Identification and Characterization of an OSH1 Thiol Reductase from Populus trichocarpa


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Cultivation, Gene Isolation, and Vector Construction
2.2. Prokaryotic Expression, Purification, and Activity of PtOSH1 In Vitro
2.3. Overexpression of PtOSH1 in Arabidopsis Thaliana (A. Thaliana)
2.4. Polymerse Chain Reaction (PCR) and Quantitative Reverse Transcription-PCR
3. Results
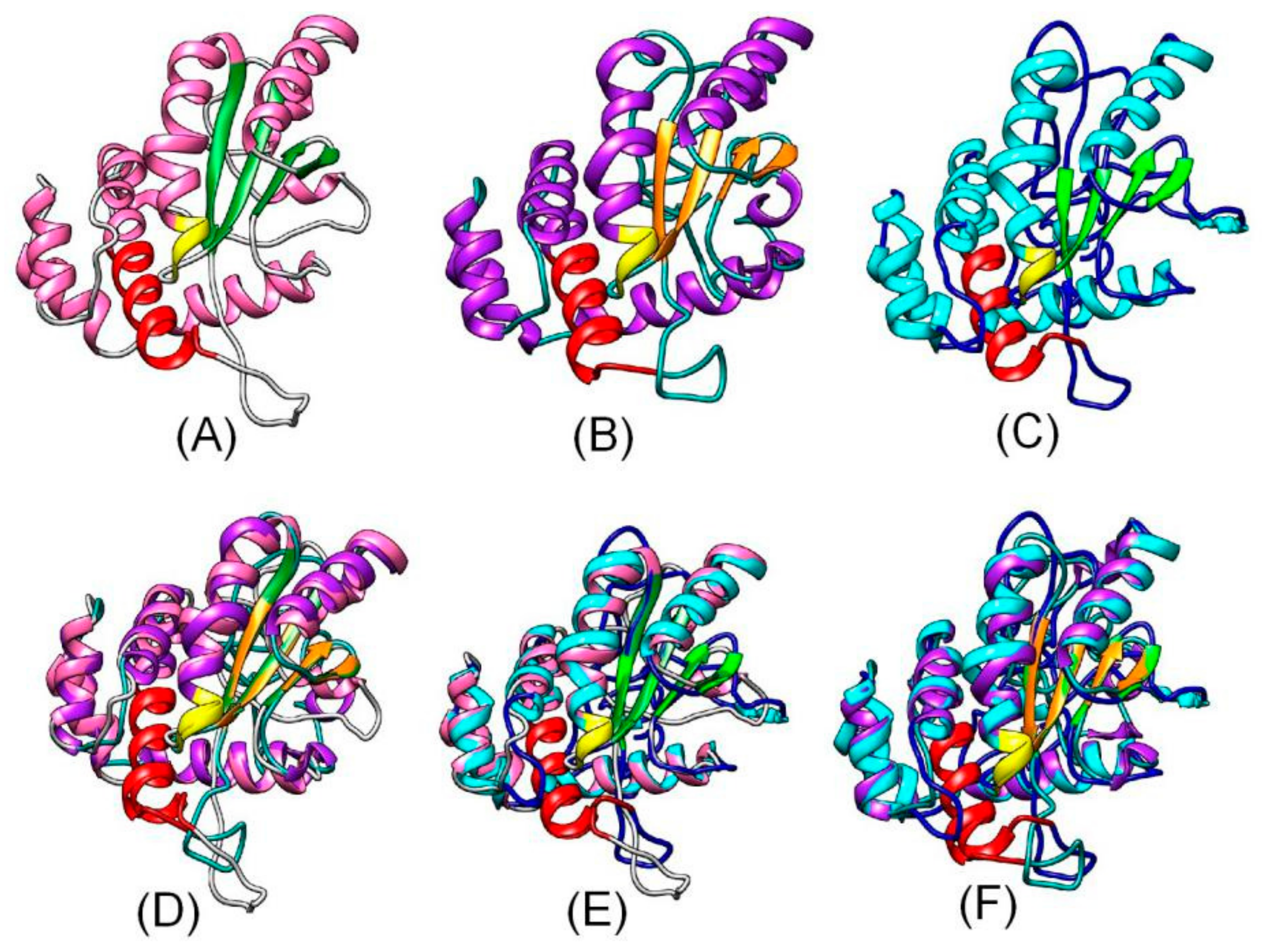
3.1. Molecular Characterization of a Poplar OSH1
3.2. Transcript Levels of PtOSH1 in Tissues and Under Cd2+ Stress
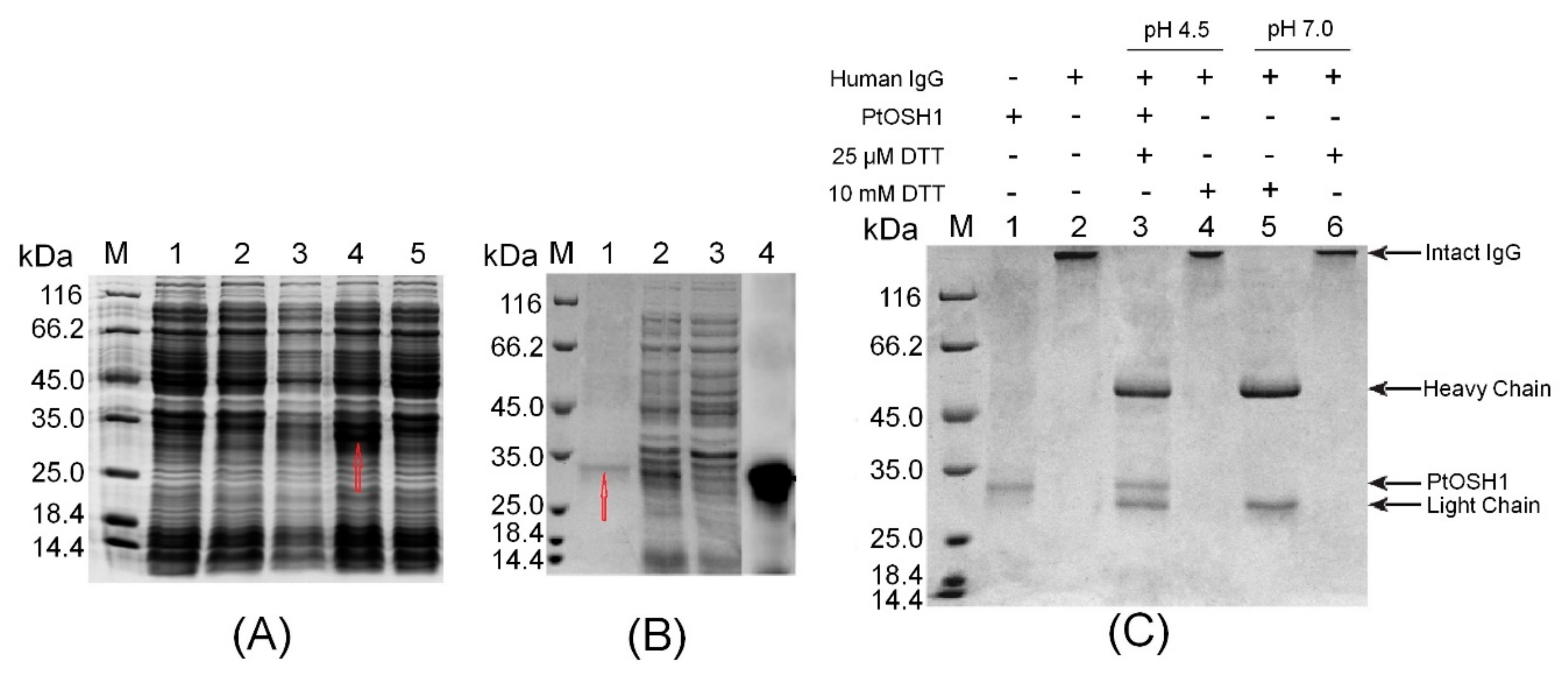
3.3. Expression, Purification, and Functional Analysis of Recombinant PtOSH1
3.4. Characterization of Transgenic A. Thaliana Lines
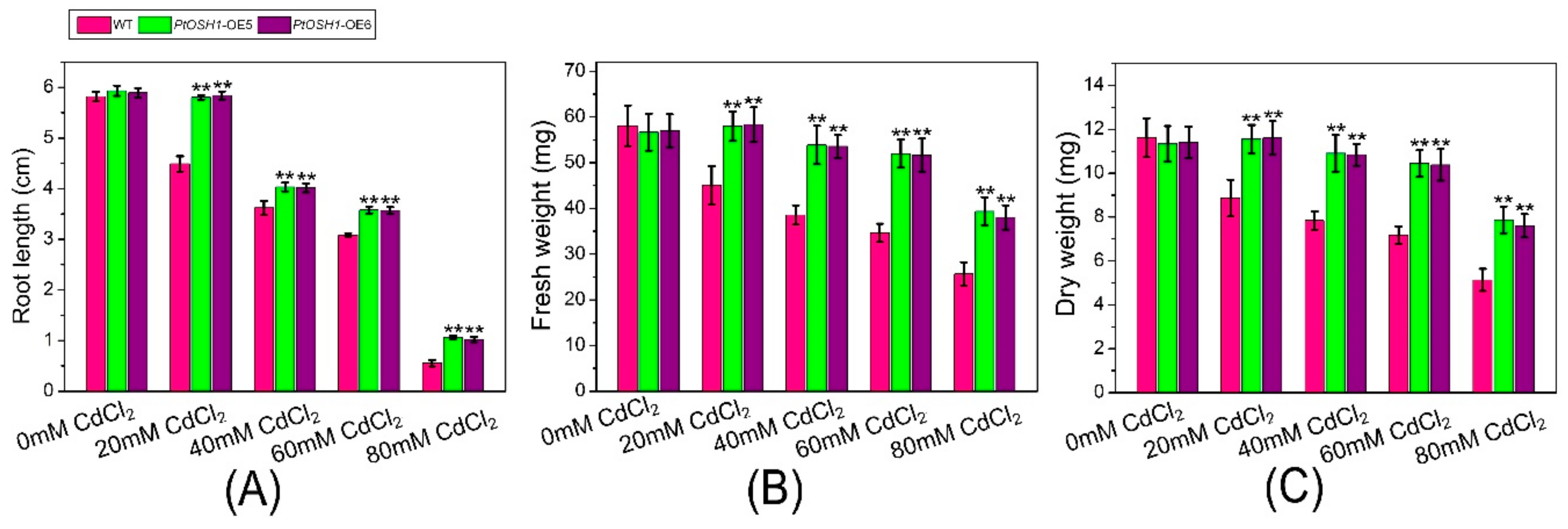
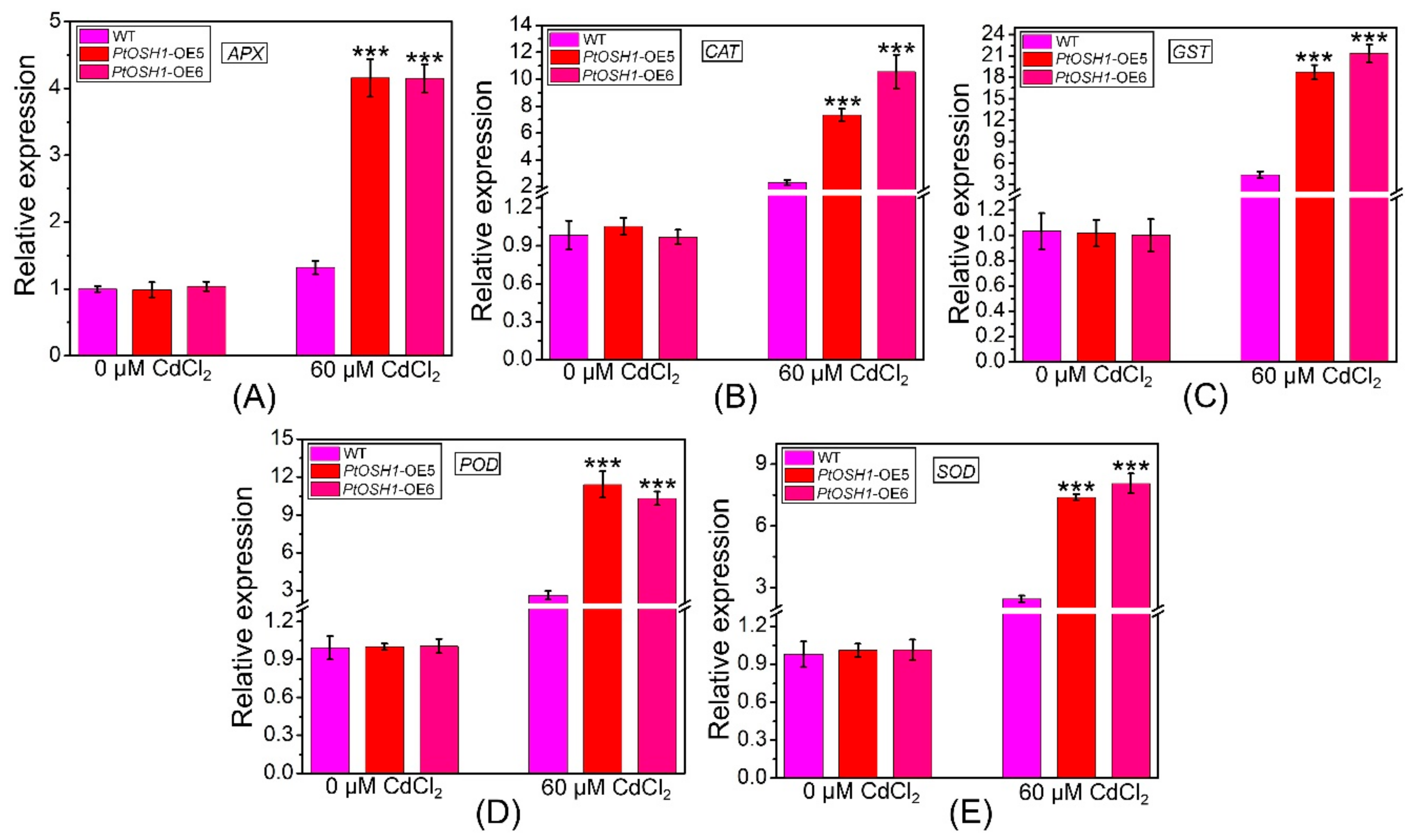
3.5. Response of Transgenic Plants to Cd2+ Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Luster, A.D.; Weinshank, R.L.; Feinman, R.; Ravetch, J.V. Molecular and biochemical characterization of a novel gamma-interferon-inducible protein. J. Biol. Chem. 1988, 263, 12036–12043. [Google Scholar] [PubMed]
- Sigal, L.J.; Crotty, S.; Andino, R.; Rock, K.L. Cytotoxic T-cell immunity to virus-infected non-haematopoietic cells requires presentation of exogenous antigen. Nature 1999, 398, 77. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, B.; Phan, U.T.; Geuze, H.J.; Cresswell, P. Enzymatic reduction of disulfide bonds in lysosomes: Characterization of a gamma-interferon-inducible lysosomal thiol reductase (GILT). Proc. Natl. Acad. Sci. USA 2000, 97, 745–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lackman, R.L.; Jamieson, A.M.; Griffith, J.M.; Geuze, H.; Cresswell, P. Innate immune recognition triggers secretion of lysosomal enzymes by macrophages. Traffic 2007, 8, 1179–1189. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Cresswell, P. Defective cross-presentation of viral antigens in GILT-free mice. Science 2010, 328, 1394–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rausch, M.P.; Hastings, K.T. Diverse cellular and organismal functions of the lysosomal thiol reductase GILT. Mol. Immunol. 2015, 68, 124–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, G.Q.; Touloukian, C.E.; Yang, J.C.; Restifo, N.P.; Sherry, R.M.; Hwu, P.; Morton, K.E. Immunization of patients with metastatic melanoma using both class I-and class II-restricted peptides from melanoma-associated antigens. J. Immunother. 2003, 26, 349. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Zhou, D.; Kinraide, T.B.; Luo, X.; Li, L.; Li, D.; Zhang, H. Cell membrane surface potential (ψ0) plays a dominant role in the phytotoxicity of copper and arsenate. Plant Physiol. 2008, 148, 2134–2143. [Google Scholar] [CrossRef] [Green Version]
- Chiang, H.S.; Maric, M. Lysosomal thiol reductase negatively regulates autophagy by altering glutathione synthesis and oxidation. Free. Radic. Biol. Med. 2011, 51, 688–699. [Google Scholar] [CrossRef]
- Zhang, C.; Wu, L.; Luo, Y.; Zhang, H.; Christie, P. Identifying sources of soil inorganic pollutants on a regional scale using a multivariate statistical approach: Role of pollutant migration and soil physicochemical properties. Environ. Pollut. 2008, 151, 470–476. [Google Scholar] [CrossRef] [Green Version]
- Conesa, J.A.; Gálvez, A.; Mateos, F.; Martín-Gullón, I.; Font, R. Organic and inorganic pollutants from cement kiln stack feeding alternative fuels. J. Hazard. Mater. 2008, 158, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.P.; Batish, D.R.; Kaur, G.; Arora, K.; Kohli, R.K. Nitric oxide (as sodium nitroprusside) supplementation ameliorates Cd toxicity in hydroponically grown wheat roots. Environ. Exp. Bot. 2008, 63, 158–167. [Google Scholar] [CrossRef]
- Fashola, M.; Ngole-Jeme, V.; Babalola, O. Heavy metal pollution from gold mines: Environmental effects and bacterial strategies for resistance. Int. J. Environ. Res. Public Health 2016, 13, 1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Chen, W. Role of salicylic acid in alleviating photochemical damage and autophagic cell death induction of cadmium stress in Arabidopsis thaliana. Photochem. Photobiol. Sci. 2011, 10, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Loake, G.J.; Chu, C. Cross-talk of nitric oxide and reactive oxygen species in plant programed cell death. Front. Plant Sci. 2013, 4, 314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, R.; Bashir, K.; Ishimaru, Y.; Nishizawa, N.K.; Nakanishi, H. The role of heavy-metal ATPases, HMAs, in zinc and cadmium transport in rice. Plant Signal. Behav. 2012, 7, 1605–1607. [Google Scholar] [CrossRef]
- Gielen, H.; Remans, T.; Vangronsveld, J.; Cuypers, A. Toxicity responses of Cu and Cd: The involvement of miRNAs and the transcription factor SPL7. BMC Plant Biol. 2016, 16, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Wei, X.; Yu, P.; Deng, X.; Xu, W.; Ma, M.; Zhang, H. Expression of cadR enhances its specific activity for Cd detoxification and accumulation in Arabidopsis. Plant Cell Physiol. 2016, 57, 1720–1731. [Google Scholar] [CrossRef] [Green Version]
- Rahoui, S.; Ben, C.; Chaoui, A.; Martinez, Y.; Yamchi, A.; Rickauer, M.; El Ferjani, E. Oxidative injury and antioxidant genes regulation in cadmium-exposed radicles of six contrasted Medicago truncatula genotypes. Environ. Sci. Pollut. Res. 2014, 21, 8070–8083. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.; Tran, L.S.P. Impacts of priming with silicon on the growth and tolerance of maize plants to alkaline stress. Front. Plant Sci. 2016, 7, 243. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Khan, A.L.; Waqas, M.; Lee, I.J. Silicon regulates antioxidant activities of crop plants under abiotic-induced oxidative stress: A review. Front. Plant Sci. 2017, 8, 510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lillig, C.H.; Holmgren, A. Thioredoxin and related molecules–from biology to health and disease. Antioxid. Redox Signal. 2007, 9, 25–47. [Google Scholar] [CrossRef] [PubMed]
- Meyer, Y.; Vignols, F.; Reichheld, J.P. Classification of plant thioredoxins by sequence similarity and intron position. Methods Enzymol. 2002, 347, 394. [Google Scholar] [PubMed]
- Collin, V.; Issakidis-Bourguet, E.; Marchand, C.; Hirasawa, M.; Lancelin, J.M.; Knaff, D.B.; Miginiac-Maslow, M. The Arabidopsis plastidial thioredoxins New functions and new insights into specificity. J. Biol. Chem. 2003, 278, 23747–23752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laloi, C.; Rayapuram, N.; Chartier, Y.; Grienenberger, J.M.; Bonnard, G.; Meyer, Y. Identification and characterization of a mitochondrial thioredoxin system in plants. Proc. Natl. Acad. Sci. USA 2001, 98, 14144–14149. [Google Scholar] [CrossRef] [Green Version]
- Serrato, A.J.; Cejudo, F.J. Type-h thioredoxins accumulate in the nucleus of developing wheat seed tissues suffering oxidative stress. Planta 2003, 217, 392–399. [Google Scholar] [CrossRef]
- Balmer, Y.; Vensel, W.H.; Tanaka, C.K.; Hurkman, W.J.; Gelhaye, E.; Rouhier, N.; Buchanan, B.B. Thioredoxin links redox to the regulation of fundamental processes of plant mitochondria. Proc. Natl. Acad. Sci. USA 2004, 101, 2642–2647. [Google Scholar] [CrossRef] [Green Version]
- Holmgren, A. Thioredoxin structure and mechanism: Conformational changes on oxidation of the active-site sulfhydryls to a disulfide. Structure 1995, 3, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Miki, H.; Funato, Y. Regulation of intracellular signalling through cysteine oxidation by reactive oxygen species. J. Biochem. 2012, 151, 255–261. [Google Scholar] [CrossRef] [Green Version]
- Cooper, C.E.; Patel, R.P.; Brookes, P.S.; Darley-Usmar, V.M. Nanotransducers in cellular redox signaling: Modification of thiols by reactive oxygen and nitrogen species. Trends Biochem. Sci. 2002, 27, 489–492. [Google Scholar] [CrossRef]
- Nakamura, H.; Nakamura, K.; Yodoi, J. Redox regulation of cellular activation. Annu. Rev. Immunol. 1997, 15, 351–369. [Google Scholar] [CrossRef] [PubMed]
- Sahaf, B.; Söderberg, A.; Spyrou, G.; Barral, A.M.; Pekkari, K.; Holmgren, A.; Rosén, A. Thioredoxin expression and localization in human cell lines: Detection of full-length and truncated species. Exp. Cell Res. 1997, 236, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Schenk, H.; Klein, M.; Erdbrügger, W.; Dröge, W.; Schulze-Osthoff, K. Distinct effects of thioredoxin and antioxidants on the activation of transcription factors NF-kappa B and AP-1. Proc. Natl. Acad. Sci. USA 1994, 91, 1672–1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirota, K.; Murata, M.; Sachi, Y.; Nakamura, H.; Takeuchi, J.; Mori, K.; Yodoi, J. Distinct roles of thioredoxin in the cytoplasm and in the nucleus a two-step mechanism of redox regulation of transcription factor NF-κB. J. Biol. Chem. 1999, 274, 27891–27897. [Google Scholar] [CrossRef] [Green Version]
- Rollin-Genetet, F.; Berthomieu, C.; Davin, A.H.; Quéméneur, E. Escherichia coli thioredoxin inhibition by cadmium: Two mutually exclusive binding sites involving Cys32 and Asp26. Eur. J. Biochem. 2004, 271, 1299–1309. [Google Scholar] [CrossRef]
- Lemaire, S.; Keryer, E.; Stein, M.; Schepens, I.; Issakidis-Bourguet, E.; Gérard-Hirne, C.; Jacquot, J.P. Heavy-Metal Regulation of Thioredoxin Gene Expression in Chlamydomonas reinhardtii. Plant Physiol. 1999, 120, 773–778. [Google Scholar] [CrossRef] [Green Version]
- Ohkama-Ohtsu, N.; Kasajima, I.; Fujiwara, T.; Naito, S. Isolation and characterization of an Arabidopsis mutant that overaccumulates O-acetyl-L-Ser. Plant Physiol. 2004, 136, 3209–3222. [Google Scholar] [CrossRef] [Green Version]
- Tuskan, G.A.; Difazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Schein, J. The genome of black cottonwood, Populus trichocarpa (Torr. Gray). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Zang, D.; Wang, C.; Ji, X.; Wang, Y. Tamarix hispida zinc finger protein ThZFP1 participates in salt and osmotic stress tolerance by increasing proline content and SOD and POD activities. Plant Sci. 2015, 235, 111–121. [Google Scholar] [CrossRef]
- Maric, M.; Arunachalam, B.; Phan, U.T.; Dong, C.; Garrett, W.S.; Cannon, K.S.; Cresswell, P. Defective antigen processing in GILT-free mice. Science 2001, 294, 1361–1365. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Cao, X.; Ji, X.; Liu, H.; Wu, H.; Gu, W.; Zhang, S. Molecular structure, tissue distribution and functional characterization of interferon-γ-inducible lysosomal thiol reductase (GILT) gene in chicken (Gallus gallus). Vet. Immunol. Immunopathol. 2013, 153, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Woods, I.G.; Wilson, C.; Friedlander, B.; Chang, P.; Reyes, D.K.; Nix, R.; Talbot, W.S. The zebrafish gene map defines ancestral vertebrate chromosomes. Genome Res. 2005, 15, 1307–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Zhang, S.; Liu, Z.; Gaowa, S.; Wang, Y. Characterization and expression of gamma-interferon-inducible lysosomal thiol reductase (GILT) gene in amphioxus Branchiostoma belcheri with implications for GILT in innate immune response. Mol. Immunol. 2007, 44, 2631–2637. [Google Scholar] [CrossRef]
- Dan, W.B.; Wang, S.L.; Liang, J.Q.; Zhang, S.Q. Molecular cloning and expression analysis of porcine γ-interferon-inducible lysosomal thiol reductase (GILT). Vet. Immunol. Immunopathol. 2008, 126, 163–167. [Google Scholar] [CrossRef]
- Zheng, W.; Chen, X. Cloning and expression analysis of interferon-γ-inducible-lysosomal thiol reductase gene in large yellow croaker (Pseudosciaena crocea). Mol. Immunol. 2006, 43, 2135–2141. [Google Scholar] [CrossRef]
- Kongton, K.; McCall, K.; Phongdara, A. Identification of gamma-interferon-inducible lysosomal thiol reductase (GILT) homologues in the fruit fly Drosophila melanogaster. Dev. Comp. Immunol. 2014, 44, 389–396. [Google Scholar] [CrossRef]
- Phan, U.T.; Maric, M.; Dick, T.P.; Cresswell, P. Multiple species express thiol oxidoreductases related to GILT. Immunogenetics 2001, 53, 342–346. [Google Scholar] [CrossRef]
- Lackman, R.L.; Cresswell, P. Exposure of the promonocytic cell line THP-1 to Escherichia coli induces IFN-γ-inducible lysosomal thiol reductase expression by inflammatory cytokines. J. Immunol. 2006, 177, 4833–4840. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Haque, M.A.; Blum, J.S. Role of disulfide bonds in regulating antigen processing and epitope selection. J. Immunol. 2002, 169, 2444–2450. [Google Scholar] [CrossRef]
- Hastings, K.T.; Cresswell, P. Disulfide reduction in the endocytic pathway: Immunological functions of gamma-interferon-inducible lysosomal thiol reductase. Antioxid. Redox Signal. 2011, 15, 657–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McInnis, S.M.; Desikan, R.; Hancock, J.T.; Hiscock, S.J. Production of reactive oxygen species and reactive nitrogen species by angiosperm stigmas and pollen: Potential signalling crosstalk? New Phytol. 2006, 172, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.F. Enhancing antitumor immune responses: Intracellular peptide delivery and identification of MHC class II-restricted tumor antigens. Immunol. Rev. 2002, 188, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Navrot, N.; Rouhier, N.; Gelhaye, E.; Jacquot, J.P. Reactive oxygen species generation and antioxidant systems in plant mitochondria. Physiol. Plant. 2007, 129, 185–195. [Google Scholar] [CrossRef]
- Pitzschke, A.; Forzani, C.; Hirt, H. Reactive oxygen species signaling in plants. Antioxid. Redox Signal. 2006, 8, 1757–1764. [Google Scholar] [CrossRef]
- Jacquot, J.P.; Lancelin, J.M.; Meyer, Y. Thioredoxins: Structure and function in plant cells. New Phytol. 1997, 136, 543–570. [Google Scholar] [CrossRef]
- Schürmann, P.; Jacquot, J.P. Plant thioredoxin systems revisited. Annu. Rev. Plant. Biol. 2000, 51, 371–400. [Google Scholar] [CrossRef] [Green Version]
- Hossain, Z.; Komatsu, S. Contribution of proteomic studies towards understanding plant heavy metal stress response. Front. Plant. Sci. 2013, 3, 310. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, C.V.; Rey, P. Plant thioredoxins are key actors in the oxidative stress response. Trends Plant. Sci. 2006, 11, 329–334. [Google Scholar] [CrossRef]
- Zhou, R.; Tardivel, A.; Thorens, B.; Choi, I.; Tschopp, J. Thioredoxin-interacting protein links oxidative stress to inflammasome activation. Nat. Immunol. 2010, 11, 136. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant. Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salt, D.E.; Wagner, G.J. Cadmium transport across tonoplast of vesicles from oat roots. Evidence for a Cd2+/H+ antiport activity. J. Biol. Chem. 1993, 268, 12297–12302. [Google Scholar] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, H.; Zhou, J.; Xu, C.; Movahedi, A.; Sun, W.; Li, D.; Zhuge, Q. Identification and Characterization of an OSH1 Thiol Reductase from Populus trichocarpa. Cells 2020, 9, 76. https://doi.org/10.3390/cells9010076
Wei H, Zhou J, Xu C, Movahedi A, Sun W, Li D, Zhuge Q. Identification and Characterization of an OSH1 Thiol Reductase from Populus trichocarpa. Cells. 2020; 9(1):76. https://doi.org/10.3390/cells9010076
Chicago/Turabian StyleWei, Hui, Jie Zhou, Chen Xu, Ali Movahedi, Weibo Sun, Dawei Li, and Qiang Zhuge. 2020. "Identification and Characterization of an OSH1 Thiol Reductase from Populus trichocarpa" Cells 9, no. 1: 76. https://doi.org/10.3390/cells9010076
APA StyleWei, H., Zhou, J., Xu, C., Movahedi, A., Sun, W., Li, D., & Zhuge, Q. (2020). Identification and Characterization of an OSH1 Thiol Reductase from Populus trichocarpa. Cells, 9(1), 76. https://doi.org/10.3390/cells9010076