Precision Medicine in Neurodegenerative Diseases: Some Promising Tips Coming from the microRNAs’ World


Abstract
:1. Introduction
2. MiRNAs as Pharmacoepigenomic Targets for NDDs
3. Alzheimer’s Disease
4. Parkinson’s Disease
5. Multiple Sclerosis
6. MiRNAs Involved in MS Drug Resistance
7. Potential Role of miRNA Polymorphisms in Drug Response and Efficacy
8. The Therapeutic Potential of miRNAs
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Strafella, C.; Caputo, V.; Galota, M.R.; Zampatti, S.; Marella, G.; Mauriello, S.; Cascella, R.; Giardina, E. Application of precision medicine in neurodegenerative diseases. Front. Neurol. 2018, 9, 701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, L.; Tan, E.K.; Zeng, L. MicroRNAs and neurodegenerative diseases. In MicroRNA: Medical Evidence: From Molecular Biology to Clinical Practice; Santulli, G., Ed.; Advances in Experimental Medicine and Biology, Springer International Publishing: Cham, Switzerland, 2015; pp. 85–105. ISBN 978-3-319-22671-2. [Google Scholar]
- Lewis, P.A.; Spillane, J.E. Chapter 7—Multiple sclerosis. In The Molecular and Clinical Pathology of Neurodegenerative Disease; Lewis, P.A., Spillane, J.E., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 221–251. ISBN 978-0-12-811069-0. [Google Scholar]
- Junn, E.; Mouradian, M.M. MicroRNAs in neurodegenerative diseases and their therapeutic potential. Pharmacol. Ther. 2012, 133, 142–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elangovan, S.; Holsinger, D.M.R. Personalized medicine in neurodegenerative diseases (SJNDT). S. J. Neurol. Disord. Ther. 2018, 1–4. [Google Scholar]
- Gupta, S.; Verma, S.; Mantri, S.; Berman, N.E.; Sandhir, R. Targeting microRNAs in prevention and treatment of neurodegenerative disorders. Drug Dev. Res. 2015, 76, 397–418. [Google Scholar] [CrossRef] [PubMed]
- Basak, I.; Patil, K.S.; Alves, G.; Larsen, J.P.; Møller, S.G. MicroRNAs as neuroregulators, biomarkers and therapeutic agents in neurodegenerative diseases. Cell. Mol. Life Sci. 2016, 73, 811–827. [Google Scholar] [CrossRef]
- Espinoza-Lewis, R.A.; Wang, D.-Z. Chapter ten—MicroRNAs in heart development. In Current Topics in Developmental Biology; Bruneau, B.G., Ed.; Heart Development, Academic Press: Cambridge, MA, USA, 2012; Volume 100, pp. 279–317. [Google Scholar]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2008, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The rosetta stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Qin, H.; Zhao, Q.; He, X.-X. Emerging role of transcription factor-microRNA-target gene feed-forward loops in cancer. Biomed. Rep. 2015, 3, 611–616. [Google Scholar] [CrossRef] [Green Version]
- Freiesleben, S.; Hecker, M.; Zettl, U.K.; Fuellen, G.; Taher, L. Analysis of microRNA and gene expression profiles in multiple sclerosis: Integrating interaction data to uncover regulatory mechanisms. Sci. Rep. 2016, 6, 34512. [Google Scholar] [CrossRef]
- McKeever, P.M.; Schneider, R.; Taghdiri, F.; Weichert, A.; Multani, N.; Brown, R.A.; Boxer, A.L.; Karydas, A.; Miller, B.; Robertson, J.; et al. MicroRNA expression levels are altered in the cerebrospinal fluid of patients with young-onset Alzheimer’s disease. Mol. Neurobiol. 2018, 55, 8826–8841. [Google Scholar] [CrossRef] [Green Version]
- Leggio, L.; Vivarelli, S.; L’Episcopo, F.; Tirolo, C.; Caniglia, S.; Testa, N.; Marchetti, B.; Iraci, N. MicroRNAs in parkinson’s disease: From pathogenesis to novel diagnostic and therapeutic approaches. Int. J. Mol. Sci. 2017, 18, 2698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liguori, M.; Nuzziello, N.; Introna, A.; Consiglio, A.; Licciulli, F.; D’Errico, E.; Scarafino, A.; Distaso, E.; Simone, I.L. Dysregulation of microRNAs and target genes networks in peripheral blood of patients with sporadic amyotrophic lateral sclerosis. Front. Mol. Neurosci. 2018, 11, 288. [Google Scholar] [CrossRef]
- Juźwik, C.A.; Drake, S.S.; Zhang, Y.; Paradis-Isler, N.; Sylvester, A.; Amar-Zifkin, A.; Douglas, C.; Morquette, B.; Moore, C.S.; Fournier, A.E. MicroRNA dysregulation in neurodegenerative diseases: A systematic review. Prog. Neurobiol. 2019, 182, 101664. [Google Scholar] [CrossRef] [PubMed]
- Rojo Arias, J.E.; Busskamp, V. Challenges in microRNAs’ targetome prediction and validation. Neural Regen. Res. 2019, 14, 1672–1677. [Google Scholar] [PubMed]
- Marangon, D.; Raffaele, S.; Fumagalli, M.; Lecca, D. MicroRNAs change the games in central nervous system pharmacology. Biochem. Pharmacol. 2019, 168, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Dolan, M.E. Emerging role of microRNAs in drug response. Curr. Opin. Mol. 2010, 12, 695–702. [Google Scholar]
- Latini, A.; Borgiani, P.; Novelli, G.; Ciccacci, C. MiRNAs in drug response variability: Potential utility as biomarkers for personalized medicine. Pharmacogenomics 2019, 20, 1049–1059. [Google Scholar] [CrossRef]
- Yu, A.-M.; Tian, Y.; Tu, M.-J.; Ho, P.Y.; Jilek, J.L. MicroRNA pharmacoepigenetics: Posttranscriptional regulation mechanisms behind variable drug disposition and strategy to develop more effective therapy. Drug Metab. Dispos. 2016, 44, 308–319. [Google Scholar] [CrossRef] [Green Version]
- Peedicayil, J. Pharmacoepigenetics and pharmacoepigenomics: An overview. Curr. Drug Discov. Technol. 2019, 16, 392–399. [Google Scholar] [CrossRef]
- Peedicayil, J. Pharmacoepigenetics and pharmacoepigenomics. Pharmacogenomics 2008, 9, 1785–1786. [Google Scholar] [CrossRef] [Green Version]
- Landgrave-Gómez, J.; Mercado-Gómez, O.; Guevara-Guzmán, R. Epigenetic mechanisms in neurological and neurodegenerative diseases. Front. Cell. Neurosci. 2015, 9, 58. [Google Scholar] [PubMed] [Green Version]
- Woodcock, D.M.; Crowther, P.J.; Diver, W.P. The majority of methylated deoxycytidines in human DNA are not in the CpG dinucleotide. Biochem. Biophys. Res. Commun. 1987, 145, 888–894. [Google Scholar] [CrossRef]
- Xylaki, M.; Atzler, B.; Outeiro, T.F. Epigenetics of the synapse in neurodegeneration. Curr. Neurol. Neurosci. Rep. 2019, 19, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lister, R.; Mukamel, E.A.; Nery, J.R.; Urich, M.; Puddifoot, C.A.; Johnson, N.D.; Lucero, J.; Huang, Y.; Dwork, A.J.; Schultz, M.D.; et al. Global epigenomic reconfiguration during mammalian brain development. Science 2013, 341, 1237905. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Sarathlal, K.C.; Taliyan, R. Epigenetics in neurodegenerative diseases: The role of histone DEacetylases. CNS Neurol. Disord. Drug Targets 2019, 18, 11–18. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, S.; Yu, L.; Jin, J.; Ye, X.; Liu, Y.; Xu, Y. HDAC3 negatively regulates spatial memory in a mouse model of Alzheimer’s disease. Aging Cell 2017, 16, 1073–1082. [Google Scholar] [CrossRef]
- Harrison, I.F.; Smith, A.D.; Dexter, D.T. Pathological histone acetylation in Parkinson’s disease: Neuroprotection and inhibition of microglial activation through SIRT 2 inhibition. Neurosci. Lett. 2018, 666, 48–57. [Google Scholar] [CrossRef]
- Garbes, L.; Riessland, M.; Wirth, B. Histone acetylation as a potential therapeutic target in motor neuron degenerative diseases. Curr. Pharm. Des. 2013, 19, 5093–5104. [Google Scholar] [CrossRef]
- He, Y.; Chevillet, J.R.; Liu, G.; Kim, T.K.; Wang, K. The effects of microRNA on the absorption, distribution, metabolism and excretion of drugs. Br. J. Pharm. 2015, 172, 2733–2747. [Google Scholar] [CrossRef] [Green Version]
- Nakano, M.; Nakajima, M. Current knowledge of microRNA-mediated regulation of drug metabolism in humans. Expert Opin. Drug Metab. Toxicol. 2018, 14, 493–504. [Google Scholar] [CrossRef]
- Massoud, F.; Gauthier, S. Update on the pharmacological treatment of Alzheimer’s disease. Curr. Neuropharmacol. 2010, 8, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Cedazo-Mínguez, A. Apolipoprotein E and Alzheimer’s disease: Molecular mechanisms and therapeutic opportunities. J. Cell. Mol. Med. 2007, 11, 1227–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verheijen, J.; Sleegers, K. Understanding Alzheimer disease at the interface between genetics and transcriptomics. Trends Genet. 2018, 34, 434–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, L.W. Alzheimer’s disease: Early diagnosis and treatment. Hong Kong Med. J. 2012, 18, 228–237. [Google Scholar] [PubMed]
- Nikolac Perkovic, M.; Pivac, N. Genetic markers of Alzheimer’s disease. In Frontiers in Psychiatry: Artificial Intelligence, Precision Medicine, and Other Paradigm Shifts; Advances in Experimental Medicine and Biology; Kim, Y.-K., Ed.; Springer: Singapore, 2019; pp. 27–52. ISBN 978-981-329-721-0. [Google Scholar]
- Gauthier, S. Pharmacological treatment of Alzheimer’s disease. Alzheimer’s Dement J. Alzheimer’s Assoc. 2012, 8, P2. [Google Scholar] [CrossRef]
- Weller, J.; Budson, A. Current understanding of Alzheimer’s disease diagnosis and treatment. F1000Research 2018, 7, F1000. [Google Scholar] [CrossRef] [Green Version]
- Mangialasche, F.; Solomon, A.; Winblad, B.; Mecocci, P.; Kivipelto, M. Alzheimer’s disease: Clinical trials and drug development. Lancet Neurol. 2010, 9, 702–716. [Google Scholar] [CrossRef]
- Godyń, J.; Jończyk, J.; Panek, D.; Malawska, B. Therapeutic strategies for Alzheimer’s disease in clinical trials. Pharm. Rep. 2016, 68, 127–138. [Google Scholar] [CrossRef]
- Wang, C.-N.; Wang, Y.-J.; Wang, H.; Song, L.; Chen, Y.; Wang, J.-L.; Ye, Y.; Jiang, B. The anti-dementia effects of donepezil involve miR-206-3p in the hippocampus and cortex. Biol. Pharm. Bull. 2017, 40, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Tian, N.; Cao, Z.; Zhang, Y. MiR-206 decreases brain-derived neurotrophic factor levels in a transgenic mouse model of Alzheimer’s disease. Neurosci. Bull. 2014, 30, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Yan, Y.; Jiao, Y.; Gao, Z.; Xia, Y.; Kong, L.; Yao, Y.; Tao, Z.; Song, J.; Yan, Y.; et al. Neuroprotective effect of osthole on neuron synapses in an Alzheimer’s disease cell model via upregulation of microRNA-9. J. Mol. Neurosci. 2016, 60, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, S.; Kishi, T.; Iwata, N. Combination therapy with cholinesterase inhibitors and memantine for Alzheimer’s disease: A systematic review and meta-analysis. Int. J. Neuropsychopharmacol. 2014, 18, pyu115. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Li, Z.; Zhao, L.; Zhao, W. Simvastatin ameliorate memory deficits and inflammation in clinical and mouse model of Alzheimer’s disease via modulating the expression of miR-106b. Biomed. Pharm. 2017, 92, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Kong, L.; Yao, Y.; Li, S.; Tao, Z.; Yan, Y.; Yang, J. Osthole decreases beta amyloid levels through up-regulation of miR-107 in Alzheimer’s disease. Neuropharmacology 2016, 108, 332–344. [Google Scholar] [CrossRef]
- Zhang, Z.-R.; Leung, W.N.; Cheung, H.Y.; Chan, C.W. Osthole: A review on its bioactivities, pharmacological properties, and potential as alternative medicine. Evid.-Based Complement. Altern. Med. 2015, 2015, 919616. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Wen, Q.; Liang, W.; Kang, T.; Ren, L.; Zhang, N.; Zhao, D.; Sun, D.; Yang, J. Osthole reverses beta-amyloid peptide cytotoxicity on neural cells by enhancing cyclic AMP response element-binding protein phosphorylation. Biol. Pharm. Bull. 2013, 36, 1950–1958. [Google Scholar] [CrossRef] [Green Version]
- Chao, X.; Zhou, J.; Chen, T.; Liu, W.; Dong, W.; Qu, Y.; Jiang, X.; Ji, X.; Zhen, H.; Fei, Z. Neuroprotective effect of osthole against acute ischemic stroke on middle cerebral ischemia occlusion in rats. Brain Res. 2010, 1363, 206–211. [Google Scholar] [CrossRef]
- Lin, Y.; Liang, X.; Yao, Y.; Xiao, H.; Shi, Y.; Yang, J. Osthole attenuates APP-induced Alzheimer’s disease through up-regulating miRNA-101a-3p. Life Sci. 2019, 225, 117–131. [Google Scholar] [CrossRef]
- Yao, Y.; Gao, Z.; Liang, W.; Kong, L.; Jiao, Y.; Li, S.; Tao, Z.; Yan, Y.; Yang, J. Osthole promotes neuronal differentiation and inhibits apoptosis via Wnt/β-catenin signaling in an Alzheimer’s disease model. Toxicol. Appl. Pharm. 2015, 289, 474–481. [Google Scholar] [CrossRef]
- Li, S.-H.; Gao, P.; Wang, L.-T.; Yan, Y.-H.; Xia, Y.; Song, J.; Li, H.-Y.; Yang, J.-X. Osthole stimulated neural stem cells differentiation into neurons in an Alzheimer’s disease cell model via upregulation of microRNA-9 and rescued the functional impairment of hippocampal neurons in APP/PS1 transgenic mice. Front. Neurosci. 2017, 11, 340. [Google Scholar] [CrossRef] [Green Version]
- Shi, R.; Zhang, S.; Cheng, G.; Yang, X.; Zhao, N.; Chen, C. Ginsenoside Rg1 and acori graminei rhizoma attenuates neuron cell apoptosis by promoting the expression of miR-873-5p in Alzheimer’s disease. Neurochem. Res. 2018, 43, 1529–1538. [Google Scholar] [CrossRef] [PubMed]
- Sherer, T.B.; Chowdhury, S.; Peabody, K.; Brooks, D.W. Overcoming obstacles in Parkinson’s disease. Mov. Disord. 2012, 27, 1606–1611. [Google Scholar] [CrossRef] [PubMed]
- Beitz, J.M. Parkinson’s disease: A review. Front. Biosci. 2014, 6, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Margis, R.; Margis, R.; Rieder, C.R.M. Identification of blood microRNAs associated to Parkinsońs disease. J. Biotechnol. 2011, 152, 96–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef]
- Chaudhuri, K.R.; Odin, P. The challenge of non-motor symptoms in Parkinson’s disease. Prog. Brain Res. 2010, 184, 325–341. [Google Scholar]
- Caggiu, E.; Paulus, K.; Mameli, G.; Arru, G.; Sechi, G.P.; Sechi, L.A. Differential expression of miRNA 155 and miRNA 146a in Parkinson’s disease patients. eNeurologicalSci 2018, 13, 1–4. [Google Scholar] [CrossRef]
- Serafin, A.; Foco, L.; Zanigni, S.; Blankenburg, H.; Picard, A.; Zanon, A.; Giannini, G.; Pichler, I.; Facheris, M.F.; Cortelli, P.; et al. Overexpression of blood microRNAs 103a, 30b, and 29a in L-dopa-treated patients with PD. Neurology 2015, 84, 645–653. [Google Scholar] [CrossRef]
- Filatova, E.V.; Alieva, A.K.; Shadrina, M.I.; Slominsky, P.A. MicroRNAs: Possible role in pathogenesis of Parkinson’s disease. Biochemistry (Mosc) 2012, 77, 813–819. [Google Scholar] [CrossRef]
- Liu, X.; Chen, J.; Guan, T.; Yao, H.; Zhang, W.; Guan, Z.; Wang, Y. MiRNAs and target genes in the blood as biomarkers for the early diagnosis of Parkinson’s disease. BMC Syst. Biol. 2019, 13, 10. [Google Scholar]
- Shinde, S.; Mukhopadhyay, S.; Mohsen, G.; Khoo, S.K. Biofluid-based microRNA biomarkers for Parkinsons disease: An overview and update. AIMS Med. Sci. 2015, 2, 15. [Google Scholar] [CrossRef]
- Kalinderi, K.; Fidani, L.; Katsarou, Z.; Bostantjopoulou, S. Pharmacological treatment and the prospect of pharmacogenetics in Parkinson’s disease. Int. J. Clin. Pract. 2011, 65, 1289–1294. [Google Scholar] [CrossRef] [PubMed]
- Rizek, P.; Kumar, N.; Jog, M.S. An update on the diagnosis and treatment of Parkinson disease. CMAJ 2016, 188, 1157–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connolly, B.S.; Lang, A.E. Pharmacological treatment of Parkinson disease: A review. JAMA 2014, 311, 1670–1683. [Google Scholar] [CrossRef] [PubMed]
- Schwienbacher, C.; Foco, L.; Picard, A.; Corradi, E.; Serafin, A.; Panzer, J.; Zanigni, S.; Blankenburg, H.; Facheris, M.F.; Giannini, G.; et al. Plasma and white blood cells show different miRNA expression profiles in Parkinson’s disease. J. Mol. Neurosci. 2017, 62, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Alieva, A.K.; Filatova, E.V.; Karabanov, A.V.; Illarioshkin, S.N.; Limborska, S.A.; Shadrina, M.I.; Slominsky, P.A. MiRNA expression is highly sensitive to a drug therapy in Parkinson’s disease. Parkinsonism Relat. Disord. 2015, 21, 72–74. [Google Scholar] [CrossRef] [PubMed]
- Johns, P. Chapter 14—Multiple sclerosis. In Clinical Neuroscience; Johns, P., Ed.; Churchill Livingstone: London, UK, 2014; pp. 181–196. ISBN 978-0-443-10321-6. [Google Scholar]
- Schaeffer, J.; Cossetti, C.; Mallucci, G.; Pluchino, S. Chapter 30—Multiple sclerosis. In Neurobiology of Brain Disorders; Zigmond, M.J., Rowland, L.P., Coyle, J.T., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 497–520. ISBN 978-0-12-398270-4. [Google Scholar]
- Li, K.X.; Picheca, L. Second-Line Therapy for Patients with Relapsing-Remitting Multiple Sclerosis: A Review of Guidelines; CADTH Rapid Response Reports; Canadian Agency for Drugs and Technologies in Health: Ottawa, ON, Canada, 2019. [Google Scholar]
- Cotsapas, C.; Mitrovic, M.; Hafler, D. Chapter 46—Multiple sclerosis. In Handbook of Clinical Neurology; Neurogenetics, Part II; Geschwind, D.H., Paulson, H.L., Klein, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 148, pp. 723–730. [Google Scholar]
- Montalban, X.; Gold, R.; Thompson, A.J.; Otero-Romero, S.; Amato, M.P.; Chandraratna, D.; Clanet, M.; Comi, G.; Derfuss, T.; Fazekas, F.; et al. ECTRIMS/EAN guideline on the pharmacological treatment of people with multiple sclerosis. Mult. Scler. 2018, 24, 96–120. [Google Scholar] [CrossRef] [Green Version]
- Tur, C.; Moccia, M.; Barkhof, F.; Chataway, J.; Sastre-Garriga, J.; Thompson, A.J.; Ciccarelli, O. Assessing treatment outcomes in multiple sclerosis trials and in the clinical setting. Nat. Rev. Neurol. 2018, 14, 75–93. [Google Scholar] [CrossRef]
- Markowitz, C.E. Interferon-beta: Mechanism of action and dosing issues. Neurology 2007, 68, S8–S11. [Google Scholar] [CrossRef]
- Hecker, M.; Thamilarasan, M.; Koczan, D.; Schröder, I.; Flechtner, K.; Freiesleben, S.; Füllen, G.; Thiesen, H.-J.; Zettl, U.K. MicroRNA expression changes during interferon-beta treatment in the peripheral blood of multiple sclerosis patients. Int. J. Mol. Sci. 2013, 14, 16087–16110. [Google Scholar] [CrossRef] [Green Version]
- Coenen, M.; Hinze, A.V.; Mengel, M.; Fuhrmann, C.; Lüdenbach, B.; Zimmermann, J.; Dykstra, V.; Fimmers, R.; Viviani, R.; Stingl, J.; et al. Immune- and miRNA-response to recombinant interferon beta-1a: A biomarker evaluation study to guide the development of novel type I interferon- based therapies. BMC Pharmacol. Toxicol. 2015, 16, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehtesham, N.; Khorvash, F.; Kheirollahi, M. MiR-145 and miR20a-5p potentially mediate pleiotropic effects of interferon-beta through mitogen-activated protein kinase signaling pathway in multiple sclerosis patients. J. Mol. Neurosci. 2017, 61, 16–24. [Google Scholar] [CrossRef]
- Waschbisch, A.; Atiya, M.; Linker, R.A.; Potapov, S.; Schwab, S.; Derfuss, T. Glatiramer acetate treatment normalizes deregulated microRNA expression in relapsing remitting multiple sclerosis. PLoS ONE 2011, 6, e24604. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Deshpande, M.; Suhail, H.; Rattan, R.; Giri, S. Targeted stage-specific inflammatory microRNA profiling in urine during disease progression in experimental autoimmune encephalomyelitis: Markers of disease progression and drug response. J. Neuroimmune Pharm. 2016, 11, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Bergman, P.; Piket, E.; Khademi, M.; James, T.; Brundin, L.; Olsson, T.; Piehl, F.; Jagodic, M. Circulating miR-150 in CSF is a novel candidate biomarker for multiple sclerosis. Neurol. Neuroimmunol. Neuroinflamm. 2016, 3, e219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenoglio, C.; De Riz, M.; Pietroboni, A.M.; Calvi, A.; Serpente, M.; Cioffi, S.M.G.; Arcaro, M.; Oldoni, E.; Scarpini, E.; Galimberti, D. Effect of fingolimod treatment on circulating miR-15b, miR23a and miR-223 levels in patients with multiple sclerosis. J. Neuroimmunol. 2016, 299, 81–83. [Google Scholar] [CrossRef]
- Sáenz-Cuesta, M.; Alberro, A.; Muñoz-culla, M.; Osorio-Querejeta, I.; Id, M.F.; Lopetegui, I.; Tainta, M.; Prada, Á.; Castillo-triviño, T.; Manuel, J.; et al. The first dose of fingolimod affects circulating extracellular vesicles in multiple sclerosis patients. Int. J. Mol. Sci. 2018, 19, 2448. [Google Scholar] [CrossRef] [Green Version]
- Meira, M.; Sievers, C.; Hoffmann, F.; Derfuss, T.; Kuhle, J.; Kappos, L.; Lindberg, R.L. MiR-126: A novel route for natalizumab action? Mult. Scler. 2014, 20, 1363–1370. [Google Scholar] [CrossRef]
- Meira, M.; Sievers, C.; Hoffmann, F.; Rasenack, M.; Kuhle, J.; Derfuss, T.; Kappos, L.; Lindberg, R.L.P. Unraveling natalizumab effects on deregulated miR-17 expression in CD4+ T cells of patients with relapsing-remitting multiple sclerosis. J. Immunol. Res. 2014, 2014, 897249. [Google Scholar] [CrossRef] [Green Version]
- Sievers, C.; Meira, M.; Hoffmann, F.; Fontoura, P.; Kappos, L.; Lindberg, R.L. Altered microRNA expression in B lymphocytes in multiple sclerosis: Towards a better understanding of treatment effects. Clin. Immunol. 2012, 144, 70–79. [Google Scholar] [CrossRef]
- Ingwersen, J.; Menge, T.; Wingerath, B.; Kaya, D.; Graf, J.; Prozorovski, T.; Keller, A.; Backes, C.; Beier, M.; Scheffler, M.; et al. Natalizumab restores aberrant miRNA expression profile in multiple sclerosis and reveals a critical role for miR-20b. Ann. Clin. Transl. Neurol. 2015, 2, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Mameli, G.; Arru, G.; Caggiu, E.; Niegowska, M.; Leoni, S.; Madeddu, G.; Babudieri, S.; Sechi, G.P.; Sechi, L.A. Natalizumab therapy modulates miR-155, miR-26a and proinflammatory cytokine expression in MS patients. PLoS ONE 2016, 11, e0157153. [Google Scholar] [CrossRef] [PubMed]
- Michell-Robinson, M.A.; Moore, C.S.; Healy, L.M.; Osso, L.A.; Zorko, N.; Grouza, V.; Touil, H.; Poliquin-Lasnier, L.; Trudelle, A.-M.; Giacomini, P.S.; et al. Effects of fumarates on circulating and CNS myeloid cells in multiple sclerosis. Ann. Clin. Transl. Neurol. 2016, 3, 27–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaknin-Dembinsky, A.; Charbit, H.; Brill, L.; Abramsky, O.; Gur-Wahnon, D.; Ben-Dov, I.Z.; Lavon, I. Circulating microRNAs as biomarkers for rituximab therapy, in neuromyelitis optica (NMO). J. Neuroinflamm. 2016, 13, 179. [Google Scholar] [CrossRef] [Green Version]
- Eftekharian, M.M.; Komaki, A.; Mazdeh, M.; Arsang-Jang, S.; Taheri, M.; Ghafouri-Fard, S. Expression profile of selected microRNAs in the peripheral blood of multiple sclerosis patients: A multivariate statistical analysis with ROC curve to find new biomarkers for fingolimod. J. Mol. Neurosci. 2019, 68, 153–161. [Google Scholar] [CrossRef]
- Ebrahimkhani, S.; Beadnall, H.N.; Wang, C.; Suter, C.M.; Barnett, M.H.; Buckland, M.E.; Vafaee, F. Serum exosome microRNAs predict multiple sclerosis disease activity after fingolimod treatment. Mol. Neurobiol. 2019, 1–14. [Google Scholar] [CrossRef]
- Manna, I.; Iaccino, E.; Dattilo, V.; Barone, S.; Vecchio, E.; Mimmi, S.; Filippelli, E.; Demonte, G.; Polidoro, S.; Granata, A.; et al. Exosome-associated miRNA profile as a prognostic tool for therapy response monitoring in multiple sclerosis patients. FASEB J. 2018, 32, 4241–4246. [Google Scholar] [CrossRef] [Green Version]
- Fattahi, M.; Rezaei, N.; Nematalahi, F.S.; Shaygannejad, V.; Fouladi, S.; Karimi, L.; Fathi, F.; Dehghani, L.; Mirmosayyeb, O.; Eskandari, N. MicroRNA-29b variants and MxA expression change during interferon beta therapy in patients with relapsing-remitting multiple sclerosis. Mult. Scler. Relat. Disord. 2019, 35, 241–245. [Google Scholar] [CrossRef]
- Fattahi, M.; Eskandari, N.; Sotoodehnejadnematalahi, F.; Shaygannejad, V.; Kazemi, M. Comparison of the expression of miR-326 between interferon beta responders and non-responders in relapsing-remitting multiple sclerosis. Cell J. 2020, 22, 92–95. [Google Scholar]
- De Felice, B.; Mondola, P.; Sasso, A.; Orefice, G.; Bresciamorra, V.; Vacca, G.; Biffali, E.; Borra, M.; Pannone, R. Small non-coding RNA signature in multiple sclerosis patients after treatment with interferon-β. BMC Med. Genom. 2014, 7, 26. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Culla, M.; Irizar, H.; Castillo-Triviño, T.; Sáenz-Cuesta, M.; Sepúlveda, L.; Lopetegi, I.; de Munain, A.L.; Olascoaga, J.; Baranzini, S.; Otaegui, D. Blood miRNA expression pattern is a possible risk marker for natalizumab-associated progressive multifocal leukoencephalopathy in multiple sclerosis patients. Mult. Scler. 2014, 20, 1851–1859. [Google Scholar] [CrossRef]
- Li, M.-P.; Hu, Y.-D.; Hu, X.-L.; Zhang, Y.-J.; Yang, Y.-L.; Jiang, C.; Tang, J.; Chen, X.-P. MiRNAs and miRNA polymorphisms modify drug response. Int. J. Environ. Res. Public Health 2016, 13, 1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, E.; Lv, Y.; Meng, F.; Yu, X.; Zhang, Y.; Wang, S.; Liu, X.; Liu, D.; Wang, J.; Li, X.; et al. CREAM: A database for chemotherapy resistance-associated miRSNP. Cell Death Dis. 2014, 5, e1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, P.J. MicroRNA polymorphisms: A giant leap towards personalized medicine. Per Med 2009, 6, 119–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labib, D.A.; Shaker, O.G.; El Refai, R.M.; Ghoniem, S.A.; Elmazny, A. Association between miRNA-146a and polymorphisms of its target gene, IRAK1, regarding susceptibility to and clinical features of systemic lupus erythematous and multiple sclerosis. Lab. Med. 2019, 50, 34–41. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Z.G.; Lu, M.; Wang, X.; Shang, X.; Elias, S.B.; Chopp, M. MiR-146a promotes remyelination in a cuprizone model of demyelinating injury. Neuroscience 2017, 348, 252–263. [Google Scholar] [CrossRef]
- Wang, G.; van der Walt, J.M.; Mayhew, G.; Li, Y.-J.; Züchner, S.; Scott, W.K.; Martin, E.R.; Vance, J.M. Variation in the miRNA-433 binding site of FGF20 confers risk for Parkinson disease by overexpression of α-synuclein. Am. J. Hum. Genet. 2008, 82, 283–289. [Google Scholar] [CrossRef] [Green Version]
- Polymeropoulos, M.H.; Lavedan, C.; Leroy, E.; Ide, S.E.; Dehejia, A.; Dutra, A.; Pike, B.; Root, H.; Rubenstein, J.; Boyer, R.; et al. Mutation in the α-synuclein gene identified in families with Parkinson’s Disease. Science 1997, 276, 2045–2047. [Google Scholar] [CrossRef] [Green Version]
- Ghanbari, M.; Ikram, M.A.; de Looper, H.W.J.; Hofman, A.; Erkeland, S.J.; Franco, O.H.; Dehghan, A. Genome-wide identification of microRNA-related variants associated with risk of Alzheimer’s disease. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Cacabelos, R. (Ed.) Pharmacoepigenetics; Academic Press: Cambridge, MA, USA, 2019; ISBN 978-0-12-813940-0. [Google Scholar]
- Hanna, J.; Hossain, G.S.; Kocerha, J. The potential for microRNA therapeutics and clinical research. Front. Genet. 2019, 10, 478. [Google Scholar] [CrossRef] [Green Version]
- Wen, M.M. Getting miRNA therapeutics into the target cells for neurodegenerative diseases: A mini-review. Front. Mol. Neurosci. 2016, 9, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonneau, E.; Neveu, B.; Kostantin, E.; Tsongalis, G.J.; De Guire, V. How close are miRNAs from clinical practice? A perspective on the diagnostic and therapeutic market. EJIFCC 2019, 30, 114–127. [Google Scholar] [PubMed]



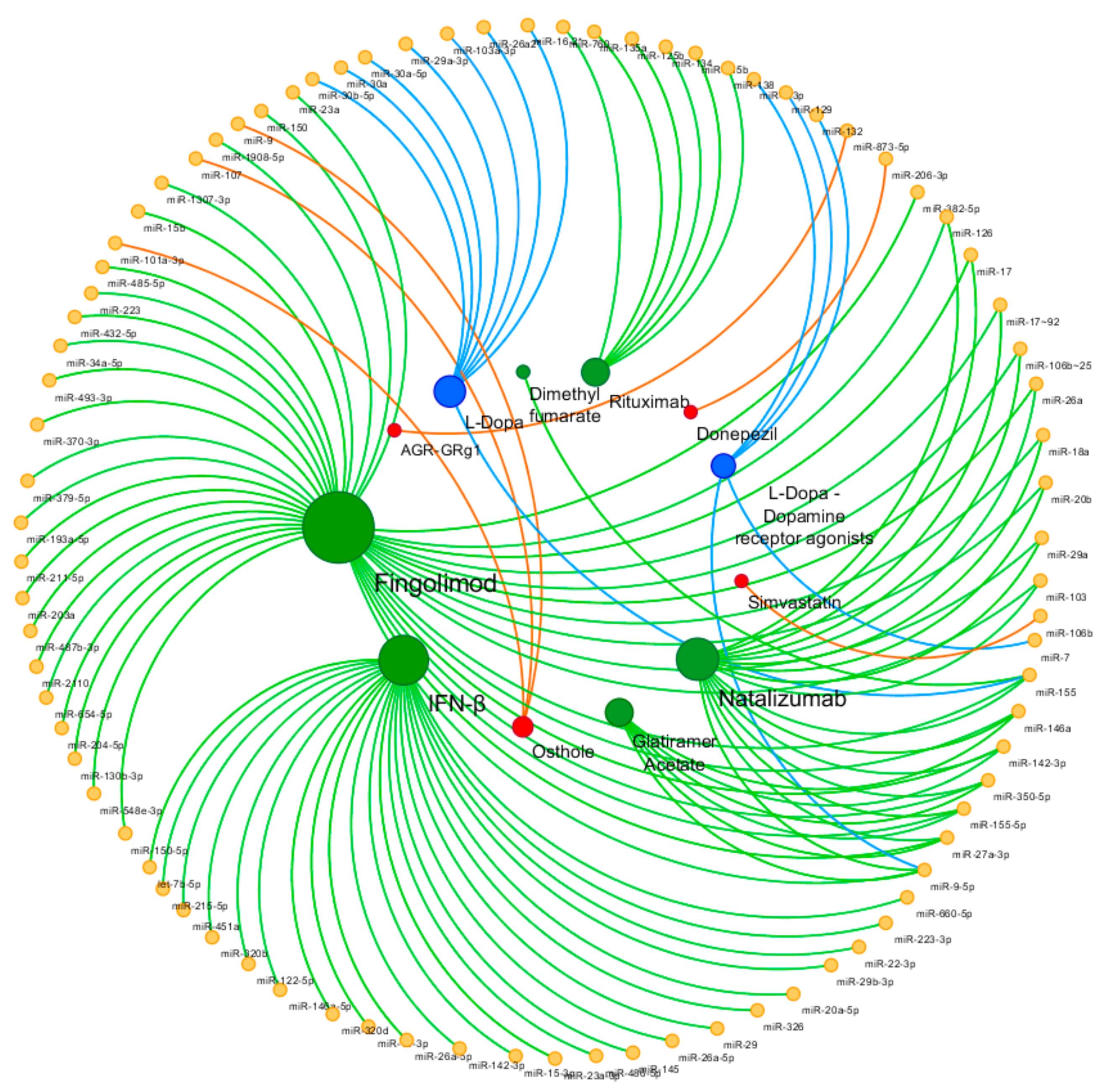{kind=link}
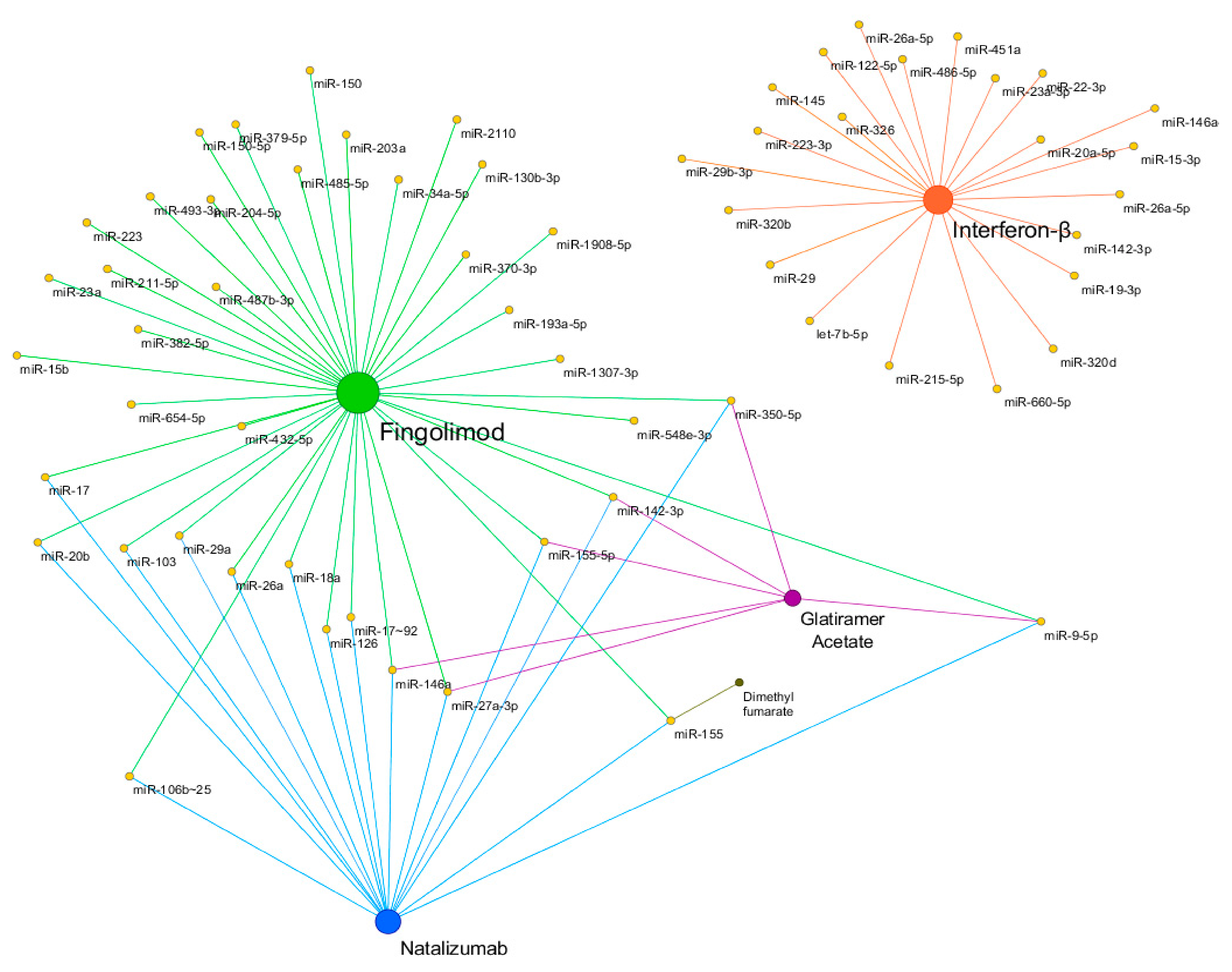{kind=link}
| Disease | Drug | miRNA | Source | Comparison | Ref. |
|---|---|---|---|---|---|
| Alzheimer’s Disease | Donepezil | miR-206-3p | Hippocampus, cortex - Mouse | Treat vs Non treat | [43] |
| Simvastatin | miR-106b | SH-SY5Y cells; APP/PS1 mice brain tissues | Treat vs Non treat | [47] | |
| Osthole | miR-9 | APP-overexpressed cells | Treat vs Non treat | [45,54] | |
| miR-107 | APP-overexpressed cells; APP/PS1 mice | Treat vs Non treat | [48] | ||
| miR-101a-3p | APP-overexpressed cells; APP/PS1 mice | Treat vs Non treat | [52] | ||
| AGR-GRg1 | miR-873-5p | Hippocampus - Mouse | Treat vs Non treat | [55] | |
| Parkinson’s disease | l-Dopa | miR-30b-5p, miR-30a-5p | Plasma | Treat vs Non treat | [69] |
| miR-29a-3p, miR-30b-5p, miR-103a-3p | PBMCs | Treat vs Non treat | [62] | ||
| miR-16-2-3p, miR-26a-2-3p, miR-30a | Peripheral blood | Treat vs Non treat | [58] | ||
| miR-155 | PBMCs | Treat vs Non treat | [61] | ||
| l-Dopa Dopamine receptor agonists Amantadine | miR-7, miR-9-3p, miR-9-5p, miR-129, miR-132 | Peripheral blood | Treat vs Non treat | [70] | |
| Multiple Sclerosis | Interferon-β | miR-29 | PBMCs | Treat vs Non treat | [78] |
| miR-145, miR-20a-5p | Whole blood | Treat vs Non treat | [80] | ||
| miR-22-3p, miR-660-5p, miR-486-5p, miR-451a, let-7b-5p, miR-320b, miR-122-5p, miR-215-5p, miR-320d, miR-19-3p, miR-26a-5p, miR-142-3p, miR-146a-5p, miR-15-3p, miR-23a-3p, miR-223-3p | Exosome | Res vs Non res | [95] | ||
| miR-29b-3p | PBMCs | Res vs Non res | [96] | ||
| miR-326 | PBMCs | Res vs Non res | [97] | ||
| miR-26a-5p | PBMCs | Res vs Non res | [98] | ||
| Glatiramer acetate | miR-146a, miR-142-3p | PBMCs | Treat vs Non treat | [81] | |
| miR-155-5p, miR-27a-3p, miR-9-5p, miR-350-5p | Plasma and urine exosome | Treat vs Non treat | [82] | ||
| Natalizumab | miR-150 | CSF, Plasma | Treat vs Non treat | [83] | |
| miR-126, miR-17 | CD4 + T cells | Treat vs Non treat | [86,87] | ||
| miR-17~92, miR-106b~25 | B lymphocytes | Treat vs Non treat | [88] | ||
| miR-18a, miR-20b, miR-29a, miR-103 | Blood, CD4 + T cells | Treat vs Non treat | [89] | ||
| miR-26a, miR-155 | PBMCs | Treat vs Non treat | [90] | ||
| miR-155 | Monocytes | Treat vs Non treat | [91] | ||
| Dimethyl fumarate | miR-155 | Monocytes | Treat vs Non treat | [91] | |
| Fingolimod | miR-150 | Plasma | Treat vs Non treat | [83] | |
| miR-23a, miR-15b, miR-223 | Blood | Treat vs Non treat | [84] | ||
| miR-34a-5p, miR-211-5p, miR-204-5p | Peripheral blood | Res vs Non res | [93] | ||
| miR-150-5p, miR-548e-3p, miR-130b-3p, miR-654-5p, miR-487b-3p, miR-203a, miR-193a-5p, miR-379-5p, miR-370-3p, miR-382-5p, miR-493-3p, miR-432-5p, miR-485-5p, miR-2110, miR-1307-3p, miR-1908-5p | Serum exosomal | Stable res vs Positive res | [94] | ||
| Progressive Multifocal Leukoencephalopathy | Natalizumab | miR-320, miR-320b, miR-629 | Blood | Treat vs Non treat | [99] |
| Neuromyelitis Optica | Rituximab | miR-125b, miR-760, miR-135a, miR-134, miR-138, miR-135b | Blood | Treat vs Non treat | [92] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nuzziello, N.; Ciaccia, L.; Liguori, M. Precision Medicine in Neurodegenerative Diseases: Some Promising Tips Coming from the microRNAs’ World. Cells 2020, 9, 75. https://doi.org/10.3390/cells9010075
Nuzziello N, Ciaccia L, Liguori M. Precision Medicine in Neurodegenerative Diseases: Some Promising Tips Coming from the microRNAs’ World. Cells. 2020; 9(1):75. https://doi.org/10.3390/cells9010075
Chicago/Turabian StyleNuzziello, Nicoletta, Loredana Ciaccia, and Maria Liguori. 2020. "Precision Medicine in Neurodegenerative Diseases: Some Promising Tips Coming from the microRNAs’ World" Cells 9, no. 1: 75. https://doi.org/10.3390/cells9010075
APA StyleNuzziello, N., Ciaccia, L., & Liguori, M. (2020). Precision Medicine in Neurodegenerative Diseases: Some Promising Tips Coming from the microRNAs’ World. Cells, 9(1), 75. https://doi.org/10.3390/cells9010075