Mutant Huntingtin Affects Diabetes and Alzheimer’s Markers in Human and Cell Models of Huntington’s Disease


Abstract
:
1. Introduction
1.1. Biology of Huntingtin and Identification of DNA Polymorphism Causing HD
1.2. Neurodegeneration and Metabolic Diseases
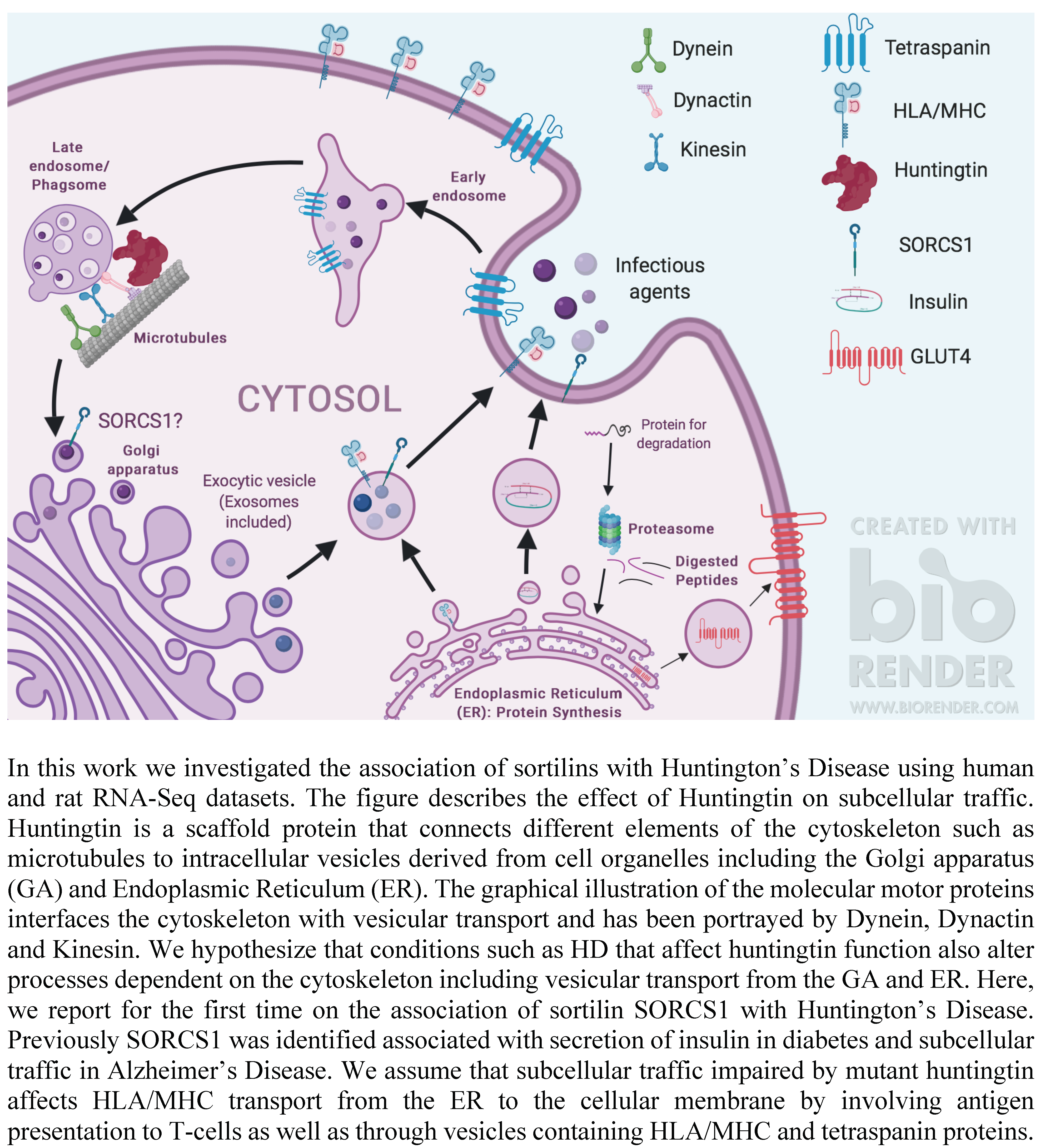
1.3. VPS10P-Domain Receptors or Sortilins: Regulators of Subcellular Protein Trafficking and Markers of Diabetes and Neurodegeneration
2. Materials and Methods
2.1. Library Preparation and Sequencing
2.2. Human and Rat GENOMES
2.3. Genome-Wide Association Study (GWAS)
2.4. Gene Expression Omnibus (GEO) RNA-Seq Datasets
2.5. Post-GATK/GWAS Processing
2.6. Statistical Analysis
2.7. Manhattan Plots
2.8. Linkage Visualization Using Haploview
2.9. Validation of GATK/GWAS DNA Variants Identification by Pyrosequencing
3. Results
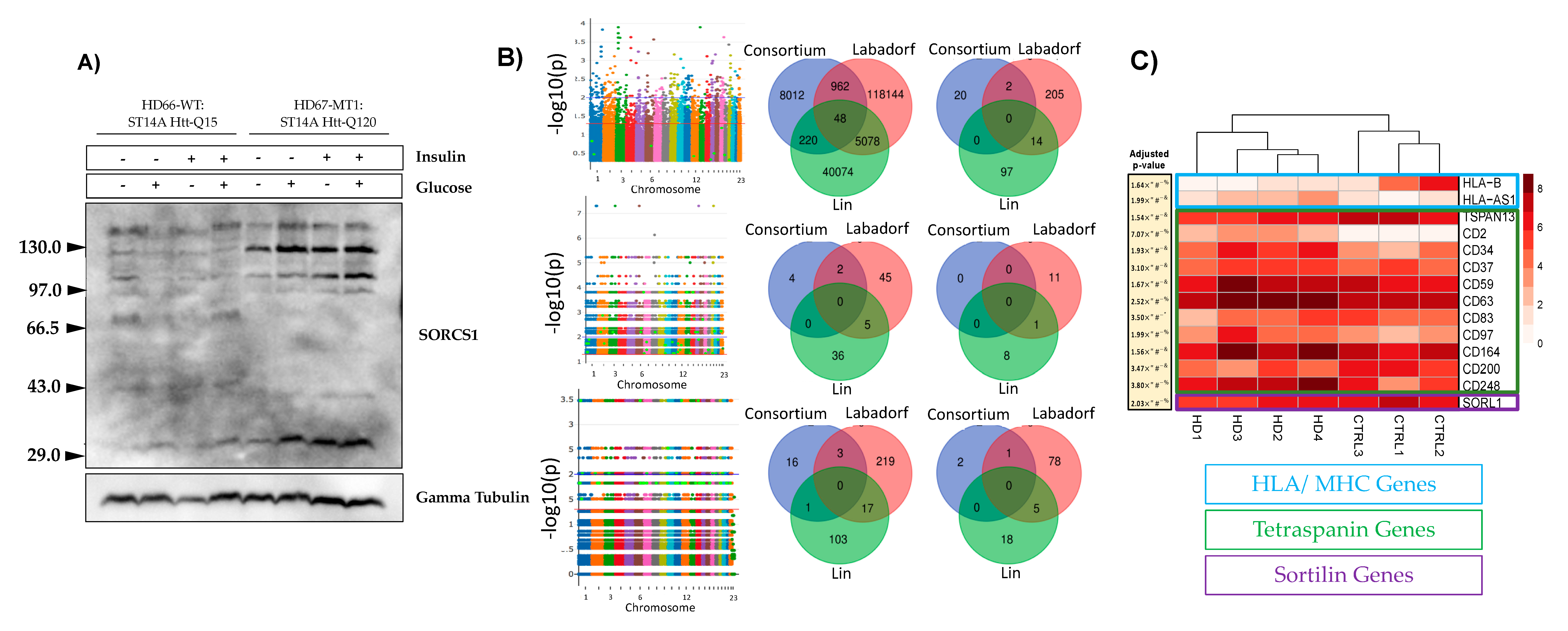
3.1. mHTT Is Associated with SORCS1 Protein Up-Regulation and Sortilins SNPs
3.2. mHTT Affects Pathways Important for Immunological Function
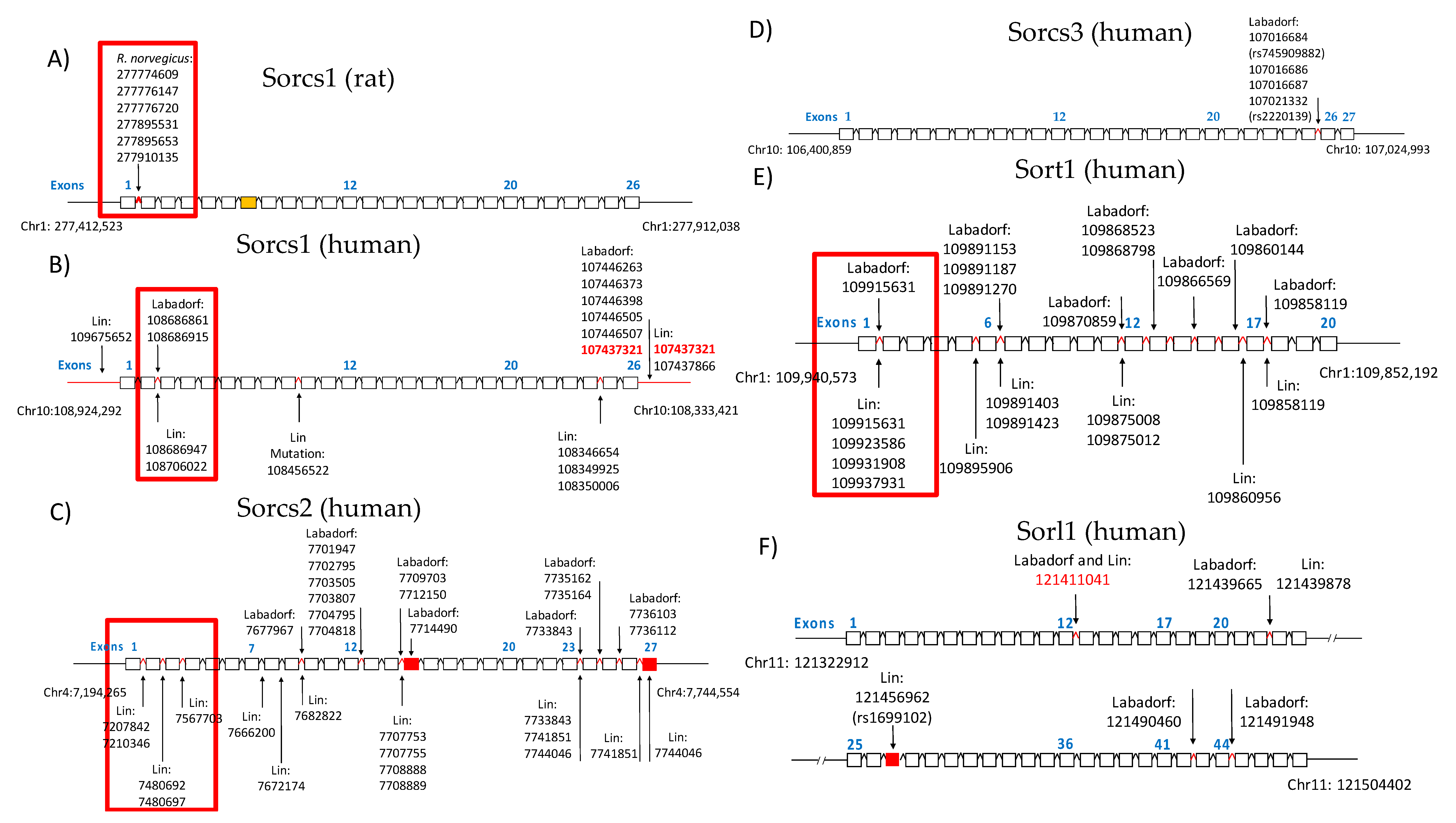
3.3. Validation of GATK/GWAS DNA Variants Identification by Pyrosequencing Methodology
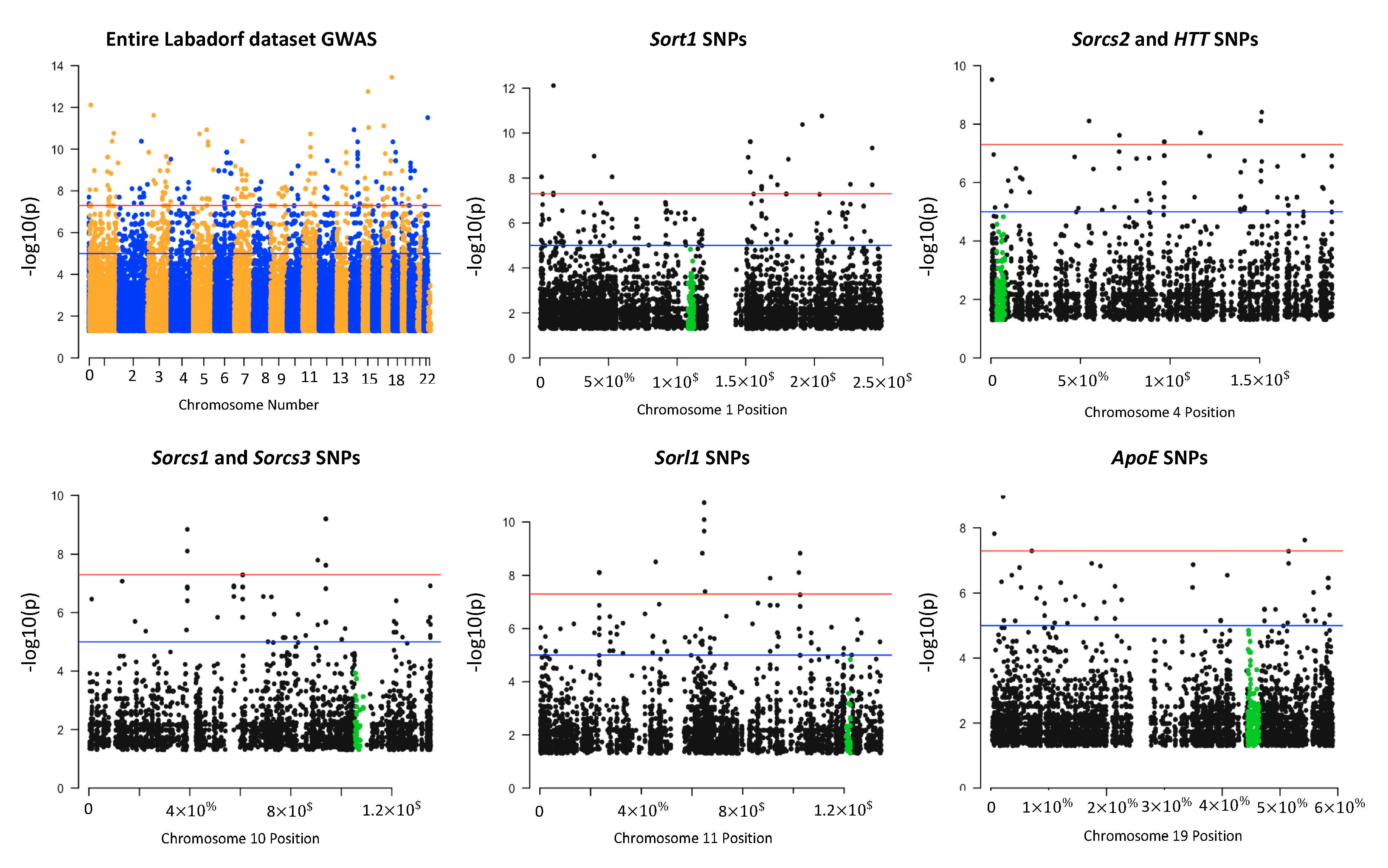
3.4. Sortilin and HTT SNPs Identified by GATK/GWAS Pipeline of RNA-Seq Datasets of Human Origin Are in Linkage Disequilibrium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Montojo, M.; Aganzo, M.; González, N. Huntington’s Disease and Diabetes: Chronological Sequence of its Association. J. Huntington’s Dis. 2017, 6, 179–188. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, M.E.; Ambrose, C.M.; Duyao, M.P.; Myers, R.H.; Lin, C.; Srinidhi, L.; Barnes, G.; Taylor, S.A.; James, M.; Groot, N.; et al. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. Cell 1993, 72, 971–983. [Google Scholar] [CrossRef]
- Palidwor, G.A.; Shcherbinin, S.; Huska, M.R.; Rasko, T.; Stelzl, U.; Arumughan, A.; Foulle, R.; Porras, P.; Sanchez-Pulido, L.; Wanker, E.E.; et al. Detection of Alpha-Rod Protein Repeats Using a Neural Network and Application to Huntingtin. Plos Comput. Biol. 2009, 5, e1000304. [Google Scholar] [CrossRef] [PubMed]
- Gusella, J.F.; Wexler, N.S.; Conneally, M.P.; Naylor, S.L.; Anderson, M.; Tanzi, R.E.; Watkins, P.C.; Ottina, K.; Wallace, M.R.; Sakaguchi, A.Y.; et al. A polymorphic DNA marker genetically linked to Huntington’s disease. Nature 1983, 306, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Warby, S.C.; Montpetit, A.; Hayden, A.R.; Carroll, J.B.; Butland, S.L.; Visscher, H.; Collins, J.A.; Semaka, A.; Hudson, T.J.; Hayden, M.R. CAG Expansion in the Huntington Disease Gene Is Associated with a Specific and Targetable Predisposing Haplogroup. Am. J. Hum. Genet. 2009, 84, 351–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podolsky, S.; Leopold, N.; Sax, D. Increased frequency of diabetes mellitus in patients with huntington’s chorea. Lancet 1972, 299, 1356–1359. [Google Scholar] [CrossRef]
- Farrer, L.A. Diabetes mellitus in Huntington disease. Clin. Genet. 1985, 27, 62–67. [Google Scholar] [CrossRef]
- Saudou, F.; Humbert, S. The Biology of Huntingtin. Neuron 2016, 89, 910–926. [Google Scholar] [CrossRef] [Green Version]
- Zala, D.; Hinckelmann, M.-V.; Yu, H.; Lyra da Cunha, M.; Liot, G.; Cordelières, F.P.; Marco, S.; Saudou, F. Vesicular Glycolysis Provides On-Board Energy for Fast Axonal Transport. Cell 2013, 152, 479–491. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, K.A.; Richardson, C.C.; Ravishankar, A.; Brigatti, C.; Liberati, D.; Lampasona, V.; Piemonti, L.; Morgan, D.; Feltbower, R.G.; Christie, M.R. Identification of Tetraspanin-7 as a Target of Autoantibodies in Type 1 Diabetes. Diabetes 2016, 65, 1690–1698. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Pessin, J.E. Ins (endocytosis) and outs (exocytosis) of GLUT4 trafficking. Curr. Opin. Cell Biol. 2007, 19, 466–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kebede, M.A.; Oler, A.T.; Gregg, T.; Balloon, A.J.; Johnson, A.; Mitok, K.; Rabaglia, M.; Schueler, K.; Stapleton, D.; Thorstenson, C.; et al. SORCS1 is necessary for normal insulin secretory granule biogenesis in metabolically stressed β cells. J. Clin. Investig. 2014, 124, 4240–4256. [Google Scholar] [CrossRef] [PubMed]
- Marcusson, E.G.; Horazdovsky, B.F.; Cereghino, J.; Gharakhanian, E.; Emr, S.D. The sorting receptor for yeast vacuolar carboxypeptidase Y is encoded by the VPS10 gene. Cell 1994, 77, 579–586. [Google Scholar] [CrossRef]
- Reitz, C. The role of the retromer complex in aging-related neurodegeneration: A molecular and genomic review. Mol. Genet. Genomics 2015, 290, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Reitz, C.; Tosto, G.; Vardarajan, B.; Rogaeva, E.; Ghani, M.; Rogers, R.S.; Conrad, C.; Haines, J.L.; Pericak-Vance, M.A.; Fallin, M.D.; et al. Independent and epistatic effects of variants in VPS10-d receptors on Alzheimer disease risk and processing of the amyloid precursor protein (APP). Transl. Psychiatry 2013, 3, e256. [Google Scholar] [CrossRef] [PubMed]
- Reitz, C.; Tokuhiro, S.; Clark, L.N.; Conrad, C.; Vonsattel, J.; Hazrati, L.; Palotás, A.; Lantigua, R.; Medrano, M.; Jiménez-Velázquez, I.Z.; et al. SORCS1 alters amyloid precursor protein processing and variants may increase Alzheimer’s disease risk. Ann. Neurol. 2011, 69, 47–64. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, C.L.; Rodgers, A.; Herbst, S.; Coade, S.; Gronow, A.; Guzman, C.A.; Wilson, M.S.; Kanzaki, M.; Nykjaer, A.; Gutierrez, M.G. The proneurotrophin receptor sortilin is required for Mycobacterium tuberculosis control by macrophages. Sci. Rep. 2016, 6, 29332. [Google Scholar] [CrossRef] [Green Version]
- Reuter, E.; Weber, J.; Paterka, M.; Ploen, R.; Breiderhoff, T.; van Horssen, J.; Willnow, T.E.; Siffrin, V.; Zipp, F. Role of Sortilin in Models of Autoimmune Neuroinflammation. J. Immunol. 2015, 195, 5762–5769. [Google Scholar] [CrossRef] [Green Version]
- Chaves, G.; Özel, R.; Rao, N.V.; Hadiprodjo, H.; Costa, Y.; Tokuno, Z.; Pourmand, N. Metabolic and transcriptomic analysis of Huntington’s disease model reveal changes in intracellular glucose levels and related genes. Heliyon 2017, 3, e00381. [Google Scholar] [CrossRef]
- Picelli, S.; Faridani, O.R.; Björklund, Å.K.; Winberg, G.; Sagasser, S.; Sandberg, R. Full-length RNA-seq from single cells using Smart-seq. Nat. Protoc. 2014, 9, 171. [Google Scholar] [CrossRef]
- Nvigen. Available online: www.nvigen.com (accessed on 1 June 2019).
- Ensembl Genome Browser. Available online: https://grch37.ensembl.org/Homo_sapiens/Info/Index; https://mar2015.archive.ensembl.org/Rattus_norvegicus/Info/Index (accessed on 1 June 2019).
- Haploview. Available online: https://learn.gencore.bio.nyu.edu/variant-calling/; broadinstitute.org/haploview/haploview (accessed on 1 June 2019).
- DePristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491. [Google Scholar] [CrossRef] [PubMed]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. Current Protocols in Bioinformatics; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2013. [Google Scholar]
- Visscher, P.M.; Wray, N.R.; Zhang, Q.; Sklar, P.; McCarthy, M.I.; Brown, M.A.; Yang, J. 10 Years of GWAS Discovery: Biology, Function, and Translation. Am. J. Hum. Genetics 2017, 101, 5–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labadorf, A.; Hoss, A.G.; Lagomarsino, V.; Latourelle, J.C.; Hadzi, T.C.; Bregu, J.; MacDonald, M.E.; Gusella, J.F.; Chen, J.-F.; Akbarian, S.; et al. RNA Sequence Analysis of Human Huntington Disease Brain Reveals an Extensive Increase in Inflammatory and Developmental Gene Expression. PLoS ONE 2015, 10, e0143563. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Park, J.; Ramachandran, S.; Zhang, Y.; Tseng, Y.-T.; Shen, S.; Waldvogel, H.J.; Curtis, M.A.; Faull, R.L.; Troncoso, J.C.; et al. Transcriptome sequencing reveals aberrant alternative splicing in Huntington’s disease. Hum. Mol. Genet. 2016, 25, 3454–3466. [Google Scholar] [CrossRef] [PubMed]
- Consortium, T.H.; Lim, R.G.; Salazar, L.L.; Wilton, D.K.; King, A.R.; Stocksdale, J.T.; Sharifabad, D.; Lau, A.L.; Stevens, B.; Reidling, J.C.; et al. Developmental alterations in Huntington’s disease neural cells and pharmacological rescue in cells and mice. Nat. Neurosci. 2017, 20, 648. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; The Sequence Alignment/Map format and SAMtools. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Turner, S.D. qqman: An R package for visualizing GWAS results using Q-Q and manhattan plots. Biorxiv 2014, 005165. [Google Scholar] [CrossRef]
- Barrett, J.; Fry, B.; Maller, J.; Daly, M. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Pourmand, N.; Elahi, E.; Davis, R.W.; Ronaghi, M. Multiplex Pyrosequencing. Nucleic Acids Res. 2002, 30, e31. [Google Scholar] [CrossRef] [PubMed]
- Royo, J.; Hidalgo, M.; Ruiz, A. Pyrosequencing protocol using a universal biotinylated primer for mutation detection and SNP genotyping. Nat. Protoc. 2007, 2, 1734–1739. [Google Scholar] [CrossRef] [PubMed]
- Schächter, F.; Faure-Delanef, L.; Guénot, F.; Rouger, H.; Froguel, P.; Lesueur-Ginot, L.; Cohen, D. Genetic associations with human longevity at the APOE and ACE loci. Nat. Genet. 1994, 6, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Nejentsev, S.; Howson, J.M.; Walker, N.M.; Szeszko, J.; Field, S.F.; Stevens, H.E.; Reynolds, P.; Hardy, M.; King, E.; Masters, J.; et al. Localization of type 1 diabetes susceptibility to the MHC class I genes HLA-B and HLA-A. Nature 2007, 450, 887. [Google Scholar] [CrossRef]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Cellular and Molecular Immunology, 6th ed.; WB Saunders: St. Louis, MO, USA, 2010. [Google Scholar]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Bu, G. Apolipoprotein E and its receptors in Alzheimer’s disease: Pathways, pathogenesis and therapy. Nat. Rev. Neurosci. 2009, 10, 333–344. [Google Scholar] [CrossRef]
- Scherzer, C.R.; Offe, K.; Gearing, M.; Rees, H.D.; Fang, G.; Heilman, C.J.; Schaller, C.; Bujo, H.; Levey, A.I.; Lah, J.J. Loss of Apolipoprotein E Receptor LR11 in Alzheimer Disease. Arch. Neurol-chicago 2004, 61, 1200–1205. [Google Scholar] [CrossRef] [Green Version]
- Leloup, N.; Lössl, P.; Meijer, D.H.; Brennich, M.; Heck, A.J.; Thies-Weesie, D.M.; Janssen, B.J. Low pH-induced conformational change and dimerization of sortilin triggers endocytosed ligand release. Nat. Commun. 2017, 8, 1708. [Google Scholar] [CrossRef]
- Östensson, M.; Montén, C.; Bacelis, J.; Gudjonsdottir, A.H.; Adamovic, S.; Ek, J.; Ascher, H.; Pollak, E.; Arnell, H.; Browaldh, L.; et al. A Possible Mechanism behind Autoimmune Disorders Discovered By Genome-Wide Linkage and Association Analysis in Celiac Disease. PLoS ONE 2013, 8, e70174. [Google Scholar] [CrossRef]
- Foerster, K.; Freudenberg, J. HLA antigen frequencies in patients with huntington’s chorea and their relatives. J. Neurol. 1980, 223, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Madsen, H.; Nielsen, L.; Sørensen, S. An association study of Huntington’s disease and HLA. J. Med. Genet. 1982, 19, 452–454. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
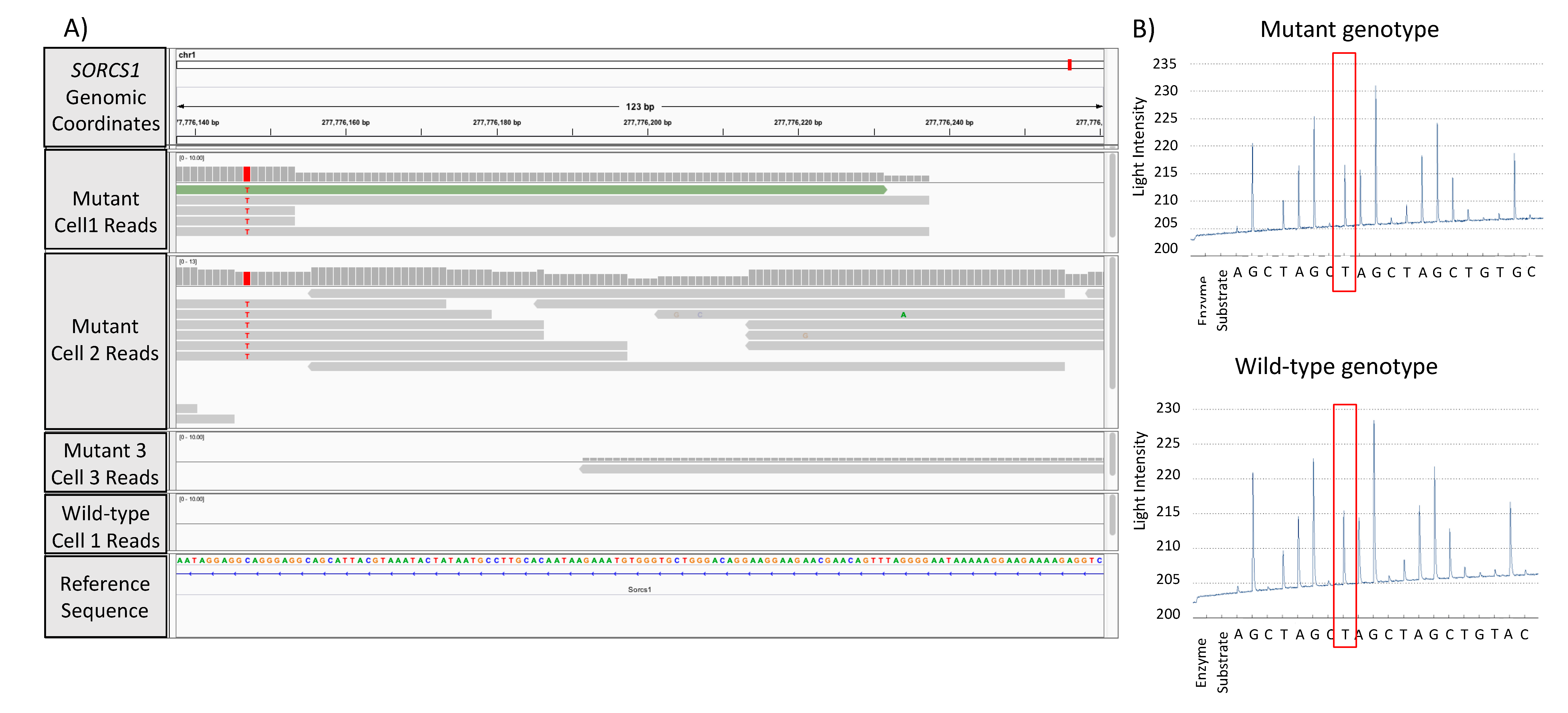{kind=link}
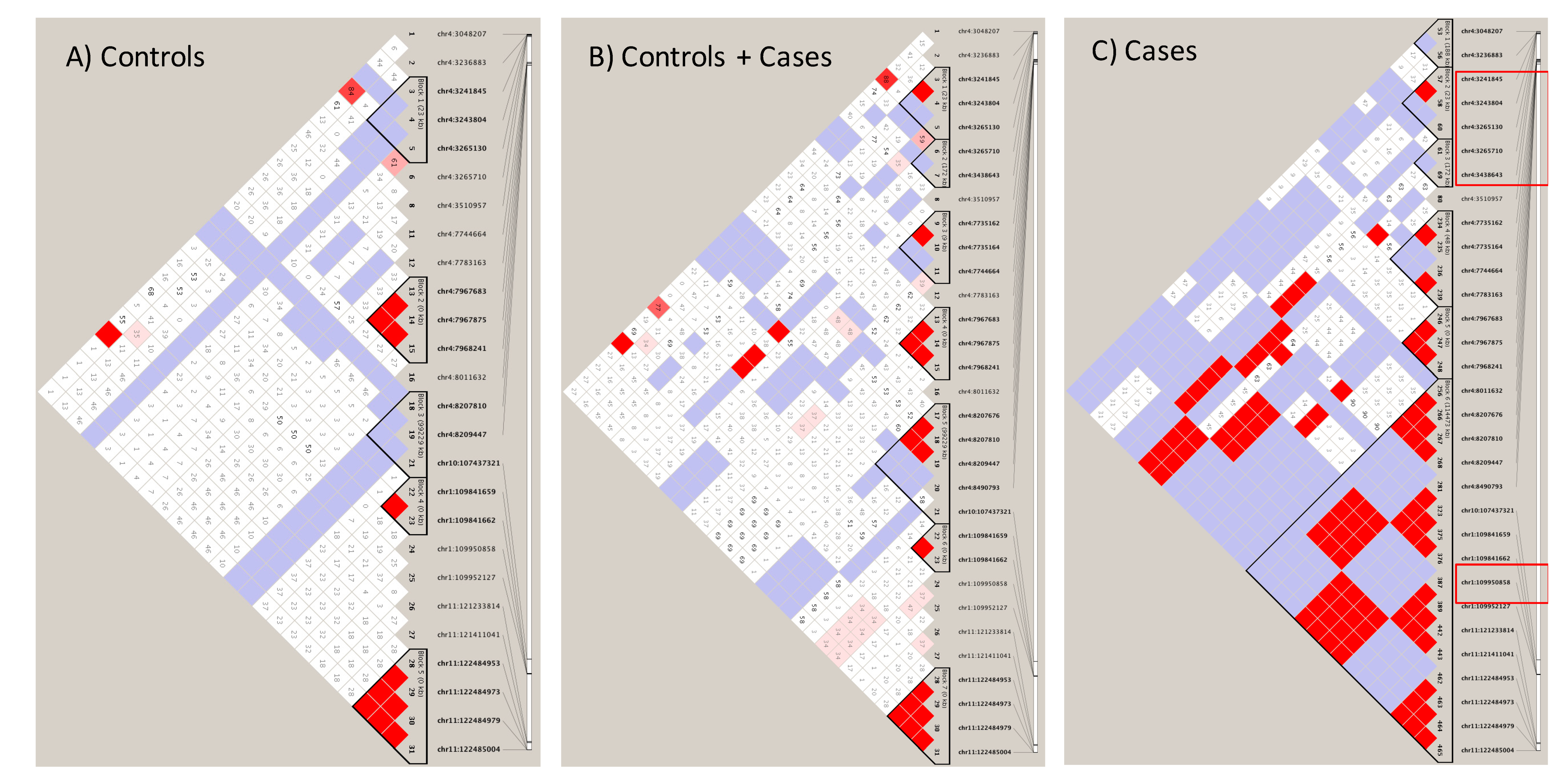{kind=link}
| Dataset | Total Number of SNPs Identified | Significant SNPs (p < 0.05) | Sortilin Gene Body Size (bp) | SNPs in Gene Body (p < 0.05) | SNPs in Vicinity (±106 bp Away from Gene Body) | |
|---|---|---|---|---|---|---|
| Rattus norvegicus (ST14A cells) | 222997 | 8464 (3.80%) | SORCS1 | 499515 | 6 | 13 |
| SORCS2 | 371129 | 0 | 8 | |||
| SORCS3 | 641740 | 0 | 2 | |||
| SORT1 | 89539 | 0 | 11 | |||
| SORL1 | 170117 | 0 | 5 | |||
| Labadorf | 2658838 | 84349 (3.17%) | SORCS1 | 590871 | 2 | 8 |
| SORCS2 | 550289 | 15 | 171 | |||
| SORCS3 | 624134 | 4 | 53 | |||
| SORT1 | 88381 | 9 | 153 | |||
| SORL1 | 181490 | 4 | 38 | |||
| Lin | 3172675 | 44669 (1.4%) | SORCS1 | 590871 | 6 | 9 |
| SORCS2 | 550289 | 10 | 121 | |||
| SORCS3 | 624134 | 0 | 23 | |||
| SORT1 | 88381 | 11 | 111 | |||
| SORL1 | 181490 | 4 | 41 | |||
| HD iPSC Consortium (iPSC Cells) | 238013 | 8979 (3.38%) | SORCS1 | 590871 | 0 | 0 |
| SORCS2 | 550289 | 0 | 20 | |||
| SORCS3 | 624134 | 0 | 3 | |||
| SORT1 | 88381 | 0 | 22 | |||
| SORL1 | 181490 | 3 | 6 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaves, G.; Stanley, J.; Pourmand, N. Mutant Huntingtin Affects Diabetes and Alzheimer’s Markers in Human and Cell Models of Huntington’s Disease. Cells 2019, 8, 962. https://doi.org/10.3390/cells8090962
Chaves G, Stanley J, Pourmand N. Mutant Huntingtin Affects Diabetes and Alzheimer’s Markers in Human and Cell Models of Huntington’s Disease. Cells. 2019; 8(9):962. https://doi.org/10.3390/cells8090962
Chicago/Turabian StyleChaves, Gepoliano, John Stanley, and Nader Pourmand. 2019. "Mutant Huntingtin Affects Diabetes and Alzheimer’s Markers in Human and Cell Models of Huntington’s Disease" Cells 8, no. 9: 962. https://doi.org/10.3390/cells8090962
APA StyleChaves, G., Stanley, J., & Pourmand, N. (2019). Mutant Huntingtin Affects Diabetes and Alzheimer’s Markers in Human and Cell Models of Huntington’s Disease. Cells, 8(9), 962. https://doi.org/10.3390/cells8090962