The FtsZ Homolog, FszB, Inhibits Mitochondrial Dynamics in Dictyostelium discoideum


Abstract
:1. Introduction
2. Materials and Methods
2.1. Creation of FszB-GFP Strain
2.2. Imaging Mitochondrial Dynamics
2.3. Disruption of Actin with Latrunculin-B
2.4. Disruption of the Microtubules with Nocodazole
2.5. Analyzing Morphology and Size
2.6. Kymographs and Quantifying Motility Rates
2.7. Counting Rates of Fission, Fusion
2.8. Determining FszB Localization
3. Results
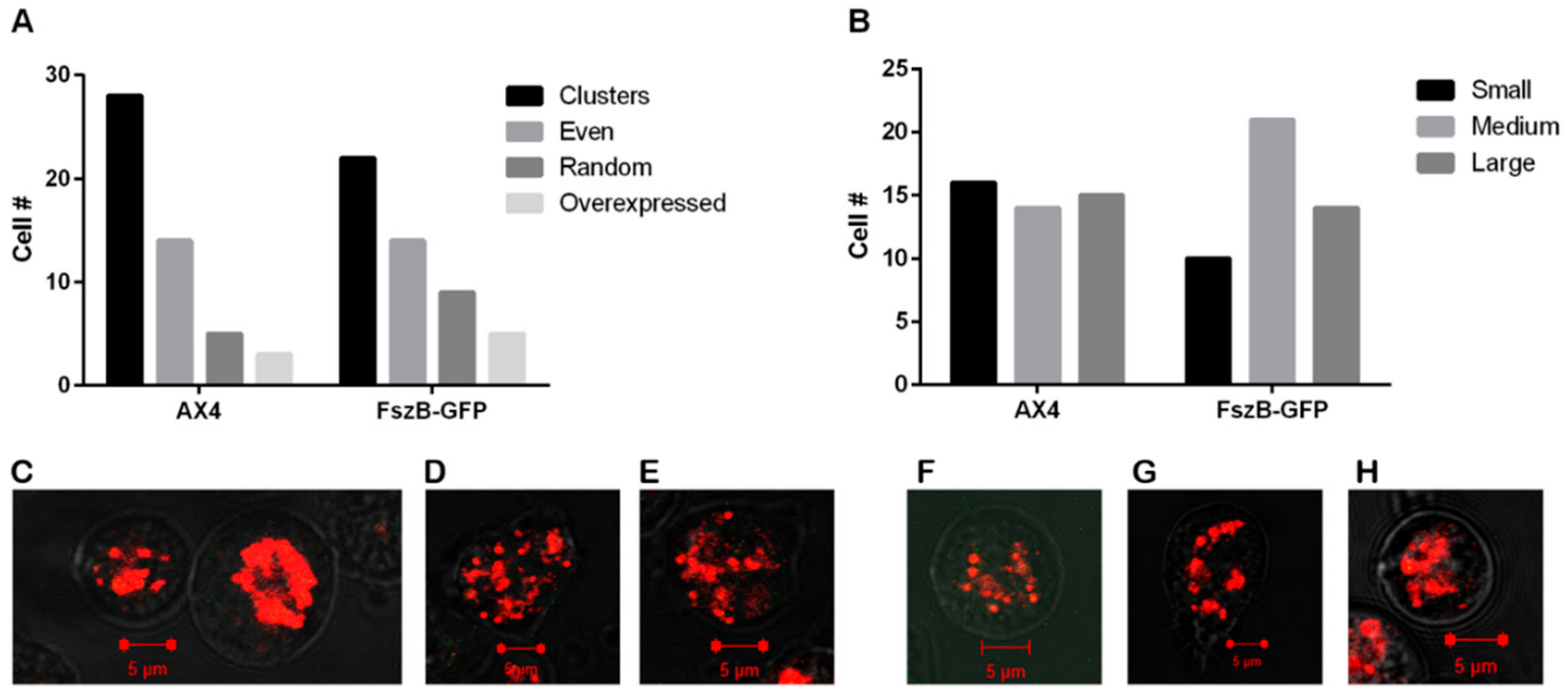
3.1. FszB-GFP Overexpression Does Not Change Mitochondrial Distribution nor Size
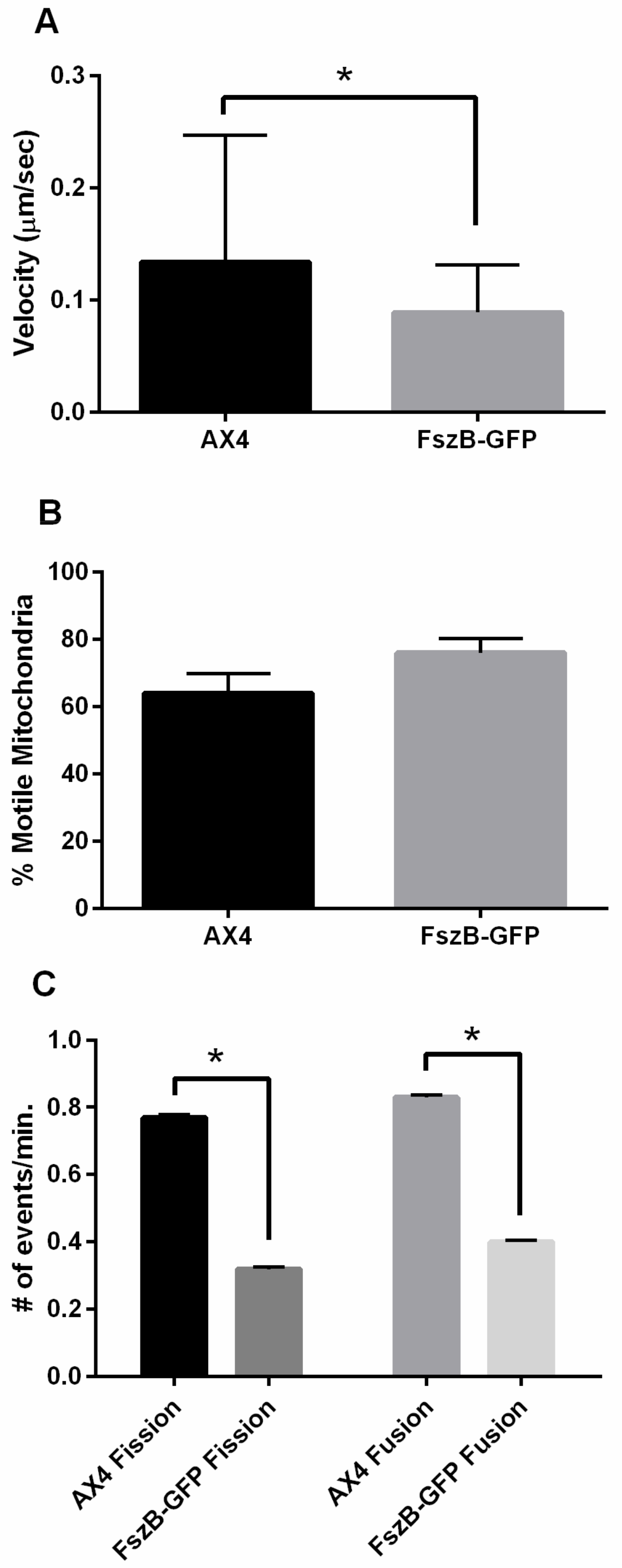
3.2. Overexpressed FszB-GFP Inhibits Mitochondrial Dynamics but Localizes to Fission and Fusion Events
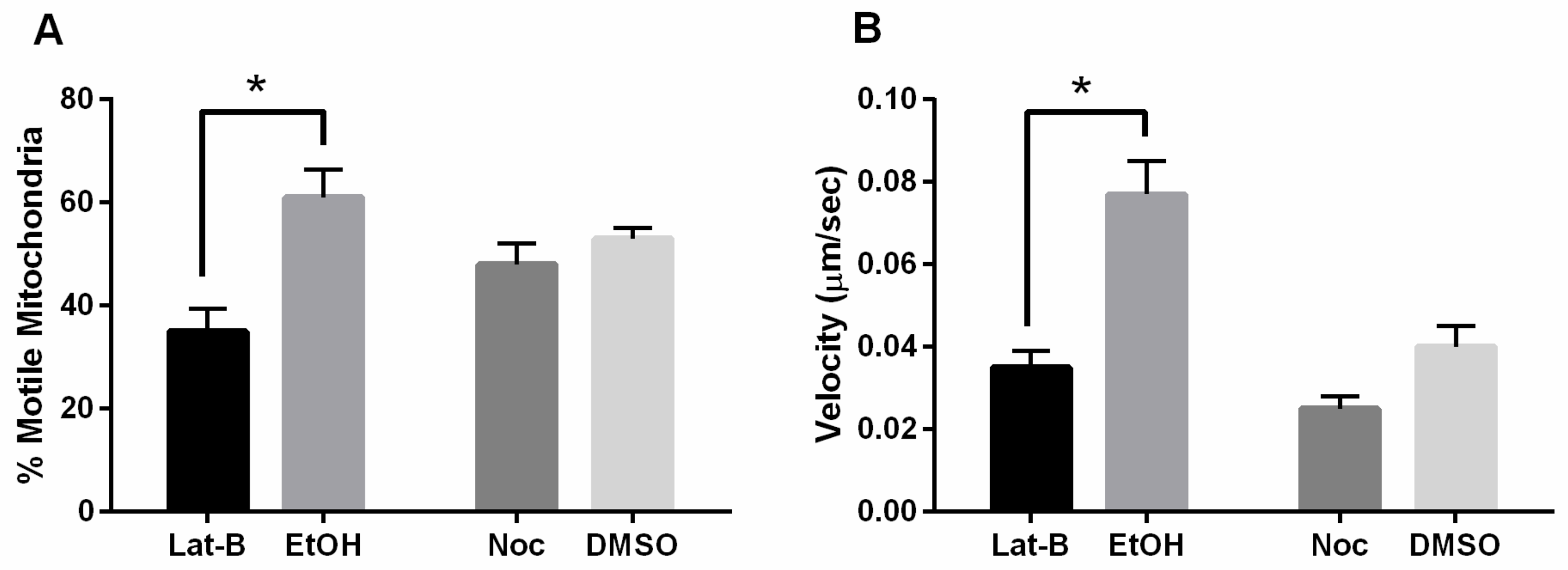
3.3. Mitochondrial Motility Is Decreased with FszB-GFP and Disrupted Actin Cytoskeleton
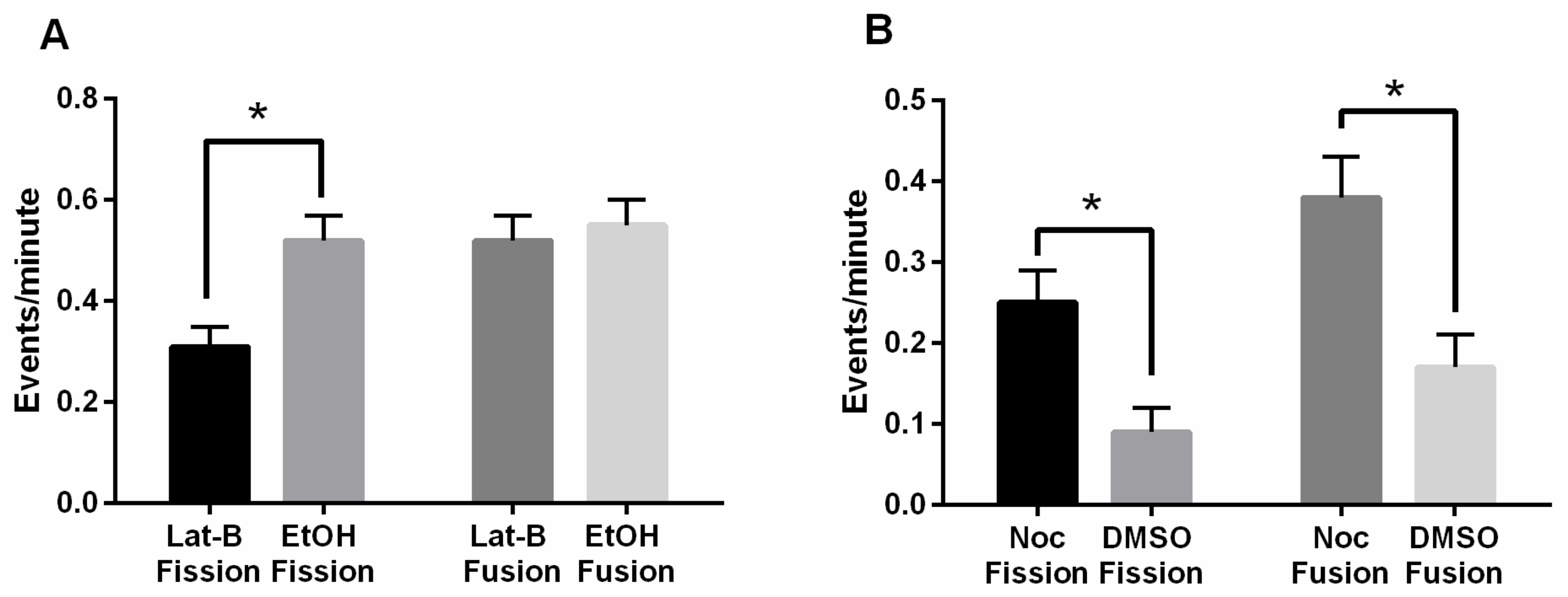
3.4. Disruption of the Actin Cytoskeleton When FszB-GFP Is Overexpressed Unbalances Fission and Fusion
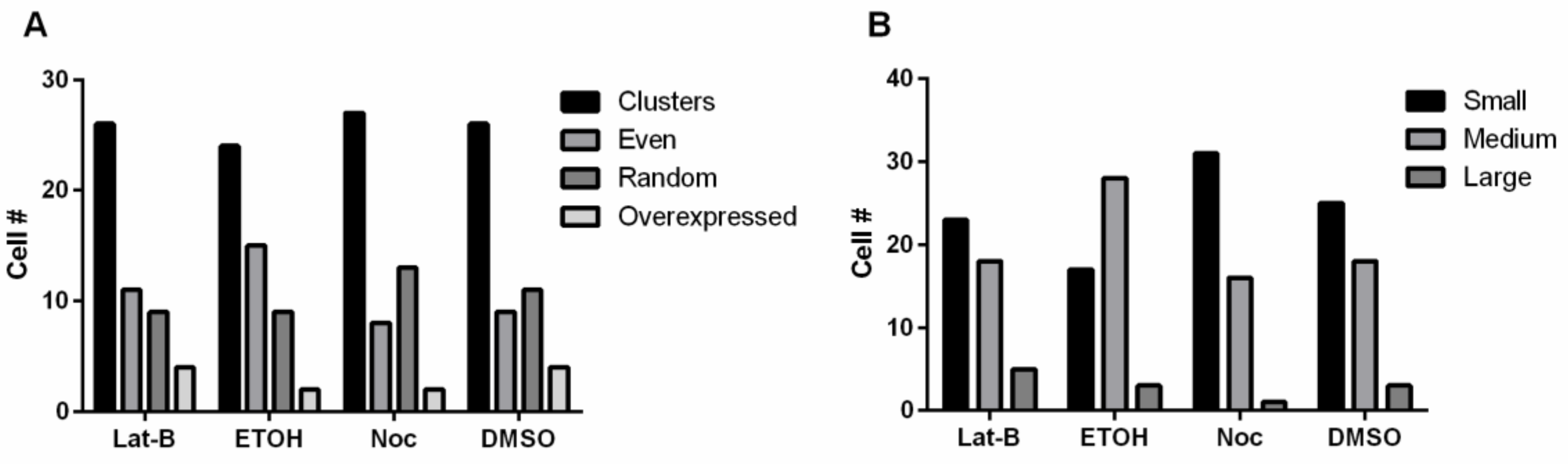
3.5. Disruption of the Cytoskeleton and Overexpressing FszB-GFP Does Not Alter Morphology or Size
4. Discussion
4.1. FszB-GFP Inhibits Fission, Fusion, and Motility
4.2. Actin Counteracts FszB-GFP to Mediate Fission Events
4.3. Microtubules and FszB-GFP Mediate Opposing Function in the Process of Mitochondrial Motility
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vyas, S.; Zaganjor, E.; Haigis, M.C. Mitochondria and Cancer. Cell 2016, 166, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Nunnari, J.; Marshall, W.; Straight, A.; Murray, A.; Sedat, J.; Walter, P. Mitochondrial transmission during mating in S. cerevisiae is determined by mitochondrial fusion and fission and the intramitochondrial segregation of mtDNA. Mol. Biol. Cell 1997, 8, 1233–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Wu, M.; Kalyanasundaram, A. Structural and biomechanical basis of mitochondrial movement in eukaryotic cells. Int. J. Nanomed. 2013, 8, 4033–4042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twig, G.; Elorza, A.; A Molina, A.J.; Mohamed, H.; Wikstrom, J.D.; Walzer, G.; Stiles, L.; E Haigh, S.; Katz, S.; Las, G.; et al. Fission and selective fusion govern mitochondrial segregation and elimination by autophagy. EMBO J. 2008, 27, 433–446. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Xie, Z.; Turkson, S.; Zhuang, X. A53T human α-synuclein overexpression in transgenic mice induces pervasive mitochondria macroautophagy defects preceding dopamine neuron degeneration. J. Neurosci. 2015, 35, 890–905. [Google Scholar] [CrossRef]
- Youle, R.J.; van der Bliek, A.M. Mitochondrial Fission, Fusion, and Stress. Science 2012, 337, 1062–1065. [Google Scholar] [CrossRef] [Green Version]
- Ashrafi, G.; Schwarz, T.L. The pathways of mitophagy for quality control and clearance of mitochondria. Cell Death Differ. 2013, 20, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Chan, D.C. Mitochondrial dynamics--fusion, fission, movement, and mitophagy--in neurodegenerative diseases. Hum. Mol. Genet. 2009, 18, R169–R176. [Google Scholar] [CrossRef]
- Hoppins, S. The regulation of mitochondrial dynamics. Curr. Opin. Cell Boil. 2014, 29, 46–52. [Google Scholar] [CrossRef]
- Horbay, R.; Bilyy, R. Mitochondrial dynamics during cell cycling. Apoptosis 2016, 21, 1327–1335. [Google Scholar] [CrossRef]
- Westrate, L.M.; Drocco, J.A.; Martin, K.R.; Hlavacek, W.S.; MacKeigan, J.P. Mitochondrial Morphological Features Are Associated with Fission and Fusion Events. PLoS ONE 2014, 9, e95265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cagalinec, M.; Safiulina, D.; Liiv, M.; Liiv, J.; Choubey, V.; Wareski, P.; Veksler, V.; Kaasik, A. Principles of the mitochondrial fusion and fission cycle in neurons. J. Cell Sci. 2013, 126, 2187–2197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sajic, M.; Mastrolia, V.; Lee, C.Y.; Trigo, D.; Sadeghian, M.; Mosley, A.J.; Gregson, N.A.; Duchen, M.R.; Smith, K.J. Impulse Conduction Increases Mitochondrial Transport in Adult Mammalian Peripheral Nerves In Vivo. PLoS Biol. 2014, 11, 1001754. [Google Scholar] [CrossRef] [PubMed]
- Woods, L.C.; Berbusse, G.W.; Naylor, K. Microtubules Are Essential for Mitochondrial Dynamics–Fission, Fusion, and Motility–in Dictyostelium discoideum. Front. Cell Dev. Boil. 2016, 4, 73. [Google Scholar] [CrossRef] [Green Version]
- Berbusse, G. Investigating the Importance of Mitochondrial Dynamics: Fission, Fusion, Trafficking and Fragmentation in the Single-Celled Organism Dictyostelium Discoideum and CD-1 Mice In Vitro; In Research; University of Central Arkansas: Conway, SC, USA, 2014. [Google Scholar]
- Morris, R.L.; Hollenbeck, P.J. Axonal transport of mitochondria along microtubules and F-actin in living vertebrate neurons. J. Cell Boil. 1995, 131, 1315–1326. [Google Scholar] [CrossRef] [Green Version]
- Detmer, S.A.; Chan, D.C. Functions and dysfunctions of mitochondrial dynamics. Nat. Rev. Mol. Cell Boil. 2007, 8, 870–879. [Google Scholar] [CrossRef]
- Karbowski, M.; Youle, R.J. Dynamics of mitochondrial morphology in healthy cells and during apoptosis. Cell Death Differ. 2003, 10, 870–880. [Google Scholar] [CrossRef] [Green Version]
- Nunnari, J.; Suomalainen, A. Mitochondria: In sickness and in health. Cell 2012, 148, 1145–1159. [Google Scholar] [CrossRef] [Green Version]
- Liesa, M.; Palacin, M.; Zorzano, A. Mitochondrial Dynamics in Mammalian Health and Disease. Physiol. Rev. 2009, 89, 799–845. [Google Scholar] [CrossRef] [Green Version]
- Tieu, K.; Imm, J. Mitochondrial Dynamics as a Potential Therapeutic Target for Parkinson’s Disease? Adv. Clin. Neurosci. Rehabil. 2014, 14, 6–8. [Google Scholar]
- Oliver, D.; Reddy, P.H. Dynamics of Dynamin-Related Protein 1 in Alzheimer’s Disease and Other Neurodegenerative Diseases. Cells 2019, 8, 961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, P.H. Abnormal tau, mitochondrial dysfunction, impaired axonal transport of mitochondria, and synaptic deprivation in Alzheimer’s disease. Brain Res. 2011, 1415, 136–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loomis, W.F. A better way to discover gene function in the social amoeba Dictyostelium discoideum. Genome Res. 2016, 26, 1161–1164. [Google Scholar] [CrossRef] [Green Version]
- Müller-Taubenberger, A.; Kortholt, A.; Eichinger, L. Simple system – substantial share: The use of Dictyostelium in cell biology and molecular medicine. Eur. J. Cell Boil. 2013, 92, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Chernivec, E.; Cooper, J.; Naylor, K. Exploring the Effect of Rotenone—A Known Inducer of Parkinson’s Disease—On Mitochondrial Dynamics in Dictyostelium discoideum. Cells 2018, 7, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annesley, S.J.; Fisher, P.R. Dictyostelium discoideum—A model for many reasons. Mol. Cell. Biochem. 2009, 329, 73–91. [Google Scholar] [CrossRef]
- Schimmel, B.G.; Berbusse, G.W.; Naylor, K. Mitochondrial fission and fusion in Dictyostelium discoideum: A search for proteins involved in membrane dynamics. BMC Res. Notes 2012, 5, 505. [Google Scholar] [CrossRef] [Green Version]
- Adams, D.W.; Errington, J. Bacterial cell division: Assembly, maintenance and disassembly of the Z ring. Nat. Rev. Genet. 2009, 7, 642–653. [Google Scholar] [CrossRef]
- And, J.L.; Addinall, S.G. Bacterial Cell Division and The Z Ring. Annu. Rev. Biochem. 1997, 66, 93–116. [Google Scholar]
- Beech, P.L.; Gilson, P.R. FtsZ and Organelle Division in Protists. Protist 2000, 151, 11–16. [Google Scholar] [CrossRef]
- Gilson, P.R.; Yu, X.-C.; Hereld, D.; Barth, C.; Savage, A.; Kiefel, B.R.; Lay, S.; Fisher, P.R.; Margolin, W.; Beech, P.L. Two Dictyostelium Orthologs of the Prokaryotic Cell Division Protein FtsZ Localize to Mitochondria and Are Required for the Maintenance of Normal Mitochondrial Morphology. Eukaryot. Cell 2003, 2, 1315–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilson, P.R.; Beech, P.L. Cell division protein FtsZ: Running rings around bacteria, chloroplasts and mitochondria. Res. Microbiol. 2001, 152, 3–10. [Google Scholar] [CrossRef]
- Takahara, M.; Takahashi, H.; Matsunaga, S.; Miyagishima, S.; Takano, H.; Sakai, A.; Kawano, S.; Kuroiwa, T. A putative mitochondrial ftsZ gene is present in the unicellular primitive red alga Cyanidioschyzon merolae. Mol. Genet. Genom. 2000, 264, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Osteryoung, K.W. The Division of Endosymbiotic Organelles. Science 2003, 302, 1698–1704. [Google Scholar] [CrossRef] [Green Version]
- Fey, P.; Dodson, R.J.; Basu, S.; Chisholm, R.L. One stop shop for everything Dictyostelium: DictyBase and the Dicty Stock Center in 2012. In Dictyostelium Discoideum Protocols; Humana Press: Totowa, NJ, USA, 2013; pp. 59–92. [Google Scholar]
- Johnson, L.V.; Walsh, M.L.; Chen, L.B. Localization of mitochondria in living cells with rhodamine 123. Proc. Natl. Acad. Sci. USA 1980, 77, 990–994. [Google Scholar] [CrossRef] [Green Version]
- Fukushima, N.H.; Brisch, E.; Keegan, B.R.; Bleazard, W.; Shaw, J.M. The GTPase Effector Domain Sequence of the Dnm1p GTPase Regulates Self-Assembly and Controls a Rate-limiting Step in Mitochondrial Fission. Mol. Biol. Cell 2001, 12, 2756–2766. [Google Scholar] [CrossRef] [Green Version]
- Labrousse, A.M.; Zappaterra, M.D.; Rube, D.A.; Van Der Bliek, A.M. C. elegans dynamin-related protein DRP-1 controls severing of the mitochondrial outer membrane. Mol. Cell 1999, 4, 815–826. [Google Scholar] [CrossRef]
- Moore, A.S.; Wong, Y.C.; Simpson, C.L.; Holzbaur, E.L.F. Dynamic actin cycling through mitochondrial subpopulations locally regulates the fission–fusion balance within mitochondrial networks. Nat. Commun. 2016, 7, 12886. [Google Scholar] [CrossRef]
- Friedman, J.R.; Lackner, L.L.; West, M.; DiBenedetto, J.R.; Nunnari, J.; Voeltz, G.K. ER tubules mark sites of mitochondrial division. Science 2011, 334, 358–362. [Google Scholar] [CrossRef] [Green Version]
- Korobova, F.; Ramabhadran, V.; Higgs, H.N. An actin-dependent step in mitochondrial fission mediated by the ER-associated formin INF2. Science 2013, 339, 464–467. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Event | Lat-B | EtOH | Noc | DMSO |
|---|---|---|---|---|
| Fission | 89% | 100% | 97% | 100% |
| Fusion | 83% | 100% | 89% | 82% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vogel, E.; Pittman, P.B.; Naylor, K. The FtsZ Homolog, FszB, Inhibits Mitochondrial Dynamics in Dictyostelium discoideum. Cells 2020, 9, 64. https://doi.org/10.3390/cells9010064
Vogel E, Pittman PB, Naylor K. The FtsZ Homolog, FszB, Inhibits Mitochondrial Dynamics in Dictyostelium discoideum. Cells. 2020; 9(1):64. https://doi.org/10.3390/cells9010064
Chicago/Turabian StyleVogel, Ericka, Pristine Bay Pittman, and Kari Naylor. 2020. "The FtsZ Homolog, FszB, Inhibits Mitochondrial Dynamics in Dictyostelium discoideum" Cells 9, no. 1: 64. https://doi.org/10.3390/cells9010064
APA StyleVogel, E., Pittman, P. B., & Naylor, K. (2020). The FtsZ Homolog, FszB, Inhibits Mitochondrial Dynamics in Dictyostelium discoideum. Cells, 9(1), 64. https://doi.org/10.3390/cells9010064