Mesenchymal Stem Cells as a Bio Organ for Treatment of Female Infertility

 and
and
Abstract
:1. Introduction
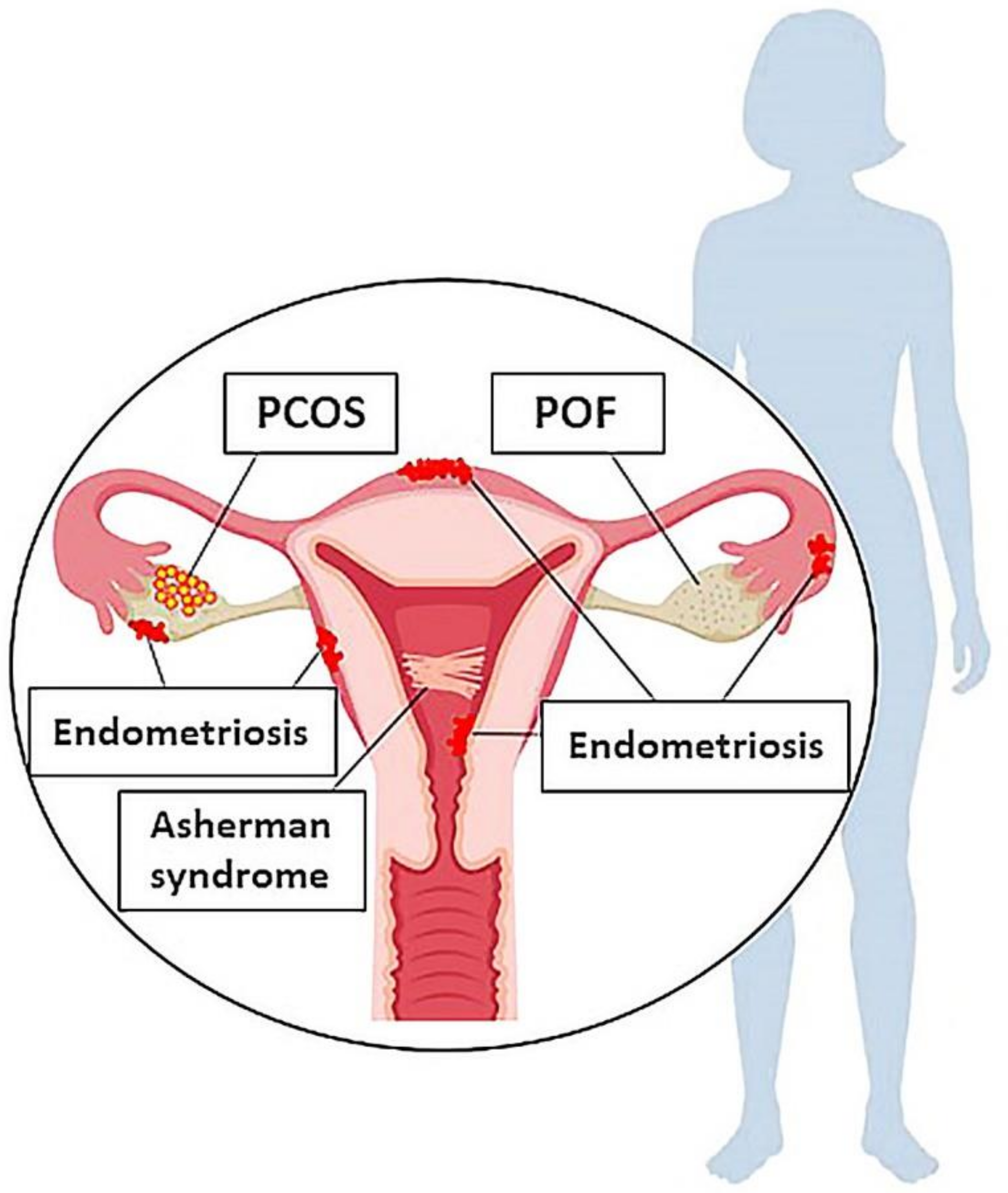
2. Female Reproductive System Disorders
2.1. POF
2.2. PCOS
2.3. Endometriosis
2.4. Asherman syndrome
2.5. Preeclampsia
3. Mesenchymal Stem cells (MSCs)
3.1. Bone Marrow Stromal Cells (BMSCs)
3.2. Adipose-Derived Stem Cells (ADSCs)
3.3. Menstrual Blood-Derived Mesenchymal Stem Cells (MenSCs)
3.4. Umbilical Cord Mesenchymal Stem Cells (UC-MSCs)
3.5. Amniotic Fluid Stem Cells (AFSCs)
3.6. Amnion-Derived Mesenchymal Stem Cells (AD-MSCs)
3.7. Placenta Mesenchymal Stem Cells (PMSCs)
3.8. Pluripotent Stem Cell-derived MSCs (PSC-MSCs)
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hull, M.; Glazener, C.M.; Kelly, N.J.; Conway, D.I.; Foster, P.A.; Hinton, R.A.; Coulson, C.; Lambert, P.A.; Watt, E.M.; Desai, K.M. Population study of causes, treatment, and outcome of infertility. Br. Med. J. (Clin. Res. Ed.) 1985, 291, 1693–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Gong, T.T.; Jiang, Y.T.; Zhang, S.; Zhao, Y.H.; Wu, Q.J. Global, regional, and national prevalence and disability-adjusted life-years for infertility in 195 countries and territories, 1990–2017: Results from a global burden of disease study, 2017. Aging 2019, 11, 10952–10991. [Google Scholar] [CrossRef] [PubMed]
- Jobling, P.; O’Hara, K.; Hua, S. Female reproductive tract pain: Targets, challenges, and outcomes. Front. Pharmacol. 2014, 5, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crain, D.A.; Janssen, S.J.; Edwards, T.M.; Heindel, J.; Ho, S.M.; Hunt, P.; Iguchi, T.; Juul, A.; McLachlan, J.A.; Schwartz, J.; et al. Female reproductive disorders: The roles of endocrine-disrupting compounds and developmental timing. Fertil. Steril. 2008, 90, 911–940. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.X.; Chen, S.R.; Su, P.P.; Huang, F.H.; Shi, Y.C.; Shi, Q.Y.; Lin, S. Using Mesenchymal Stem Cells to Treat Female Infertility: An Update on Female Reproductive Diseases. Stem Cells Int. 2019, 2019, 9071720. [Google Scholar] [CrossRef]
- Naji, A.; Rouas-Freiss, N.; Durrbach, A.; Carosella, E.D.; Sensébé, L.; Deschaseaux, F. Concise review: Combining human leukocyte antigen G and mesenchymal stem cells for immunosuppressant biotherapy. Stem Cells (Dayt. Ohio) 2013, 31, 2296–2303. [Google Scholar] [CrossRef]
- Squillaro, T.; Peluso, G.; Galderisi, U. Clinical Trials With Mesenchymal Stem Cells: An Update. Cell Transplant. 2016, 25, 829–848. [Google Scholar] [CrossRef] [Green Version]
- Galipeau, J.; Sensébé, L. Mesenchymal Stromal Cells: Clinical Challenges and Therapeutic Opportunities. Cell Stem Cell 2018, 22, 824–833. [Google Scholar] [CrossRef] [Green Version]
- Trounson, A.; McDonald, C. Stem Cell Therapies in Clinical Trials: Progress and Challenges. Cell Stem Cell 2015, 17, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human mesenchymal stem cells—Current trends and future prospective. Biosci. Rep. 2015, 35, e00191. [Google Scholar] [CrossRef]
- Spitzhorn, L.; Megges, M.; Wruck, W.; Rahman, S.; Otte, J.; Degistirici, O.; Meisel, R.; Sorg, R.V.; Oreffo, R.O.C.; Adjave, J. Human iPSC-derived MSCs (iMSCs) from aged individuals acquire a rejuvenation signature. Stem Cell Res. Ther. 2019, 10, 100. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.L.; Zhang, Y.; Li, X.; Fu, Q.L. Mechanisms underlying the protective effects of mesenchymal stem cell-based therapy. Cell. Mol. Life Sci. 2020, 77, 2771–2794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rungsiwiwut, R.; Virutamasen, P.; Pruksananonda, K. Mesenchymal stem cells for restoring endometrial function: An infertility perspective. Reprod Med. Biol. 2020, 00, 1–7. [Google Scholar] [CrossRef]
- Yoon, S.Y. Mesenchymal stem cells for restoration of ovarian function. Clin. Exp. Reprod. Med. 2019, 46, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C. The roles of different stem cells on premature ovarian failure. Curr. Stem Cell Res. Ther. 2020, 15, 473–481. [Google Scholar]
- Chen, L.; Guo, S.; Wei, C.; Li, H.; Wang, H.; Xu, Y. Effect of stem cell transplantation of premature ovarian failure in animal models and patients: A meta-analysis and case report. Exp. Ther. Med. 2018, 15, 4105–4118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoek, A.; Schoemaker, J.; Drexhage, H.A. Premature Ovarian Failure and Ovarian Autoimmunity. Endocr. Rev. 1997, 18, 107–134. [Google Scholar]
- Rocha, A.L.; Oliveira, F.R.; Azevedo, R.C.; Silva, V.A.; Peres, T.M.; Candido, A.L.; Gomes, K.B.; Reis, F.M. Recent advances in the understanding and management of polycystic ovary syndrome. F1000Research 2019, 8. F1000 Faculty Rev-565. [Google Scholar] [CrossRef] [Green Version]
- Ndefo, U.A.; Eaton, A.; Green, M.R. Polycystic ovary syndrome: A review of treatment options with a focus on pharmacological approaches. Pharm. Ther. 2013, 38, 336–355. [Google Scholar]
- Dallenbach-Hellweg, G. The endometrium of infertility. A review. Pathol. Res. Pract. 1984, 178, 527–537. [Google Scholar] [CrossRef]
- Maybin, J.A.; Critchley, H.O. Menstrual physiology: Implications for endometrial pathology and beyond. Hum. Reprod. Update 2015, 21, 748–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasson, I.E.; Taylor, H.S. Stem cells and the pathogenesis of endometriosis. Ann. N. Y. Acad. Sci. 2008, 1127, 106–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceccaroni, M.; Roviglione, G.; Rosenberg, P.; Pesci, A.; Clarizia, R.; Bruni, F.; Zardini, C.; Ruffo, G.; Placci, A.; Crippa, S. Pericardial, pleural and diaphragmatic endometriosis in association with pelvic peritoneal and bowel endometriosis: A case report and review of the literature. Wideochirurgia I Inne Tech. Maloinwazyjne = Videosurg. Other Miniinvasive Tech. 2012, 7, 122–131. [Google Scholar] [CrossRef]
- Klemmt, P.; Starzinski-Powitz, A. Molecular and Cellular Pathogenesis of Endometriosis. Curr. Women’s Health Rev. 2018, 14, 106–116. [Google Scholar] [CrossRef]
- Dreisler, E.; Kjer, J.J. Asherman’s syndrome: Current perspectives on diagnosis and management. Int. J. Women’s Health 2019, 11, 191–198. [Google Scholar] [CrossRef] [PubMed]
- March, C.M. Asherman’s syndrome. Semin. Reprod. Med. 2011, 29, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Lemoine, E.; Granger, J.P.; Karumanchi, S.A. Preeclampsia: Pathophysiology, Challenges, and Perspectives. Circ. Res. 2019, 124, 1094–1112. [Google Scholar] [CrossRef] [PubMed]
- Uzan, J.; Carbonnel, M.; Piconne, O.; Asmar, R.; Ayoubi, J.M. Pre-eclampsia: Pathophysiology, diagnosis, and management. Vasc. Health Risk Manag. 2011, 7, 467–474. [Google Scholar]
- Phipps, E.; Prasanna, D.; Brima, W.; Jim, B. Preeclampsia: Updates in Pathogenesis, Definitions, and Guidelines. Clin. J. Am. Soc. Nephrol. 2016, 11, 1102–1113. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.j.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Krampera, M.; Galipeau, J.; Shi, Y.; Tarte, K.; Sensebe, L.; MSC Committee of the International Society for Cellular Therapy (ISCT). Immunological characterization of multipotent mesenchymal stromal cells--The International Society for Cellular Therapy (ISCT) working proposal. Cytotherapy 2013, 15, 1054–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galipeau, J.; Krampera, M.; Barrett, J.; Dazzi, F.; Deans, R.J.; DeBruijn, J.; Dominici, M.; Fibbe, W.E.; Gee, A.P.; Gimble, J.M.; et al. International Society for Cellular Therapy perspective on immune functional assays for mesenchymal stromal cells as potency release criterion for advanced phase clinical trials. Cytotherapy 2016, 18, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrell, C.R.; Fellabaum, C.; Jovicic, N.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Molecular Mechanisms Responsible for Therapeutic Potential of Mesenchymal Stem Cell-Derived Secretome. Cells 2019, 8, 467. [Google Scholar] [CrossRef] [Green Version]
- Ling, L.; Feng, X.; Wei, T.; Wang, Y.; Wang, Y.; Wang, Z.; Tang, D.; Luo, Y.; Xiong, Z. Human amnion-derived mesenchymal stem cell (hAD-MSC) transplantation improves ovarian function in rats with premature ovarian insufficiency (POI) at least partly through a paracrine mechanism. Stem Cell Res. Ther. 2019, 10, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Chiu, S.; Liang, X.; Gao, F.; Zhang, Z.; Liao, S.; Liang, Y.; Chai, Y.H.; Low, D.J.; Tse, H.F. Rap1-mediated nuclear factor-kappaB (NF-κB) activity regulates the paracrine capacity of mesenchymal stem cells in heart repair following infarction. Cell Death Discov. 2015, 1, 15007. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.; Xiong, X.L.; Xiao, N.; He, K.; Chen, M.S.; Peng, J.; Su, X.; Mei, H.; Dai, Y.N.; Wei, D.; et al. Mesenchymal Stem Cells Alleviate DHEA-Induced Polycystic Ovary Syndrome (PCOS) by Inhibiting Inflammation in Mice. Stem Cells Int. 2019, 12. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.B.; Hao, J.X.; Meng, T.G.; Guo, L.; Dong, M.Z.; Fan, L.H.; Ouyang, Y.C.; Wang, G.; Sun, Q.Y.; Ou, X.H.; et al. Transfer of autologous mitochondria from adipose tissue-derived stem cells rescues oocyte quality and infertility in aged mice. Aging 2017, 9, 2480–2488. [Google Scholar] [CrossRef] [Green Version]
- Court, A.C.; Le-Gatt, A.; Luz-Crawford, P.; Parra, E.; Aliaga-Tobar, V.; Bátiz, L.F.; Contreras, R.A.; Ortúzar, M.I.; Kurte, M.; Elizondo-Vega, R. Mitochondrial transfer from MSCs to T cells induces Treg differentiation and restricts inflammatory response. EMBO Rep. 2020, 21, e48052. [Google Scholar] [CrossRef]
- Owen, M.; Friedenstein, A.J. Stromal stem cells: Marrow-derived osteogenic precursors. Ciba Found. Symp. 1988, 136, 42–60. [Google Scholar]
- Altaner, C.; Altanerova, V.; Cihova, M.; Hunakova, L.; Kaiserova, K.; Klepanec, A.; Vulev, I.; Madaric, J. Characterization of mesenchymal stem cells of “no-options” patients with critical limb ischemia treated by autologous bone marrow mononuclear cells. PLoS ONE 2013, 8, e73722. [Google Scholar] [CrossRef]
- Gao, L.; Huang, Z.; Lin, H.; Tian, Y.; Li, P.; Lin, S. Bone Marrow Mesenchymal Stem Cells (BMSCs) Restore Functional Endometrium in the Rat Model for Severe Asherman Syndrome. Reprod. Sci. (Thousand Oakscalif.) 2019, 26, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tal, R.; Pluchino, N.; Mamillapalli, R.; Taylor, H.S. Systemic administration of bone marrow-derived cells leads to better uterine engraftment than use of uterine-derived cells or local injection. J. Cell. Mol. Med. 2018, 22, 67–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tepper, O.M.; Sealove, B.A.; Murayama, T.; Asahara, T. Newly emerging concepts in blood vessel growth: Recent discovery of endothelial progenitor cells and their function in tissue regeneration. J. Investig. Med. 2003, 51, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Besikcioglu, H.E.; Sarıbas, G.S.; Ozogul, C.; Tiryaki, M.; Kilic, S.; Pınarlı, F.A.; Gulbahar, O. Determination of the effects of bone marrow derived mesenchymal stem cells and ovarian stromal stem cells on follicular maturation in cyclophosphamide induced ovarian failure in rats. Taiwan. J. Obstet. Gynecol. 2019, 58, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Yang, X.; Han, Z.P.; Qu, F.F.; Shao, L.; Shi, Y.F. Mesenchymal stem cells: A new trend for cell therapy. Acta Pharmacol. Sin. 2013, 34, 747–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abd-Allah, S.H.; Shalaby, S.M.; Pasha, H.F.; El-Shal, A.S.; Raafat, N.; Shabrawy, S.M.; Awad, H.A.; Amer, M.G.; Gharib, M.A.; El Gendy, E.A.; et al. Mechanistic action of mesenchymal stem cell injection in the treatment of chemically induced ovarian failure in rabbits. Cytotherapy 2013, 15, 64–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, S.A.; Shalaby, S.M.; Abdelaziz, M.; Brakta, S.; Hill, W.D.; Ismail, N.; Al-Hendy, A. Human Mesenchymal Stem Cells Partially Reverse Infertility in Chemotherapy-Induced Ovarian Failure. Reprod. Sci. (Thousand Oakscalif.) 2018, 25, 51–63. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; He, Y.; Wang, X.; Peng, D.; Chen, X.; Li, X.; Wang, Q. Overexpression of miR-21 in stem cells improves ovarian structure and function in rats with chemotherapy-induced ovarian damage by targeting PDCD4 and PTEN to inhibit granulosa cell apoptosis. Stem Cell Res. Ther. 2017, 8, 187. [Google Scholar] [CrossRef] [Green Version]
- Badawy, A.; Sobh, M.A.; Ahdy, M.; Abdelhafez, M.S. Bone marrow mesenchymal stem cell repair of cyclophosphamide-induced ovarian insufficiency in a mouse model. Int. J. Women’s Health 2017, 9, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Taylor, H.S. Endometrial cells derived from donor stem cells in bone marrow transplant recipients. JAMA 2004, 292, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Nagori, C.B.; Panchal, S.Y.; Patel, H. Endometrial regeneration using autologous adult stem cells followed by conception by in vitro fertilization in a patient of severe Asherman’s syndrome. J. Hum. Reprod. Sci. 2011, 4, 43–48. [Google Scholar] [PubMed]
- Wang, J.; Ju, B.; Pan, C.; Gu, Y.; Zhang, Y.; Sun, L.; Zhang, B.; Zhang, Y. Application of Bone Marrow-Derived Mesenchymal Stem Cells in the Treatment of Intrauterine Adhesions in Rats. Cell. Physiol. Biochem. 2016, 39, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Mohanty, S.; Seth, T.; Shankar, M.; Bhaskaran, S.; Dharmendra, S. Autologous stem cell transplantation in refractory Asherman’s syndrome: A novel cell based therapy. J. Hum. Reprod. Sci. 2014, 7, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, X.; Cabanillas, S.; Cervelló, I.; Arbona, C.; Raga, F.; Ferro, J.; Palmero, J.; Remohí, J.; Pellicer, A.; Simón, C. Autologous cell therapy with CD133+ bone marrow-derived stem cells for refractory Asherman’s syndrome and endometrial atrophy: A pilot cohort study. Hum. Reprod. (Oxf. Engl. ) 2016, 31, 1087–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervelló, I.; Gil-Sanchis, C.; Santamaría, X.; Cabanillas, S.; Díaz, A.; Faus, A.; Pellicer, A.; Simón, C. Human CD133(+) bone marrow-derived stem cells promote endometrial proliferation in a murine model of Asherman syndrome. Fertil. Steril. 2015, 104, 1552–1560.e603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alawadhi, F.; Du, H.; Cakmak, H.; Taylor, H.S. Bone Marrow-Derived Stem Cell (BMDSC) transplantation improves fertility in a murine model of Asherman’s syndrome. PLoS ONE 2014, 9, e96662. [Google Scholar] [CrossRef]
- Zhao, G.; Cao, Y.; Zhu, X.; Tang, X.; Ding, L.; Sun, H.; Li, J.; Li, X.; Dai, C.; Ru, T. Transplantation of collagen scaffold with autologous bone marrow mononuclear cells promotes functional endometrium reconstruction via downregulating ΔNp63 expression in Asherman’s syndrome. Sci. China. Life Sci. 2017, 60, 404–416. [Google Scholar] [CrossRef] [Green Version]
- Gangaraju, V.K.; Lin, H. MicroRNAs: Key regulators of stem cells. Nat. Reviews. Mol. Cell Biol. 2009, 10, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Tian, C.; Liu, L.; Ye, X.; Fu, H.; Sheng, X.; Wang, L.; Wang, H.; Heng, D.; Liu, L. Functional Oocytes Derived from Granulosa Cells. Cell Rep. 2019, 29, 4256–4267. [Google Scholar] [CrossRef] [Green Version]
- Aghabozorgi, A.S.; Ahangari, N.; Eftekhaari, T.E.; Torbati, P.N.; Bahiraee, A.; Ebrahimi, R.; Pasdar, A. Circulating exosomal miRNAs in cardiovascular disease pathogenesis: New emerging hopes. J. Cell. Physiol. 2019, 234, 21796–21809. [Google Scholar] [CrossRef]
- Sun, B.; Ma, Y.; Wang, F.; Hu, L.; Sun, Y. miR-644-5p carried by bone mesenchymal stem cell-derived exosomes targets regulation of p53 to inhibit ovarian granulosa cell apoptosis. Stem Cell Res. Ther. 2019, 10, 360. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Lin, L.; Sha, C.; Li, T.; Zhao, D.; Wei, H.; Chen, Q.; Liu, Y.; Chen, X.; Xu, W. Bone marrow mesenchymal stem cell-derived exosomal miR-144-5p improves rat ovarian function after chemotherapy-induced ovarian failure by targeting PTEN. Lab. Investig. 2020, 100, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Esfandyari, S.; Al-Hendy, A. Evolving role of microRNAs in uterine fibroid pathogenesis: Filling the gap! Fertil. Steril. 2020, 113, 1167–1168. [Google Scholar] [CrossRef] [PubMed]
- Lendeckel, S.; Jödicke, A.; Christophis, P.; Heidinger, K.; Wolff, J.; Fraser, J.K.; Hedrick, M.H.; Berthold, L.; Howaldt, H.P. Autologous stem cells (adipose) and fibrin glue used to treat widespread traumatic calvarial defects: Case report. J. Cranio-Maxillo-Facial Surg. 2004, 32, 370–373. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.A.; Chung, H.M.; Won, C.H.; Sung, J.H. Potential application of adipose-derived stem cells and their secretory factors to skin: Discussion from both clinical and industrial viewpoints. Expert Opin. Biol. Ther. 2010, 10, 495–503. [Google Scholar] [CrossRef]
- Ra, J.C.; Jeong, E.C.; Kang, S.K.; Lee, S.J.; Choi, K.H. A Prospective, Nonrandomized, no Placebo-Controlled, Phase I/II Clinical Trial Assessing the Safety and Efficacy of Intramuscular Injection of Autologous Adipose Tissue-Derived Mesenchymal Stem Cells in Patients With Severe Buerger’s Disease. Cell Med. 2016, 9, 87–102. [Google Scholar] [CrossRef]
- Choudhery, M.S.; Badowski, M.; Muise, A.; Pierce, J.; Harris, D.T. Donor age negatively impacts adipose tissue-derived mesenchymal stem cell expansion and differentiation. J. Transl. Med. 2014, 12, 8. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.H.; Kim, B.; Choi, I.; Kim, H.; Choi, H.S.; Suh, K.; Bae, Y.C.; Jung, J.S. Characterization and expression analysis of mesenchymal stem cells from human bone marrow and adipose tissue. Cell. Physiol. Biochem. 2004, 14, 311–324. [Google Scholar] [CrossRef]
- Gesellschaft, D. Transfusion Medicine and Hemotherapy: Offizielles Organ der Deutschen Gesellschaft für Transfusionsmedizin und Immunhämatologie; Interdisziplinäre Zeitschrift für Infusions-und Hämotherapie, Immunhämatologie und Klinische Hämostaseologie; Karger: Berlin, Germany, 2003. [Google Scholar]
- Damous, L.L.; Nakamuta, J.S.; Carvalho, A.E.; Carvalho, K.C.; Soares, J.M., Jr.; Simões, M.; Krieger, J.E.; Baracat, E.C. Does adipose tissue-derived stem cell therapy improve graft quality in freshly grafted ovaries. Reprod. Biol. Endocrinol. 2015, 13, 108. [Google Scholar] [CrossRef] [Green Version]
- Terraciano, P.; Garcez, T.; Ayres, L.; Durli, I.; Baggio, M.; Kuhl, C.P.; Laurino, C.; Passos, E.; Paz, A.H.; Cirne-Lima, E. Cell therapy for chemically induced ovarian failure in mice. Stem Cells Int. 2014, 720753. [Google Scholar] [CrossRef]
- Sun, M.; Wang, S.; Li, Y.; Yu, L.; Gu, F.; Wang, C.; Yao, Y. Adipose-derived stem cells improved mouse ovary function after chemotherapy-induced ovary failure. Stem Cell Res. Ther. 2013, 4, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, J.; Ding, L.; Cheng, J.; Yang, J.; Li, X.; Yan, G.; Sun, H.; Dai, J.; Hu, Y. Transplantation of adipose-derived stem cells combined with collagen scaffolds restores ovarian function in a rat model of premature ovarian insufficiency. Hum. Reprod. (Oxf. Engl. ) 2016, 31, 1075–1086. [Google Scholar] [CrossRef] [PubMed]
- Abomaray, F.; Gidlöf, S.; Bezubik, B.; Engman, M.; Götherström, C. Mesenchymal Stromal Cells Support Endometriotic Stromal Cells In Vitro. Stem Cells Int. 2018, 7318513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilic, S.; Yuksel, B.; Pinarli, F.; Albayrak, A.; Boztok, B.; Delibasi, T. Effect of stem cell application on Asherman syndrome, an experimental rat model. J. Assist. Reprod. Genet. 2014, 31, 975–982. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Ichim, T.E.; Zhong, J.; Rogers, A.; Yin, Z.; Jackson, J.; Wang, H.; Ge, W.; Bogin, V.; Chan, K.W.; et al. Endometrial regenerative cells: A novel stem cell population. J. Transl. Med. 2007, 5, 57. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.; Patel, A.N.; Ichim, T.E.; Riordan, N.H.; Wang, H.; Min, W.P.; Woods, E.J.; Reid, M.; Mansilla, E.; Marin, G.H. Feasibility investigation of allogeneic endometrial regenerative cells. J. Transl. Med. 2009, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Wolff, E.F.; Mutlu, L.; Massasa, E.E.; Elsworth, J.D.; Eugene Redmond, D., Jr.; Taylor, H.S. Endometrial stem cell transplantation in MPTP- exposed primates: An alternative cell source for treatment of Parkinson’s disease. J. Cell. Mol. Med. 2015, 19, 249–256. [Google Scholar] [CrossRef]
- Wolff, E.F.; Gao, X.B.; Yao, K.V.; Andrews, Z.B.; Du, H.; Elsworth, J.D.; Taylor, H.S. Endometrial stem cell transplantation restores dopamine production in a Parkinson’s disease model. J. Cell. Mol. Med. 2011, 15, 747–755. [Google Scholar] [CrossRef]
- Liu, T.; Huang, Y.; Zhang, J.; Qin, W.; Chi, H.; Chen, J.; Yu, Z.; Chen, C. Transplantation of human menstrual blood stem cells to treat premature ovarian failure in mouse model. Stem Cells Dev. 2014, 23, 1548–1557. [Google Scholar] [CrossRef] [Green Version]
- Manshadi, M.D.; Navid, S.; Hoshino, Y.; Daneshi, E.; Noory, P.; Abbasi, M. The effects of human menstrual blood stem cells-derived granulosa cells on ovarian follicle formation in a rat model of premature ovarian failure. Microscopy Res. Techniquey 2019, 82, 635–642. [Google Scholar] [CrossRef]
- Liebermann, D.A.; Tront, J.S.; Sha, X.; Mukherjee, K.; Mohamed-Hadley, A.; Hoffman, B. Gadd45 stress sensors in malignancy and leukemia. Crit. Rev. Oncog. 2011, 16, 129–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyka-Nouspikel, N.; Desmarais, J.; Gokhale, P.J.; Jones, M.; Meuth, M.; Andrews, P.W.; Nouspikel, T. Deficient DNA damage response and cell cycle checkpoints lead to accumulation of point mutations in human embryonic stem cells. Stem Cells (Dayt. Ohio) 2012, 30, 1901–1910. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Lou, I.C.; Conolly, R.B. Computational modeling of signaling pathways mediating cell cycle checkpoint control and apoptotic responses to ionizing radiation-induced DNA damage. Dose-Response 2012, 10, 251–273. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Guo, F.; Yuan, Q.; Shao, Y.; Zhang, Y.; Wang, H.; Hao, S.; Du, X. Endometrial mesenchymal stem cells isolated from menstrual blood repaired epirubicin-induced damage to human ovarian granulosa cells by inhibiting the expression of Gadd45b in cell cycle pathway. Stem Cell Res. Ther. 2019, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, Y.; Yang, T.; Li, J.; Yang, X. Study of the reparative effects of menstrual-derived stem cells on premature ovarian failure in mice. Stem Cell Res. Ther. 2017, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Chan, R.W.; Ng, E.H.; Yeung, W.S. Identification of cells with colony-forming activity, self-renewal capacity, and multipotency in ovarian endometriosis. Am. J. Pathol. 2011, 178, 2832–2844. [Google Scholar] [CrossRef] [Green Version]
- Kao, A.P.; Wang, K.H.; Chang, C.C.; Lee, J.N.; Long, C.Y.; Chen, H.S.; Tsai, C.F.; Hsieh, T.H.; Tsai, E.M. Comparative study of human eutopic and ectopic endometrial mesenchymal stem cells and the development of an in vivo endometriotic invasion model. Fertil. Steril. 2011, 95, 1308–1315. [Google Scholar] [CrossRef]
- Zhang, S.; Li, P.; Yuan, Z.; Tan, J. Platelet-rich plasma improves therapeutic effects of menstrual blood-derived stromal cells in rat model of intrauterine adhesion. Stem Cell Res. Ther. 2019, 10, 61. [Google Scholar] [CrossRef]
- Zheng, S.X.; Wang, J.; Wang, X.L.; Ali, A.; Wu, L.M.; Liu, Y.S. Feasibility analysis of treating severe intrauterine adhesions by transplanting menstrual blood-derived stem cells. Int. J. Mol. Med. 2018, 41, 2201–2212. [Google Scholar] [CrossRef]
- Tan, J.; Li, P.; Wang, Q.; Li, Y.; Li, X.; Zhao, D.; Xu, X.; Kong, L. Autologous menstrual blood-derived stromal cells transplantation for severe Asherman’s syndrome. Hum. Reprod. (Oxf. Engl.) 2016, 31, 2723–2729. [Google Scholar] [CrossRef] [Green Version]
- Domnina, A.; Novikova, P.; Obidina, J.; Fridlyanskaya, I.; Alekseenko, L.; Kozhukharova, I.; Lyublinskaya, O.; Zenin, V.; Nikolsky, N. Human mesenchymal stem cells in spheroids improve fertility in model animals with damaged endometrium. Stem Cell Res. Ther. 2018, 9, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paduano, F.; Marrelli, M.; Palmieri, F.; Tatullo, M. CD146 Expression Influences Periapical Cyst Mesenchymal Stem Cell Properties. Stem Cell Rev. Rep. 2016, 12, 592–603. [Google Scholar] [CrossRef] [PubMed]
- Nagamura-Inoue, T.; He, H. Umbilical cord-derived mesenchymal stem cells: Their advantages and potential clinical utility. World J. Stem Cells 2014, 6, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.B.; Song, Y.; Chen, Y.; Zhang, F.; Qi, F.Z. Differentiation of umbilical cord mesenchymal stem cells into hepatocytes in comparison with bone marrow mesenchymal stem cells. Mol. Med. Rep. 2018, 18, 2009–2016. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Yun, M.M.; Han, X.; Zhao, R.; Zhou, E.; Yun, S. Human umbilical cord mesenchymal stromal cells suppress MHC class II expression on rat vascular endothelium and prolong survival time of cardiac allograft. Int. J. Clin. Exp. Med. 2014, 7, 1760–1767. [Google Scholar]
- Mohamed, S.A.; Shalaby, S.; Brakta, S.; Elam, L.; Elsharoud, A.; Al-Hendy, A. Umbilical Cord Blood Mesenchymal Stem Cells as an Infertility Treatment for Chemotherapy Induced Premature Ovarian Insufficiency. Biomedicines 2019, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Zhong, Y.; Qian, C.; Zou, Q.; Ou, J.; Shi, Y.; Gao, L.; Wang, G.; Liu, Z.; Li, H. Human Umbilical Cord Mesenchymal Stem Cells Therapy in Cyclophosphamide-Induced Premature Ovarian Failure Rat Model. BioMed Res. Int. 2016, 2517514. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.F.; Hu, H.B.; Xu, H.Y.; Fu, X.F.; Peng, D.X.; Su, W.Y.; He, Y.L. Human umbilical cord mesenchymal stem cell transplantation restores damaged ovaries. J. Cell. Mol. Med. 2015, 19, 2108–2117. [Google Scholar] [CrossRef]
- Wang, S.; Yu, L.; Sun, M.; Mu, S.; Wang, C.; Wang, D.; Yao, Y. The therapeutic potential of umbilical cord mesenchymal stem cells in mice premature ovarian failure. BioMed Res. Int. 2013, 690491. [Google Scholar] [CrossRef]
- Jalalie, L.; Rezaie, M.J.; Jalili, A.; Rezaee, M.A.; Vahabzadeh, Z.; Rahmani, M.R.; Karimipoor, M.; Hakhamaneshi, M.S. Distribution of the CM-Dil-Labeled Human Umbilical Cord Vein Mesenchymal Stem Cells Migrated to the Cyclophosphamide-Injured Ovaries in C57BL/6 Mice. Iran. Biomed. J. 2019, 23, 200–208. [Google Scholar] [CrossRef] [Green Version]
- Elfayomy, A.K.; Almasry, S.M.; El-Tarhouny, S.A.; Eldomiaty, M.A. Human umbilical cord blood-mesenchymal stem cells transplantation renovates the ovarian surface epithelium in a rat model of premature ovarian failure: Possible direct and indirect effects. Tissue Cell 2016, 48, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Yan, G.; Wang, B.; Xu, L.; Gu, Y.; Ru, T.; Cui, X.; Lei, L.; Liu, J.; Sheng, X. Transplantation of UC-MSCs on collagen scaffold activates follicles in dormant ovaries of POF patients with long history of infertility. Sci. China. Life Sci. 2018, 61, 1554–1565. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lei, L.; Wang, S.; Sheng, X.; Yan, G.; Xu, L.; Liu, J.; Liu, M.; Zhen, X.; Ding, L. Transplantation of umbilical cord-derived mesenchymal stem cells on a collagen scaffold improves ovarian function in a premature ovarian failure model of mice. Vitr. Cell. Dev. Biology. Anim. 2019, 55, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Mao, Q.; He, J.; She, H.; Zhang, Z.; Yin, C. Human umbilical cord mesenchymal stem cells improve the reserve function of perimenopausal ovary via a paracrine mechanism. Stem Cell Res. Ther. 2017, 8, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Zhang, M.; Zhang, Y.; Li, W.; Yang, B. Mesenchymal stem cells derived from Wharton jelly of the human umbilical cord ameliorate damage to human endometrial stromal cells. Fertil. Steril. 2011, 96, 1029–1036. [Google Scholar] [CrossRef]
- Fan, D.; Wu, S.; Ye, S.; Wang, W.; Guo, X.; Liu, Z. Umbilical cord mesenchyme stem cell local intramuscular injection for treatment of uterine niche: Protocol for a prospective, randomized, double-blinded, placebo-controlled clinical trial. Medicine 2017, 96, e8480. [Google Scholar] [CrossRef]
- Shi, Q.; Gao, J.; Jiang, Y.; Sun, B.; Lu, W.; Su, M.; Xu, Y.; Yang, X.; Zhang, Y. Differentiation of human umbilical cord Wharton’s jelly-derived mesenchymal stem cells into endometrial cells. Stem Cell Res. Ther. 2017, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, Y.; Guan, C.Y.; Tian, S.; Lv, X.D.; Li, J.H.; Ma, X.; Xia, H.F. Therapeutic effect of human umbilical cord-derived mesenchymal stem cells on injured rat endometrium during its chronic phase. Stem Cell Res. Ther. 2018, 9, 36. [Google Scholar] [CrossRef] [Green Version]
- Xin, L.; Lin, X.; Pan, Y.; Zheng, X.; Shi, L.; Zhang, Y.; Ma, L.; Gao, C.; Zhang, S. A collagen scaffold loaded with human umbilical cord-derived mesenchymal stem cells facilitates endometrial regeneration and restores fertility. Acta Biomater. 2019, 92, 160–171. [Google Scholar] [CrossRef]
- Xu, L.; Ding, L.; Wang, L.; Cao, Y.; Zhu, H.; Lu, J.; Li, X.; Song, T.; Hu, Y.; Dai, J. Umbilical cord-derived mesenchymal stem cells on scaffolds facilitate collagen degradation via upregulation of MMP-9 in rat uterine scars. Stem Cell Res. Ther. 2017, 8, 84. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Sun, H.; Zhu, H.; Zhu, X.; Tang, X.; Yan, G.; Wang, J.; Bai, D.; Wang, J.; Wang, L.; et al. Allogeneic cell therapy using umbilical cord MSCs on collagen scaffolds for patients with recurrent uterine adhesion: A phase I clinical trial. Stem Cell Res. Ther. 2018, 9, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.L.; Yu, Y.; Guan, H.B.; Qiao, C. Effect of Human Umbilical Cord Mesenchymal Stem Cell Transplantation in a Rat Model of Preeclampsia. Reprod. Sci. (Thousand Oakscalif.) 2016, 23, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Liu, Y.; Zhang, D.; Xie, J.; Guan, H.; Shang, T. Beneficial effect of human umbilical cord-derived mesenchymal stem cells on an endotoxin-induced rat model of preeclampsia. Exp. Ther. Med. 2015, 10, 1851–1856. [Google Scholar] [CrossRef]
- Zhang, D.; Fu, L.; Wang, L.; Lin, L.; Yu, L.; Zhang, L.; Shang, T. Therapeutic benefit of mesenchymal stem cells in pregnant rats with angiotensin receptor agonistic autoantibody-induced hypertension: Implications for immunomodulation and cytoprotection. Hypertens. Pregnancy 2017, 36, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wu, Y.; Chang, X.; Li, Y.; Wang, K.; Duan, T. Effects of Human Umbilical Cord Mesenchymal Stem Cells on Human Trophoblast Cell Functions In Vitro. Stem Cells Int. 2016, 9156731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Z.H.; Wei, J.; Lu, M.Q.; Jin, M.Y.; Geng, H.L. Protective effect of human umbilical cord mesenchymal stem cell exosomes on preserving the morphology and angiogenesis of placenta in rats with preeclampsia. Biomed. Pharmacother. 2018, 105, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Loukogeorgakis, S.P.; De Coppi, P. Concise Review: Amniotic Fluid Stem Cells: The Known, the Unknown, and Potential Regenerative Medicine Applications. Stem Cells (Dayt. Ohio) 2017, 35, 1663–1673. [Google Scholar] [CrossRef] [Green Version]
- Steigman, S.A.; Ahmed, A.; Shanti, R.M.; Tuan, R.S.; Valim, C.; Fauza, D.O. Sternal repair with bone grafts engineered from amniotic mesenchymal stem cells. J. Pediatric Surg. 2009, 44, 1120–1126. [Google Scholar] [CrossRef] [Green Version]
- Kunisaki, S.M.; Freedman, D.A.; Fauza, D.O. Fetal tracheal reconstruction with cartilaginous grafts engineered from mesenchymal amniocytes. J. Pediatric Surg. 2006, 41, 675–682. [Google Scholar] [CrossRef]
- Kunisaki, S.M.; Fuchs, J.R.; Kaviani, A.; Oh, J.T.; LaVan, D.A.; Vacanti, J.P.; Wilson, J.M.; Fauza, D.O. Diaphragmatic repair through fetal tissue engineering: A comparison between mesenchymal amniocyte- and myoblast-based constructs. J. Pediatric Surg. 2006, 41, 34–39. [Google Scholar] [CrossRef]
- Fuchs, J.R.; Kaviani, A.; Oh, J.T.; LaVan, D.; Udagawa, T.; Jennings, R.W.; Wilson, J.M.; Fauza, D.O. Diaphragmatic reconstruction with autologous tendon engineered from mesenchymal amniocytes. J. Pediatric Surg. 2004, 39, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.Y.; Liu, I.H.; Cheng, C.C.; Chang, C.C.; Lee, Y.H.; Cheng, W.T.; Wu, S.C. Amniotic fluid stem cells prevent follicle atresia and rescue fertility of mice with premature ovarian failure induced by chemotherapy. PLoS ONE 2014, 9, e106538. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Huang, Y.; Guo, L.; Cheng, W.; Zou, G. CD44+/CD105+ human amniotic fluid mesenchymal stem cells survive and proliferate in the ovary long-term in a mouse model of chemotherapy-induced premature ovarian failure. Int. J. Med Sci. 2012, 9, 592–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, G.Y.; Cheng, C.C.; Chiang, Y.S.; Cheng, W.T.; Liu, I.H.; Wu, S.C. Exosomal miR-10a derived from amniotic fluid stem cells preserves ovarian follicles after chemotherapy. Sci. Rep. 2016, 6, 23120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schickel, R.; Boyerinas, B.; Park, S.M.; Peter, M.E. MicroRNAs: Key players in the immune system, differentiation, tumorigenesis and cell death. Oncogene 2008, 27, 5959–5974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.; Ling, L.; Zhang, W.; Liu, X.; Wang, Y.; Luo, Y.; Xiong, Z. Effects of Human Amnion-Derived Mesenchymal Stem Cell (hAD-MSC) Transplantation In Situ on Primary Ovarian Insufficiency in SD Rats. Reprod. Sci. (Thousand Oakscalif.) 2020, 27, 1502–1512. [Google Scholar] [CrossRef]
- Ling, L.; Feng, X.; Wei, T.; Wang, Y.; Wang, Y.; Zhang, W.; He, L.; Wang, Z.; Zeng, Q.; Xiong, Z. Effects of low-intensity pulsed ultrasound (LIPUS)-pretreated human amnion-derived mesenchymal stem cell (hAD-MSC) transplantation on primary ovarian insufficiency in rats. Stem Cell Res. Ther. 2017, 8, 283. [Google Scholar] [CrossRef]
- Hwang, J.H.; Lee, M.J.; Seok, O.S.; Paek, Y.C.; Cho, G.J.; Seol, H.J.; Lee, J.K.; Oh, M.J. Cytokine expression in placenta-derived mesenchymal stem cells in patients with pre-eclampsia and normal pregnancies. Cytokine 2010, 49, 95–101. [Google Scholar] [CrossRef]
- Yin, N.; Zhao, W.; Luo, Q.; Yuan, W.; Luan, X.; Zhang, H. Restoring Ovarian Function With Human Placenta-Derived Mesenchymal Stem Cells in Autoimmune-Induced Premature Ovarian Failure Mice Mediated by Treg Cells and Associated Cytokines. Reprod. Sci. (Thousand Oakscalif.) 2018, 25, 1073–1082. [Google Scholar] [CrossRef]
- Zhang, H.; Luo, Q.; Lu, X.; Yin, N.; Zhou, D.; Zhang, L.; Zhao, W.; Wang, D.; Du, P.; Hou, Y. Effects of hPMSCs on granulosa cell apoptosis and AMH expression and their role in the restoration of ovary function in premature ovarian failure mice. Stem Cell Res. Ther. 2018, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Yin, N.; Wang, Y.; Lu, X.; Liu, R.; Zhang, L.; Zhao, W.; Yuan, W.; Luo, Q.; Wu, H.; Luan, X.; et al. hPMSC transplantation restoring ovarian function in premature ovarian failure mice is associated with change of Th17/Tc17 and Th17/Treg cell ratios through the PI3K/Akt signal pathway. Stem Cell Res. Ther. 2018, 9, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Zhao, W.; Wang, L.; Luo, Q.; Yin, N.; Lu, X.; Hou, Y.; Cui, J.; Zhang, H. Human placenta-derived mesenchymal stem cells inhibit apoptosis of granulosa cells induced by IRE1α pathway in autoimmune POF mice. Cell Biol. Int. 2019, 43, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Choi, J.H.; Jun, Y.; Lim, S.M.; Park, S.L.; Paek, J.Y.L.; Lee, S.H.; Hwang, J.Y.; Kim, G.J. 3D-cultured human placenta-derived mesenchymal stem cell spheroids enhance ovary function by inducing folliculogenesis. Sci. Rep. 2018, 8, 15313. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.P. Placental villous mesenchymal cells trigger trophoblast invasion. Cell Adhes. Migr. 2014, 8, 94–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolfo, A.; Giuffrida, D.; Nuzzo, A.M.; Pierobon, D.; Cardaropoli, S.; Piccoli, E.; Giovarelli, M.; Todros, T. Pro-inflammatory profile of preeclamptic placental mesenchymal stromal cells: New insights into the etiopathogenesis of preeclampsia. PLoS ONE 2013, 8, e59403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.P.; Huang, J.P.; Chu, T.Y.; Aplin, J.D.; Chen, C.Y.; Wu, Y.H. Human placental multipotent mesenchymal stromal cells modulate trophoblast migration via Rap1 activation. Placenta 2013, 34, 913–923. [Google Scholar] [CrossRef]
- Nuzzo, A.M.; Giuffrida, D.; Zenerino, C.; Piazzese, A.; Olearo, E.; Todros, T.; Rolfo, A. JunB/cyclin-D1 imbalance in placental mesenchymal stromal cells derived from preeclamptic pregnancies with fetal-placental compromise. Placenta 2014, 35, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Qu, H.M.; Qu, L.P.; Pan, X.Z.; Mu, L.S. Upregulated miR-222 targets BCL2L11 and promotes apoptosis of mesenchymal stem cells in preeclampsia patients in response to severe hypoxia. Int. J. Clin. Exp. Pathol. 2018, 11, 110–119. [Google Scholar]
- Nuzzo, A.M.; Giuffrida, D.; Masturzo, B.; Mele, P.; Piccoli, E.; Eva, C.; Todros, T.; Rolfo, A. Altered expression of G1/S phase cell cycle regulators in placental mesenchymal stromal cells derived from preeclamptic pregnancies with fetal-placental compromise. Cell Cycle (Georget. Tex.) 2017, 16, 200–212. [Google Scholar] [CrossRef]
- Lian, Q.; Zhang, Y.; Zhang, J.; Zhang, H.K.; Wu, X.; Zhang, Y.; Lam, F.F.; Kang, S.; Xia, J.C.; Lai, W.H. Functional mesenchymal stem cells derived from human induced pluripotent stem cells attenuate limb ischemia in mice. Circulation 2010, 121, 1113–1123. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Chan, Y.C.; Ho, J.C.; Siu, C.W.; Lian, Q.; Tse, H.F. Regulation of cell proliferation of human induced pluripotent stem cell-derived mesenchymal stem cells via ether-à-go-go 1 (hEAG1) potassium channel. Am. J. Physiol. Cell Physiol. 2012, 303, C115–C125. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.Y.; Lee, Y.W.; Hsu, P.J.; Wang, H.H.; Wang, Y.; Liou, J.Y.; Hsu, S.H.; Wu, K.K.; Yen, B.L. Human pluripotent stem cell (PSC)-derived mesenchymal stem cells (MSCs) show potent neurogenic capacity which is enhanced with cytoskeletal rearrangement. Oncotarget 2016, 7, 43949–43959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloor, A.; Patel, A.; Griffin, J.E.; Gilleece, M.H.; Radia, R.; Yeung, D.T.; Drier, D.; Larson, L.S.; Uenishi, G.I.; Hei, D.; et al. Production, safety and efficacy of iPSC-derived mesenchymal stromal cells in acute steroid-resistant graft versus host disease: A phase I, multicenter, open-label, dose-escalation study. Nat. Med. 2020. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| MSC Types | Disease | Model | Main Effect | References |
|---|---|---|---|---|
| BMSC | POF | Rabbits | Increasing the secretion of VEGF | [46] |
| POF | Mice | Formation of new primordial follicles | [49] | |
| POF | Rat | Increasing ovarian weight, follicle counts and E2 levels | [48] | |
| POF | Rat | Inhibition of GCs apoptosis | [61] | |
| POF | Rat | Increasing follicle counts, E2 and AMH levels | [62] | |
| Asherman syndrome | Human | Restoration of menstruation in endometrium | [53] | |
| Asherman syndrome | Rat | Reconstruction of functional endometrium | [41] | |
| Asherman syndrome | Murine | Regeneration of endometrium | [56] | |
| Asherman syndrome | Human | Reconstruction of functional endometrium | [57] | |
| ADSC | POF | Mice | Increasing the number of follicles with normal structure | [71] |
| POF | Mice | Increasing follicles at different stages and ovulation | [72] | |
| POF | Rat | Increasing follicle counts, E2 levels and pregnancy rates | [73] | |
| Endometriosis | Cell culture | Supporting the development of endometriosis | [74] | |
| Asherman syndrome | Rat | Regeneration of endometrium | [66] | |
| MenSC | POF | Mice | Increasing ovarian weight, follicle counts and E2 levels | [80] |
| POF | Rat | Increasing AMH, E2 and progesterone levels | [81] | |
| POF | Mice | Inhibition of GCs apoptosis | [77] | |
| Endometriosis | Mice | Increasing the invasion and angiogenesis | [86] | |
| Asherman syndrome | Human | Regeneration of endometrium | [91] | |
| Asherman syndrome | Rat | Increasing the secretion of angiogenic and anti-inflammatory factors | [92] | |
| UC-MSC | POF | Murine | Increasing ovarian weight, follicle counts, AMH levels, expression of Inhibin A and FSHR in growing follicles | [97] |
| POF | Rat | Inhibition of GCs apoptosis, improvement of endocrine secretion system and folliculogenesis, | [98] | |
| POF | Rat | Recovery of estrous cycle, levels of sex hormones and fertility | [99] | |
| POF | Mice | Inhibition of GCs apoptosis, increasing the level of sex hormons and number of follicles | [100] | |
| POF | Human | Activation of primordial follicles | [103] | |
| POF | Mice | Increasing ovarian volume and angiogenesis, number of antral follicles, AMH and E2 levels | [104] | |
| POF | Rat | Increasing follicle counts and E2 levels | [102] | |
| PCOS | Mice | Inhibition of ovarian local and systemic inflammatory responses | [36] | |
| Preeclampsia | Rat | Inhibition of inflammation | [113] | |
| Preeclampsia | Rat | Inhibition of inflammation | [114] | |
| Preeclampsia | Rat | Inhibition of hypertension and inflammation | [115] | |
| Preeclampsia | Rat | Improvement of morphology and angiogenesis of placenta | [117] | |
| AFSC | POF | Mice | Inhibition of follicular atresia and preserving the healthy follicles | [123] |
| POF | Mice | Regeneration of ovarian cells | [124] | |
| POF | Mice | Inhibition of GCs apoptosis and follicular atresia | [125] | |
| AD-MSC | POF | Rat | Inhibition of GCs apoptosis, increasing ovarian angiogenesis and follicular development | [34] |
| POF | Rat | Inhibition of GCs apoptosis, increasing follicular numbers and AMH levels | [127] | |
| POF | Rat | Inhibition of inflammation | [128] | |
| PMSC | POF | Mice | Inhibition of GCs apoptosis, increasing E2 levels | [130] |
| POF | Mice | Inhibition of GCs apoptosis, improvement of ovarian function | [131] | |
| POF | Mice | Increasing follicular numbers, E2 and AMH levels | [132] | |
| POF | Mice | Inhibition of GCs apoptosis | [133] | |
| Preeclampsia | Human | Increasing the production of HGF | [137] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esfandyari, S.; Chugh, R.M.; Park, H.-s.; Hobeika, E.; Ulin, M.; Al-Hendy, A. Mesenchymal Stem Cells as a Bio Organ for Treatment of Female Infertility. Cells 2020, 9, 2253. https://doi.org/10.3390/cells9102253
Esfandyari S, Chugh RM, Park H-s, Hobeika E, Ulin M, Al-Hendy A. Mesenchymal Stem Cells as a Bio Organ for Treatment of Female Infertility. Cells. 2020; 9(10):2253. https://doi.org/10.3390/cells9102253
Chicago/Turabian StyleEsfandyari, Sahar, Rishi Man Chugh, Hang-soo Park, Elie Hobeika, Mara Ulin, and Ayman Al-Hendy. 2020. "Mesenchymal Stem Cells as a Bio Organ for Treatment of Female Infertility" Cells 9, no. 10: 2253. https://doi.org/10.3390/cells9102253
APA StyleEsfandyari, S., Chugh, R. M., Park, H. -s., Hobeika, E., Ulin, M., & Al-Hendy, A. (2020). Mesenchymal Stem Cells as a Bio Organ for Treatment of Female Infertility. Cells, 9(10), 2253. https://doi.org/10.3390/cells9102253