Visfatin: A Possible Role in Cardiovasculo-Metabolic Disorders

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Visfatin’s Tissue Expression
3. Functional Roles
3.1. Intracellular Visfatin (iNAMPT)
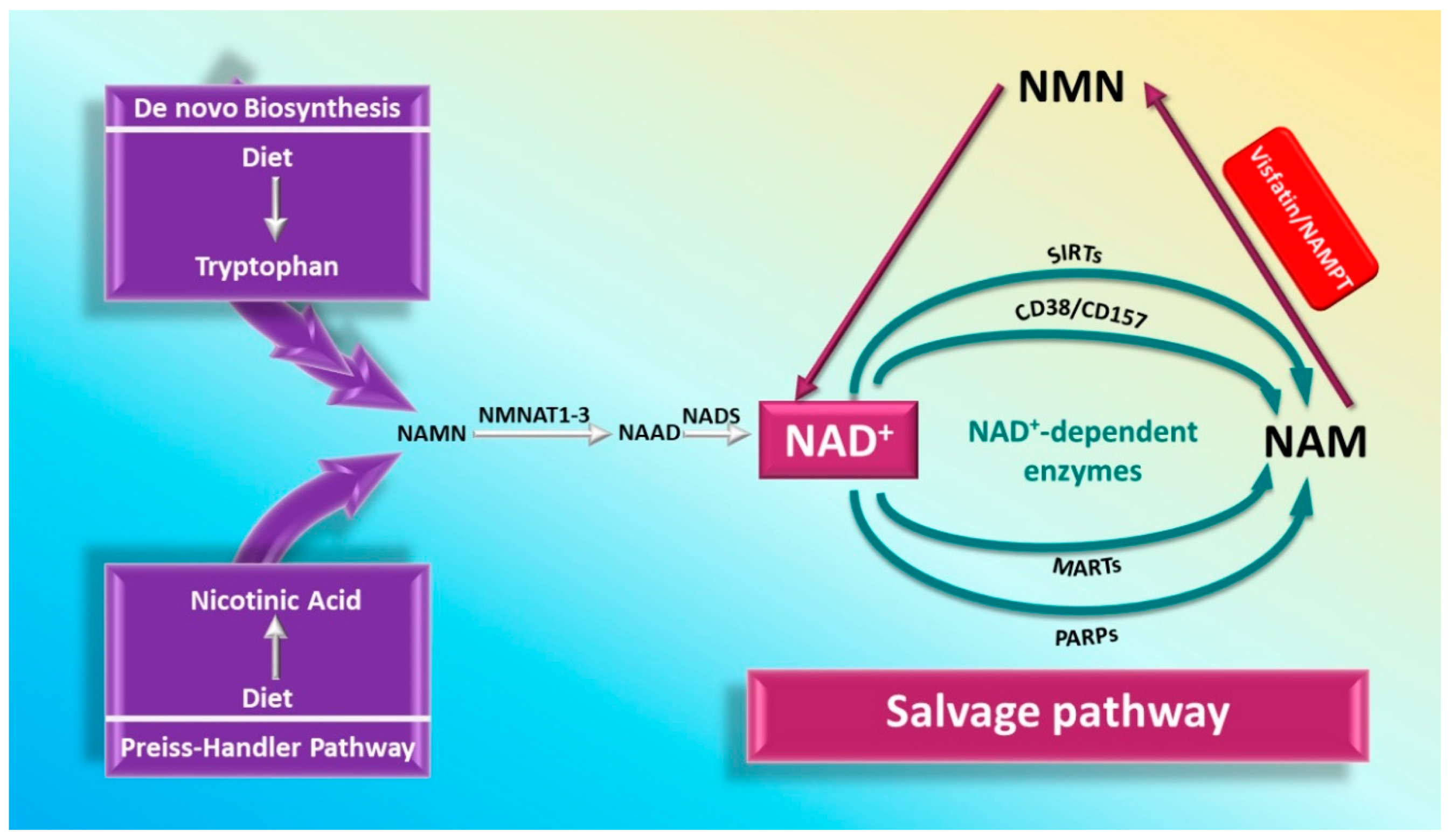
3.1.1. Role of iNAMPT in NAD Biosynthesis
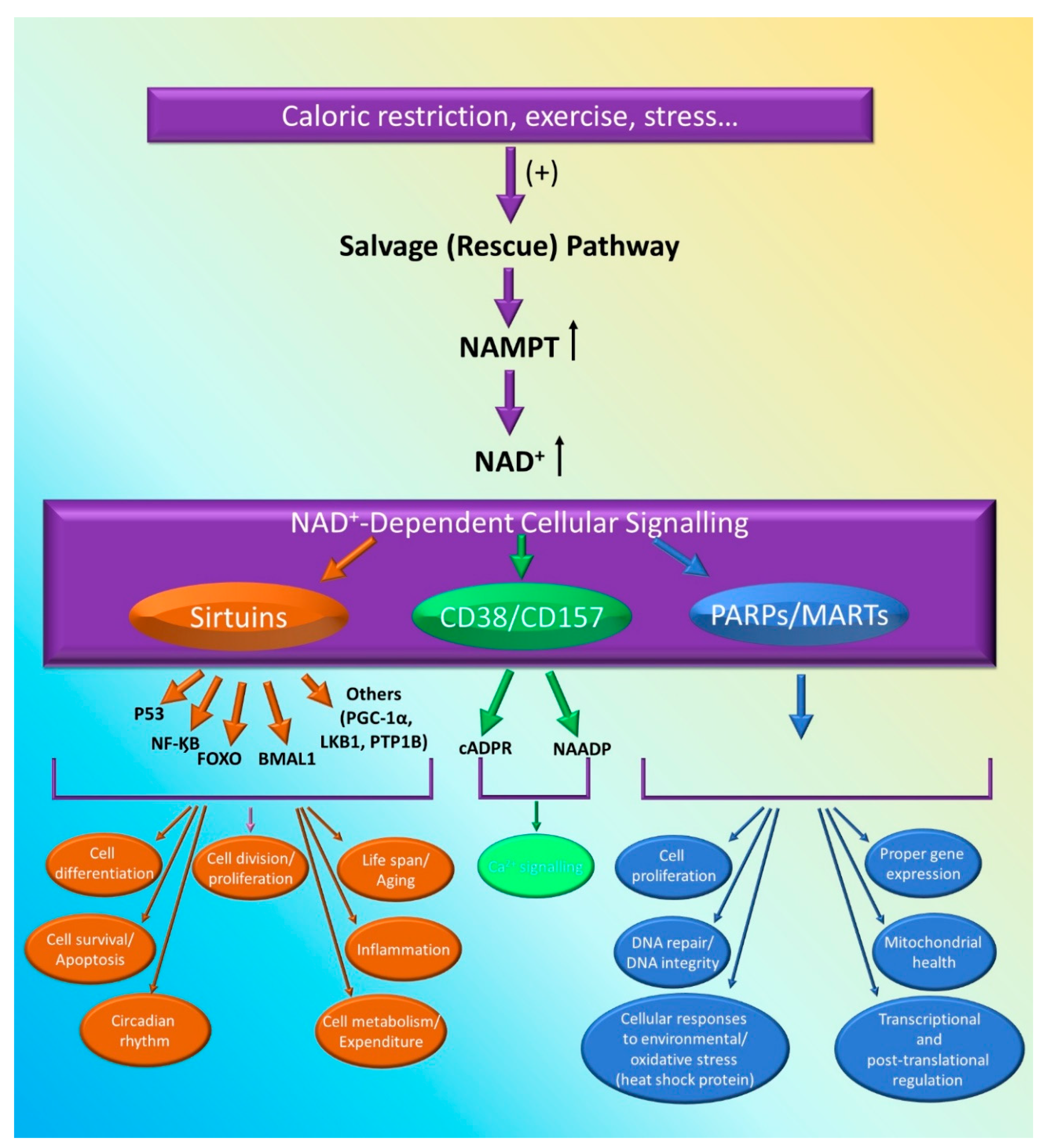
3.1.2. Visfatin and NAD-Dependent Enzymes
Sirtuins
PARPs/MARTs
CD38 and CD157
3.2. Extracellular Visfatin (eNAMPT)
3.2.1. eNAMPT/Visfatin Acting as PBEF
3.2.2. eNAMPT/Visfatin Acting as a Cytokine
3.2.3. eNAMPT/Visfatin Acting as Insulinomimetic Adipokine
4. Relevance of Visfatin to Metabolic Diseases
4.1. Visfatin and Diabetes
4.2. Visfatin and Obesity
4.3. Visfatin and PCOS
4.4. Visfatin in Clinical Studies
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Murphy, K.; Bloom, S.R. Are all fats created equal? Nat. Med. 2006, 12, 32–33. [Google Scholar] [CrossRef]
- Hassan, M.; Latif, N.; Yacoub, M. Adipose tissue: Friend or foe? Nat. Rev. Cardiol. 2012, 9, 689–702. [Google Scholar] [CrossRef] [PubMed]
- Fain, J.N.; Madan, A.K.; Hiler, M.L.; Cheema, P.; Bahouth, S.W. Comparison of the Release of Adipokines by Adipose Tissue, Adipose Tissue Matrix, and Adipocytes from Visceral and Subcutaneous Abdominal Adipose Tissues of Obese Humans. Endocrinology 2004, 145, 2273–2282. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.J.; Wu, C.; Lin, H.; Lee, I.; Tseng, J.; Chou, M.M.; Sheu, W.H.H. Visfatin-induced expression of inflammatory mediators in human endothelial cells through the NF-κB pathway. Int. J. Obes. 2009, 33, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, S.H.; Li, Y.; Parodo, J.; Kapus, A.; Fan, L.; Rotstein, O.D.; Marshall, J.C. Pre–B cell colony–enhancing factor inhibits neutrophil apoptosis in experimental inflammation and clinical sepsis. J. Clin. Investig. 2004, 113, 318–1327. [Google Scholar] [CrossRef]
- Belo, V.A.; Luizon, M.R.; Lacchini, R.; Miranda, J.A.; Lanna, C.M.M.; Souza-Costa, D.C.; Tanus-Santos, J.E. The effects of NAMPT haplotypes and metabolic risk factors on circulating visfatin/NAMPT levels in childhood obesity. Int. J. Obes. 2013, 39, 130–135. [Google Scholar] [CrossRef]
- Dogru, T.; Sonmez, A.; Tasci, I.; Yilmaz, M.; Erdem, G.; Erturk, H.; Bingol, N.; Kilic, S.; Ozgurtas, T. Plasma visfatin levels in young male patients with uncomplicated and newly diagnosed hypertension. J. Hum. Hypertens. 2006, 21, 173–175. [Google Scholar] [CrossRef] [PubMed]
- Gunes, F.; Akbal, E.; Cakir, E.; Akyurek, O.; Altunbas, M.; Ozbek, M. Visfatin may be a novel marker for identifying stages of essential hypertension in advanced age patients. Intern. Med. 2012, 51, 553–557. [Google Scholar] [CrossRef] [Green Version]
- Dahl, T.B.; Yndestad, A.; Skjelland, M.; Øie, E.; Dahl, A.; Michelsen, A.; Damås, J.K.; Tunheim, S.H.; Ueland, T.; Smith, C.; et al. Increased Expression of Visfatin in Macrophages of Human Unstable Carotid and Coronary Atherosclerosis. Circulation 2007, 115, 972–980. [Google Scholar] [CrossRef]
- Anwar, M.; Saleh, A.I.; Al Olabi, R.; Al Shehabi, T.S.; Eid, A.H. Glucocorticoid-induced fetal origins of adult hypertension: Association with epigenetic events. Vasc. Pharmacol. 2016, 82, 41–50. [Google Scholar] [CrossRef]
- Samal, B.; Sun, Y.; Stearns, G.; Xie, C.; Suggs, S.; Mcniece, I. Cloning and characterization of the cDNA encoding a novel human pre-B-cell colony-enhancing factor. Mol. Cell. Biol. 1994, 14, 1431–1437. [Google Scholar] [CrossRef]
- Martin, P.R.; Shea, R.J.; Mulks, M.H. Identification of a Plasmid-Encoded Gene fromHaemophilus ducreyi Which Confers NAD Independence. J. Bacteriol. 2001, 183, 1168–1174. [Google Scholar] [CrossRef] [Green Version]
- Rongvaux, A.; Shea, R.J.; Mulks, M.H.; Gigot, D.; Urbain, J.; Leo, O.; Andris, F. Pre-B-cell colony-enhancing factor, whose expression is up-regulated in activated lymphocytes, is a nicotinamide phosphoribosyltransferase, a cytosolic enzyme involved in NAD biosynthesis. Eur. J. Immunol. 2002, 32, 3225–3234. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, X.; Bheda, P.; Revollo, J.R.; Imai, S.-I.; Wolberger, C. Structure of Nampt/PBEF/visfatin, a mammalian NAD+ biosynthetic enzyme. Nat. Struct. Mol. Biol. 2006, 13, 661–662. [Google Scholar] [CrossRef]
- Fukuhara, A.; Matsuda, M.; Nishizawa, M.; Segawa, K.; Tanaka, M.; Kishimoto, K.; Matsuki, Y.; Murakami, M.; Ichisaka, T.; Murakami, H.; et al. Visfatin: A Protein Secreted by Visceral Fat That Mimics the Effects of Insulin. Science 2005, 307, 426–430. [Google Scholar] [CrossRef] [PubMed]
- Revollo, J.R.; Körner, A.; Mills, K.F.; Satoh, A.; Wang, T.; Garten, A.; Dasgupta, B.; Sasaki, Y.; Wolberger, C.; Townsend, R.R.; et al. Nampt/PBEF/Visfatin Regulates Insulin Secretion in β Cells as a Systemic NAD Biosynthetic Enzyme. Cell Metab. 2007, 6, 363–375. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Xia, T.; Zhou, L.; Chen, X.; Gan, L.; Yao, W.; Peng, Y.; Yang, Z. Gene organization, alternate splicing and expression pattern of porcine visfatin gene. Domest. Anim. Endocrinol. 2007, 32, 235–245. [Google Scholar] [CrossRef]
- Chang, L.; Yang, R.; Wang, M.; Liu, J.; Wang, Y.; Zhang, H.; Li, Y. Angiotensin II Type-1 Receptor-JAK/STAT Pathway Mediates the Induction of Visfatin in Angiotensin II-Induced Cardiomyocyte Hypertrophy. Am. J. Med Sci. 2012, 343, 220–226. [Google Scholar] [CrossRef]
- Chen, J.; Weng, J.-F.; Hong, W.-C.; Luo, L.-F.; Yu, W.; Luo, S.-D. Change in plasma visfatin level after severe traumatic brain injury. Peptides 2012, 38, 8–12. [Google Scholar] [CrossRef]
- Xiao, H.; Cheng, M.; Zhang, L.-J.; Liu, K. Visfatin expression and genetic polymorphism in patients with traumatic brain injury. Int. J. Clin. Exp. Med. 2015, 8, 9799–9804. [Google Scholar]
- Dahl, T.B.; Holm, S.; Aukrust, P.; Halvorsen, B. Visfatin/NAMPT: A Multifaceted Molecule with Diverse Roles in Physiology and Pathophysiology. Annu. Rev. Nutr. 2012, 32, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Verdin, E. NAD+ in aging, metabolism, and neurodegeneration. Science 2015, 350, 1208–1213. [Google Scholar] [CrossRef]
- Belenky, P.; Bogan, K.L.; Brenner, C. NAD+ metabolism in health and disease. Trends Biochem. Sci. 2007, 32, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Menzies, K.J.; Auwerx, J. NAD+ Metabolism and the Control of Energy Homeostasis: A Balancing Act between Mitochondria and the Nucleus. Cell Metab. 2015, 22, 31–53. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Sauve, A.A. NAD + metabolism: Bioenergetics, signaling and manipulation for therapy. Biochim. Biophys. Acta 2016, 1864, 1787–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agledal, L.; Niere, M.; Ziegler, M. The phosphate makes a difference: Cellular functions of NADP. Redox Rep. 2010, 15, 2–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, W. NAD+/NADH and NADP+/NADPH in Cellular Functions and Cell Death: Regulation and Biological Consequences. Antioxid. Redox. Signal. 2007, 10, 179–206. [Google Scholar] [CrossRef] [Green Version]
- Bogan, K.L.; Brenner, C. Nicotinic Acid, Nicotinamide, and Nicotinamide Riboside: A Molecular Evaluation of NAD+Precursor Vitamins in Human Nutrition. Annu. Rev. Nutr. 2008, 28, 115–130. [Google Scholar] [CrossRef] [Green Version]
- Hassa, P.O.; Haenni, S.S.; Elser, M.; Hottiger, M.O. Nuclear ADP-Ribosylation Reactions in Mammalian Cells: Where Are We Today and Where Are We Going? Microbiol. Mol. Biol. Rev. 2006, 70, 789–829. [Google Scholar] [CrossRef] [Green Version]
- Chowdhry, S.; Zanca, C.; Rajkumar, U.; Koga, T.; Diao, Y.; Raviram, R.; Liu, F.; Turner, K.; Yang, H.; Brunk, E.; et al. NAD metabolic dependency in cancer is shaped by gene amplification and enhancer remodelling. Nat. Cell Biol. 2019, 569, 570–575. [Google Scholar] [CrossRef]
- Houtkooper, R.H.; Cantó, C.; Wanders, R.J.; Auwerx, J. The Secret Life of NAD+: An Old Metabolite Controlling New Metabolic Signaling Pathways. Endocr. Rev. 2010, 31, 194–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Miao, C.-Y. NAMPT as a Therapeutic Target against Stroke. Trends Pharmacol. Sci. 2015, 36, 891–905. [Google Scholar] [CrossRef]
- Imai, S.-I.; Guarente, L. NAD+ and sirtuins in aging and disease. Trends Cell Biol. 2014, 24, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Garten, A.; Schuster, S.; Penke, M.; Gorski, T.; De Giorgis, T.; Kiess, W. Physiological and pathophysiological roles of NAMPT and NAD metabolism. Nat. Rev. Endocrinol. 2015, 11, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Houtkooper, R.H.; Pirinen, E.; Auwerx, J. Sirtuins as regulators of metabolism and healthspan. Nat. Rev. Mol. Cell Biol. 2012, 13, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Michan, S.; Sinclair, D.A. Sirtuins in mammals: Insights into their biological function. Biochem. J. 2007, 404, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kane, A.E.; Sinclair, D.A. Sirtuins and NAD+ in the Development and Treatment of Metabolic and Cardiovascular Diseases. Circ. Res. 2018, 123, 868–885. [Google Scholar] [CrossRef]
- Rutanen, J.; Yaluri, N.; Modi, S.; Pihlajamäki, J.; Vänttinen, M.; Itkonen, P.; Kainulainen, S.; Yamamoto, H.; Lagouge, M.; Sinclair, D.A.; et al. SIRT1 mRNA Expression May Be Associated with Energy Expenditure and Insulin Sensitivity. Diabetes 2010, 59, 829–835. [Google Scholar] [CrossRef] [Green Version]
- Nakahata, Y.; Sahar, S.; Astarita, G.; Kaluzova, M.; Sassone-Corsi, P. Circadian Control of the NAD+ Salvage Pathway by CLOCK-SIRT1. Science 2009, 324, 654–657. [Google Scholar] [CrossRef]
- Lüscher, B.; Bütepage, M.; Eckei, L.; Krieg, S.; Verheugd, P.; Shilton, B.H. ADP-Ribosylation, a Multifaceted Posttranslational Modification Involved in the Control of Cell Physiology in Health and Disease. Chem. Rev. 2018, 118, 1092–1136. [Google Scholar] [CrossRef]
- Vyas, S.; Matic, I.; Uchima, L.; Rood, J.; Zaja, R.; Hay, R.T.; Ahel, I.; Chang, P. Family-wide analysis of poly(ADP-ribose) polymerase activity. Nat. Commun. 2014, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- D’Amours, D.; Desnoyers, S.; D’Silva, I.; Poirier, G.G. Poly(ADP-ribosyl)ation reactions in the regulation of nuclear functions. Biochem. J. 1999, 342, 249–268. [Google Scholar]
- Amé, J.-C.; Spenlehauer, C.; De Murcia, G. The PARP superfamily. BioEssays 2004, 26, 882–893. [Google Scholar] [CrossRef]
- Schreiber, V.; Dantzer, F.; Ame, J.-C.; De Murcia, G. Poly(ADP-ribose): Novel functions for an old molecule. Nat. Rev. Mol. Cell Biol. 2006, 7, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Vyas, S.; Chesarone-Cataldo, M.; Todorova, T.; Huang, Y.-H.; Chang, P. A systematic analysis of the PARP protein family identifies new functions critical for cell physiology. Nat. Commun. 2013, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Pittelli, M.; Formentini, L.; Faraco, G.; Lapucci, A.; Rapizzi, E.; Cialdai, F.; Romano, G.; Moneti, G.; Moroni, F.; Chiarugi, A. Inhibition of Nicotinamide Phosphoribosyltransferase. J. Biol. Chem. 2010, 285, 34106–34114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rongvaux, A.; Galli, M.; Denanglaire, S.; Van Gool, F.; Drèze, P.L.; Szpirer, C.; Bureau, F.; Andris, F.; Leo, O. Nicotinamide phosphoribosyl transferase/pre-B cell colony-enhancing factor/visfatin is required for lymphocyte development and cellular resistance to genotoxic stress. J. Immunol. 2008, 181, 4685–4695. [Google Scholar] [CrossRef] [Green Version]
- Dahl, T.B.; Haukeland, J.W.; Yndestad, A.; Ranheim, T.; Gladhaug, I.P.; Damås, J.K.; Haaland, T.; Løberg, E.M.; Arntsen, B.; Birkeland, K.; et al. Intracellular Nicotinamide Phosphoribosyltransferase Protects against Hepatocyte Apoptosis and Is Down-Regulated in Nonalcoholic Fatty Liver Disease. J. Clin. Endocrinol. Metab. 2010, 95, 3039–3047. [Google Scholar] [CrossRef]
- Xu, R.; Yuan, Z.; Yang, L.; Li, L.; Li, D.; Lv, C. Inhibition of NAMPT decreases cell growth and enhances susceptibility to oxidative stress. Oncol. Rep. 2017, 38, 1767–1773. [Google Scholar] [CrossRef] [Green Version]
- Alessio, M.; Roggero, S.; Funaro, A.; De Monte, L.B.; Peruzzi, L.; Geuna, M.; Malavasi, F. CD38 molecule: Structural and biochemical analysis on human T lymphocytes, thymocytes, and plasma cells. J. Immunol. 1990, 145, 878–884. [Google Scholar] [PubMed]
- Malavasi, F.; Funaro, A.; Alessio, M.; Demonte, L.B.; Ausiello, C.M.; Dianzani, U.; Lanza, F.; Magrini, E.; Momo, M.; Roggero, S. CD38: A multi-lineage cell activation molecule with a split personality. Int. J. Clin. Lab. Res. 1992, 22, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Mehta, K.; Shahid, U.; Malavasi, F. Human CD38, a cell-surface protein with multiple functions. FASEB J. 1996, 10, 1408–1417. [Google Scholar] [CrossRef]
- Fernàndez, J.E.; Deaglio, S.; Donati, D.; Beusan, I.S.; Corno, F.; Aranega, A.; Forni, M.; Falini, B.; Malavasi, F. Analysis of the distribution of human CD38 and of its ligand CD31 in normal tissues. J. Boil. Regul. Homeost. Agents 1998, 12, 81–91. [Google Scholar]
- Takasawa, S.; Tohgo, A.; Noguchi, N.; Koguma, T.; Nata, K.; Sugimoto, T.; Yonekura, H.; Okamoto, H. Synthesis and hydrolysis of cyclic ADP-ribose by human leukocyte antigen CD38 and inhibition of the hydrolysis by ATP. J. Biol. Chem. 1993, 268, 26052–26054. [Google Scholar]
- Aksoy, P.; White, T.A.; Thompson, M.; Chini, E.N. Regulation of intracellular levels of NAD: A novel role for CD38. Biochem. Biophys. Res. Commun. 2006, 345, 1386–1392. [Google Scholar] [CrossRef]
- Chini, E.N.; Chini, C.C.S.; Kato, I.; Takasawa, S.; Okamoto, H. CD38 is the major enzyme responsible for synthesis of nicotinic acid-adenine dinucleotide phosphate in mammalian tissues. Biochem. J. 2002, 362, 125–130. [Google Scholar]
- Dousa, T.P.; Chini, E.N.; Beers, K.W. Adenine nucleotide diphosphates: Emerging second messengers acting via intracellular Ca2+ release. Am. J. Physiol. Physiol. 1996, 271, C1007–C1024. [Google Scholar] [CrossRef]
- Howard, M.; Grimaldi, J.C.; Bazan, J.F.; Lund, F.E.; Santos-Argumedo, L.; Parkhouse, R.M.; Walseth, T.F.; Lee, H.C. Formation and hydrolysis of cyclic ADP-ribose catalyzed by lymphocyte antigen CD38. Science 1993, 262, 1056–1059. [Google Scholar] [CrossRef]
- Lee, H.C.; Aarhus, R. A Derivative of NADP Mobilizes Calcium Stores Insensitive to Inositol Trisphosphate and Cyclic ADP-ribose. J. Biol. Chem. 1995, 270, 2152–2157. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.C.; Aarhus, R.; Levitt, D. The crystal structure of cyclic ADP-ribose. Nat. Genet. 1994, 1, 143–144. [Google Scholar] [CrossRef]
- Lee, H.C.; Walseth, T.F.; Bratt, G.T.; Hayes, R.N.; Clapper, D.L. Structural determination of a cyclic metabolite of NAD+ with intracellular Ca2+-mobilizing activity. J. Biol. Chem. 1989, 264, 1608–1615. [Google Scholar]
- Liang, M.; Chini, E.N.; Cheng, J.; Dousa, T.P. Synthesis of NAADP and cADPR in Mitochondria. Arch. Biochem. Biophys. 1999, 371, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Khoo, K.M.; Han, M.-K.; Park, J.B.; Chae, S.W.; Kim, U.H.; Lee, H.C.; Bay, B.H.; Chang, C.F. Localization of the Cyclic ADP-ribose-dependent Calcium Signaling Pathway in Hepatocyte Nucleus. J. Biol. Chem. 2000, 275, 24807–24817. [Google Scholar] [CrossRef] [Green Version]
- Adebanjo, O.A.; Anandatheerthavarada, H.K.; Koval, A.P.; Moonga, B.S.; Biswas, G.; Sun, L.; Sodam, B.R.; Bevis, P.J.; Huang, C.L.; Epstein, S.; et al. A new function for CD38/ADP-ribosyl cyclase in nuclear Ca2+ homeostasis. Nat. Cell Biol. 1999, 1, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Mizuguchi, M.; Otsuka, N.; Ikeda, K.; Takahashi, H. Ultrastructural localization of CD38 immunoreactivity in rat brain. Brain Res. 1997, 756, 52–60. [Google Scholar] [CrossRef]
- Yalcintepe, L.; Albeniz, I.; Cinar, S.; Tiryaki, D.; Bermek, E.; Graeff, R.M.; Lee, H.C. Nuclear CD38 in retinoic acid-induced HL-60 cells. Exp. Cell Res. 2005, 303, 14–21. [Google Scholar] [CrossRef]
- Sun, L.; Adebanjo, O.A.; Koval, A.; Anandatheerthavarada, H.K.; Iqbal, J.; Wu, X.Y.; Moonga, B.S.; Wu, X.B.; Biswas, G.; Bevis, P.J.; et al. A novel mechanism for coupling cellular intermediary metabolism to cytosolic Ca2+ signaling via CD38/ADP-ribosyl cyclase, a putative intracellular NAD+ sensor. FASEB J. 2002, 16, 302–314. [Google Scholar] [CrossRef] [Green Version]
- Orciani, M.; Trubiani, O.; Guarnieri, S.; Ferrero, E.; Di Primio, R. CD38 is constitutively expressed in the nucleus of human hematopoietic cells. J. Cell. Biochem. 2008, 105, 905–912. [Google Scholar] [CrossRef]
- Chmielewski, J.P.; Bowlby, S.C.; Wheeler, F.B.; Shi, L.; Sui, G.; Davis, A.L.; Howard, T.D.; Agostino, R.B.; Miller, L.D.; Sirintrapun, S.J.; et al. CD38 Inhibits Prostate Cancer Metabolism and Proliferation by Reducing Cellular NAD+ Pools. Mol. Cancer Res. 2018, 16, 1687. [Google Scholar] [CrossRef] [Green Version]
- Quarona, V.; Zaccarello, G.; Chillemi, A.; Brunetti, E.; Singh, V.K.; Ferrero, E.; Funaro, A.; Horenstein, A.L.; Malavasi, F. CD38 and CD157: A long journey from activation markers to multifunctional molecules. Cytom. Part. B Clin. Cytom. 2013, 84, 207–217. [Google Scholar] [CrossRef]
- Moschen, A.R.; Kaser, A.; Enrich, B.; Mosheimer, B.; Theurl, M.; Niederegger, H.; Tilg, H. Visfatin, an Adipocytokine with Proinflammatory and Immunomodulating Properties. J. Immunol. 2007, 178, 1748–1758. [Google Scholar] [CrossRef] [Green Version]
- Hirata, Y.; Kimura, N.; Sato, K.; Ohsugi, Y.; Takasawa, S.; Okamoto, H.; Ishikawa, J.; Kaisho, T.; Ishihara, K.; Hirano, T. ADP ribosyl cyclase activity of a novel bone marrow stromal cell surface molecule, BST-1. FEBS Lett. 1994, 356, 244–248. [Google Scholar] [CrossRef] [Green Version]
- Kover, K.; Tong, P.Y.; Watkins, D.; Clements, M.; Stehno-Bittel, L.; Novikova, L.; Bittel, D.; Kibiryeva, N.; Stuhlsatz, J.; Yan, Y.; et al. Expression and Regulation of Nampt in Human Islets. PLoS ONE 2013, 8, e58767. [Google Scholar] [CrossRef]
- Stephens, J.M.; Vidal-Puig, A.J. An update on visfatin/pre-B cell colony-enhancing factor, an ubiquitously expressed, illusive cytokine that is regulated in obesity. Curr. Opin. Lipidol. 2006, 17, 128–131. [Google Scholar] [CrossRef]
- Hug, C.; Lodish, H.F. MEDICINE: Visfatin: A New Adipokine. Science 2005, 307, 366–367. [Google Scholar] [CrossRef]
- De Fougerolles, A.R.; Chi-Rosso, G.; Bajardi, A.; Gotwals, P.; Green, C.D.; Koteliansky, V.E. Global Expression Analysis of Extracellular Matrix–Integrin Interactions in Monocytes. Immunity 2000, 13, 749–758. [Google Scholar] [CrossRef] [Green Version]
- Newburger, P.E.; Subrahmanyam, Y.; Weissman, S.M. Global analysis of neutrophil gene expression. Curr. Opin. Hematol. 2000, 7, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Nau, G.J.; Richmond, J.F.L.; Schlesinger, A.; Jennings, E.G.; Lander, E.S.; Young, R.A. Human macrophage activation programs induced by bacterial pathogens. Proc. Natl. Acad. Sci. USA 2002, 99, 1503–1508. [Google Scholar] [CrossRef] [Green Version]
- Adya, R.; Tan, B.K.; Chen, J.; Randeva, H.S. Pre-B cell colony enhancing factor (PBEF)/visfatin induces secretion of MCP-1 in human endothelial cells: Role in visfatin-induced angiogenesis. Atherosclerosis 2009, 205, 113–119. [Google Scholar] [CrossRef]
- Adya, R.; Tan, B.K.; Chen, J.; Randeva, H.S. Nuclear Factor-κB Induction by Visfatin in Human Vascular Endothelial Cells. Its role in MMP-2/9 production and activation. Diabetes Care 2008, 31, 758–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-R.; Bae, Y.-H.; Bae, S.-K.; Choi, K.-S.; Yoon, K.-H.; Koo, T.H.; Jang, H.-O.; Yun, I.; Kim, K.-W.; Kwon, Y.-G.; et al. Visfatin enhances ICAM-1 and VCAM-1 expression through ROS-dependent NF-κB activation in endothelial cells. Biochim. Biophys. Acta (BBA) Bioenerg. 2008, 1783, 886–895. [Google Scholar] [CrossRef] [Green Version]
- Adya, R.; Tan, B.K.; Punn, A.; Chen, J.; Randeva, H.S. Visfatin induces human endothelial VEGF and MMP-2/9 production via MAPK and PI3K/Akt signalling pathways: Novel insights into visfatin-induced angiogenesis. Cardiovasc. Res. 2007, 78, 356–365. [Google Scholar] [CrossRef]
- Bae, Y.-H.; Bae, M.-K.; Kim, S.-R.; Lee, J.H.; Wee, H.-J.; Bae, S.-K. Upregulation of fibroblast growth factor-2 by visfatin that promotes endothelial angiogenesis. Biochem. Biophys. Res. Commun. 2009, 379, 206–211. [Google Scholar] [CrossRef]
- Haider, D.G.; Schaller, G.; Kapiotis, S.; Maier, C.; Luger, A.; Wolzt, M. The release of the adipocytokine visfatin is regulated by glucose and insulin. Diabetologia 2006, 49, 1909–1914. [Google Scholar] [CrossRef]
- Fukuhara, A.; Matsuda, M.; Nishizawa, M.; Segawa, K.; Tanaka, M.; Kishimoto, K.; Matsuki, Y.; Murakami, M.; Ichisaka, T.; Murakami, H.; et al. Retraction. Science 2007, 318, 565. [Google Scholar] [CrossRef]
- Harasim-Symbor, E.; Chabowski, A.; Gorski, J. Lack of downstream insulin-mimetic effects of visfatin/eNAMPT on glucose and fatty acid metabolism in skeletal muscles. Acta Physiol. 2011, 202, 21–28. [Google Scholar] [CrossRef]
- Chang, Y.-H.; Chang, D.-M.; Lin, K.-C.; Shin, S.-J.; Lee, Y.-J. Visfatin in overweight/obesity, type 2 diabetes mellitus, insulin resistance, metabolic syndrome and cardiovascular diseases: A meta-analysis and systemic review. Diabetes/Metabolism Res. Rev. 2011, 27, 515–527. [Google Scholar] [CrossRef]
- Krzyzanowska, K.; Krugluger, W.; Mittermayer, F.; Rahman, R.; Haider, M.; Shnawa, N.; Schernthaner, G. Increased visfatin concentrations in women with gestational diabetes mellitus. Clin. Sci. 2006, 110, 605–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Bermejo, A.; Chico-Julià, B.; Fernàndez-Balsells, M.; Recasens, M.; Esteve, E.; Casamitjana, R.; Ricart, W.; Fernández-Real, J.-M. Serum Visfatin Increases with Progressive β-Cell Deterioration. Diabetes 2006, 55, 2871–2875. [Google Scholar] [CrossRef] [Green Version]
- Haider, D.G.; Pleiner, J.; Francesconi, M.; Wiesinger, G.F.; Müller, M.; Wolzt, M. Exercise Training Lowers Plasma Visfatin Concentrations in Patients with Type 1 Diabetes. J. Clin. Endocrinol. Metab. 2006, 91, 4702–4704. [Google Scholar] [CrossRef] [Green Version]
- Retnakaran, R.; Youn, B.-S.; Liu, Y.; Hanley, A.J.G.; Lee, N.S.; Park, J.W.; Song, E.S.; Vu, V.; Kim, W.; Tungtrongchitr, R.; et al. Correlation of circulating full-length visfatin (PBEF/NAMPT) with metabolic parameters in subjects with and without diabetes: A cross-sectional study. Clin. Endocrinol. 2008, 69, 885–893. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.-P.; Chung, F.-M.; Chang, D.-M.; Tsai, J.C.-R.; Huang, H.-F.; Shin, S.-J.; Lee, Y.-J. Elevated Plasma Level of Visfatin/Pre-B Cell Colony-Enhancing Factor in Patients with Type 2 Diabetes Mellitus. J. Clin. Endocrinol. Metab. 2006, 91, 295–299. [Google Scholar] [CrossRef] [Green Version]
- Sandeep, S.; Velmurugan, K.; Deepa, R.; Mohan, V. Serum visfatin in relation to visceral fat, obesity, and type 2 diabetes mellitus in Asian Indians. Metabolism 2007, 56, 565–570. [Google Scholar] [CrossRef]
- Dogru, T.; Sonmez, A.; Tasci, I.; Bozoglu, E.; Yilmaz, M.I.; Genc, H.; Erdem, G.; Gok, M.; Bingol, N.; Kiliç, S.; et al. Plasma visfatin levels in patients with newly diagnosed and untreated type 2 diabetes mellitus and impaired glucose tolerance. Diabetes Res. Clin. Pract. 2007, 76, 24–29. [Google Scholar] [CrossRef]
- Chan, T.-F.; Chen, Y.-L.; Lee, C.-H.; Chou, F.-H.; Wu, L.-C.; Jong, S.-B.; Tsai, E.-M. Decreased Plsma Visfatin Concentrations in Women with Gestational Diabetes Mellitus. J. Soc. Gynecol. Investig. 2006, 13, 364–367. [Google Scholar] [CrossRef]
- Ingelsson, E.; Larson, M.G.; Fox, C.S.; Yin, X.; Wang, T.J.; Lipinska, I.; Pou, K.M.; Hoffmann, U.; Benjamin, E.J.; Keaney, J.F.; et al. Clinical Correlates of Circulating Visfatin Levels in a Community-Based Sample. Diabetes Care 2007, 30, 1278–1280. [Google Scholar] [CrossRef] [Green Version]
- Takebayashi, K.; Suetsugu, M.; Wakabayashi, S.; Aso, Y.; Inukai, T. Association between plasma visfatin and vascular endothelial function in patients with type 2 diabetes mellitus. Metabolism 2007, 56, 451–458. [Google Scholar] [CrossRef]
- Berndt, J.; Klöting, N.; Kralisch, S.; Kovacs, P.; Fasshauer, M.; Schön, M.R.; Stumvoll, M.; Blüher, M. Plasma Visfatin Concentrations and Fat Depot-Specific mRNA Expression in Humans. Diabetes 2005, 54, 2911–2916. [Google Scholar] [CrossRef] [Green Version]
- Pagano, C.; Pilon, C.; Olivieri, M.; Mason, P.; Fabris, R.; Serra, R.; Milan, G.; Rossato, M.; Federspil, G.; Vettor, R. Reduced Plasma Visfatin/Pre-B Cell Colony-Enhancing Factor in Obesity Is Not Related to Insulin Resistance in Humans. J. Clin. Endocrinol. Metab. 2006, 91, 3165–3170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jian, W.-X.; Luo, T.-H.; Gu, Y.-Y.; Zhang, H.-L.; Zheng, S.; Dai, M.; Han, J.-F.; Zhao, Y.; Li, G.; Luo, M. The visfatin gene is associated with glucose and lipid metabolism in a Chinese population. Diabet. Med. 2006, 23, 967–973. [Google Scholar] [CrossRef]
- Varma, V.; Yao-Borengasser, A.; Rasouli, N.; Bodles, A.M.; Phanavanh, B.; Lee, M.-J.; Starks, T.; Kern, L.M.; Spencer, H.J.; McGehee, R.E.; et al. Human Visfatin Expression: Relationship to Insulin Sensitivity, Intramyocellular Lipids, and Inflammation. J. Clin. Endocrinol. Metab. 2007, 92, 666–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, H.; Tang, S.-Y.; Luo, X.-H.; Huang, J.; Cui, R.-R.; Yuan, L.-Q.; Zhou, H.-D.; Wu, X.-P.; Liao, E.-Y. Insulin-Like Effects of Visfatin on Human Osteoblasts. Calcif. Tissue Int. 2007, 80, 201–210. [Google Scholar] [CrossRef]
- Pham, D.-V.; Park, P.-H. Recent insights on modulation of inflammasomes by adipokines: A critical event for the pathogenesis of obesity and metabolism-associated diseases. Arch. Pharmacal Res. 2020, 43, 997–1016. [Google Scholar] [CrossRef]
- Koka, S.; Xia, M.; Zhang, C.; Zhang, Y.; Li, P.-L.; Boini, K.M. Podocyte NLRP3 Inflammasome Activation and Formation by Adipokine Visfatin. Cell. Physiol. Biochem. 2019, 53, 355–365. [Google Scholar] [CrossRef]
- Xia, M.; Boini, K.M.; Abais, J.M.; Xu, M.; Zhang, Y.; Li, P.-L. Endothelial NLRP3 Inflammasome Activation and Enhanced Neointima Formation in Mice by Adipokine Visfatin. Am. J. Pathol. 2014, 184, 1617–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romacho, T.; Valencia, I.; Ramos-González, M.; Vallejo, S.; López-Esteban, M.; Lorenzo, O.; Cannata, P.; Romero, A.; Hipólito-Luengo, A.S.; Gómez-Cerezo, J.F.; et al. Visfatin/eNampt induces endothelial dysfunction in vivo: A role for Toll-Like Receptor 4 and NLRP3 inflammasome. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Pitzer, A.L.; Li, X.; Li, P.-L.; Wang, L.; Zhang, Y. Instigation of endothelial Nlrp3 inflammasome by adipokine visfatin promotes inter-endothelial junction disruption: Role of HMGB1. J. Cell. Mol. Med. 2015, 19, 2715–2727. [Google Scholar] [CrossRef]
- Ezzati-Mobaser, S.; Malekpour-Dehkordi, Z.; Nourbakhsh, M.; Tavakoli-Yaraki, M.; Ahmadpour, F.; Golpour, P.; Nourbakhsh, M. The up-regulation of markers of adipose tissue fibrosis by visfatin in pre-adipocytes as well as obese children and adolescents. Cytokine 2020, 134, 155193. [Google Scholar] [CrossRef]
- Bannigida, D.M.; Nayak, S.B.; R., V. Serum visfatin and adiponectin – markers in women with polycystic ovarian syndrome. Arch. Physiol. Biochem. 2018, 126, 1–4. [Google Scholar] [CrossRef]
- Ozkaya, M.; Cakal, E.; Ustun, Y.; Engin-Ustun, Y. Effect of metformin on serum visfatin levels in patients with polycystic ovary syndrome. Fertil. Steril. 2010, 93, 880–884. [Google Scholar] [CrossRef]
- Seow, K.-M.; Hwang, J.-L.; Wang, P.-H.; Ho, L.-T.; Juan, C.-C. Expression of visfatin mRNA in peripheral blood mononuclear cells is not correlated with visfatin mRNA in omental adipose tissue in women with polycystic ovary syndrome. Hum. Reprod. 2011, 26, 2869–2873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plati, E.; Kouskouni, E.; Malamitsi-Puchner, A.; Boutsikou, M.; Kaparos, G.; Baka, S. Visfatin and leptin levels in women with polycystic ovaries undergoing ovarian stimulation. Fertil. Steril. 2010, 94, 1451–1456. [Google Scholar] [CrossRef]
- Kim, J.J.; Choi, Y.M.; Hong, M.A.; Kim, M.J.; Chae, S.J.; Kim, S.M.; Hwang, K.R.; Yoon, S.H.; Ku, S.Y.; Suh, C.S.; et al. Serum visfatin levels in non-obese women with polycystic ovary syndrome and matched controls. Obstet. Gynecol. Sci. 2018, 61, 253–260. [Google Scholar] [CrossRef]
- Tsouma, I.; Kouskouni, E.; Demeridou, S.; Boutsikou, M.; Hassiakos, D.; Chasiakou, A.; Hassiakou, S.; Baka, S. Correlation of visfatin levels and lipoprotein lipid profiles in women with polycystic ovary syndrome undergoing ovarian stimulation. Gynecol. Endocrinol. 2014, 30, 516–519. [Google Scholar] [CrossRef]
- Pepene, C.E. Evidence for visfatin as an independent predictor of endothelial dysfunction in polycystic ovary syndrome. Clin. Endocrinol. 2011, 76, 119–125. [Google Scholar] [CrossRef]
- Lajunen, T.K.; Purhonen, A.-K.; Haapea, M.; Ruokonen, A.; Puukka, K.; Hartikainen, A.-L.; Savolainen, M.J.; Morin-Papunen, L.; Tapanainen, J.S.; Franks, S.; et al. Full-length visfatin levels are associated with inflammation in women with polycystic ovary syndrome. Eur. J. Clin. Investig. 2011, 42, 321–328. [Google Scholar] [CrossRef]
- Dambala, K.; Vavilis, D.; Bili, E.; Goulis, D.G.; Tarlatzis, B.C. Serum visfatin, vascular endothelial growth factor and matrix metalloproteinase-9 in women with polycystic ovary syndrome. Gynecol. Endocrinol. 2017, 33, 529–533. [Google Scholar] [CrossRef] [PubMed]
- DeRosa, G.; D’Angelo, A.; Romano, D.; Maffioli, P. Effects of metformin extended release compared to immediate release formula on glycemic control and glycemic variability in patients with type 2 diabetes. Drug Des. Dev. Ther. 2017, 11, 1481–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahra, M.; Shah, M.; Ali, A.; Rahim, R. Effects of Metformin on Endocrine and Metabolic Parameters in Patients with Polycystic Ovary Syndrome. Horm. Metab. Res. 2016, 49, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.-H.; He, C.-T.; Lee, C.-H.; Wu, L.-Y.; Hung, Y.-J. Both slow-release and regular-form metformin improve glycemic control without altering plasma visfatin level in patients with type 2 diabetes mellitus. Metabolism 2007, 56, 1087–1092. [Google Scholar] [CrossRef]
- Kadoglou, N.P.; Tsanikidis, H.; Kapelouzou, A.; Vrabas, I.; Vitta, I.; Karayannacos, P.E.; Liapis, C.D.; Sailer, N. Effects of rosiglitazone and metformin treatment on apelin, visfatin, and ghrelin levels in patients with type 2 diabetes mellitus. Metabolism 2010, 59, 373–379. [Google Scholar] [CrossRef]
- Aigner, E.; Bachofner, N.; Klein, K.; De Geyter, C.; Hohla, F.; Patsch, W.; Datz, C. Retinol-Binding Protein 4 in Polycystic Ovary Syndrome—Association with Steroid Hormones and Response to Pioglitazone Treatment. J. Clin. Endocrinol. Metab. 2009, 94, 1229–1235. [Google Scholar] [CrossRef] [Green Version]
- Erdem, G.; Dogru, T.; Tasci, I.; Bozoglu, E.; Muhsiroglu, O.; Tapan, S.; Ercin, C.N.; Sonmez, A. The effects of pioglitazone and metformin on plasma visfatin levels in patients with treatment naive type 2 diabetes mellitus. Diabetes Res. Clin. Pract. 2008, 82, 214–218. [Google Scholar] [CrossRef]
- El-Sheikh, H.M.; El-Haggar, S.M.; Elbedewy, T.A. Comparative study to evaluate the effect of l-carnitine plus glimepiride versus glimepiride alone on insulin resistance in type 2 diabetic patients. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 167–173. [Google Scholar] [CrossRef]
- DeRosa, G.; Maffioli, P.; Ferrari, I.; D’Angelo, A.; Fogari, R.; Palumbo, I.; Randazzo, S.; Cicero, A.F.G. Orlistat and L-carnitine compared to orlistat alone on insulin resistance in obese diabetic patients. Endocr. J. 2010, 57, 777–786. [Google Scholar] [CrossRef] [Green Version]
- DeRosa, G.; Maffioli, P.; Salvadeo, S.A.T.; Ferrari, I.; Gravina, A.; Mereu, R.; D’Angelo, A.; Palumbo, I.; Randazzo, S.; Cicero, A.F.G. Sibutramine and L-Carnitine Compared to Sibutramine Alone on Insulin Resistance in Diabetic Patients. Intern. Med. 2010, 49, 1717–1725. [Google Scholar] [CrossRef] [Green Version]
- Saddi-Rosa, P.; Oliveira, C.S.V.; Giuffrida, F.M.; Reis, A.F. Visfatin, glucose metabolism and vascular disease: A review of evidence. Diabetol. Metab. Syndr. 2010, 2, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherer, P.E.; Hill, J.A. Obesity, Diabetes, and Cardiovascular Diseases. Circ. Res. 2016, 118, 1703–1705. [Google Scholar] [CrossRef] [Green Version]
- Sethi, J.K. Is PBEF/visfatin/Nampt an authentic adipokine relevant to the metabolic syndrome? Curr. Hypertens. Rep. 2007, 9, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Körner, A.; Garten, A.; Blüher, M.; Tauscher, R.; Kratzsch, J.; Kiess, W. Molecular Characteristics of Serum Visfatin and Differential Detection by Immunoassays. J. Clin. Endocrinol. Metab. 2007, 92, 4783–4791. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| NAD+-Dependent Cellular Signaling | Functional Outcomes |
|---|---|
| Sirtuins | Cell division/proliferation Cell differentiation Cell survival/apoptosis Life span Inflammation Cell metabolism/expenditure |
| CD38/CD157 | Ca2+ signaling |
| PARPs/MARTs | Cell proliferation DNA repair/DNA integrity Cellular responses to environmental/oxidative stress Proper gene expression Mitochondrial health Transcriptional and post-translational regulation |
| Functional Role | Possible Outcomes |
|---|---|
| PBEF | Enhances murine pre-B-cell colony formation Upregulates SCF and IL-7 |
| Cytokine | Inflammatory pathways: NF-κB, MAPK, PI3 Vascular remodeling: ↑ MCP-1, ↑ MMP, ↑ VGEF, ↑ FGF-2 |
| Insulin-mimetic | Binds insulin receptor Increases insulin sensitivity and glucose lowering effects Enhances glucose uptake/transport Lipogenesis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dakroub, A.; A. Nasser, S.; Younis, N.; Bhagani, H.; Al-Dhaheri, Y.; Pintus, G.; Eid, A.A.; El-Yazbi, A.F.; Eid, A.H. Visfatin: A Possible Role in Cardiovasculo-Metabolic Disorders. Cells 2020, 9, 2444. https://doi.org/10.3390/cells9112444
Dakroub A, A. Nasser S, Younis N, Bhagani H, Al-Dhaheri Y, Pintus G, Eid AA, El-Yazbi AF, Eid AH. Visfatin: A Possible Role in Cardiovasculo-Metabolic Disorders. Cells. 2020; 9(11):2444. https://doi.org/10.3390/cells9112444
Chicago/Turabian StyleDakroub, Ali, Suzanne A. Nasser, Nour Younis, Humna Bhagani, Yusra Al-Dhaheri, Gianfranco Pintus, Assaad A. Eid, Ahmed F. El-Yazbi, and Ali H. Eid. 2020. "Visfatin: A Possible Role in Cardiovasculo-Metabolic Disorders" Cells 9, no. 11: 2444. https://doi.org/10.3390/cells9112444
APA StyleDakroub, A., A. Nasser, S., Younis, N., Bhagani, H., Al-Dhaheri, Y., Pintus, G., Eid, A. A., El-Yazbi, A. F., & Eid, A. H. (2020). Visfatin: A Possible Role in Cardiovasculo-Metabolic Disorders. Cells, 9(11), 2444. https://doi.org/10.3390/cells9112444