The Rare IL22RA2 Signal Peptide Coding Variant rs28385692 Decreases Secretion of IL-22BP Isoform-1, -2 and -3 and Is Associated with Risk for Multiple Sclerosis

 , , ,
, , ,  , ,
, ,  , ,
, , 

 add
Show full author list
add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Controls
2.2. Selection and Genotyping of SNPs
2.3. Statistical Analysis
2.4. Functional Annotation of SNPs
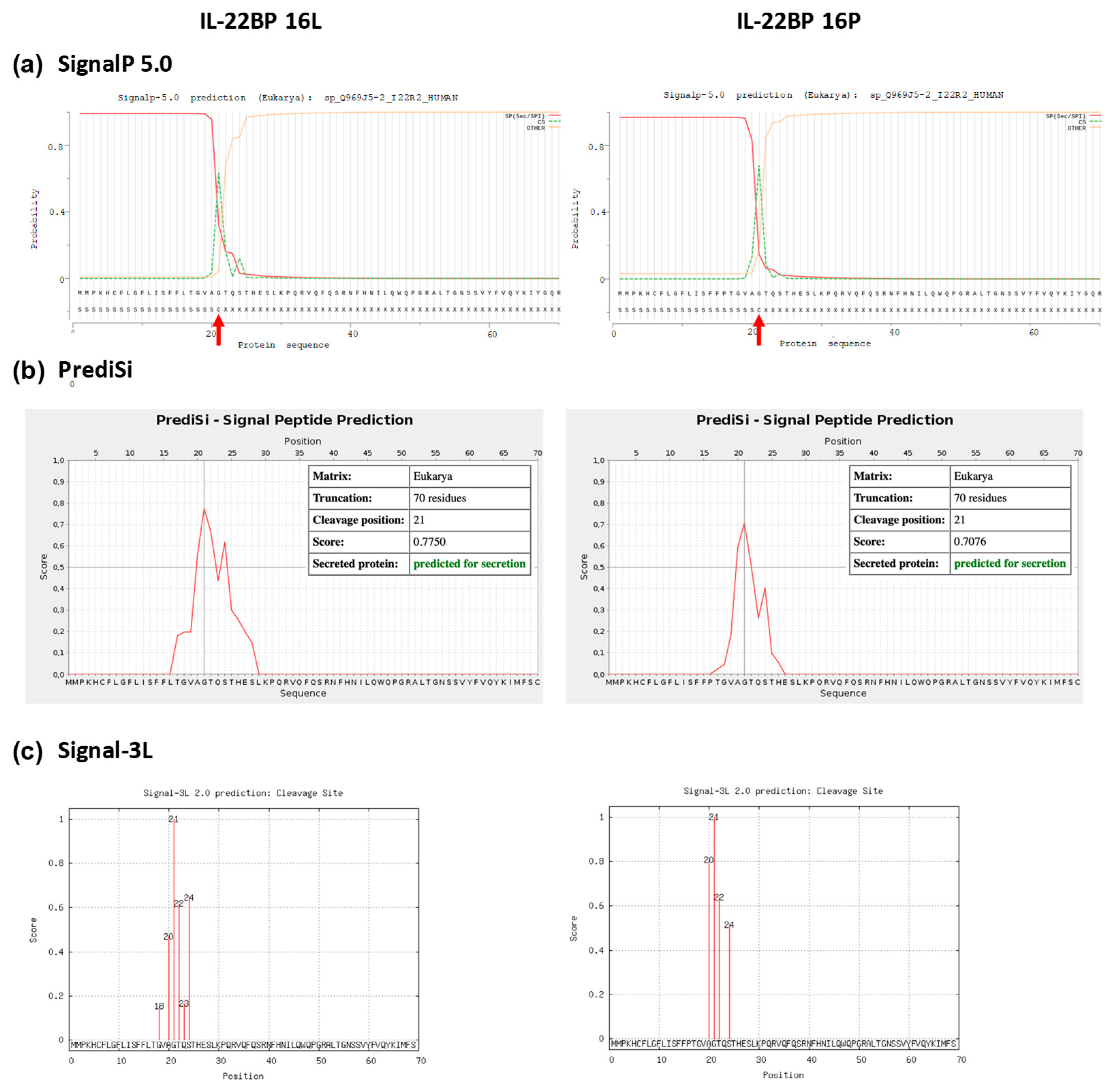
2.5. In silico Analysis of the Effect of the Leu to Pro Transition Coded by rs28385692 on Signal Peptide Characteristics
2.6. p.Leu16Pro Mutagenesis of the Three IL-22BP Isoforms
2.7. Assessment of Effect of p.Leu16Pro Variant on Secretion of IL-22BP Isoforms
2.8. Replacement of the SP of IL-22BPi2 with the IL17A SP
2.9. Flow Cytometry Analysis
3. Results
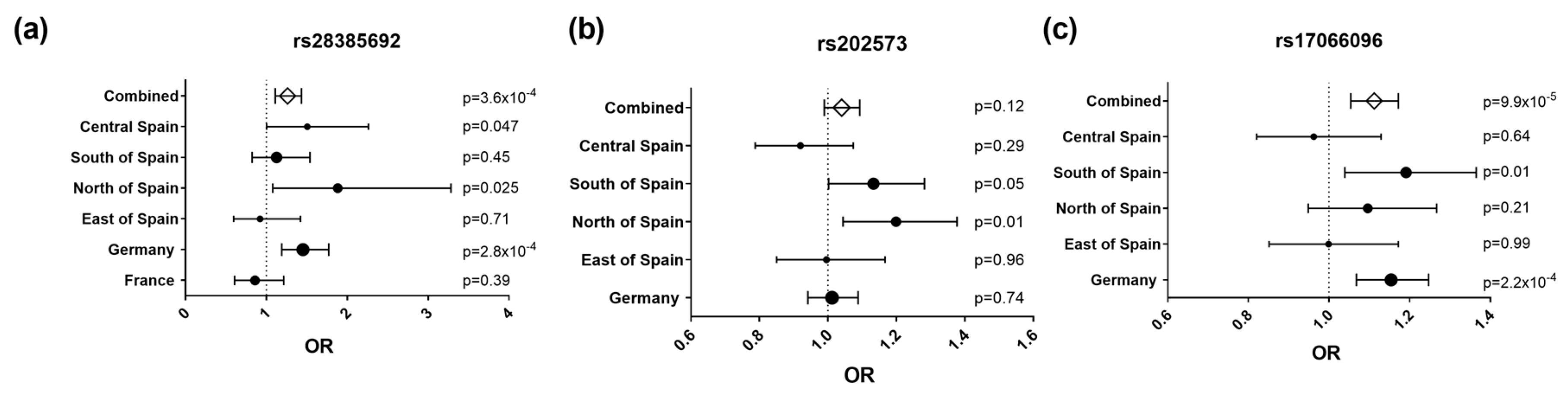
3.1. The p.Leu16Pro coding SNP rs28385692 is Associated with Risk for Multiple Sclerosis
3.2. In Silico Analysis of the p.Leu16Pro Variant
3.3. The p.Leu16Pro Variant Decreases Secretion of IL-22BPi1, IL-22BPi2 and IL-22BPi3
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AP | Acetone precipitate |
| CL | Cell lysate |
| EAE | Experimental autoimmune encephalomyelitis |
| EDSS | Expanded Disability Status Scale |
| GWAS | Genome-wide association screen |
| IL-22BPi1, 2 or 3 | Interleukin 22 binding protein isoform-1, -2 or -3 |
| IL22RA2 | Interleukin 22 receptor subunit alpha 2 |
| LD | Linkage disequilibrium |
| MAF | Minor allele frequency |
| OR | Odds ratio |
| PP | Primary progressive |
| RAF | Risk allele frequency |
| RR | Relapsing-remitting |
| ScP | Secondary progressive |
| SFM | Serum-free medium |
| SNP | Single nucleotide polymorphism |
| SP | Signal peptide |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP | OR 1 (95% CI 2) | p |
|---|---|---|
| rs4896239 | 0.95 (0.798–1.131) | 0.5638 |
| rs17066096 | 1.059 (0.8746–1.282) | 0.5569 |
| rs12194034 | 1.025 (0.8412–1.249) | 0.8055 |
| rs1543509 | 0.9821 (0.7719–1.25) | 0.8835 |
| rs28366 | 1.047 (0.8594–1.275) | 0.6497 |
| rs276466 | 0.9779 (0.7803–1.225) | 0.8458 |
| rs10484798 | 0.8452 (0.6874–1.039) | 01105 |
| rs13217897 | 0.8249 (0.6608–1.03) | 0.08906 |
| rs2064501 | 0.8337 (0.6674–1.041) | 0.109 |
| rs11154914 | 1.043 (0.7864–1.384) | 0.7687 |
| rs28385692 | 1.799 (0.8654–3.739) | 0.1158 |
| rs13197049 | 0.846 (0.6766–1.058) | 0.1423 |
| rs6570136 | 0.8669 (0.7018–1.071) | 0.1855 |
| rs7745487 | 1.001 (0.7436–1.347) | 0.9947 |
| Block | Haplotype | Frequency | Case, Control Frequencies | p |
|---|---|---|---|---|
| rs4896239 + rs28385692 | TA | 0.501 | 0.479, 0.519 | 0.102 |
| CG | 0.267 | 0.273, 0.262 | 0.6031 | |
| CA | 0.232 | 0.248, 0.219 | 0.1642 | |
| rs12194034 + rs1543509+rs28366 + rs276466 | TTTA | 0.408 | 0.404, 0.411 | 0.763 |
| TTTG | 0.22 | 0.216, 0.224 | 0.7026 | |
| ATCA | 0.217 | 0.215, 0.218 | 0.8637 | |
| TCTA | 0.148 | 0.160, 0.139 | 0.2266 | |
| rs132117897 + rs202573 + rs2064501 + rs11154914 + rs13197049 | GGCAA | 0.495 | 0.475, 0.512 | 0.1298 |
| AGTAT | 0.193 | 0.178, 0.206 | 0.1565 | |
| GATGA | 0.172 | 0.192, 0.156 | 0.0561 | |
| GATAA | 0.124 | 0.139, 0.111 | 0.0806 | |
| rs6570136 +rs7745487 | GG | 0.551 | 0.543, 0.558 | 0.5396 |
| AG | 0.278 | 0.265, 0.289 | 0.2853 | |
| AA | 0.171 | 0.192, 0.153 | 0.0373 |
| ID Server Name | Individual or Consensus Tool | Website | MLT That Based on | Input Parameters | Deleterious Threshold & Outputs | Ref. |
|---|---|---|---|---|---|---|
| Meta RL & Meta SVM | Consensus | http://annovar.openbioinformatics.org/en/latest/ | MMAF, linear kernel, radial kernel and polynomial kernel | A score of SIFT, PolyPhen-2, GERPþþ, Mutation Taster, Mutation Assessor, FATHMM, LRT, SiPhy and PhyloP | D (Deleterious), N (Neutral) and U (Unknown) | [50] |
| Meta-SNP | Consensus | http://snps.biofold.org/meta-snp/ | RF | A score of PANTHER, PhD-SNP, SIFT, and SNAP | Disease related or Polymorphic non-synonymous SNVs | [32] |
| PredictSNP | Consensus | https://loschmidt.chemi.muni.cz/predictsnp/ | Weighted majority vote consensus | A score of PolyPhen-1, PolyPhen-2, SIFT, MAPP, PhD-SNP and SNAP | Confidence scores and neutral or deleterious | [31] |
| REVEL | Consensus | https://omictools.com/revel-tool | RF | A score of MutPred, FATHMM, VEST, Poly-Phen, SIFT, PROVEAN, Mutation Assessor, Mutation Taster, LRT, GERP, SiPhy, phyloP, and phastCons | Disease variants or rare neutral variants | [51] |
| CADD | Individual | http://cadd.gs.washington.edu/score | A linear kernel support vector machine (SVM) | SNVs | Functional, deleterious, and pathogenic variants | [52] |
| MAPP | Individual | http://mendel.stanford.edu/SidowLab/downloads/MAPP/index.html | Physicochemical properties and alignment score | The original amino acid, the position of the substitution and the new amino acid. | Score (0–1). The predicted is damaging if the score <=0.05 and tolerated if the score >0.05 | [53] |
| Mutation Assessor | Individual | http://mutationassessor.org/r3/ | Conservation method | Genome build, chromosome position, reference allele and substituted allele or Protein ID and variant | (VC) Variant conservation score and (VS) Variant specificity score. Level of functional impact (high, medium, low, neutral) | [54] |
| SIFT | Individual | http://sift.jcvi.org/www/SIFT_help.html#SIFT_OUTPUT | Alignment scores | The original amino acid, the position of the substitution and the new amino acid | Score (0–1). The predicted is damaging if the score <=0.05 and tolerated if the score >0.05 | [55] |
| SNAP2 | Individual | https://rostlab.org/services/snap2web/ | ANN | Protein sequence | Non-neutral and neutral, Score and accuracy | [56] |
| PANTHER | Individual | http://www.pantherdb.org/about.jsp | Alignment Scores HMM | Protein sequence and substitution | All GO annotations & Phylogenetic annotation | [57] |
| PhD-SNPg | Individual | http://snps.biofold.org/phd-snpg/method.html | Gradient boosting algorithm | Chromosome position, and protein variation (position, and first amino acid, and second amino acid variant) | If the probability is >0.5 then the SNV is predicted to be Pathogenic otherwise Benign | [58] |
| Polyphen-2 | Individual | http://genetics.bwh.harvard.edu/pph2/ | Empirical rules | Protein, SNP identifier or Protein sequence in FASTA format and positions of the substitution | Probably damaging or Benign, or Possibly damaging, Sensitivity, specificity, Multiple sequence alignment and 3D Visualization | [59] |






References
- Patsopoulos, N.A.; Baranzini, S.E.; Santaniello, A.; Shoostari, P.; Cotsapas, C.; Wong, G.; Beecham, A.H.; James, T.; Replogle, J.; Vlachos, I.S.; et al. Multiple sclerosis genomic map implicates peripheral immune cells and microglia in susceptibility. Science 2019, 365, eaav7188. [Google Scholar] [CrossRef] [Green Version]
- Beyeen, A.D.; Adzemovic, M.Z.; Ockinger, J.; Stridh, P.; Becanovic, K.; Laaksonen, H.; Lassmann, H.; Harris, R.A.; Hillert, J.; Alfredsson, L.; et al. IL-22RA2 Associates with Multiple Sclerosis and Macrophage Effector Mechanisms in Experimental Neuroinflammation. J. Immunol. 2010, 185, 6883–6890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenbroeck, K.; Alvarez, J.; Swaminathan, B.; Alloza, I.; Matesanz, F.; Urcelay, E.; Comabella, M.; Alcina, A.; Fedetz, M.; Ortiz, M.A.; et al. A cytokine gene screen uncovers SOCS1 as genetic risk factor for multiple sclerosis. Genes Immun. 2012, 13, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beecham, A.H.; Patsopoulos, N.A.; Xifara, D.K.; Davis, M.F.; Kemppinen, A.; Cotsapas, C.; Shah, T.S.; Spencer, C.; Booth, D.; Goris, A.; et al. Analysis of immune-related loci identifies 48 new susceptibility variants for multiple sclerosis. Nat. Genet. 2013, 45, 1353–1362. [Google Scholar] [PubMed]
- Bush, W.S.; Sawcer, S.J.; De Jager, P.L.; The International Multiple Sclerosis Genetics Consortium (IMSGC). A systems biology approach uncovers cell-specific gene regulatory effects of genetic associations in multiple sclerosis. Am. J. Nat. Commun. 2019, 10, 2236. [Google Scholar] [CrossRef] [Green Version]
- Sawcer, S.; Hellenthal, G.; Pirinen, M.; Spencer, C.C.A.; Patsopoulos, N.A.; Moutsianas, L.; Dilthey, A.; Su, Z.; Freeman, C.; Hunt, S.E.; et al. Genetic risk and a primary role for cell-mediated immune mechanisms in multiple sclerosis. Nature 2011, 476, 214–219. [Google Scholar] [PubMed]
- Wolk, K.; Kunz, S.; Witte, E.; Friedrich, M.; Asadullah, K.; Sabat, R. IL-22 Increases the Innate Immunity of Tissues. Immunity 2004, 21, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Wolk, K.; Sabat, R. Interleukin-22: A novel T- and NK-cell derived cytokine that regulates the biology of tissue cells. Cytokine Growth Factor Rev. 2006, 17, 367–380. [Google Scholar] [CrossRef]
- Laaksonen, H.; Guerreiro-Cacais, A.O.; Adzemovic, M.Z.; Parsa, R.; Zeitelhofer, M.; Jagodic, M.; Olsson, T. The multiple sclerosis risk gene IL22RA2 contributes to a more severe murine autoimmune neuroinflammation. Genes Immun. 2014, 15, 457–465. [Google Scholar] [CrossRef]
- Kebir, H.; Kreymborg, K.; Ifergan, I.; Dodelet-Devillers, A.; Cayrol, R.; Bernard, M.; Giuliani, F.; Arbour, N.; Becher, B.; Prat, A. Human TH17 lymphocytes promote blood-brain barrier disruption and central nervous system inflammation. Nat. Med. 2007, 13, 1173–1175. [Google Scholar] [CrossRef] [Green Version]
- Almolda, B.; Costa, M.; Montoya, M.; González, B.; Castellano, B. Increase in Th17 and T-reg Lymphocytes and Decrease of IL22 Correlate with the Recovery Phase of Acute EAE IN Rat. PLoS ONE 2011, 6, e27473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Li, R.; Dai, Y.; Wu, A.; Wang, H.; Cheng, C.; Qiu, W.; Lu, Z.; Zhong, X.; Shu, Y.; et al. IL-22 secreting CD4+ T cells in the patients with neuromyelitis optica and multiple sclerosis. J. Neuroimmunol. 2013, 261, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Perriard, G.; Mathias, A.; Enz, L.; Canales, M.; Schluep, M.; Gentner, M.; Schaeren-Wiemers, N.; Du Pasquier, R.A. Interleukin-22 is increased in multiple sclerosis patients and targets astrocytes. J. Neuroinflammation 2015, 12, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotenko, S.V.; Izotova, L.S.; Mirochnitchenko, O.V.; Esterova, E.; Dickensheets, H.; Donnelly, R.P.; Pestka, S. Identification, Cloning, and Characterization of a Novel Soluble Receptor That Binds IL-22 and Neutralizes Its Activity. J. Immunol. 2001, 166, 7096–7103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumoutier, L.; Lejeune, D.; Colau, D.; Renauld, J.-C. Cloning and Characterization of IL-22 Binding Protein, a Natural Antagonist of IL-10-Related T Cell-Derived Inducible Factor/IL-22. J. Immunol. 2001, 166, 7090–7095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, B.C.; Logsdon, N.J.; Walter, M.R. Structure of IL-22 Bound to Its High-Affinity IL-22R1 Chain. Structure 2008, 16, 1333–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logsdon, N.J.; Jones, B.C.; Josephson, K.; Cook, J.; Walter, M.R. Comparison of Interleukin-22 and Interleukin-10 Soluble Receptor Complexes. J. Interf. Cytokine Res. 2002, 22, 1099–1112. [Google Scholar] [CrossRef] [PubMed]
- Wolk, K.; Witte, E.; Hoffmann, U.; Doecke, W.-D.W.-D.W.-D.; Endesfelder, S.; Asadullah, K.; Sterry, W.; Volk, H.-D.H.-D.H.-D.; Wittig, B.M.; Sabat, R. IL-22 Induces Lipopolysaccharide-Binding Protein in Hepatocytes: A Potential Systemic Role of IL-22 in Crohn’s Disease. J. Immunol. 2007, 178, 5973–5981. [Google Scholar] [CrossRef]
- Lim, C.; Hong, M.; Savan, R. Human IL-22 binding protein isoforms act as a rheostat for IL-22 signaling. Sci. Signal. 2016, 9, ra95. [Google Scholar] [CrossRef]
- Gómez-Fernández, P.; Urtasun, A.; Paton, A.W.; Paton, J.C.; Borrego, F.; Dersh, D.; Argon, Y.; Alloza, I.; Vandenbroeck, K. Long Interleukin-22 Binding Protein Isoform-1 Is an Intracellular Activator of the Unfolded Protein Response. Front. Immunol. 2018, 9, 2934. [Google Scholar] [CrossRef]
- McDonald, W.I.; Compston, A.; Edan, G.; Goodkin, D.; Hartung, H.P.; Lublin, F.D.; McFarland, H.F.; Paty, D.W.; Polman, C.H.; Reingold, S.C.; et al. Recommended diagnostic criteria for multiple sclerosis: Guidelines from the International Panel on the diagnosis of multiple sclerosis. Ann. Neurol. 2001, 50, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Poser, C.M.; Paty, D.W.; Scheinberg, L.; McDonald, W.I.; Davis, F.A.; Ebers, G.C.; Johnson, K.P.; Sibley, W.A.; Silberberg, D.H.; Tourtellotte, W.W. New diagnostic criteria for multiple sclerosis: Guidelines for research protocols. Ann. Neurol. 1983, 13, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Cherny, S.S.; Sham, P.C. Genetic Power Calculator: Design of linkage and association genetic mapping studies of complex traits. Bioinformatics 2003, 19, 149–150. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, S.B.; Schaffner, S.F.; Nguyen, H.; Moore, J.M.; Roy, J.; Blumenstiel, B.; Higgins, J.; DeFelice, M.; Lochner, A.; Faggart, M.; et al. The structure of haplotype blocks in the human genome. Science 2002, 296, 2225–2229. [Google Scholar] [CrossRef] [Green Version]
- Skol, A.D.; Scott, L.J.; Abecasis, G.R.; Boehnke, M. Joint analysis is more efficient than replication-based analysis for two-stage genome-wide association studies. Nat. Genet. 2006, 38, 209–213. [Google Scholar] [CrossRef]
- Johnson, A.D.; Handsaker, R.E.; Pulit, S.L.; Nizzari, M.M.; O’Donnell, C.J.; de Bakker, P.I.W. SNAP: A web-based tool for identification and annotation of proxy SNPs using HapMap. Bioinformatics 2008, 24, 2938–2939. [Google Scholar] [CrossRef] [Green Version]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [Green Version]
- Boyle, A.P.; Hong, E.L.; Hariharan, M.; Cheng, Y.; Schaub, M.A.; Kasowski, M.; Karczewski, K.J.; Park, J.; Hitz, B.C.; Weng, S.; et al. Annotation of functional variation in personal genomes using RegulomeDB. Genome Res. 2012, 22, 1790–1797. [Google Scholar] [CrossRef] [Green Version]
- Bendl, J.; Stourac, J.; Salanda, O.; Pavelka, A.; Wieben, E.D.; Zendulka, J.; Brezovsky, J.; Damborsky, J. PredictSNP: Robust and Accurate Consensus Classifier for Prediction of Disease-Related Mutations. PLoS Comput. Biol. 2014, 10, e1003440. [Google Scholar] [CrossRef] [PubMed]
- Capriotti, E.; Altman, R.B.; Bromberg, Y. Collective judgment predicts disease-associated single nucleotide variants. BMC Genomics 2013, 14, S2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, F.; Achuthan, P.; Akanni, W.; Allen, J.; Amode, M.R.; Armean, I.M.; Bennett, R.; Bhai, J.; Billis, K.; Boddu, S.; et al. Ensembl 2019. Nucleic Acids Res. 2019, 47, D745–D751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bendtsen, J.D.; Nielsen, H.; von Heijne, G.; Brunak, S. Improved prediction of signal peptides: SignalP 3.0. J. Mol. Biol. 2004, 340, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Kall, L.; Krogh, A.; Sonnhammer, E.L.L. Advantages of combined transmembrane topology and signal peptide prediction—The Phobius web server. Nucleic Acids Res. 2007, 35, W429–W432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchan, D.W.A.; Jones, D.T. The PSIPRED Protein Analysis Workbench: 20 years on. Nucleic Acids Res. 2019, 47, 4068–4085. [Google Scholar] [CrossRef] [Green Version]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Hiller, K.; Grote, A.; Scheer, M.; Munch, R.; Jahn, D. PrediSi: Prediction of signal peptides and their cleavage positions. Nucleic Acids Res. 2004, 32, W375–W379. [Google Scholar] [CrossRef]
- Buchan, D.W.A.A.; Minneci, F.; Nugent, T.C.O.O.; Bryson, K.; Jones, D.T. Scalable web services for the PSIPRED Protein Analysis Workbench. Nucleic Acids Res. 2013, 41, 349–357. [Google Scholar] [CrossRef]
- Wang, S.; Li, W.; Liu, S.; Xu, J. RaptorX-Property: A web server for protein structure property prediction. Nucleic Acids Res. 2016, 44, W430–W435. [Google Scholar] [CrossRef]
- Adamczak, R.; Porollo, A.; Meller, J. Accurate prediction of solvent accessibility using neural networks-based regression. Proteins Struct. Funct. Bioinform. 2004, 56, 753–767. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.S.; Shaalan, A.A.; Dessouky, M.I.; Abdelnaiem, A.E.; Abdel-Haleem, D.A.; ElHefnawi, M. Predicting Non-Synonymous Single Nucleotide Variants Pathogenic Effects in Human Diseases. In Encyclopedia of Bioinformatics and Computational Biology; Elsevier Ltd.: Amserdam, The Netherlands, 2018; pp. 400–409. ISBN 9780128096338. [Google Scholar]
- Owji, H.; Nezafat, N.; Negahdaripour, M.; Hajiebrahimi, A.; Ghasemi, Y. A comprehensive review of signal peptides: Structure, roles, and applications. Eur. J. Cell Biol. 2018, 97, 422–441. [Google Scholar] [CrossRef] [PubMed]
- Hegde, R.S.; Bernstein, H.D. The surprising complexity of signal sequences. Trends Biochem. Sci. 2006, 31, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Symoens, S.; Malfait, F.; Renard, M.; André, J.; Hausser, I.; Loeys, B.; Coucke, P.; De Paepe, A. COL5A1 signal peptide mutations interfere with protein secretion and cause classic Ehlers-Danlos syndrome. Hum. Mutat. 2009, 30, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, G.C.; Wang, Q.; Ramamoorthy, P.; Whim, M.D. A Common Single Nucleotide Polymorphism Alters the Synthesis and Secretion of Neuropeptide Y. J. Neurosci. 2008, 28, 14428–14434. [Google Scholar] [CrossRef]
- Lee, S.-K.; Lee, K.-E.; Song, S.J.; Hyun, H.-K.; Lee, S.-H.; Kim, J.-W. A DSPP Mutation Causing Dentinogenesis Imperfecta and Characterization of the Mutational Effect. Biomed. Res. Int. 2013, 2013, 1–7. [Google Scholar]
- Morris, G.E.; Braund, P.S.; Moore, J.S.; Samani, N.J.; Codd, V.; Webb, T.R. Coronary Artery Disease–Associated LIPA Coding Variant rs1051338 Reduces Lysosomal Acid Lipase Levels and Activity in Lysosomes. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1050–1057. [Google Scholar] [CrossRef] [Green Version]
- Lill, C.M.; Schilling, M.; Ansaloni, S.; Schröder, J.; Jaedicke, M.; Luessi, F.; Schjeide, B.-M.M.; Mashychev, A.; Graetz, C.; Akkad, D.A.; et al. Assessment of microRNA-related SNP effects in the 3′ untranslated region of the IL22RA2 risk locus in multiple sclerosis. Neurogenetics 2014, 15, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Wei, P.; Jian, X.; Gibbs, R.; Boerwinkle, E.; Wang, K.; Liu, X. Comparison and integration of deleteriousness prediction methods for nonsynonymous SNVs in whole exome sequencing studies. Hum. Mol. Genet. 2015, 24, 2125–2137. [Google Scholar] [CrossRef] [Green Version]
- Ioannidis, N.M.; Rothstein, J.H.; Pejaver, V.; Middha, S.; McDonnell, S.K.; Baheti, S.; Musolf, A.; Li, Q.; Holzinger, E.; Karyadi, D.; et al. REVEL: An Ensemble Method for Predicting the Pathogenicity of Rare Missense Variants. Am. J. Hum. Genet. 2016, 99, 877–885. [Google Scholar] [CrossRef]
- Rentzsch, P.; Witten, D.; Cooper, G.M.; Shendure, J.; Kircher, M. CADD: Predicting the deleteriousness of variants throughout the human genome. Nucleic Acids Res. 2019, 47, D886–D894. [Google Scholar] [CrossRef] [PubMed]
- Stone, E.A.; Sidow, A. Physicochemical constraint violation by missense substitutions mediates impairment of protein function and disease severity. Genome Res. 2005, 15, 978–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reva, B.; Antipin, Y.; Sander, C. Predicting the functional impact of protein mutations: Application to cancer genomics. Nucleic Acids Res. 2011, 39, e118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.; Henikoff, S.; Ng, P.C. Predicting the effects of coding non-synonymous variants on protein function using the SIFT algorithm. Nat. Protoc. 2009, 4, 1073–1081. [Google Scholar] [CrossRef]
- Bromberg, Y.; Rost, B. SNAP: Predict effect of non-synonymous polymorphisms on function. Nucleic Acids Res. 2007, 35, 3823–3835. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.D.; Kejariwal, A. Coding single-nucleotide polymorphisms associated with complex vs. Mendelian disease: Evolutionary evidence for differences in molecular effects. Proc. Natl. Acad. Sci. USA 2004, 101, 15398–15403. [Google Scholar] [CrossRef] [Green Version]
- Capriotti, E.; Calabrese, R.; Casadio, R. Predicting the insurgence of human genetic diseases associated to single point protein mutations with support vector machines and evolutionary information. Bioinformatics 2006, 22, 2729–2734. [Google Scholar] [CrossRef] [Green Version]
- Adzhubei, I.A.; Schmidt, S.; Peshkin, L.; Ramensky, V.E.; Gerasimova, A.; Bork, P.; Kondrashov, A.S.; Sunyaev, S.R. A method and server for predicting damaging missense mutations. Nat. Methods 2010, 7, 248–249. [Google Scholar] [CrossRef] [Green Version]





| Population | Number (% Female) | Age, Average ± SD 1 | RR 2 & ScP 3/PP 4/Other/ND 5 | Age at Onset, Average ± SD | EDSS 6, Mean ± SD | |
|---|---|---|---|---|---|---|
| Bilbao | Cases | 647 (72.3) | 42.5 ± 12.01 | 79.6/9/1.4/10 | 30.42 ± 10.17 | 2.9 ± 2.3 |
| Controls | 573 (60.3) | 44.2 ± 9 | - | - | - | |
| Donostia | Cases | 572 (64.8) | 46.4 ± 4.8 | 84.8/3.8/4.8/6.6 | 33.01 ± 11.05 | 2.79 ± 2.7 |
| Controls | 250 (66) | 50.52 ± 13.26 | - | - | - | |
| Barcelona | Cases | 676 (63.3) | 40.17 ± 12.93 | 81.5/14.8/3.7 | 31.6 ± 9.9 | 3.91 ± 2.5 |
| Controls | 910 (52.7) | 40.2 ± 12.9 | - | - | - | |
| Madrid | Cases | 899 (63.7) | 44.8 ± 10.55 | 79.7/6.9/4.7/8.7 | 29.8 ± 8.65 | 2.56 ± 2.13 |
| Controls | 697 (55.1) | 40.96 ± 16.71 | - | - | - | |
| Andalucía | Cases | 1474 (61) | 43 ± 12 | 47.4/1/9/42.6 | 28.87 ± 10.25 | ND |
| Controls | 1777 (64.4) | 40.22 ± 12.9 | - | - | - | |
| Germany | Cases | 3762 (70.2) | 42.2 ± 13.6 | ND | ND | ND |
| Controls | 2972 (60.1) | 41.1 ± 14.05 | - | - | - | |
| France | Cases | 1344 (63.6) | 44.3 ± 11.8 | ND | ND | ND |
| Controls | 768 (60.4) | 39.6 ± 13 | - | - | - | |
| SNP | Position 1 | Risk Allele | RAF 2 Cases | RAF Controls | Other Allele | p | OR 3 (95% CI 4) |
|---|---|---|---|---|---|---|---|
| rs4896239 | 137,448,873 | C | 0.52 | 0.50 | T | 0.19 | 1.116 (0.942–1.31) |
| rs17066096 | 137,452,908 | G | 0.29 | 0.27 | A | 0.26 | 1.132 (0.92–1.34) |
| rs12194034 | 137,458,262 | A | 0.23 | 0.22 | T | 0.65 | 1.047 (0.86–1.274) |
| rs1543509 | 137,465,656 | C | 0.15 | 0.14 | T | 0.92 | 1.012 (0.797–1.285) |
| rs28366 | 137,466,087 | C | 0.24 | 0.23 | T | 0.52 | 1.066 (0.88–1.297) |
| rs276466 | 137,466,614 | A | 0.78 | 0.78 | G | 0.99 | 1.001 (0.799–1.25) |
| rs10484798 | 137,470,756 | A | 0.76 | 0.72 | G | 0.05 | 1.23 (1.0–1.508) |
| rs13217897 | 137,471,327 | G | 0.83 | 0.79 | A | 0.02 | 1.291 (1.05–1.591) |
| rs202573 | 137,473,672 | A | 0.33 | 0.28 | G | 0.007 | 1.273 (1.067–1.518) |
| rs2064501 | 137,477,823 | T | 0.50 | 0.49 | C | 0.65 | 1.039 (0.879–1.226) |
| rs11154914 | 137,480,411 | G | 0.19 | 0.16 | A | 0.06 | 1.23 (0.99–1.524) |
| rs28385692 | 137,482,840 | C | 0.02 | 0.01 | T | 0.05 | 1.972 (0.983–3.954) |
| rs13197049 | 137,491,211 | A | 0.83 | 0.80 | T | 0.03 | 1.260 (1.021–1.556) |
| rs6570136 | 137,494,622 | A | 0.46 | 0.45 | G | 0.85 | 1.017 (0.847–1.222) |
| rs7745487 | 137,496,672 | A | 0.18 | 0.15 | G | 0.10 | 1.201 (0.96–1.496) |
| Conditioned to rs17066096 | Conditioned to rs28385692 | ||||
|---|---|---|---|---|---|
| SNP | Reference (minor) Allele | p | OR 1 (95% CI 2) | p | OR (95% CI) |
| rs17066096 | G | NA | NA | 0.001042 | 1.098 (1.039–1.162) |
| rs202573 | A | 0.2424 | 1.029 (0.981–1.079) | 0.3093 | 1.033 (0.9702–1.1) |
| rs28385692 | C | 0.001146 | 1.098 (1.101–1.476) | NA | NA |
| SNP | Proxy | Major Allele | Minor Allele (Frequency) 1 | Ensembl Consequence | SIFT | PolyPhen | RegulomeDB |
|---|---|---|---|---|---|---|---|
| rs10484798 | rs28362847 | G | A (0.21) | regulatory_region_variant | - | - | 5: TF binding or DNase peak |
| rs10484798 | G | A (0.21) | intron_variant | - | - | 6: other | |
| rs13197049 | rs13217897 | G | A (0.17) | intron_variant | - | - | 3a: TF binding + any motif + DNase peak |
| rs17175239 | A | G (0.17) | intergenic_variant | - | - | 5: TF binding or DNase peak | |
| rs1961618 | C | T (0.17) | intron_variant | - | - | 5: TF binding or DNase peak | |
| rs12664889 | C | A (0.17) | intron_variant | - | - | 7: no data | |
| rs13197049 | A | T (0.17) | intron variant | - | - | 7: no data | |
| rs11154913 | A | G (0.17) | intron_variant | - | - | 5: TF binding or DNase peak | |
| rs13193435 | C | A (0.17) | intron_variant | - | - | 5: TF binding or DNase peak | |
| rs7749054 | T | G (0.17) | intergenic_variant | - | - | 6: other | |
| rs13197049 | A | T (0.17) | intron_variant | - | - | 7: no data | |
| rs7766677 | A | C (0.17) | intergenic_variant | - | - | 7: no data | |
| rs13217897 | rs13217897 | G | A (0.17) | intron_variant | - | - | 3a: TF binding + any motif + DNase peak |
| rs13193435 | C | A (0.17) | intron variant | - | - | 5: TF binding or DNase peak | |
| rs1961618 | C | T (0.17) | intron_variant | - | - | 5: TF binding or DNase peak | |
| rs17175239 | A | G (0.17) | intergenic_variant | - | - | 5: TF binding or DNase peak | |
| rs7766677 | A | C (0.17) | intergenic_variant | - | - | 6: other | |
| rs11154913 | A | G (0.17) | intron_variant | - | - | 7: no data | |
| rs7749054 | T | G (0.17) | intergenic_variant | - | - | 7: no data | |
| rs12664889 | C | A (0.17) | intron_variant | - | - | 7: no data | |
| rs13197049 | A | T (0.17) | intron_variant | - | - | 7: no data | |
| rs17066096 | rs17066063 | G | A (0.23) | TF_binding_site_variant | - | - | 3a: TF binding + any motif + DNase peak |
| rs62420820 | G | A (0.23) | regulatory_region_variant | - | - | 3a: TF binding + any motif + DNase peak | |
| rs72975618 | C | T (0.23) | TF_binding_site_variant | - | - | 4: TF binding + DNase peak | |
| rs1322553 | A | G (0.23) | regulatory_region_variant | - | - | 5: TF binding or DNase peak | |
| rs12214115 | G | T (0.23) | regulatory_region_variant | - | - | 5: TF binding or DNase peak | |
| rs12214014 | C | T (0.23) | regulatory_region_variant | - | - | 5: TF binding or DNase peak | |
| rs17066096 | A | G (0.23) | intergenic_variant | - | - | 6: other | |
| rs202573 | rs202571 | T | C (0.31) | intron_variant | - | - | 7: no data |
| rs202573 | G | A (0.31) | intron_variant | - | - | 7: no data | |
| rs28385692 | rs28385692 | T | C (0.03) | missense_variant | Tolerated (0.11) | Benign (0.376) | 5: TF binding or DNase peak |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Fernández, P.; Lopez de Lapuente Portilla, A.; Astobiza, I.; Mena, J.; Urtasun, A.; Altmann, V.; Matesanz, F.; Otaegui, D.; Urcelay, E.; Antigüedad, A.; et al. The Rare IL22RA2 Signal Peptide Coding Variant rs28385692 Decreases Secretion of IL-22BP Isoform-1, -2 and -3 and Is Associated with Risk for Multiple Sclerosis. Cells 2020, 9, 175. https://doi.org/10.3390/cells9010175
Gómez-Fernández P, Lopez de Lapuente Portilla A, Astobiza I, Mena J, Urtasun A, Altmann V, Matesanz F, Otaegui D, Urcelay E, Antigüedad A, et al. The Rare IL22RA2 Signal Peptide Coding Variant rs28385692 Decreases Secretion of IL-22BP Isoform-1, -2 and -3 and Is Associated with Risk for Multiple Sclerosis. Cells. 2020; 9(1):175. https://doi.org/10.3390/cells9010175
Chicago/Turabian StyleGómez-Fernández, Paloma, Aitzkoa Lopez de Lapuente Portilla, Ianire Astobiza, Jorge Mena, Andoni Urtasun, Vivian Altmann, Fuencisla Matesanz, David Otaegui, Elena Urcelay, Alfredo Antigüedad, and et al. 2020. "The Rare IL22RA2 Signal Peptide Coding Variant rs28385692 Decreases Secretion of IL-22BP Isoform-1, -2 and -3 and Is Associated with Risk for Multiple Sclerosis" Cells 9, no. 1: 175. https://doi.org/10.3390/cells9010175
APA StyleGómez-Fernández, P., Lopez de Lapuente Portilla, A., Astobiza, I., Mena, J., Urtasun, A., Altmann, V., Matesanz, F., Otaegui, D., Urcelay, E., Antigüedad, A., Malhotra, S., Montalban, X., Castillo-Triviño, T., Espino-Paisán, L., Aktas, O., Buttmann, M., Chan, A., Fontaine, B., Gourraud, P. -A., ... Vandenbroeck, K. (2020). The Rare IL22RA2 Signal Peptide Coding Variant rs28385692 Decreases Secretion of IL-22BP Isoform-1, -2 and -3 and Is Associated with Risk for Multiple Sclerosis. Cells, 9(1), 175. https://doi.org/10.3390/cells9010175