Neutrophil Extracellular Traps (NETs) in the Cerebrospinal Fluid Samples from Children and Adults with Central Nervous System Infections

 , ,
, , 
 ,
,  and
and
Abstract
:1. Introduction
2. Patients and Methods
2.1. Paediatric Patients
2.2. Adult Patients
2.3. NET Remnant Assay (NETs)
2.4. Neutrophil Elastase (NE) Activity Assay
2.5. Cytokine and Chemokine Assays
2.6. Statistics
2.7. Ethics
3. Results
3.1. CSF-NETs in Relation to Patient Diagnoses
3.2. CSF-NE Activity
3.3. NETs in Relation to Clinical Outcome in Children
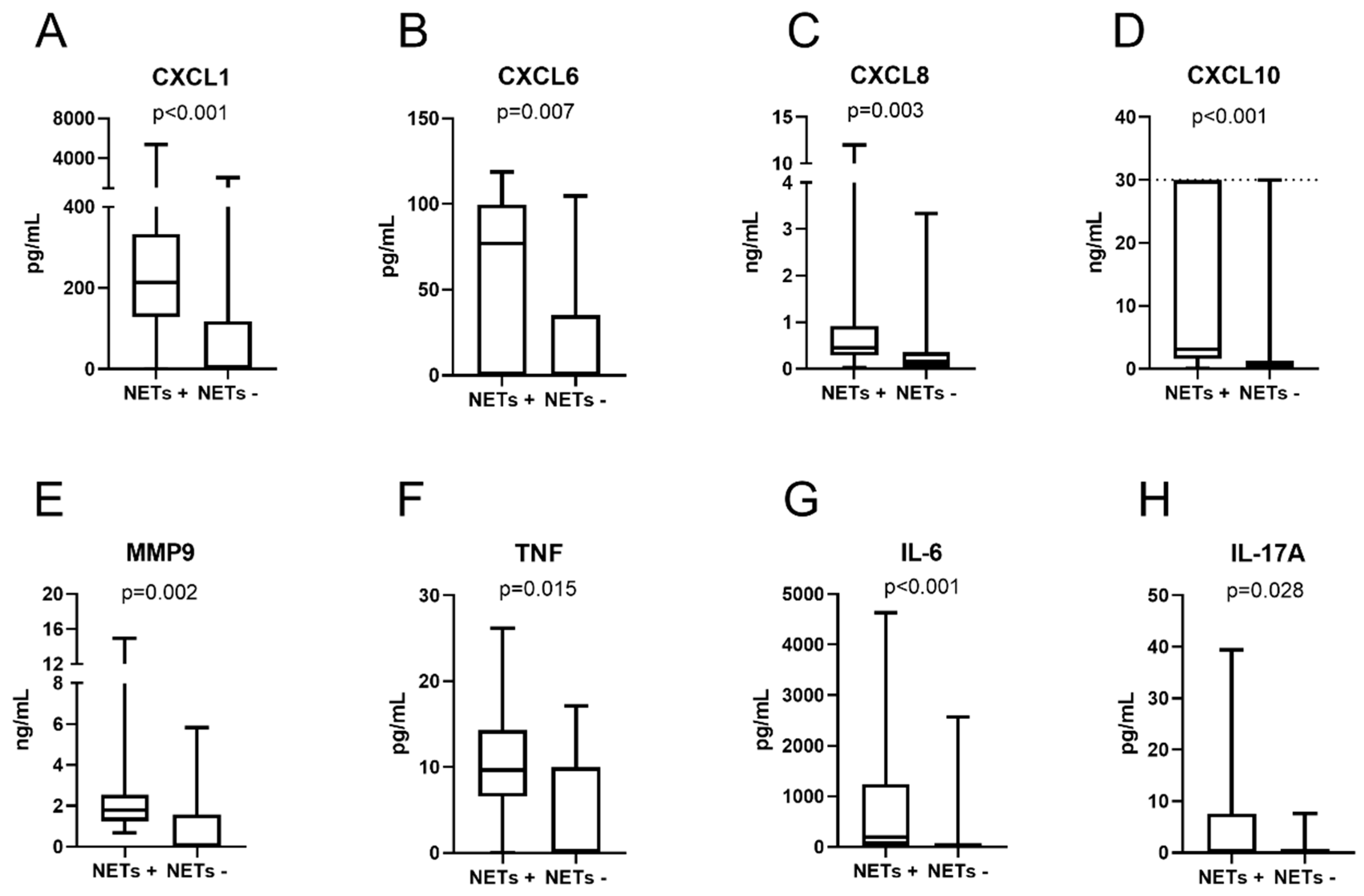
3.4. Cytokines and Chemokines
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Halperin, J.J. Lyme neuroborreliosis. Curr. Opin. Infect. Dis. 2019, 32, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Henningsson, A.J.; Malmvall, B.E.; Ernerudh, J.; Matussek, A.; Forsberg, P. Neuroborreliosis-An epidemiological, clinical and healthcare cost study from an endemic area in the south-east of Sweden. Clin. Microbiol. Infect. 2010, 16, 1245–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skogman, B.H.; Croner, S.; Nordwall, M.; Eknefelt, M.; Ernerudh, J.; Forsberg, P. Lyme neuroborreliosis in children: A prospective study of clinical features, prognosis, and outcome. Pediatr. Infect. Dis. J. 2008, 27, 1089–1094. [Google Scholar] [CrossRef] [PubMed]
- Sodermark, L.; Sigurdsson, V.; Nas, W.; Wall, P.; Trollfors, B. Neuroborreliosis in Swedish Children: A Population-based Study on Incidence and Clinical Characteristics. Pediatr. Infect. Dis. J. 2017, 36, 1052–1056. [Google Scholar] [CrossRef] [PubMed]
- Dahl, V.; Wisell, K.T.; Giske, C.G.; Tegnell, A.; Wallensten, A. Lyme neuroborreliosis epidemiology in Sweden 2010 to 2014: Clinical microbiology laboratories are a better data source than the hospital discharge diagnosis register. Euro Surveill. 2019, 24, 20. [Google Scholar] [CrossRef] [Green Version]
- Pietikainen, A.; Oksi, J.; Hytonen, J. Point-of-care testing for CXCL13 in Lyme neuroborreliosis. Diagn Microbiol. Infect. Dis. 2018, 91, 226–228. [Google Scholar] [CrossRef]
- Mygland, A.; Ljostad, U.; Fingerle, V.; Rupprecht, T.; Schmutzhard, E.; Steiner, I.; European Federation of Neurological Societies. EFNS guidelines on the diagnosis and management of European Lyme neuroborreliosis. Eur. J. Neurol. 2010, 17, 8–16. [Google Scholar] [CrossRef]
- Shah, S.S.; Zaoutism, T.E.; Turnquist, J.; Hodinka, R.L.; Coffin, S.E. Early differentiation of Lyme from enteroviral meningitis. Pediatr. Infect. Dis. J. 2005, 24, 542–545. [Google Scholar] [CrossRef]
- Salazar, J.C.; Pope, C.D.; Sellati, T.J.; Feder, H.M.; Jr Kiely, T.G.; Dardick, K.R.; Buckman, R.L.; Moore, M.W.; Caimano, M.J.; Pope, J.G.; et al. Coevolution of markers of innate and adaptive immunity in skin and peripheral blood of patients with erythema migrans. J. Immunol. 2003, 171, 2660–2670. [Google Scholar] [CrossRef] [Green Version]
- Liew, P.X.; Kubes, P. The Neutrophil’s Role During Health and Disease. Physiol. Rev. 2019, 99, 1223–1248. [Google Scholar] [CrossRef]
- Kruger, P.; Saffarzadeh, M.; Weber, A.N.; Rieber, N.; Radsak, M.; von Bernuth, H.; Benarafa, C.; Roos, D.; Skokowa, J. Hartl DNeutrophils: Between host defence immune modulation,, tissue, injury. PLoS Pathog. 2015, 11, e1004651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V. Neutrophil Extracellular Traps in the Second Decade. J. Innate Immun. 2018, 10, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, A.; Gruneboom, A.; Petru, L.; Podolska, M.J.; Kling, L.; Maueroder, C.; Dahms, F.; Christiansen, S.; Günter, L.; Krenn, V.; et al. Frontline Science: Aggregated neutrophil extracellular traps prevent inflammation on the neutrophil-rich ocular surface. J. Leukoc. Biol. 2019, 105, 1087–1098. [Google Scholar] [CrossRef]
- Munoz, L.E.; Boeltz, S.; Bilyy, R.; Schauer, C.; Mahajan, A.; Widulin, N.; Grüneboom, A.; Herrmann, I.; Boada, E.; Rauh, M.; et al. Neutrophil Extracellular Traps Initiate Gallstone Formation. Immunity 2019, 51, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Schonrich, G.; Raftery, M.J. Neutrophil Extracellular Traps Go Viral. Front. Immunol. 2016, 7, 366. [Google Scholar] [CrossRef] [Green Version]
- Menten-Dedoyart, C.; Faccinetto, C.; Golovchenko, M.; Dupiereux, I.; Van Lerberghe, P.B.; Dubois, S.; Desmet, C.; Elmoualij, B.; Baron, F.; Rudenko, N.; et al. Neutrophil extracellular traps entrap and kill Borrelia burgdorferi sensu stricto spirochetes and are not affected by Ixodes ricinus tick saliva. J. Immunol. 2012, 189, 5393–5401. [Google Scholar] [CrossRef] [Green Version]
- De Buhr, N.; Reuner, F.; Neumann, A.; Stump-Guthier, C.; Tenenbaum, T.; Schroten, H.; Ishikawa, H.; Müller, K.; Beineke, A.; Hennig-Pauka, I.; et al. Neutrophil extracellular trap formation in the Streptococcus suis-infected cerebrospinal fluid compartment. Cell Microbiol. 2017, 19, 12649. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, T.; Fisher, J.; Bakochi, A.; Neumann, A.; Cardoso, J.F.P.; Karlsson, C.A.Q.; Pavan, C.; Lundgaard, I.; Nilson, B.; Peter, R.; et al. Neutrophil extracellular traps in the central nervous system hinder bacterial clearance during pneumococcal meningitis. Nat. Commun. 2019, 10, 1667. [Google Scholar] [CrossRef]
- Knudtzen, F.C.; Andersen, N.S.; Jensen, T.G.; Skarphedinsson, S. Characteristics and Clinical Outcome of Lyme Neuroborreliosis in a High Endemic Area, 1995–2014: A Retrospective Cohort Study in Denmark. Clin. Infect. Dis. 2017, 65, 1489–1495. [Google Scholar] [CrossRef] [PubMed]
- Schauer, C.; Janko, C.; Munoz, L.E.; Zhao, Y.; Kienhofer, D.; Frey, B. Aggregated neutrophil extracellular traps limit inflammation by degrading cytokines and chemokines. Nat. Med. 2014, 20, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Backman, K.; Skogman, B.H. Occurrence of erythema migrans in children with Lyme neuroborreliosis and the association with clinical characteristics and outcome—A prospective cohort study. BMC Pediatr. 2018, 18, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, K.; Lebech, A.M. Lyme neuroborreliosis: A new sensitive diagnostic assay for intrathecal synthesis of Borrelia burgdorferi-specific immunoglobulin GA. and M. Ann. Neurol. 1991, 30, 197–205. [Google Scholar] [CrossRef]
- Rupprecht, T.A.; Manz, K.M.; Fingerle, V.; Lechner, C.; Klein, M.; Pfirrmann, M.; Koedel, U. Diagnostic value of cerebrospinal fluid CXCL13 for acute Lyme neuroborreliosis. A systematic review and meta-analysis. Clin. Microbiol. Infect. 2018, 24, 1234–1240. [Google Scholar] [CrossRef] [Green Version]
- Soderberg, D.; Kurz, T.; Motamedi, A.; Hellmark, T.; Eriksson, P.; Segelmark, M. Increased levels of neutrophil extracellular trap remnants in the circulation of patients with small vessel vasculitis, but an inverse correlation to anti-neutrophil cytoplasmic antibodies during remission. Rheumatolog (Oxf.) 2015, 54, 2085–2094. [Google Scholar] [CrossRef] [Green Version]
- Kessenbrock, K.; Krumbholz, M.; Schonermarck, U.; Back, W.; Gross, W.L.; Werb, Z.; Gröne, H.J.; Brinkmann, V.; Jenne, D.E. Netting neutrophils in autoimmune small-vessel vasculitis. Nat. Med. 2009, 15, 623–625. [Google Scholar] [CrossRef]
- Appelgren, D.; Dahle, C.; Knopf, J.; Bilyy, R.; Vovk, V.; Sundgren, P.C.; Bengtsson, A.A.; Wetterö, J.; Muñoz, L.E.; Herrmann, M.; et al. Active NET formation in Libman-Sacks endocarditis without antiphospholipid antibodies: A dramatic onset of systemic lupus erythematosus. Autoimmunity 2018, 51, 310–318. [Google Scholar] [CrossRef] [Green Version]
- Rumble, J.M.; Huber, A.K.; Krishnamoorthy, G.; Srinivasan, A.; Giles, D.A.; Zhang, X.; Wang, L.; Segal, B.M. Neutrophil-related factors as biomarkers in EAE and MS. J. Exp. Med. 2015, 212, 23–35. [Google Scholar] [CrossRef] [Green Version]
- Michael, M.; Vermeren, S. A neutrophil-centric view of chemotaxis. Essays Biochem. 2019, 63, 607–618. [Google Scholar]
- Isailovic, N.; Daigo, K.; Mantovani, A.; Selmi, C. Interleukin-17 and innate immunity in infections and chronic inflammation. J. Autoimmun. 2015, 60, 1–11. [Google Scholar] [CrossRef]
- Hellberg, S.; Eklund, D.; Gawel, D.R.; Kopsen, M.; Zhang, H.; Nestor, C.E.; Kockum, I.; Olsson, T.; Skogh, T.; Kastbom, A.; et al. Dynamic Response Genes in CD4+ T Cells Reveal a Network of Interactive Proteins that Classifies Disease Activity in Multiple Sclerosis. Cell Rep. 2016, 16, 2928–2939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neil, L.J.; Kaplan, M.J.; Carmona-Rivera, C. The Role of Neutrophils and Neutrophil Extracellular Traps in Vascular Damage in Systemic Lupus Erythematosus. J. Clin. Med. 2019, 8, 1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papayannopoulos, V.; Metzler, K.D.; Hakkim, A.; Zychlinsky, A. Neutrophil elastase and myeloperoxidase regulate the formation of neutrophil extracellular traps. J. Cell Biol. 2010, 191, 677–691. [Google Scholar] [CrossRef] [Green Version]
- Makis, A.; Shipway, D.; Hatzimichael, E.; Galanakis, E.; Pshezhetskiy, D.; Chaliasos, N.; Stebbing, J.; Siamopoulou, A. Cytokine and adhesion molecule expression evolves between the neutrophilic and lymphocytic phases of viral meningitis. J. Interferon Cytokine Res. 2010, 30, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Gyllemark, P.; Forsberg, P.; Ernerudh, J.; Henningsson, A.J. Intrathecal Th17- and B cell-associated cytokine and chemokine responses in relation to clinical outcome in Lyme neuroborreliosis: A large retrospective study. J. Neuroinflamm. 2017, 14, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietikainen, A.; Maksimow, M.; Kauko, T.; Hurme, S.; Salmi, M.; Hytonen, J. Cerebrospinal fluid cytokines in Lyme neuroborreliosis. J. Neuroinflamm. 2016, 13, 273. [Google Scholar] [CrossRef] [Green Version]
- Cerar, T.; Ogrinc, K.; Lotric-Furlan, S.; Kobal, J.; Levicnik-Stezinar, S.; Strle, F.; Ruzić-Sabljic, E. Diagnostic value of cytokines and chemokines in lyme neuroborreliosis. Clin. Vaccine Immunol. 2013, 20, 1578–1584. [Google Scholar] [CrossRef]
- Eckman, E.A.; Pacheco-Quinto, J.; Herdt, A.R.; Halperin, J.J. Neuroimmunomodulators in Neuroborreliosis and Lyme Encephalopathy. Clin. Infect. Dis. 2018, 67, 80–88. [Google Scholar] [CrossRef]
- Park, S.E.; Shin, K.; Song, D.; Nam, S.O.; Kim, K.M.; Lyu, S.Y. Comparison of Cerebrospinal Fluid Cytokine Levels in Children of Enteroviral Meningitis With Versus Without Pleocytosis. J. Interferon Cytokine Res. 2018, 38, 348–355. [Google Scholar] [CrossRef] [Green Version]
- Maric, L.S.; Lepej, S.Z.; Gorenec, L.; Grgic, I.; Trkulja, V.; Rode, O.D.; Roglic, S.; Grmoja, T.; Barisic, N.; Tesovic, G. Chemokines CXCL10, CXCL11, and CXCL13 in acute disseminated encephalomyelitis, non-polio enterovirus aseptic meningitis, and neuroborreliosis: CXCL10 as initial discriminator in diagnostic algorithm? Neurol Sci. 2018, 39, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Brissette, C.A.; Kees, E.D.; Burke, M.M.; Gaultney, R.A.; Floden, A.M.; Watt, J.A. The multifaceted responses of primary human astrocytes and brain microvascular endothelial cells to the Lyme disease spirochete, Borrelia burgdorferi. ASN Neuro 2013, 5, 221–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavcic, A.; Tesovic, G.; Gorenec, L.; Grgic, I.; Benic, B.; Lepej, S.Z. Concentration gradient of CXCL10 and CXCL11 between the cerebrospinal fluid and plasma in children with enteroviral aseptic meningitis. Eur. J. Paediatr. Neurol. 2011, 15, 502–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henningsson, A.J.; Tjernberg, I.; Malmvall, B.E.; Forsberg, P.; Ernerudh, J. Indications of Th1 and Th17 responses in cerebrospinal fluid from patients with Lyme neuroborreliosis: A large retrospective study. J. Neuroinflamm. 2011, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- Zajkowska, J.; Moniuszko-Malinowska, A.; Pancewicz, S.A.; Muszynska-Mazur, A.; Kondrusik, M.; Grygorczuk, S.; Swierzbińska-Pijanowska, R.; Dunaj, J.; Czupryna, P. Evaluation of CXCL10, CXCL11, CXCL12 and CXCL13 chemokines in serum and cerebrospinal fluid in patients with tick borne encephalitis (TBE). Adv. Med. Sci. 2011, 56, 311–317. [Google Scholar] [CrossRef]
- Evans, S.S.; Repasky, E.A.; Fisher, D.T. Fever and the thermal regulation of immunity: The immune system feels the heat. Nat. Rev. Immunol. 2015, 15, 335–349. [Google Scholar] [CrossRef]
- Sadik, C.D.; Kim, N.D.; Luster, A.D. Neutrophils cascading their way to inflammation. Trends Immunol. 2011, 32, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Simmons, S.B.; Liggitt, D.; Goverman, J.M. Cytokine-regulated neutrophil recruitment is required for brain but not spinal cord inflammation during experimental autoimmune encephalomyelitis. J. Immunol. 2014, 193, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Morichi, S.; Urabe, T.; Morishita, N.; Takeshita, M.; Ishida, Y.; Oana, S.; Yamanaka, G.; Kashiwagi, Y.; Kawashima, H. Pathological analysis of children with childhood central nervous system infection based on changes in chemokines and interleukin-17 family cytokines in cerebrospinal fluid. J. Clin. Lab Anal. 2018, 32, 521784686. [Google Scholar] [CrossRef] [Green Version]
- Hartiala, P.; Hytonen, J.; Suhonen, J.; Lepparanta, O.; Tuominen-Gustafsson, H.; Viljanen, M.K. Borrelia burgdorferi inhibits human neutrophil functions. Microbes Infect. 2008, 10, 60–68. [Google Scholar] [CrossRef]
- Carrasco, S.E.; Troxell, B.; Yang, Y.; Brandt, S.L.; Li, H.; Sandusky, G.E.; Condon, K.W.; Serezani, C.H.; Yang, X.F. Outer surface protein OspC is an antiphagocytic factor that protects Borrelia burgdorferi from phagocytosis by macrophages. Infect. Immun. 2015, 83, 4848–4860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doster, R.S.; Rogers, L.M.; Gaddy, J.A.; Aronoff, D.M. Macrophage Extracellular Traps: A Scoping Review. J. Innate Immun. 2018, 10, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Granger, V.; Faille, D.; Marani, V.; Noel, B.; Gallais, Y.; Szely, N.; Flament, H.; Pallardy, M.; Chollet-Martin, S.; de Chaisemartin, L. Human blood monocytes are able to form extracellular traps. J. Leukoc. Biol. 2017, 102, 775–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| On Admission | LNB (N = 28) | TBE (N = 3) | EVM (N = 7) | OVI (N = 4) | Other Disorder (N = 69) | |
|---|---|---|---|---|---|---|
| Age | Median (range) | 10 (3–15) | 10 (3–15) | 10 (4–15) | 4 (1–14) | 13 (0–19) |
| Sex | Female, N (%) | 14 (50) | 0 | 2 (30) | 3 (75) | 44 (64) |
| Duration of symptoms | <1 week, N (%) | 16 (59) | 1 (50) | 5 (83) | 1 (100) | 9 (22) |
| 1–4 weeks, N (%) | 9 (33) | 1 (50) | 1 (17) | 0 | 11 (27) | |
| 1–2 months, N (%) | 1 (4) | 0 | 0 | 0 | 5 (12) | |
| >2 months, N (%) | 1 (4) | 0 | 0 | 0 | 16 (39) | |
| Clinical features | Facial nerve palsy, N (%) | 22 (79) | 0 (0) | 0 (0) | 1 (25) | 17 (25) |
| Meningeal symptoms ¤, N (%) | 22 (79) | 2 (67) | 7 (100) | 1(25) | 50 (72) | |
| Fever >38 °C, N (%) | 13 (46) | 2 (67) | 7 (100) | 2 (50) | 7 (10) | |
| Fatigue, N (%) | 23 (82) | 3 (100) | 6 (86) | 4 (100) | 38 (55) | |
| Laboratory findings | Pleocytosis *, median (range) | 157 (20–486) | 100 (0–130) | 156 (20–634) | 0 (0–6) | 0 (0–74) |
| CSF mono, median (range) | 149 (8–484) | 77 (0–116) | 86 (16–610) | 6 | 0 (0–40) | |
| CSF poly, median (range) | 4 (0–30) | 14 (0–28) | 24 (4–164) | 0 | 0 (0–34) | |
| Recovery at follow-up: | ||||||
| 2 months | Yes, N (%) | 25 (89) | 2 (67) | 7 (100) | 2 (50) | 45 (65) |
| On Admission | LNB (N = 32) | LD Unspec. (N = 6) | TBE (N = 3) | OVM (N = 2) | Other Disorders (N = 21) | |
|---|---|---|---|---|---|---|
| Age | Median (range) | 58 (18–82) | 52 (30–80) | 50 (46–57) | 60 (50–69) | 51 (24–88) |
| Sex | Female, N (%) | 15 (47) | 6 (100) | 1 (33) | 1 (50) | 13 (62) |
| Duration of symptoms | <1 week, N (%) | 4 (13) | 0 (0) | 0 | 0 | 1 (5) |
| 1–4 weeks, N (%) | 7 (23) | 1 (20) | 3 (100) | 1 (50) | 1 (5) | |
| 1–2 months, N (%) | 10 (31) | 1 (20) | 0 | 0 | 1 (5) | |
| >2 months, N (%) | 11 (34) | 3 (60) | 0 | 1 (50) | 18 (86) | |
| Clinical features | Facial nerve palsy, N (%) | 5 (16) | 1 (17) | 0 | 0 | 0 |
| Meningeal symptoms ¤, N (%) | 15 (47) | 3 (50) | 2 (67) | 2 (100) | 10 (48) | |
| Fever >38 °C, N (%) | 3 (9) | 0 | 3 (100) | 1 (50) | 3 (14) | |
| Fatigue, N (%) | 17 (53) | 5 (83) | 3 (100) | 1 (50) | 9 (43) | |
| Radiating pain, N (%) | 22 (69) | 2 (33) | 0 | 1 (50) | 9 (43) | |
| Laboratory findings | Pleocytosis *, median (range) | 43 (6–390) | 5 (4–5) | 82 (51–131) | 33 (18–47) | 6 (0–292) |
| CSF mono, median (range) | 67 (5–355) | NA | 62 (49–118) | 26 (6–45) | 54 (17–91) & | |
| CSF poly, median (range) | 3 (0–45) | NA | 13 (2–20) | 7 (2–12) | 106 (10–202) & | |
| Recovery at follow-up: | ||||||
| 3 weeks | Yes, N (%) | 12 (38) | 3 (50) | 0 | 1 (50) | 4 (20) |
| 6 months | Yes, N (%) | 24 (77) | 6 (100) | 2 (67) | 1 (50) | 8 (42) |
| NETs (Remnants) | NETs (Elastase) † | Cells (Total) | Polynuclear Cells | Mononuclear Cells | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Analyte | Quantified * | rs | p-Value | rs | p-Value | rs | p-Value | rs | p-Value | rs | p-Value |
| CXCL1 | 21/38 | 0.61 | <0.001 | 0.76 | <0.001 | 0.52 | 0.006 | 0.69 | <0.001 | 0.48 | 0.02 |
| CXCL6 | 14/38 | 0.52 | 0.007 | 0.65 | 0.002 | 0.72 | <0.001 | 0.74 | <0.001 | 0.67 | <0.001 |
| CXCL8 | 38/38 | 0.53 | 0.006 | 0.64 | 0.003 | 0.60 | <0.001 | 0.65 | <0.001 | 0.57 | 0.002 |
| CXCL10 | 32/38 | 0.56 | 0.002 | 0.66 | 0.002 | 0.79 | <0.001 | 0.81 | <0.001 | 0.76 | <0.001 |
| MMP9 | 25/38 | 0.53 | 0.006 | N/A | n.s. | 0.70 | <0.001 | 0.65 | <0.001 | 0.68 | <0.001 |
| TNF | 22/37 | N/A | n.s. | N/A | n.s. | 0.81 | <0.001 | 0.65 | <0.001 | 0.80 | <0.001 |
| IL-6 | 28/37 | 0.70 | <0.001 | 0.81 | <0.001 | 0.63 | <0.001 | 0.73 | <0.001 | 0.59 | <0.001 |
| IL-17A | 4/37 | N/A | n.s. | N/A | n.s. | N/A | n.s. | 0.46 | 0.03 | N/A | n.s. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Appelgren, D.; Enocsson, H.; Skogman, B.H.; Nordberg, M.; Perander, L.; Nyman, D.; Nyberg, C.; Knopf, J.; Muñoz, L.E.; Sjöwall, C.; et al. Neutrophil Extracellular Traps (NETs) in the Cerebrospinal Fluid Samples from Children and Adults with Central Nervous System Infections. Cells 2020, 9, 43. https://doi.org/10.3390/cells9010043
Appelgren D, Enocsson H, Skogman BH, Nordberg M, Perander L, Nyman D, Nyberg C, Knopf J, Muñoz LE, Sjöwall C, et al. Neutrophil Extracellular Traps (NETs) in the Cerebrospinal Fluid Samples from Children and Adults with Central Nervous System Infections. Cells. 2020; 9(1):43. https://doi.org/10.3390/cells9010043
Chicago/Turabian StyleAppelgren, Daniel, Helena Enocsson, Barbro H. Skogman, Marika Nordberg, Linda Perander, Dag Nyman, Clara Nyberg, Jasmin Knopf, Luis E. Muñoz, Christopher Sjöwall, and et al. 2020. "Neutrophil Extracellular Traps (NETs) in the Cerebrospinal Fluid Samples from Children and Adults with Central Nervous System Infections" Cells 9, no. 1: 43. https://doi.org/10.3390/cells9010043
APA StyleAppelgren, D., Enocsson, H., Skogman, B. H., Nordberg, M., Perander, L., Nyman, D., Nyberg, C., Knopf, J., Muñoz, L. E., Sjöwall, C., & Sjöwall, J. (2020). Neutrophil Extracellular Traps (NETs) in the Cerebrospinal Fluid Samples from Children and Adults with Central Nervous System Infections. Cells, 9(1), 43. https://doi.org/10.3390/cells9010043