters functionally human dermal papillary fibroblasts but not reticular fibroblasts: A new view of skin morphogenesis and aging. PLoS ONE 2008, 3, e4066. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.M.; Melrose, J. Proteoglycans in normal and healing skin. Adv. Wound Care (New Rochelle) 2015, 4, 152–173. [Google Scholar] [CrossRef] [PubMed]
- Meigel, W.N.; Gay, S.; Weber, L. Dermal architecture and collagen type distribution. Arch. Dermatol. Res. 1977, 259, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sriram, G.; Bigliardi, P.L.; Bigliardi-Qi, M. Fibroblast heterogeneity and its implications for engineering organotypic skin models in vitro. Eur. J. Cell Biol. 2015, 94, 483–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcos-Garcés, V.; Molina Aguilar, P.; Bea Serrano, C.; García Bustos, V.; BenaventSeguí, J.; FerrándezIzquierdo, A.; Ruiz-Saurí, A. Age-related dermal collagen changes during development, maturation and ageing—a morphometric and comparative study. J. Anat. 2014, 225, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Sorrell, J.M.; Caplan, A.I. Fibroblasts—a diverse population at the center of it all. Int. Rev. Cell Mol. Biol. 2009, 276, 161–214. [Google Scholar] [CrossRef]
- Driskell, R.R.; Watt, F.M. Understanding fibroblast heterogeneity in the skin. Trends Cell Biol. 2015, 25, 92–99. [Google Scholar] [CrossRef]
- Saalbach, A.; Kraft, R.; Herrmann, K.; Haustein, U.F.; Anderegg, U. The monoclonal antibody AS02 recognizes a protein on human fibroblasts being highly homologous to Thy-1. Arch. Dermatol. Res. 1998, 290, 360–366. [Google Scholar] [CrossRef]
- Philippeos, C.; Telerman, S.B.; Oulès, B.; Pisco, A.O.; Shaw, T.J.; Elgueta, R.; Lombardi, G.; Driskell, R.R.; Soldin, M.; Lynch, M.D.; et al. Spatial and single-cell transcriptional profiling identifies functionally distinct human dermal fibroblast subpopulations. J. Investig. Dermatol. 2018, 138, 811–825. [Google Scholar] [CrossRef] [PubMed]
- Harper, R.A.; Grove, G. Human skin fibroblasts derived from papillary and reticular dermis: Differences in growth potential in vitro. Science 1979, 204, 526–527. [Google Scholar] [CrossRef] [PubMed]
- Janson, D.G.; Saintigny, G.; van Adrichem, A.; Mahé, C.; El Ghalbzouri, A. Different gene expression patterns in human papillary and reticular fibroblasts. J. Investig. Dermatol. 2012, 132, 2565–2572. [Google Scholar] [CrossRef] [PubMed]
- Driskell, R.R.; Lichtenberger, B.M.; Hoste, E.; Kretzschmar, K.; Simons, B.D.; Charalambous, M.; Ferron, S.R.; Herault, Y.; Pavlovic, G.; Ferguson-Smith, A.C.; et al. Distinct fibroblast lineages determine dermal architecture in skin development and repair. Nature 2013, 504, 277–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabib, T.; Morse, C.; Wang, T.; Chen, W.; Lafyatis, R. SFRP2/DPP4 and FMO1/LSP1 define major fibroblast populations in human skin. J. Investig. Dermatol. 2018, 138, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Korosec, A.; Frech, S.; Gesslbauer, B.; Vierhapper, M.; Radtke, C.; Petzelbauer, P.; Lichtenberger, B.M. Lineage identity and location within the dermis determine the function of papillary and reticular fibroblasts in human skin. J. Investig. Dermatol. 2019, 139, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Tajima, S.; Pinnell, S.R. Collagen synthesis by human skin fibroblasts in culture: Studies of fibroblasts explanted from papillary and reticular dermis. J. Investig. Dermatol. 1981, 77, 410–412. [Google Scholar] [CrossRef] [PubMed]
- Schönherr, E.; Beavan, L.A.; Hausser, H.; Kresse, H.; Culp, L.A. Differences in decorin expression by papillary and reticular fibroblasts in vivo and in vitro. Biochem. J. 1993, 290, 893–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.J.; Pieper, J.; Schotel, R.; van Blitterswijk, C.A.; Lamme, E.N. Stimulation of skin repair is dependent on fibroblast source and presence of extracellular matrix. Tissue Eng. 2004, 10, 1054–1064. [Google Scholar] [CrossRef]
- Pageon, H.; Zucchi, H.; Asselineau, D. Distinct and complementary roles of papillary and reticular fibroblasts in skin morphogenesis and homeostasis. Eur. J. Dermatol. 2012, 22, 324–332. [Google Scholar] [CrossRef]
- Janson, D.; Rietveld, M.; Mahé, C.; Saintigny, G.; El Ghalbzouri, A. Differential effect of extracellular matrix derived from papillary and reticular fibroblasts on epidermal development in vitro. Eur. J. Dermatol. 2017, 27, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Schafer, I.A.; Pandy, M.; Ferguson, R.; Davis, B.R. Comparative observation of fibroblasts derived from the papillary and reticular dermis of infants and adults: Growth kinetics, packing density at confluence and surface morphology. Mech. Ageing Dev. 1985, 31, 275–293. [Google Scholar] [CrossRef]
- Sorrell, J.M.; Baber, M.A.; Caplan, A.I. Site-matched papillary and reticular human dermal fibroblasts differ in their release of specific growth factors/cytokines and in their interaction with keratinocytes. J. Cell Physiol. 2004, 200, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Cho, K.H. The effects of epidermal keratinocytes and dermal fibroblasts on the formation of cutaneous basement membrane in three-dimensional culture systems. Arch. Dermatol. Res. 2005, 296, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Janson, D.; Saintigny, G.; Mahé, C.; El Ghalbzouri, A. Papillary fibroblasts differentiate into reticular fibroblasts after prolonged in vitro culture. Exp. Dermatol. 2013, 22, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Janson, D.; Saintigny, G.; Zeypveld, J.; Mahé, C.; El Ghalbzouri, A. TGF-β1 induces differentiation of papillary fibroblasts to reticular fibroblasts in monolayer culture but not in human skin equivalents. Eur. J. Dermatol. 2014, 24, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Shao, M.; Schilder, J.; Guise, T.; Mohammad, K.S.; Matei, D. Tissue transglutaminase links TGF-beta, epithelial to mesenchymal transition and a stem cell phenotype in ovarian cancer. Oncogene 2012, 31, 2521–2534. [Google Scholar] [CrossRef]
- Wang, J.; Dodd, C.; Shankowsky, H.A.; Scott, P.G.; Tredget, E.E.; Wound Healing Research Group. Deep dermal fibroblasts contribute to hypertrophic scarring. Lab. Investig. 2008, 88, 1278–1290. [Google Scholar] [CrossRef] [Green Version]
- Rinkevich, Y.; Walmsley, G.G.; Hu, M.S.; Maan, Z.N.; Newman, A.M.; Drukker, M.; Januszyk, M.; Krampitz, G.W.; Gurtner, G.C.; Lorenz, H.P.; et al. Skin fibrosis. Identification and isolation of a dermal lineage with intrinsic fibrogenic potential. Science 2015, 348, aaa2151. [Google Scholar] [CrossRef]
- Woodley, D.T. Distinct fibroblasts in the papillary and reticular dermis: Implications for wound healing. Dermatol. Clin. 2017, 35, 95–100. [Google Scholar] [CrossRef]
- Jahoda, C.A.; Whitehouse, J.; Reynolds, A.J.; Hole, N. Hair follicle dermal cells differentiate into adipogenic and osteogenic lineages. Exp. Dermatol. 2003, 12, 849–859. [Google Scholar] [CrossRef] [PubMed]
- Hoogduijn, M.J.; Gorjup, E.; Genever, P.G. Comparative characterization of hair follicle dermal stem cells and bone marrow mesenchymal stem cells. Stem Cells Dev. 2006, 15, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Chermnykh, E.S.; Vorotelyak, E.A.; Gnedeva, K.Y.; Moldaver, M.V.; Yegorov, Y.E.; Vasiliev, A.V.; Terskikh, V.V. Dermal papilla cells induce keratinocyte tubulogenesis in culture. Histochem. Cell Biol. 2010, 133, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Shin, T.H.; Kim, H.S.; Choi, S.W.; Kang, K.S. Mesenchymal stem cell therapy for inflammatory skin diseases: Clinical potential and mode of action. Int. J. Mol. Sci. 2017, 18, 244. [Google Scholar] [CrossRef] [PubMed]
- Meleshina, A.V.; Rogovaya, O.S.; Dudenkova, V.V.; Sirotkina, M.A.; Lukina, M.M.; Bystrova, A.S.; Krut, V.G.; Kuznetsova, D.S.; Kalabusheva, E.P.; Vasiliev, A.V.; et al. Multimodal label-free imaging of living dermal equivalents including dermal papilla cells. Stem Cell Res. Ther. 2018, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.H.; Liu, P.; Xie, J.L.; Shu, B.; Xu, Y.B.; Ke, C.N.; Liu, X.S.; Li, T.Z. Experimental study on repairing of nude mice skin defects with composite skin consisting of xenogeneic dermis and epidermal stem cells and hair follicle dermal papilla cells. Burns 2008, 34, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Higgins, C.A.; Roger, M.F.; Hill, R.P.; Ali-Khan, A.S.; Garlick, J.A.; Christiano, A.M.; Jahoda, C.A.B. Multifaceted role of hair follicle dermal cells in bioengineered skins. Br. J. Dermatol. 2017, 176, 1259–1269. [Google Scholar] [CrossRef]
- Ma, D.; Kua, J.E.; Lim, W.K.; Lee, S.T.; Chua, A.W. In vitro characterization of human hair follicle dermal sheath mesenchymal stromal cells and their potential in enhancing diabetic wound healing. Cytotherapy 2015, 17, 1036–1051. [Google Scholar] [CrossRef]
- McElwee, K.J.; Kissling, S.; Wenzel, E.; Huth, A.; Hoffmann, R. Cultured peribulbar dermal sheath cells can induce hair follicle development and contribute to the dermal sheath and dermal papilla. J. Investig. Dermatol. 2003, 121, 1267–1275. [Google Scholar] [CrossRef]
- Osada, A.; Iwabuchi, T.; Kishimoto, J.; Hamazaki, T.S.; Okochi, H. Long-term culture of mouse vibrissal dermal papilla cells and de novo hair follicle induction. Tissue Eng. 2007, 13, 975–982. [Google Scholar] [CrossRef]
- Rahmani, W.; Abbasi, S.; Hagner, A.; Raharjo, E.; Kumar, R.; Hotta, A.; Magness, S.; Metzger, D.; Biernaskie, J. Hair follicle dermal stem cells regenerate the dermal sheath.; repopulate the dermal papilla, and modulate hair type. Dev. Cell 2014, 31, 543–558. [Google Scholar] [CrossRef] [PubMed]
- Havlickova, B.; Bíró, T.; Mescalchin, A.; Tschirschmann, M.; Mollenkopf, H.; Bettermann, A.; Pertile, P.; Lauster, R.; Bodó, E.; Paus, R. A human folliculoid microsphere assay for exploring epithelial- mesenchymal interactions in the human hair follicle. J. Investig. Dermatol. 2009, 129, 972–983. [Google Scholar] [CrossRef] [PubMed]
- Yen, C.M.; Chan, C.C.; Lin, S.J. High-throughput reconstitution of epithelial-mesenchymal interaction in folliculoid microtissues by biomaterial-facilitated self-assembly of dissociated heterotypic adult cells. Biomaterials 2010, 31, 4341–4352. [Google Scholar] [CrossRef] [PubMed]
- Higgins, C.A.; Chen, J.C.; Cerise, J.E.; Jahoda, C.A.; Christiano, A.M. Microenvironmental reprogramming by three-dimensional culture enables dermal papilla cells to induce de novo human hair-follicle growth. Proc. Natl. Acad Sci. USA 2013, 110, 19679–19688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalabusheva, E.; Terskikh, V.; Vorotelyak, E. Hair germ model in vitro via human postnatal keratinocyte-dermal papilla interactions: Impact of hyaluronic acid. Stem Cells Int. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Leirós, G.J.; Kusinsky, A.G.; Drago, H.; Bossi, S.; Sturla, F.; Castellanos, M.L.; Stella, I.Y.; Balañá, M.E. Dermal papilla cells improve the wound healing process and generate hair bud-like structures in grafted skin substitutes using hair follicle stem cells. Stem Cells Transl. Med. 2014, 3, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Thangapazham, R.L.; Klover, P.; Wang, J.A.; Zheng, Y.; Devine, A.; Li, S.; Sperling, L.; Cotsarelis, G.; Darling, T.N. Dissociated human dermal papilla cells induce hair follicle neogenesis in grafted dermal-epidermal composites. J. Investig. Dermatol. 2014, 134, 538–540. [Google Scholar] [CrossRef] [PubMed]
- Abaci, H.E.; Coffman, A.; Doucet, Y.; Chen, J.; Jacków, J.; Wang, E.; Guo, Z.; Shin, J.U.; Jahoda, C.A.; Christiano, A.M. Tissue engineering of human hair follicles using a biomimetic developmental approach. Nat. Commun. 2018, 9, 5301. [Google Scholar] [CrossRef] [PubMed]
- Plikus, M.V.; Guerrero-Juarez, C.F.; Ito, M.; Li, Y.R.; Dedhia, P.H.; Zheng, Y.; Shao, M.; Gay, D.L.; Ramos, R.; His, T.C.; et al. Regeneration of fat cells from myofibroblasts during wound healing. Science 2017, 355, 748–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, C.H.; Sun, Q.; Ratti, K.; Lee, S.H.; Zheng, Y.; Takeo, M.; Lee, W.; Rabbani, P.; Plikus, M.V.; Cain, J.E.; et al. Hedgehog stimulates hair follicle neogenesis by creating inductive dermis during murine skin wound healing. Nat. Commun. 2018, 9, 4903. [Google Scholar] [CrossRef] [PubMed]
- Mecklenburg, L.; Tobin, D.J.; Müller-Röver, S.; Handjiski, B.; Wendt, G.; Peters, E.M.; Pohl, S.; Moll, I.; Paus, R. Active hair growth (anagen) is associated with angiogenesis. J. Investig. Dermatol. 2000, 114, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.J.; Jing, J.; Lu, Z.F.; Zheng, M. Expression and localization of VEGFR-2 in hair follicles during induced hair growth in mice. Arch. Dermatol. Res. 2018, 310, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Fan, W.; Li, M. Angiogenin is expressed in human dermal papilla cells and stimulates hair growth. Arch. Dermatol. Res. 2009, 301, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Bassino, E.; Gasparri, F.; Giannini, V.; Munaron, L. Paracrine crosstalk between human hair follicle dermal papilla cells and microvascular endothelial cells. Exp. Dermatol. 2015, 24, 388–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huh, S.H.; Närhi, K.; Lindfors, P.H.; Häärä, O.; Yang, L.; Ornitz, D.M.; Mikkola, M.L. Fgf20 governs formation of primary and secondary dermal condensations in developing hair follicles. Genes Dev. 2013, 27, 450–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biggs, L.C.; Mäkelä, O.J.; Myllymäki, S.M.; Das Roy, R.; Närhi, K.; Pispa, J.; Mustonen, T.; Mikkola, M.L. Hair follicle dermal condensation forms via Fgf20 primed cell cycle exit, cell motility, and aggregation. Elife 2018, 7, e36468. [Google Scholar] [CrossRef] [PubMed]
- Woo, W.M.; Zhen, H.H.; Oro, A.E. Shh maintains dermal papilla identity and hair morphogenesis via a Noggin-Shh regulatory loop. Genes Dev. 2012, 26, 1235–1246. [Google Scholar] [CrossRef] [PubMed]
- Lichtenberger, B.M.; Mastrogiannaki, M.; Watt, F.M. Epidermal β-catenin activation remodels the dermis via paracrine signalling to distinct fibroblast lineages. Nat. Commun. 2016, 7, 10537. [Google Scholar] [CrossRef]
- Kaushal, G.S.; Rognoni, E.; Lichtenberger, B.M.; Driskell, R.R.; Kretzschmar, K.; Hoste, E.; Watt, F.M. Fate of prominin-1 expressing dermal papilla cells during homeostasis, wound healing and Wnt activation. J. Investig. Dermatol. 2015, 135, 2926–2934. [Google Scholar] [CrossRef]
- Sorrell, J.M.; Caplan, A.I. Fibroblast heterogeneity: More than skin deep. J. Cell Sci. 2004, 117, 667–675. [Google Scholar] [CrossRef]
- Sorrell, J.M.; Baber, M.A.; Caplan, A.I. Clonal characterization of fibroblasts in the superficial layer of the adult human dermis. Cell Tissue Res. 2007, 327, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Lo Celso, C.; Prowse, D.M.; Watt, F.M. Transient activation of beta-catenin signalling in adult mouse epidermis is sufficient to induce new hair follicles but continuous activation is required to maintain hair follicle tumours. Development 2004, 131, 1787–1799. [Google Scholar] [CrossRef] [PubMed]
- Silva-Vargas, V.; Lo Celso, C.; Giangreco, A.; Ofstad, T.; Prowse, D.M.; Braun, K.M.; Watt, F.M. Beta-catenin and Hedgehog signal strength can specify number and location of hair follicles in adult epidermis without recruitment of bulge stem cells. Dev. Cell 2005, 9, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Yang, Z.; Andl, T.; Cui, C.; Kim, N.; Millar, SE.; Cotsarelis, G. Wnt-dependent de novo hair follicle regeneration in adult mouse skin after wounding. Nature 2007, 447, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Deschene, E.R.; Myung, P.; Rompolas, P.; Zito, G.; Sun, T.Y.; Taketo, M.M.; Saotome, I.; Greco, V. β-Catenin activation regulates tissue growth non-cell autonomously in the hair stem cell niche. Science 2014, 343, 1353–1356. [Google Scholar] [CrossRef] [PubMed]
- Lam, A.P.; Gottardi, C.J. β-catenin signaling: A novel mediator of fibrosis and potential therapeutic target. Curr Opin Rheumatol 2011, 23, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Beyer, C.; Schramm, A.; Akhmetshina, A.; Dees, C.; Kireva, T.; Gelse, K.; Sonnylal, S.; de Crombrugghe, B.; Taketo, M.M.; Distler, O.; et al. β-catenin is a central mediator of pro-fibrotic Wnt signaling in systemic sclerosis. Ann. Rheum. Dis. 2012, 71, 761–767. [Google Scholar] [CrossRef]
- Hamburg, E.J.; Atit, R.P. Sustained β-catenin activity in dermal fibroblasts is sufficient for skin fibrosis. J. Investig. Dermatol. 2012, 132, 2469–2472. [Google Scholar] [CrossRef]
- Hamburg-Shields, E.; DiNuoscio, GJ.; Mullin, N.K.; Lafyatis, R.; Atit, R.P. Sustained β-catenin activity in dermal fibroblasts promotes fibrosis by up-regulating expression of extracellular matrix protein-coding genes. J. Pathol. 2015, 235, 686–697. [Google Scholar] [CrossRef]
- Mastrogiannaki, M.; Lichtenberger, B.M.; Reimer, A.; Collins, C.A.; Driskell, R.R.; Watt, F.M. β-catenin stabilization in skin fibroblasts causes fibrotic lesions by preventing adipocyte differentiation of the reticular dermis. J. Investig. Dermatol. 2016, 136, 1130–1142. [Google Scholar] [CrossRef]
- Rognoni, E.; Gomez, C.; Pisco, A.O.; Rawlins, E.L.; Simons, B.D.; Watt, F.M.; Driskell, R.R. Inhibition of β-catenin signalling in dermal fibroblasts enhances hair follicle regeneration during wound healing. Development 2016, 143, 2522–2535. [Google Scholar] [CrossRef] [PubMed]
- Longaker, M.T.; Adzick, N.S. The biology of fetal wound healing: A review. Plast. Reconstr. Surg. 1991, 87, 788–790. [Google Scholar] [CrossRef] [PubMed]
- Kishi, K.; Okabe, K.; Shimizu, R.; Kubota, Y. Fetal skin possesses the ability to regenerate completely: Complete regeneration of skin. Keio J. Med. 2012, 61, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Cass, D.L.; Bullard, K.M.; Sylvester, K.G.; Yang, E.Y.; Longaker, M.T.; Adzick, N.S. Wound size and gestational age modulate scar formation in fetal wound repair. J. Pediatr. Surg. 1997, 32, 411–415. [Google Scholar] [CrossRef]
- Bullard, K.M.; Longaker, M.T.; Lorenz, H.P. Fetal wound healing: Current biology. World J. Surg. 2003, 27, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.S.; Borrelli, M.R.; Hong, W.X.; Malhotra, S.; Cheung, A.T.M.; Ransom, R.C.; Rennert, R.C.; Morrison, S.D.; Lorenz, H.P.; Longaker, M.T. Embryonic skin development and repair. Organogenesis 2018, 14, 46–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, M.; Peled, Z.M.; Chin, G.S.; Liu, W.; Longaker, M.T. Ontogeny of expression of transforming growth factor-β 1 (TGF-β 1), TGF-β 3, and TGF-β receptors I and II in fetal rat fibroblasts and skin. Plast. Reconstr. Surg. 2001, 107, 1787–1794. [Google Scholar] [CrossRef] [PubMed]
- Carre, A.L.; James, A.W.; MacLeod, L.; Kong, W.; Kawai, K.; Longaker, MT.; Lorenz, H.P. Interaction of wingless protein (Wnt), transforming growth factor-β1, and hyaluronan production in fetal and postnatal fibroblasts. Plast. Reconstr. Surg. 2010, 125, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Armatas, A.A.; Pratsinis, H.; Mavrogonatou, E.; Angelopoulou, M.T.; Kouroumalis, A.; Karamanos, N.K.; Kletsas, D. The differential proliferative response of fetal and adult human skin fibroblasts to TGF-β is retained when cultured in the presence of fibronectin or collagen. Biochim. Biophys. Acta 2014, 1840, 2635–2642. [Google Scholar] [CrossRef] [PubMed]
- Rolfe, K.J.; Irvine, L.M.; Grobbelaar, A.O.; Linge, C. Differential gene expression in response to transforming growth factor-β1 by fetal and postnatal dermal fibroblasts. Wound Repair Regen. 2007, 15, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, M.W.; O’Kane, S. Scar-free healing: From embryonic mechanisms to adult therapeutic intervention. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Bush, J.; So, K.; Mason, T.; Occleston, N.L.; O’Kane, S.; Ferguson, M.W.J. Therapies with emerging evidence of efficacy: Avotermin for the improvement of scarring. Dermatol. Res. Pract. 2010, 2010, 690613. [Google Scholar] [CrossRef] [PubMed]
- So, K.; McGrouther, D.A.; Bush, J.A.; Durani, P.; Taylor, L.; Skotny, G.; Mason, T.; Metcalfe, A.D.; O’Kane, S.; Ferguson, M.W.J. Avotermin for scar improvement following scar revision surgery: A randomized, double-blind, within-patient, placebo-controlled, phase II clinical trial. Plast. Reconstr. Surg. 2011, 128, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Little, J.A.; Murdy, R.; Cossar, N.; Getliffe, K.M.; Hanak, J.; Ferguson, M.W. TGF β3 immunoassay standardization: Comparison of NIBSC reference preparation code 98/608 with avotermin lot 205-0505-005. J. Immunoass. Immunochem. 2012, 33, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Jackson, C.J. Extracellular matrix reorganization during wound healing and its impact on abnormal scarring. Adv. Wound Care (New Rochelle) 2015, 4, 119–136. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.Y.; Grant, M.E.; Schor, A.M. Differences between adult and foetal fibroblasts in the regulation of hyaluronate synthesis: Correlation with migratoryactivity. J. Cell Sci. 1989, 94, 577–584. [Google Scholar] [PubMed]
- Motolese, A.; Vignati, F.; Brambilla, R.; Cerati, M.; Passi, A. Interaction between a regenerative matrix and wound bed in nonhealing ulcers: Results with 16 cases. Biomed. Res. Int. 2013, 2013, 849321. [Google Scholar] [CrossRef] [PubMed]
- Frenkel, J.S. The role of hyaluronan in wound healing. Int. Wound J. 2014, 11, 159–163. [Google Scholar] [CrossRef]
- Longinotti, C. The use of hyaluronic acid based dressings to treat burns: A review. Burns Trauma 2014, 2, 162–168. [Google Scholar] [CrossRef] [Green Version]
- Hong, L.; Shen, M.; Fang, J.; Wang, Y.; Bao, Z.; Bu, S.; Zhu, Y. Hyaluronic acid (HA)—Based hydrogels for full-thickness wound repairing and skin regeneration. J. Mater. Sci. Mater. Med. 2018, 29, 150. [Google Scholar] [CrossRef]
- Yates, C.C.; Hebda, P.; Wells, A. Skin wound healing and scarring: Fetal wounds and regenerative restitution. Birth Defects Res. C Embryo Today 2012, 96, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Parekh, A.; Hebda, P.A. The contractile phenotype of dermal fetal fibroblasts in scarless wound healing. Curr. Pathobiol. Rep. 2017, 5, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Jerrell, R.J.; Leih, M.J.; Parekh, A. The altered mechanical phenotype of fetal fibroblasts hinders myofibroblast differentiation. Wound Repair Regen. 2019, 27, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.M.; Tsai, C.F.; Yen, C.M.; Lin, M.H.; Wang, W.H.; Chan, C.C.; Chen, C.L.; Phua, K.K.L.; Pan, S.H.; Plikus, M.V.; et al. Inducing hair follicle neogenesis with secreted proteins enriched in embryonic skin. Biomaterials 2018, 167, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Correa-Gallegos, D.; Christ, S.; Stefanska, A.; Liu, J.; Ramesh, P.; Rajendran, V.; De Santis, M.M.; Wagner, D.E.; Rinkevich, Y. Two succeeding fibroblastic lineages drive dermal development and the transition from regeneration to scarring. Nat. Cell Biol. 2018, 20, 422–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sgaier, S.K.; Lao, Z.; Villanueva, M.P.; Berenshteyn, F.; Stephen, D.; Turnbull, R.K.; Joyner, A.L. Genetic subdivision of the tectum and cerebellum into functionally related regions based on differential sensitivity to engrailed proteins. Development 2007, 134, 2325–2335. [Google Scholar] [CrossRef] [Green Version]
- Bayat, A.; McGrouther, D.A.; Ferguson, M.W.J. Skin Scarring. BMJ 2003, 326, 88–92. [Google Scholar] [CrossRef]
- Bran, G.M.; Goessler, U.R.; Hormann, K.; Riedel, F.; Sadick, H. Keloids: Current concepts of pathogenesis (review). Int. J. Mol. Med. 2009, 24, 283–293. [Google Scholar] [CrossRef]
- Ogawa, R.; Akaishi, S.; Huang, C.; Dohi, T.; Aoki, M.; Omori, Y.; Koike, S.; Kobe, K.; Akimoto, M.; Hyakusoku, H. Clinical applications of basic research that shows reducing skin tension could prevent and treat abnormal scarring: The importance of fascial/subcutaneous tensile reduction sutures and flap surgery for keloid and hypertrophic scar reconstruction. J. Nippon. Med. Sch. 2011, 78, 68–76. [Google Scholar] [CrossRef]
- Agha, R.; Ogawa, R.; Pietramaggiori, G.; Orgill, D.P. A review of the role of mechanical forces in cutaneous wound healing. J. Surg. Res. 2011, 171, 700–708. [Google Scholar] [CrossRef]
- Atiyeh, B.S.; Costagliola, M.; Hayek, S.N. Keloid or hypertrophic scar: The controversy: Review of the literature. Ann. Plast. Surg. 2005, 54, 676–680. [Google Scholar] [CrossRef] [PubMed]
- Butler, P.D.; Longaker, M.T.; Yang, G.P. Current progress in keloid research and treatment. J. Am. Coll. Surg. 2008, 206, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Berman, B.; Maderal, A.; Raphael, B. Keloids and Hypertrophic Scars: Pathophysiology, Classification, and Treatment. Dermatol. Surg. 2017, 43 (Suppl. 1), S3–S18. [Google Scholar] [CrossRef] [PubMed]
- Seo, B.F.; Lee, J.Y.; Jung, S.N. Models of abnormal scarring. Bio. Med. Res. Int. 2013, 2013, 423147. [Google Scholar] [CrossRef]
- Kischer, C.W.; Pindur, J.; Shetlar, M.R.; Shetlar, C.L. Implants of hypertrophic scars and keloids into the nude (athymic) mouse: Viability and morphology. J. Trauma 1989, 29, 672–677. [Google Scholar] [CrossRef] [PubMed]
- Shetlar, M.R.; Shetlar, C.L.; Kischer, C.W.; Pindur, J. Implants of keloid and hypertrophic scars into the athymic nude mouse: Changes in the glycosaminoglycans of the implants. Connect. Tissue Res. 1991, 26, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Philandrianos, C.; Gonnelli, D.; Andrac-Meyer, L.; Bruno, M.; Magalon, G.; Mordon, S. Establishment of a keloid model by transplanting human keloid onto the backs of nude mice. Ann. Chir. Plast. Esthet. 2014, 59, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.M.; Hoffren, J.; Rouda, S.; Greenbaum, S.; Fox, J.W., 4th; Moore, J.H., Jr.; Dodge, G.R. Decorin, versican, and biglycan gene expression by keloid and normal dermal fibroblasts: Differential regulation by basic fibroblast growth factor. Exp. Cell Res. 1993, 209, 200–207. [Google Scholar] [CrossRef]
- Chipev, C.C.; Simman, R.; Hatch, G.; Katz, AE.; Siegel, DM.; Simon, M. Myofibroblast phenotype and apoptosis in keloid and palmar fibroblasts in vitro. Cell Death Differ. 2000, 7, 166–176. [Google Scholar] [CrossRef]
- Phan, T.T.; Lim, I.J.; Bay, B.H.; Qi, R.; Huynh, H.T.; Lee, S.T.; Longaker, M.T. Differences in collagen production between normal and keloid-derived fibroblasts in serum-media co-culture with keloid-derived keratinocytes. J. Dermatol. Sci. 2002, 29, 26–34. [Google Scholar] [CrossRef]
- Shih, B.; Garside, E.; McGrouther, D.A.; Bayat, A. Molecular dissection of abnormal wound healing processes resulting in keloid disease. Wound Repair Regen. 2010, 18, 139–153. [Google Scholar] [CrossRef]
- Kamamoto, F.; Paggiaro, A.O.; Rodas, A.; Herson, M.R.; Mathor, M.B.; Ferreira, M.C. A wound contraction experimental model for studying keloids and wound-healing modulators. Artif. Organs 2003, 27, 701–705. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T.; Nakao, A.; Sumiyoshi, K.; Tsuchihashi, H.; Ogawa, H. SB-431542 inhibits TGF-beta-induced contraction of collagen gel by normal and keloid fibroblasts. J. Dermatol. Sci. 2005, 39, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Yamazaki, M.; Maeda, T.; Matsumura, H.; Setoguchi, Y.; Tsuboi, R. Pirfenidone suppresses keloid fibroblast-embedded collagen gel contraction. Arch. Dermatol. Res. 2012, 304, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Tucci-Viegas, V.M.; Hochman, B.; França, J.P.; Ferreira, L.M. Keloid explant culture: A model for keloid fibroblasts isolation and cultivation based on the biological differences of its specific regions. Int. Wound J. 2010, 7, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Syed, F.; Ahmadi, E.; Iqbal, S.A.; Singh, S.; McGrouther, D.A.; Bayat, A. Fibroblasts from the growing margin of keloid scars produce higher levels of collagen I and III compared with intralesional and extralesional sites: Clinical implications for lesional site-directed therapy. Br. J. Dermatol 2011, 164, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Suttho, D.; Mankhetkorn, S.; Binda, D.; Pazart, L.; Humbert, P.; Rolin, G. 3D modeling of keloid scars in vitro by cell and tissue engineering. Arch. Dermatol. Res. 2017, 309, 55–62. [Google Scholar] [CrossRef]
- Jiao, H.; Zhang, T.; Fan, J.; Xiao, R. The Superficial dermis may initiate keloid formation: Histological analysis of the keloid dermis at different depths. Front. Physiol. 2017, 8, 885. [Google Scholar] [CrossRef]
- Yan, L.; Cao, R.; Wang, L.; Liu, Y.; Pan, B.; Yin, Y.; Lv, X.; Zhuang, Q.; Sun, X.; Xiao, R. Epithelial-mesenchymal transition in keloid tissues and TGF-β1-induced hair follicle outer root sheath keratinocytes. Wound Repair Regen. 2015, 23, 601–610. [Google Scholar] [CrossRef]
- Yan, C.; Grimm, W.A.; Garner, W.L.; Qin, L.; Qin, L.; Travis, T.; Tan, N.; Han, Y.P. Epithelial to mesenchymal transition in human skin wound healing is induced by tumor necrosis factor-alpha through bone morphogenic protein-2. Am. J. Pathol. 2010, 176, 2247–2258. [Google Scholar] [CrossRef]
- Ma, X.; Chen, J.; Xu, B.; Long, X.; Qin, H.; Zhao, R.C.; Wang, X. Keloid-derived keratinocytes acquire a fibroblast-like appearance and an enhanced invasive capacity in a hypoxic microenvironment in vitro. Int. J. Mol. Med. 2015, 35, 1246–1256. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, H.; Tosa, M.; Egawa, S.; Murakami, M.; Mohammad, G.; Ogawa, R. Examination of epithelial mesenchymal transition in keloid tissues and possibility of keloid therapy target. PRS Global Open 2016, 4, e1138. [Google Scholar] [CrossRef] [PubMed]
- Stone, R.C.; Pastar, I.; Ojeh, N.; Chen, V.; Liu, S.; Garzon, K.I.; Tomic-Canic, M. Epithelial-mesenchymal transition in tissue repair and fibrosis. Cell Tissue Res. 2016, 365, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.Y.; Yu, Q.; Cheng, T.; Liao, T.; Nie, C.L.; Wang, A.Y.; Zheng, X.; Xie, X.G.; Albers, A.E.; Gao, W.Y. Role of caveolin-1 in the pathogenesis of tissue fibrosis by keloid-derived fibroblasts in vitro. Br. J. Dermatol. 2011, 164, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.K.; Lin, H.H.; Harn, H.I.; Ogawa, R.; Wang, Y.K.; Ho, Y.T.; Chen, W.R.; Lee, Y.C.; Lee, J.Y.; Shieh, S.J.; et al. Caveolin-1 controls hyperresponsiveness to mechanical stimuli and fibrogenesis-associated RUNX2 activation in keloid fibroblasts. J. Investig. Dermatol. 2018, 138, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Naitoh, M.; Kubota, H.; Ikeda, M.; Tanaka, T.; Shirane, H.; Suzuki, S.; Nagata, K. Gene expression in human keloids is altered from dermal to chondrocytic and osteogenic lineage. Genes Cells 2005, 10, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Seifert, O.; Bayat, A.; Geffers, R.; Dienus, K.; Buer, J.; Löfgren, S.; Matussek, A. Identification of unique gene expression patterns within different lesional sites of keloids. Wound Repair Regen. 2008, 16, 254–265. [Google Scholar] [CrossRef]
- Raglow, Z.; Thomas, S.M. Tumor matrix protein collagen XI alpha1 in cancer. Cancer Lett. 2015, 357, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Villa, F.; Garcia-Ocana, M.; Galvan, J.A.; Garcia-Martinez, J.; Garcia-Pravia, C.; Menendez-Rodriguez, P.; González-del Rey, C.; Barneo-Serra, L.; de Los Toyos, J.R. COL11A1/(pro)collagen 11A1 expression is a remarkable biomarker of human invasive carcinoma-associated stromal cells and carcinoma progression. Tumour Biol. 2015, 36, 2213–2222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, R. Keloid and hypertrophic scars are the result of chronic inflammation in the reticular dermis. Int. J. Mol. Sci. 2017, 18, 606. [Google Scholar] [CrossRef]
- Kruglikov, I.L.; Scherer, P.E. Dermal adipocytes: From irrelevance to metabolic targets? Trends Endocrinol. Metab. 2016, 27, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Festa, E.; Fretz, J.; Berry, R.; Schmidt, B.; Rodeheffer, M.; Horowitz, M.; Horsley, V. Adipocyte lineage cells contribute to the skin stem cell niche to drive hair cycling. Cell 2011, 146, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, H.; Engrav, L.H.; Gibran, N.S.; Yang, T.M.; Grant, J.H.; Yunusov, M.Y.; Fang, P.; Reichenbach, D.D.; Heimbach, D.M.; Isik, F.F. Cones of skin occur where hypertrophic scar occurs. Wound Repair Regen. 2001, 9, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.R.; Nicu, C.; Schneider, M.R.; Hinde, E.; Paus, R. Dermal white adipose tissue undergoes major morphological changes during the spontaneous and induced murine hair follicle cycling: A reappraisal. Arch. Dermatol. Res. 2018, 310, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, B.A.; Horsley, V. Intradermal adipocytes mediate fibroblast recruitment during skin wound healing. Development 2013, 140, 1517–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moitra, J.; Mason, M.M.; Olive, M.; Krylov, D.; Gavrilova, O.; Marcus-Samuels, B.; Feigenbaum, L.; Lee, E.; Aoyama, T.; Eckhaus, M.; et al. Life without white fat: A transgenic mouse. Genes Dev. 1998, 12, 3168–3181. [Google Scholar] [CrossRef] [PubMed]
- Marangoni, R.G.; Korman, B.D.; Wei, J.; Wood, T.A.; Graham, L.V.; Whitfield, M.L.; Scherer, P.E.; Tourtellotte, W.G.; Varga, J. Myofibroblasts in murine cutaneous fibrosis originate from adiponectin-positive intradermal progenitors. Arthr. Rheumatol. 2015, 67, 1062–1073. [Google Scholar] [CrossRef]
- Gabbiani, G.; Ryan, G.B.; Majno, G. Presence of modified fibroblasts in granulation tissue and their possible role in wound contraction. Experientia 1971, 27, 549–550. [Google Scholar] [CrossRef]
- Hinz, B. The myofibroblast: Paradigm for a mechanically active cell. J. Biomech. 2010, 43, 146–155. [Google Scholar] [CrossRef]
- Klingberg, F.; Hinz, B.; White, E.S. The myofibroblast matrix: Implications for tissue repair and fibrosis. J. Pathol. 2013, 229, 298–309. [Google Scholar] [CrossRef]
- Albeiroti, S.; Soroosh, A.; de la Motte, C.A. Hyaluronan’s Role in Fibrosis: A Pathogenic Factor or a Passive Player? Biomed. Res. Int. 2015, 2015, 790203. [Google Scholar] [CrossRef] [PubMed]
- Evanko, SP.; Potter-Perigo, S.; Petty, L.J.; Workman, G.A.; Wight, T.N. Hyaluronan Controls the Deposition of Fibronectin and Collagen and Modulates TGF-β1 Induction of Lung Myofibroblasts. Matrix Biol. 2015, 42, 74–92. [Google Scholar] [CrossRef] [PubMed]
- Serini, G.; Bochaton-Piallat, M.L.; Ropraz, P.; Geinoz, A.; Borsi, L.; Zardi, L.; Gabbiani, G. The fibronectin domain ED-A is crucial for myofibroblastic phenotype induction by transforming growth factor-beta1. J. Cell Biol. 1998, 142, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Klingberg, F.; Chau, G.; Walraven, M.; Boo, S.; Koehler, A.; Chow, M.L.; Olsen, A.L.; Im, M.; Lodyga, M.; Wells, R.G.; et al. The fibronectin ED-A domain enhances recruitment of latent TGF-β-binding protein-1 to the fibroblast matrix. J. Cell Sci. 2018, 131, 201293. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B.; Gabbiani, G.; Seemayer, T.A.; Schürch, W. Myofibroblast. In Histology for Pathologists, 4th ed.; Mills, S.E., Ed.; Lippincott-Williams & Wilkins: Philadelphia, PA, USA, 2012; pp. 131–177. [Google Scholar]
- Desmoulière, A.; Redard, M.; Darby, I.; Gabbiani, G. Apoptosis mediates the decrease in cellularity during the transition between granulation tissue and scar. Am. J. Pathol. 1995, 146, 56–66. [Google Scholar] [PubMed]
- Hecker, L.; Jagirdar, R.; Jin, T.; Thannickal, V.J. Reversible differentiation of myofibroblasts by MyoD. Exp. Cell Res. 2011, 317, 1914–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kisseleva, T.; Cong, M.; Paik, Y.; Scholten, D.; Jiang, C.; Benner, C.; Iwaisako, K.; Moore Morris, T.; Scott, B.; Tsukamoto, H.; et al. Myofibroblasts revert to an inactive phenotype during regression of liver fibrosis. Proc. Natl. Acad. Sci. USA 2012, 109, 9448–9453. [Google Scholar] [CrossRef] [Green Version]
- Troeger, J.S.; Mederacke, I.; Gwak, G.Y.; Dapito, D.H.; Mu, X.; Hsu, C.C.; Pradere, J.P.; Friedman, R.A.; Schwabe, R.F. Deactivation of hepatic stellate cells during liver fibrosis resolution in mice. Gastroenterology 2012, 143, 1073–1083.e22. [Google Scholar] [CrossRef]
- Granato, G.; Ruocco, M.R.; Iaccarino, A.; Masone, S.; Calì, G.; Avagliano, A.; Russo, V.; Bellevicine, C.; Di Spigna, G.; Fiume, G.; et al. Generation and analysis of spheroids from human primary skin myofibroblasts: An experimental system to study myofibroblasts deactivation. Cell Death Discov. 2017, 3, 17038. [Google Scholar] [CrossRef]
- Rønnov-Jessen, L.; Petersen, O.W. A function for filamentous alpha-smooth muscle actin: Retardation of motility in fibroblasts. J. Cell Biol. 1996, 134, 67–80. [Google Scholar] [CrossRef]
- Tomasek, J.J.; Gabbiani, G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 2002, 3, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Eyden, B. The myofibroblast: Phenotypic characterization as a prerequisite to understanding its functions in translational medicine. J. Cell Mol. Med. 2008, 12, 22–37. [Google Scholar] [CrossRef] [PubMed]
- Arnoldi, R.; Chaponnier, C.; Gabbiani, G.; Hinz, B. Heterogeneity of smooth muscle. In Muscle: Fundamental Biology and Mechanisms of Disease, 1st ed.; Hill, J.A., Olson, E.N., Eds.; Elsevier Academic Press: London, UK; Waltham, MA, USA; San Diego, CA, USA, 2012; Volume 3, pp. 1183–1195. [Google Scholar]
- Hinz, B. The role of myofibroblasts in wound healing. Curr Res. Transl Med. 2016, 64, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B.; Phan, S.H.; Thannickal, V.J.; Prunotto, M.; Desmouliere, A.; Varga, J.; De Wever, O.; Mareel, M.; Gabbiani, G. Recent developments in myofibroblast biology: Paradigms for connective tissue remodeling. Am. J. Pathol. 2012, 180, 1340–1355. [Google Scholar] [CrossRef] [PubMed]
- Curran, T.A.; Ghahary, A. Evidence of a role for fibrocyte and keratinocyte-like cells in the formation of hypertrophic scars. J. Burn Care Res. 2013, 34, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Duffield, J.S.; Lupher, M.; Thannickal, V.J.; Wynn, T.A. Host responses in tissue repair and fibrosis. Annu. Rev. Pathol. 2013, 8, 241–276. [Google Scholar] [CrossRef] [PubMed]
- Greenhalgh, S.N.; Iredale, J.P.; Henderson, N.C. Origins of fibrosis: Pericytes take centre stage. F1000Prime Rep. 2013, 5, 37. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, R.J.; Satish, L.; Yates, C.C.; Wells, A. Pericytes: A newly recognized player in wound healing. Wound Repair Regen. 2016, 24, 204–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.; Ma, Z.; Shankowsky, H.A.; Medina, A.; Tredget, E.E. Deep dermal fibroblast profibrotic characteristics are enhanced by bone marrow-derived mesenchymal stem cells. Wound Repair Regen. 2013, 21, 448–455. [Google Scholar] [CrossRef]
- Calvo, F.; Ege, N.; Grande-Garcia, A.; Hooper, S.; Jenkins, R.P.; Chaudhry, S.I.; Harrington, K.; Williamson, P.; Moeendarbary, E.; Charras, G.; et al. Mechanotransduction and YAP-dependent matrix remodelling is required for the generation and maintenance of cancer associated fibroblasts. Nat. Cell Biol. 2013, 15, 637–646. [Google Scholar] [CrossRef]
- Pakshir, P.; Hinz, B. The big five in fibrosis: Macrophages, myofibroblasts, matrix, mechanics, and miscommunication. Matrix Biol. 2018, 68–69, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.; Foreman, D.M.; Ferguson, M.W. Neutralisation of TGF-beta 1 and TGF-beta 2 or exogenous addition of TGF-beta 3 to cutaneous rat wounds reduces scarring. J. Cell Sci. 1995, 108, 985–1002. [Google Scholar] [PubMed]
- Serini, G.; Gabbiani, G. Modulation of alpha-smooth muscle actin expression in fibroblasts by transforming growth factor-beta isoforms: An in vivo and in vitro study. Wound Repair Regen. 1996, 4, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Rønnov-Jessen, L.; Petersen, O.W. Induction of alpha-smooth muscle actin by transforming growth factor-beta 1 in quiescent human breast gland fibroblasts. Implications for myofibroblast generation in breast neoplasia. Lab. Investig. 1993, 68, 696–707. [Google Scholar] [PubMed]
- Khouw, I.M.; van Wachem, P.B.; Plantinga, J.A.; Vujaskovic, Z.; Wissink, M.J.; de Leij, L.F.; van Luyn, M.J. TGF-beta and bFGF affect the differentiation of proliferating porcine fibroblasts into myofibroblasts in vitro. Biomaterials 1999, 20, 1815–1822. [Google Scholar] [CrossRef]
- Akasaka, Y.; Ono, I.; Tominaga, A.; Ishikawa, Y.; Ito, K.; Suzuki, T.; Imaizumi, R.; Ishiguro, S.; Jimbow, K.; Ishii, T. Basic fibroblast growth factor in an artificial dermis promotes apoptosis and inhibits expression of alpha-smooth muscle actin, leading to reduction of wound contraction. Wound Repair Regen. 2007, 15, 378–389. [Google Scholar] [CrossRef]
- Cushing, M.C.; Mariner, P.D.; Liao, J.T.; Sims, E.A.; Anseth, K.S. Fibroblast growth factor represses Smad-mediated myofibroblast activation in aortic valvular interstitial cells. FASEB J. 2008, 22, 1769–1777. [Google Scholar] [CrossRef]
- Brønnum, H.; Eskildsen, T.; Andersen, D.C.; Schneider, M.; Sheikh, S.P. IL-1β suppresses TGF-β-mediated myofibroblast differentiation in cardiac fibroblasts. Growth Factors 2013, 31, 81–89. [Google Scholar] [CrossRef]
- Mia, M.M.; Boersema, M.; Bank, R.A. Interleukin-1β attenuates myofibroblast formation and extracellular matrix production in dermal and lung fibroblasts exposed to transforming growth factor-β1. PLoS ONE 2014, 9, e91559. [Google Scholar] [CrossRef]
- Oldroyd, S.D.; Thomas, G.L.; Gabbiani, G.; El Nahas, A.M. Interferon-gamma inhibits experimental renal fibrosis. Kidney Int 1999, 56, 2116–2127. [Google Scholar] [CrossRef]
- Yokozeki, M.; Baba, Y.; Shimokawa, H.; Moriyama, K.; Kuroda, T. Interferon-gamma inhibits the myofibroblastic phenotype of rat palatal fibroblasts induced by transforming growth factor-beta1 in vitro. FEBS Lett. 1999, 442, 61–64. [Google Scholar] [CrossRef]
- Tanaka, K.; Sano, K.; Yuba, K.; Katsumura, K.; Nakano, T.; Kobayashi, M.; Ikeda, T.; Abe, M. Inhibition of induction of myofibroblasts by interferon gamma in a human fibroblast cell line. Int. Immunopharmacol. 2003, 3, 1273–1280. [Google Scholar] [CrossRef]
- Sobral, L.M.; Montan, P.F.; Martelli-Junior, H.; Graner, E.; Coletta, R.D. Opposite effects of TGF-beta1 and IFN-gamma on transdifferentiation of myofibroblast in human gingival cell cultures. J. Clin. Periodontol. 2007, 34, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Yao, P.; Lee, E.; Greenhalgh, D.; Soulika, A.M. Interferon-gamma inhibits healing post scald burn injury. Wound Repair Regen. 2012, 20, 580–591. [Google Scholar] [CrossRef] [PubMed]
- Liechty, K.W.; Adzick, N.S.; Crombleholme, T.M. Diminished interleukin 6 (IL-6) production during scarless human fetal wound repair. Cytokine 2000, 12, 671–676. [Google Scholar] [CrossRef]
- Carver, W.; Goldsmith, E.C. Regulation of tissue fibrosis by the biomechanical environment. Biomed. Res. Int. 2013, 2013, 101979. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B.; Mastrangelo, D.; Iselin, C.E.; Chaponnier, C.; Gabbiani, G. Mechanical tension controls granulation tissue contractile activity and myofibroblast differentiation. Am. J. Pathol. 2001, 159, 1009–1020. [Google Scholar] [CrossRef]
- Achterberg, V.F.; Buscemi, L.; Diekmann, H.; Smith-Clerc, J.; Schwengler, H.; Meister, J.J.; Wenck, H.; Gallinat, S.; Hinz, B. The nano-scale mechanical properties of the extracellular matrix regulate dermal fibroblast function. J. Investig. Dermatol. 2014, 134, 1862–1872. [Google Scholar] [CrossRef]
- Yeung, T.; Georges, P.C.; Flanagan, L.A.; Marg, B.; Ortiz, M.; Funaki, M.; Zahir, N.; Ming, W.; Weaver, V.; Janmey, P.A. Effects of substrate stiffness on cell morphology, cytoskeletal structure, and adhesion. Cell Motil. Banner 2005, 60, 24–34. [Google Scholar] [CrossRef]
- Ng, C.P.; Hinz, B.; Swartz, M.A. Interstitial fluid flow induces myofibroblast differentiation and collagen alignment in vitro. J. Cell Sci. 2005, 118, 4731–4739. [Google Scholar] [CrossRef] [Green Version]
- Klingberg, F.; Chow, M.L.; Koehler, A.; Boo, S.; Buscemi, L.; Quinn, T.M.; Costell, M.; Alman, B.A.; Genot, E.; Hinz, B. Prestress in the extracellular matrix sensitizes latent TGF-beta1 for activation. J. Cell Biol. 2014, 207, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Goffin, J.M.; Pittet, P.; Csucs, G.; Lussi, J.W.; Meister, J.J.; Hinz, B. Focal adhesion size controls tension-dependent recruitment of alpha-smooth muscle actin to stress fibers. J. Cell Biol. 2006, 172, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Aarabi, S.; Bhatt, K.A.; Shi, Y.; Paterno, J.; Chang, E.I.; Loh, S.A.; Holmes, J.W.; Longaker, M.T.; Yee, H.; Gurtner, G.C. Mechanical load initiates hypertrophic scar formation through decreased cellular apoptosis. FASEB J. 2007, 21, 3250–3261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wipff, P.J.; Rifkin, D.B.; Meister, J.J.; Hinz, B. Myofibroblast contraction activates latent TGF-beta1 from the extracellular matrix. J. Cell Biol. 2007, 179, 1311–1323. [Google Scholar] [CrossRef] [PubMed]
- Discher, D.E.; Janmey, P.; Wang, Y.L. Tissue cells feel and respond to the stiffness of their substrate. Science 2005, 310, 1139–1143. [Google Scholar] [CrossRef] [PubMed]
- Goldmann, W.H. Mechanosensation: A basic cellular process. Prog. Mol. Biol. Transl. Sci. 2014, 126, 75–102. [Google Scholar] [CrossRef] [PubMed]
- Paszek, M.J.; Zahir, N.; Johnson, K.R.; Lakins, J.N.; Rozenberg, G.I.; Gefen, A.; Reinhart-King, C.A.; Margulies, S.S.; Dembo, M.; Boettiger, D.; et al. Tensional homeostasis and the malignant phenotype. Cancer Cell 2005, 8, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Rosinczuk, J.; Taradaj, J.; Dymarek, R.; Sopel, M. Mechanoregulation of wound healing and skin homeostasis. Biomed. Res. Int. 2016, 2016, 3943481. [Google Scholar] [CrossRef]
- Hsu, CK.; Lin, H.H.; Harn, H.I.; Hughes, M.W.; Tang, M.J.; Yang, C.C. Mechanical forces in skin disorders. J. Dermatol. Sci. 2018, 90, 232–240. [Google Scholar] [CrossRef] [Green Version]
- Harn, H.I.; Ogawa, R.; Hsu, C.K.; Hughes, M.W.; Tang, M.J.; Chuong, C.M. The tension biology of wound healing. Exp. Dermatol. 2019, 28, 464–471. [Google Scholar] [CrossRef]
- Seifert, A.W.; Kiama, S.G.; Seifert, M.G.; Goheen, J.R.; Palmer, T.M.; Maden, M. Skin shedding and tissue regeneration in African spiny mice (Acomys). Nature 2012, 489, 561–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brant, J.O.; Lopez, M.-C.; Baker, H.V.; Barbazuk, W.B.; Maden, M.A. Comparative analysis of gene expression profiles during skin regeneration in Mus and Acomys. PLoS ONE 2015, 10, e0142931. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.; Dyson, M. Tissue replacement in the rabbit’s ear. Brit. J. Surg. 1966, 53, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Goss, R.J.; Grimes, L.N. Epidermal downgrowths in regenerating rabbit ear holes. J. Morphol 1975, 146, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Goss, R.J. Prospects for regeneration in man. Clin. Orth 1980, 151, 270–282. [Google Scholar] [CrossRef]
- Goss, R.J. Regeneration versus repair. In Wound Healing: Biochemical and Clinical Aspects; Cohen, I.K., Diegelmann, R.F., Lindblad, W.J., Eds.; WB Saunders Co: Philadelphia, PA, USA, 1992; pp. 20–39. [Google Scholar]
- Schrementi, M.E.; Ferreira, A.M.; Zender, C.; DiPietro, L.A. Site-specific production of TGF-beta in oral mucosal and cutaneous wounds. Wound Repair Regen. 2008, 16, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Mak, K.; Manji, A.; Gallant-Behm, C.; Wiebe, C.; Hart, D.A.; Larjava, H.; Häkkinen, L. Scarless healing of oral mucosa is characterized by faster resolution of inflammation and control of myofibroblast action compared to skin wounds in the red Duroc pig model. J. Dermatol. Sci. 2009, 56, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.W.; Gallant-Behm, C.; Wiebe, C.; Mak, K.; Hart, D.A.; Larjava, H.; Häkkinen, L. Wound healing in oral mucosa results in reduced scar formation as compared with skin: Evidence from the red Duroc pig model and humans. Wound Repair Regen. 2009, 17, 717–729. [Google Scholar] [CrossRef]
- Larjava, H.; Wiebe, C.; Gallant-Behm, C.; Hart, D.A.; Heino, J.; Häkkinen, L. Exploring scarless healing of oral soft tissues. J. Can. Dent. Assoc. 2011, 77, b18. [Google Scholar]
- Glim, J.E.; van Egmond, M.; Niessen, F.B.; Everts, V.; Beelen, R.H. Detrimental dermal wound healing: What can we learn from the oral mucosa? Wound Repair Regen. 2013, 21, 648–660. [Google Scholar] [CrossRef]
- Lévesque, M.; Gatien, S.; Finnson, K.; Desmeules, S.; Villiard, E.; Pilote, M.; Philip, A.; Roy, S. Transforming growth factor: Beta signaling is essential for limb regeneration in axolotls. PLoS ONE 2007, 2, e1227. [Google Scholar] [CrossRef]
- Lévesque, M.; Villiard, E.; Roy, S. Skin wound healing in axolotls: A scarless process. JEZ-B Mol. Dev. Evol. 2010, 314, 684–697. [Google Scholar] [CrossRef] [PubMed]
- Seifert, A.W.; Monaghan, J.R.; Voss, S.R.; Maden, M. Skin regeneration in adult axolotls: A blueprint for scar-free healing in vertebrates. PLoS ONE 2012, 7, e32875. [Google Scholar] [CrossRef] [PubMed]
- Yannas, I.V.; Tzeranis, D.S.; So, P.T. Surface biology of collagen scaffold explains blocking of wound contraction and regeneration of skin and peripheral nerves. Biomed. Mater. 2015, 11, 014106. [Google Scholar] [CrossRef]
- Yannas, I.V.; Tzeranis, D.S.; So, P.T. Regeneration of injured skin and peripheral nerves requires control of wound contraction, not scar formation. Wound Repair Regen. 2017, 25, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Soller, E.C.; Tzeranis, D.S.; Miu, K.; So, P.T.; Yannas, I.V. Common features of optimal collagen scaffolds that disrupt wound contraction and enhance regeneration both in peripheral nerves and in skin. Biomaterials 2012, 33, 4783–4791. [Google Scholar] [CrossRef]
- Yang, L.; Witten, T.M.; Pidaparti, R.M. A biomechanical model of wound contraction and scar formation. J. Theor. Biol. 2013, 332, 228–248. [Google Scholar] [CrossRef]
- Longaker, M.T.; Rohrich, R.J.; Greenberg, L.; Furnas, H.; Wald, R.; Bansal, V.; Seify, H.; Tran, A.; Weston, J.; Korman, J.M.; et al. A randomized controlled trial of the embrace® device to reduce incisional scar formation. Plast. Reconstr. Surg. 2014, 134, 536–546. [Google Scholar] [CrossRef]
- Lim, A.F.; Weintraub, J.; Kaplan, E.N.; Januszyk, M.; Cowley, C.; McLaughlin, P.; Beasley, B.; Gurtner, G.C.; Longaker, M.T. The embrace® device significantly decreases scarring following scar revision surgery in a randomized controlled trial. Plast. Reconstr. Surg. 2014, 133, 398–405. [Google Scholar] [CrossRef]
- Vig, K.; Chaudhari, A.; Tripathi, S.; Dixit, S.; Sahu, R.; Pillai, S.; Dennis, V.A.; Singh, S.R. Advances in skin regeneration using tissue engineering. Int. J. Mol. Sci. 2017, 18, 789. [Google Scholar] [CrossRef]
- Greenwood, J.E. The evolution of acute burn care - retiring the split skin graft. Ann. R. Coll. Surg. Engl. 2017, 99, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Haddad, A.G.; Giatsidis, G.; Orgill, D.P.; Halvorson, E.G. Skin substitutes and bioscaffolds: Temporary and permanent coverage. Clin. Plast. Surg. 2017, 44, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Kua, E.H.; Goh, C.Q.; Ting, Y.; Chua, A.; Song, C. Comparing the use of glycerol preserved and cryopreserved allogenic skin for the treatment of severe burns: Differences in clinical outcomes and in vitro tissue viability. Cell Tissue Bank. 2012, 13, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Cleland, H.; Wasiak, J.; Dobson, H.; Paul, M.; Pratt, G.; Paul, E.; Herson, M.; Akbarzadeh, S. Clinical application and viability of cryopreserved cadaveric skin allografts in severe burn: A retrospective analysis. Burns 2014, 40, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Pianigiani, E.; Tognetti, L.; Ierardi, F.; Mariotti, G.; Rubegni, P.; Cevenini, G.; Perotti, R.; Fimiani, M. Assessment of cryopreserved donor skin viability: The experience of the regional tissue bank of Siena. Cell Tissue Bank. 2016, 17, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Shen, Y.I.; Harmon, J.W. Engineering pro-regenerative hydrogels for scarless wound healing. Adv. Healthc. Mater. 2018, 14, e1800016. [Google Scholar] [CrossRef] [PubMed]
- Sun, G. Pro-regenerative hydrogel restores scarless skin during cutaneous wound healing. Adv. Healthc. Mater. 2017, 6. [Google Scholar] [CrossRef]
- Groeber, F.; Holeiter, M.; Hampel, M.; Hinderer, S.; Schenke-Layland, K. Skin tissue engineering – in vivo and in vitro applications. Adv. Drug Deliv. Rev. 2011, 63, 352–366. [Google Scholar] [CrossRef]
- Ananta, M.; Brown, R.A.; Mudera, V. A rapid fabricated living dermal equivalent for skin tissue engineering: An in vivo evaluation in an acute wound model. Tissue Eng. Part A 2012, 18, 353–361. [Google Scholar] [CrossRef]
- Heimbach, D.M.; Warden, G.D.; Luterman, A.; Jordan, M.H.; Ozobia, N.; Ryan, C.M.; Voigt, D.W.; Hickerson, W.L.; Saffle, J.R.; DeClement, F.A.; et al. Multicenter postapproval clinical trial of Integra dermal regeneration template for burn treatment. J. Burn Care Res. 2003, 24, 42–48. [Google Scholar] [CrossRef]
- Chermnykh, E.S.; Kiseleva, E.V.; Rogovaya, O.S.; Rippa, A.L.; Vasiliev, A.V.; Vorotelyak, E.A. Tissue-engineered biological dressing accelerates skin wound healing in mice via formation of provisional connective tissue. Histol. Histopathol. 2018, 30, 18006. [Google Scholar] [CrossRef]
- Yannas, I.V. Collagen and gelatin in the solid state. Rev. Macromol. Chem. 1972, 7, 49–104. [Google Scholar] [CrossRef]
- Kuo, Z.K.; Lai, P.L.; Toh, E.K.; Weng, C.H.; Tseng, H.W.; Chang, P.Z.; Chen, C.C.; Cheng, C.M. Osteogenic differentiation of preosteoblasts on a hemostatic gelatin sponge. Sci. Rep. 2016, 6, 32884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, J.S.; Snelling, C.F.; Clugston, P.; Macdonald, I.B.; Germann, E. Cultured epithelial autograft: Five years of clinical experience with twenty-eight patients. J. Trauma 1995, 39, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Still, J.; Glat, P.; Silverstein, P.; Griswold, J.; Mozingo, D. The use of a collagen sponge/living cell composite material to treat donor sites in burn patients. Burns 2003, 29, 837–841. [Google Scholar] [CrossRef]
- Nicoletti, G.; Brenta, F.; Bleve, M.; Pellegatta, T.; Malovini, A.; Faga, A.; Perugini, P. Long-term in vivo assessment of bioengineered skin substitutes: A clinical study. J. Tissue Eng. Regen. Med. 2015, 9, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Philandrianos, C.; Andrac-Meyer, L.; Mordon, S.; Feuerstein, J.M.; Sabatier, F.; Veran, J.; Magalon, G.; Casanova, D. Comparison of five dermal substitutes in full-thickness skin wound healing in a porcine model. Burns 2012, 38, 820–829. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.Q.; Potokar, T.S.; Price, P. An objective long-term evaluation of Integra (a dermal skin substitute) and split thickness skin grafts, in acute burns and reconstructive surgery. Burns 2010, 36, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Lindner, G.; Horland, R.; Wagner, I.; Atac, B.; Lauster, R. De novo formation and ultrastructural characterization of a fiber-producing human hair follicle equivalent in vitro. J. Biotechnol. 2011, 152, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, T.; Yoshimura, C.; Myasnikova, D.; Kataoka, K.; Nittami, T.; Maruo, S.; Fukuda, J. Spontaneous hair follicle germ (HFG) formation in vitro, enabling the large-scale production of HFGs for regenerative medicine. Biomaterials 2018, 154, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Wagner, I.; Atac, B.; Lindner, G.; Horland, R.; Busek, M.; Sonntag, F.; Klotzbach, U.; Thomas, A.; Lauster, R.; Marx, U. Skin and hair-on-a-chip: Hair and skin assembly versus native skin maintenance in a chip-based perfusion system. BMC Proc. 2013, 7, 93. [Google Scholar] [CrossRef]
- Mori, N.; Morimoto, Y.; Takeuchi, S. Skin integrated with perfusable vascular channels on a chip. Biomaterials 2017, 116, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Sriram, G.; Alberti, M.; Dancik, Y.; Wu, B.; Wu, R.; Feng, Z.; Ramasam, S.; Bigliardi, P.L.; Bigliardi-Qi, M.; Wang, Z. Full-thickness human skin-on-chip with enhanced epidermal morphogenesis and barrier function. Mater. Today 2018, 21, 326–340. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristics | Papillary FBs | Reticular FBs |
|---|---|---|
| Morphology | Spindle-shaped [11] | Strongly spread out over the substrate and have a stellate shape [22] |
| Extracellular matrix | Weaker alignment of collagen fibers [5], a higher content of proteoglycans [4] | Ordered network of collagen fibers and elastin strands [7] |
| Positivity for α-SMA | Small part of FBs is α-SMA positive [13] | Most part of the FBs is α-SMA positive [13,19]. |
| Functional activities in culture | High proliferative and synthetic activity [12,13,17] | Low proliferative and synthetic activity [12,13,17] |
| Expression of cell markers in mice dermis | Express genes of the WNT pathway proteins and CD39 [11]; CD26+Sca1–FBs express COL6A5, COL23A1, APCDD1, HSPB3, CD3g, CD3d, CD3ε [11] | Include Dlk1+Sca1+ and Dlk1–Sca1+ preadipocytes [11]; Dlk1+Sca1−FBs express genes of the secretoglobin superfamily [11], CD36 is upregulated in the lower reticular FBs [11] |
| Expression of cell markers in human dermis | Express genes of the WNT pathway proteins and CD39 [11]; FAP+CD90–FBs express PDPN and NTN1 [13,16] | FAP−CD90+ FBs express high levels of ACTA2, MGP, PPARγ, CD36 [16]; FBs express CD26, MFAP5, PRG4 [11], MGP and TGM2 [13] |
| Contribution to skin equivalents | Support the formation of a multilayered, more stratified and differentiated epidermis with epidermal ridges [3,11]. ECM components promote the predominant growth of keratinocytes [21] | Do not support the formation of fully-stratified epidermis [11]. The composition of the basement membrane is altered [23] |
| Scarring | May improve scar condition [30] | Generate the matrix typical for fibrosis [14] |
| Type of scaffold | Biologic Biosynthetic Synthetic |
| Substitute thickness | Split-thickness Full-thickness |
| Number of layers | Single-layered Bi-layered Multi-layered |
| Cell origin | Autologous Allogenic Xenogenic Acellular |
| Cell type included | Keratinocytes Fibroblasts Other cell types |
| The skin layer to be replaced | Epidermis Dermis Compound |
| Period of application | Temporary Permanent |
| Type of wound to be treated | Superficial Deep |
| Prospective “regenerative” criteria | Effect on contraction Vascularization Epithelialization rate Scar parameters The ability to stimulate skin appendages regeneration Long-term follow-up |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rippa, A.L.; Kalabusheva, E.P.; Vorotelyak, E.A. Regeneration of Dermis: Scarring and Cells Involved. Cells 2019, 8, 607. https://doi.org/10.3390/cells8060607
Rippa AL, Kalabusheva EP, Vorotelyak EA. Regeneration of Dermis: Scarring and Cells Involved. Cells. 2019; 8(6):607. https://doi.org/10.3390/cells8060607
Chicago/Turabian StyleRippa, Alexandra L., Ekaterina P. Kalabusheva, and Ekaterina A. Vorotelyak. 2019. "Regeneration of Dermis: Scarring and Cells Involved" Cells 8, no. 6: 607. https://doi.org/10.3390/cells8060607
APA StyleRippa, A. L., Kalabusheva, E. P., & Vorotelyak, E. A. (2019). Regeneration of Dermis: Scarring and Cells Involved. Cells, 8(6), 607. https://doi.org/10.3390/cells8060607