Nuclear Hormone Receptors and Their Ligands: Metabolites in Control of Transcription

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. A Brief History of NHRs and Their Ligands
2.1. Phenomenology (1849–1914)
2.2. Hunting for Hormones (1915–1984)
2.3. Ligands Meet Receptors (1985–1997)
2.4. Orphan NHRs Meet Their Ligands (1998–Present)
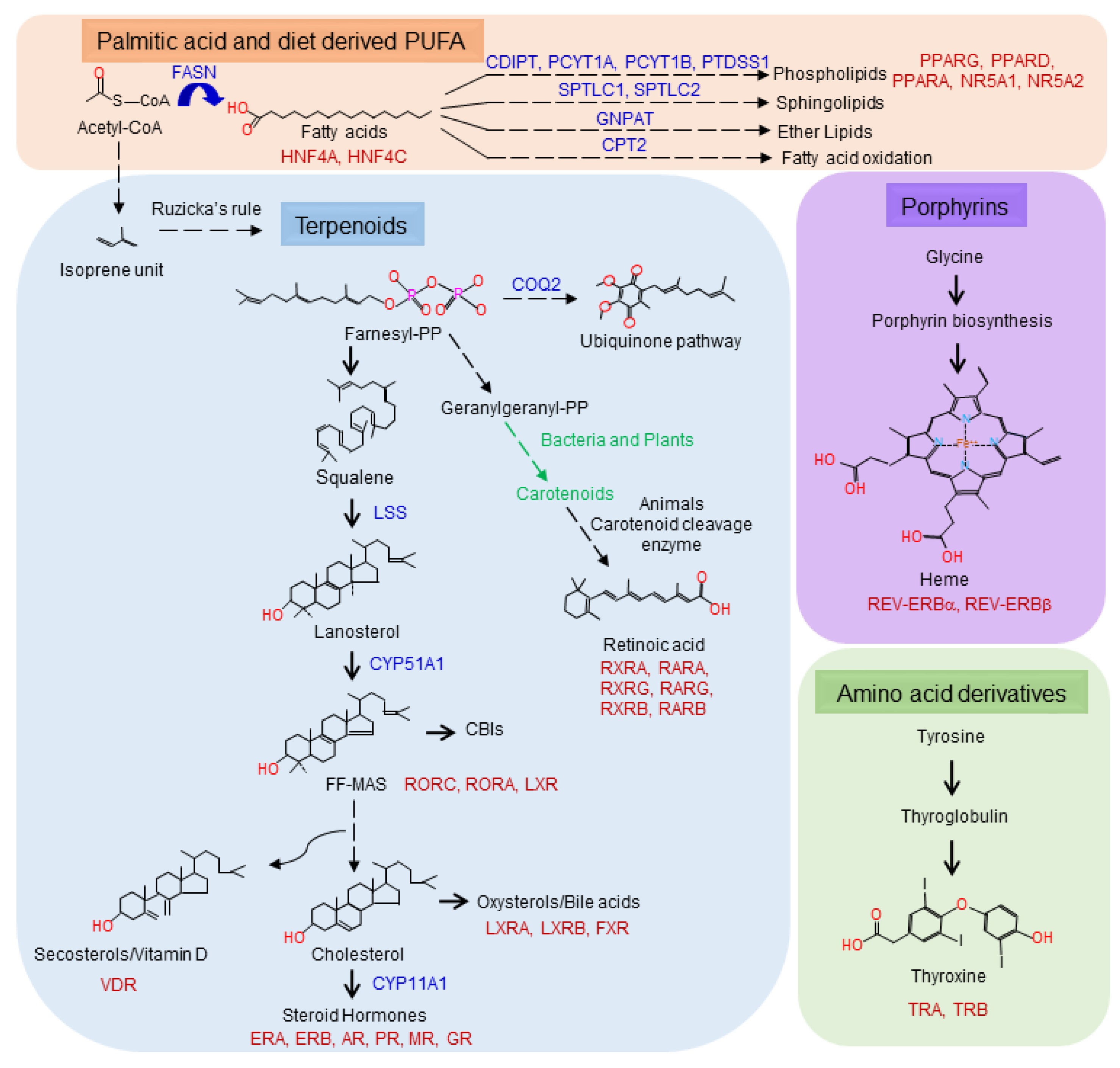
3. Metabolome as a Source of NHR Ligands
3.1. Fatty Acid Family Ligands
3.2. Terpenoid Family Ligands
3.3. Porphyrins
3.4. Amino Acid Derivatives
4. Orphan Receptors, What Ligands?
5. Common Properties of NHR Ligand Biosynthetic Pathways
Author Contributions
Funding
Conflicts of Interest
References
- Santori, F.R. Nuclear hormone receptors put immunity on sterols. Eur. J. Immunol. 2015, 45, 2730–2741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graupner, G.; Wills, K.N.; Tzukerman, M.; Zhang, X.-K.; Pfahl, M. Dual regulatory role for thyroid-hormone receptors allows control of retinoic-acid receptor activity. Nat. Cell Biol. 1989, 340, 653–656. [Google Scholar] [CrossRef] [PubMed]
- Sakai, D.D.; Helms, S.; Carlstedt-Duke, J.; A Gustafsson, J.; Rottman, F.M.; Yamamoto, K.R. Hormone-mediated repression: A negative glucocorticoid response element from the bovine prolactin gene. Genes Dev. 1988, 2, 1144–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, R.; Ursu, O.; Gaulton, A.; Bento, A.P.; Donadi, R.S.; Bologa, C.G.; Karlsson, A.; Al-Lazikani, B.; Hersey, A.; Oprea, T.; et al. A comprehensive map of molecular drug targets. Nat. Rev. Drug Discov. 2017, 16, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Shpakov, A.O.; Pertseva, M.N. Chapter 4 Signaling Systems of Lower Eukaryotes and Their Evolution. Int. Rev. Cell Mol. Biol. 2008, 269, 151–282. [Google Scholar] [CrossRef] [PubMed]
- Bridgham, J.T.; Eick, G.N.; Larroux, C.; Deshpande, K.; Harms, M.J.; Gauthier, M.-E.A.; Ortlund, E.A.; Degnan, B.M.; Thornton, J.W. Protein Evolution by Molecular Tinkering: Diversification of the Nuclear Receptor Superfamily from a Ligand-Dependent Ancestor. PLoS Biol. 2010, 8, e1000497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthold, A.A. Transplantation der hoden. Arch. Anat. Physiol. Wissen Med. 1849, 16, 42–46. [Google Scholar]
- Séquard, B. Note on the effects produced on man by subcutaneous injections of a liquid obtained from the testicles of animals. Lancet 1889, 134, 105–107. [Google Scholar] [CrossRef] [Green Version]
- Murray, G.R. Note on the Treatment of Myxœdema by Hypodermic Injections of an Extract of the Thyroid Gland of a Sheep. Br. Med. J. 1891, 2, 796–797. [Google Scholar] [CrossRef]
- Baumann, E. Ueber das Thyrojodin. Munch. Med. Wschr. 1896, 43, 309–312. [Google Scholar]
- Starling, E.H. Croonian Lecture: On the chemical correlation of the functions of the body. I. Lancet 1905, 166, 339–341. [Google Scholar] [CrossRef]
- Starling, E.H. Croonian Lecture: On the chemical correlation of the functions of the body. IV. Lancet 1905, 166, 579–583. [Google Scholar] [CrossRef]
- Kendall, E.C. The isolation in crystalline form of the compound containing iodin, which occurs in the thyroid. Its chemical nature and physiological activity. J. Am. Med Assoc. 1915, 30, 2042–2043. [Google Scholar] [CrossRef] [Green Version]
- Harington, C.R.; Barger, G. Chemistry of Thyroxine: Constitution and Synthesis of Thyroxine. Biochem. J. 1927, 21, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Doisy, E.A.; Veler, C.D.; Thayer, S.A. Folliculin from Urine of Pregnant Women; American Phsyiological Society: Rockville, MD, USA, 1929. [Google Scholar]
- Butenandt, A. Über“ Progynon ”ein krystallisiertes weibliches Sexualhormon. NW 1929, 17, 879. [Google Scholar]
- Butenandt, A.; Hanisch, G. Über die Umwandlung des Dehydro-androsterons in Δ4-Androsten-ol-(17)-0n-(3) (Testosteron); ein Weg zur Darstellung des Testosterons aus Cholesterin (Vorläuf. Mitteil.). Ber. Dtsch. Chem. Ges. (A B Ser.) 1935, 68, 1859–1862. [Google Scholar] [CrossRef]
- Ruzicka, L.; Wettstein, A. Sexualhormone VII. Über die künstliche Herstellung des Testikelhormons Testosteron (Androsten-3-on-17-ol). Helvetica Chim. Acta 1935, 18, 1264–1275. [Google Scholar] [CrossRef]
- David, K.; Dingemanse, E.; Freud, J.; Laqueur, E. Über krystallinisches männliches Hormon aus Hoden (Testosteron), wirksamer als aus Harn oder aus Cholesterin bereitetes Androsteron. Biol. Chem. 1935, 233, 281. [Google Scholar] [CrossRef]
- Butenandt, A.; Westphal, U. Zur Isolierung und Charakterisierung des Corpus-luteum-Hormons. Ber. Dtsch. Chem. Gesellschaf 1934, 67, 1440–1442. [Google Scholar] [CrossRef]
- Mason, H.L.; Hoehn, W.M.; Kendall, E.C. Chemical studies of the suprarenal cortex: IV. Structures of compounds C,D,E,F, and G. J. Biol. Chem. 1938, 124, 459–474. [Google Scholar]
- Mason, H.L.; Myers, C.S.; Kendall, E.C. Chemical studies of the suprarenal cortex: II. The identification of a substance which possesses the qualitative action of cortin; its conversion into a diketone closely related to androstenedione. J. Biol. Chem. 1936, 116, 267–276. [Google Scholar]
- Reichstein, T. Über Bestandteile der Nebennieren-Rinde VI. Trennungsmethoden, sowie Isolierung der Substanzen F. a. H und J. Helv. Chem. Acta 1936, 19, 1107–1126. [Google Scholar] [CrossRef]
- Butenandt, A. Über die chemische Untersuchung der Sexualhormone. Angew. Chem. 1931, 44, 905–908. [Google Scholar] [CrossRef]
- Simons, S.S., Jr.; Thompson, E.B. Dexamethasone 21-mesylate: An affinity label of glucocorticoid receptors from rat hepatoma tissue culture cells. Proc. Natl. Acad. Sci. USA 1981, 78, 3541–3545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gehring, U.; Hotz, A. Photoaffinity labeling and partial proteolysis of wild-type and variant glucocorticoid receptors. Biochemistry 1983, 22, 4013–4018. [Google Scholar] [CrossRef] [PubMed]
- Hollenberg, S.M.; Weinberger, C.; Ong, E.S.; Cerelli, G.; Oro, A.; Lebo, R.; Thompson, E.B.; Rosenfeld, M.G.; Evans, R.M. Primary structure and expression of a functional human glucocorticoid receptor cDNA. Nat. Cell Biol. 1985, 318, 635–641. [Google Scholar] [CrossRef]
- Giguere, V.; Ong, E.S.; Segui, P.; Evans, R.M. Identification of a receptor for the morphogen retinoic acid. Nat. Cell Biol. 1987, 330, 624–629. [Google Scholar] [CrossRef]
- Petkovich, M.; Brand, N.J.; Krust, A.; Chambon, P. A human retinoic acid receptor which belongs to the family of nuclear receptors. Nature 1987, 330, 444–450. [Google Scholar] [CrossRef]
- Clarke, N.D. Zinc Fingers in Caenorhabditis elegans: Finding Families and Probing Pathways. Science 1998, 282, 2018–2022. [Google Scholar] [CrossRef] [Green Version]
- Arenas, E. Faculty Opinions recommendation of An oxysterol signalling pathway mediated by the nuclear receptor LXR alpha. Nature 2015, 383, 728–731. [Google Scholar] [CrossRef]
- Makishima, M.; Okamoto, A.Y.; Repa, J.J.; Tu, H.; Learned, R.M.; Luk, A.; Hull, M.V.; Lustig, K.D.; Mangelsdorf, D.J.; Shan, B. Identification of a nuclear receptor for bile acids. Science 1999, 284, 1362–1365. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.J.; Blanchard, S.G.; Bledsoe, R.K.; Chandra, G.; Consler, T.G.; Kliewer, S.A.; Stimmel, J.B.; Willson, T.M.; Zavacki, A.M.; Moore, D.D.; et al. Bile acids: Natural ligands for an orphan nuclear receptor. Science 1999, 284, 1365–1368. [Google Scholar] [CrossRef] [PubMed]
- Santori, F.R.; Huang, P.; van de Pavert, S.A.; Douglass, E.F., Jr.; Leaver, D.J.; Haubrich, B.A.; Keber, R.; Lorbek, G.; Konijn, T.; Rosales, B.N.; et al. Identification of Natural RORgamma Ligands that Regulate the Development of Lymphoid Cells. Cell Metab. 2015, 21, 286–297. [Google Scholar] [CrossRef] [Green Version]
- Motola, D.L.; Cummins, C.L.; Rottiers, V.; Sharma, K.K.; Li, T.; Li, Y.; Suino-Powell, K.; Xu, H.E.; Auchus, R.J.; Antebi, A.; et al. Identification of ligands for DAF-12 that govern dauer formation and reproduction in C. elegans. Cell 2006, 124, 1209–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Zhou, X.E.; Motola, D.L.; Gao, X.; Suino-Powell, K.; Conneely, A.; Ogata, C.; Sharma, K.K.; Auchus, R.J.; Lok, J.B.; et al. Identification of the nuclear receptor DAF-12 as a therapeutic target in parasitic nematodes. Proc. Natl. Acad. Sci. USA 2009, 106, 9138–9143. [Google Scholar] [CrossRef] [Green Version]
- Retnakaran, A.; Krell, P.; Feng, Q.; Arif, B. Ecdysone agonists: Mechanism and importance in controlling insect pests of agriculture and forestry. Arch. Insect. Biochem. Physiol. 2003, 54, 187–199. [Google Scholar] [CrossRef]
- Nuclear Receptors Nomenclature Committee. A unified nomenclature system for the nuclear receptor superfamily. Cell 1999, 97, 161–163. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, B.E.; Hogenesch, J.B.; Bradfield, C.A. Mammalian Per-Arnt-Sim proteins in environmental adaptation. Ann. Rev. Phys. 2010, 72, 625–645. [Google Scholar] [CrossRef]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Bloch, K. The biological synthesis of cholesterol. Science 1965, 150, 19–28. [Google Scholar] [CrossRef]
- Villas-Boas, S.G. Metabolome Analysis: An Introduction; Wiley-Interscience: Hoboken, NJ, USA, 2007. [Google Scholar]
- Srivastava, M.; Simakov, O.; Chapman, J.; Fahey, B.; Gauthier, M.E.A.; Mitros, T.; Richards, G.S.; Conaco, C.; Dacre, M.; Hellsten, U.; et al. The Amphimedon queenslandica genome and the evolution of animal complexity. Nat. Cell Biol. 2010, 466, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Hertz, R.; Magenheim, J.; Berman, I.; Bar-Tana, J. Fatty acyl-CoA thioesters are ligands of hepatic nuclear factor-4alpha. Nature 1998, 392, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Krylova, I.N.; Sablin, E.P.; Moore, J.; Xu, R.X.; Waitt, G.M.; Mackay, J.A.; Juzumiene, D.; Bynum, J.M.; Madauss, K.; Montana, V.; et al. Structural Analyses Reveal Phosphatidyl Inositols as Ligands for the NR5 Orphan Receptors SF-1 and LRH-1. Cell 2005, 120, 343–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyman, R.A.; Mangelsdorf, D.J.; Dyck, J.A.; Stein, R.B.; Eichele, G.; Evans, R.M.; Thaller, C. 9-cis retinoic acid is a high affinity ligand for the retinoid X receptor. Cell 1992, 68, 397–406. [Google Scholar] [CrossRef]
- Levin, A.A.; Sturzenbecker, L.J.; Kazmer, S.; Bosakowski, T.; Huselton, C.; Allenby, G.; Speck, J.; Kratzeisen, C.; Rosenberger, M.; Lovey, A.; et al. 9-cis retinoic acid stereoisomer binds and activates the nuclear receptor RXR alpha. Nature 1992, 355, 359–361. [Google Scholar] [CrossRef]
- Sap, J.; Muñoz, A.; Damm, K.; Goldberg, Y.; Ghysdael, J.; Leutz, A.; Beug, H.; Vennström, B. The c-erb-A protein is a high-affinity receptor for thyroid hormone. Nature 1986, 324, 635–640. [Google Scholar] [CrossRef]
- Weinberger, C.; Thompson, C.C.; Ong, E.S.; Lebo, R.; Gruol, D.J.; Evans, R.M. The c-erb-A gene encodes a thyroid hormone receptor. Nature 1986, 324, 641–646. [Google Scholar] [CrossRef]
- Raghuram, S.; Stayrook, K.R.; Huang, P.; Rogers, P.M.; Nosie, A.K.; McClure, D.B.; Burris, L.L.; Khorasanizadeh, S.; Burris, T.P.; Rastinejad, F. Identification of heme as the ligand for the orphan nuclear receptors REV-ERBalpha and REV-ERBbeta. Nat. Struct. Mol. Biol. 2007, 14, 1207–1213. [Google Scholar] [CrossRef] [Green Version]
- Umeno, A.; Sakashita, M.; Sugino, S.; Murotomi, K.; Okuzawa, T.; Morita, N.; Tomii, K.; Tsuchiya, Y.; Yamasaki, K.; Horie, M.; et al. Comprehensive analysis of PPARgamma agonist activities of stereo-, regio-, and enantio-isomers of hydroxyoctadecadienoic acids. Biosci. Rep. 2020, 40, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Kliewer, S.A.; Lenhard, J.M.; Willson, T.M.; Patel, I.; Morris, D.C.; Lehmann, J.M. A prostaglandin J2 metabolite binds peroxisome proliferator-activated receptor gamma and promotes adipocyte differentiation. Cell 1995, 83, 813–819. [Google Scholar] [CrossRef] [Green Version]
- Forman, B.M.; Tontonoz, P.; Chen, J.; Brun, R.P.; Spiegelman, B.M.; Evans, R.M. 15-Deoxy-delta 12, 14-prostaglandin J2 is a ligand for the adipocyte determination factor PPAR gamma. Cell 1995, 83, 803–812. [Google Scholar] [CrossRef] [Green Version]
- Fahy, E.; Subramaniam, S.; Murphy, R.C.; Nishijima, M.; Raetz, C.R.H.; Shimizu, T.; Spener, F.; Van Meer, G.; Wakelam, M.J.O.; Dennis, E.A. Update of the LIPID MAPS comprehensive classification system for lipids. J. Lipid Res. 2008, 50, S9–S14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christianson, D.W. Structural and Chemical Biology of Terpenoid Cyclases. Chem. Rev. 2017, 117, 11570–11648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruzicka, L. The isoprene rule and the biogenesis of terpenic compounds. Cell. Mol. Life Sci. 1953, 9, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.L.; Brown, M.S. Regulation of the mevalonate pathway. Nat. Cell Biol. 1990, 343, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.-H.; Zhu, J.; Jiang, J.-G. Carotenoids biosynthesis and cleavage related genes from bacteria to plants. Crit. Rev. Food Sci. Nutr. 2017, 58, 2314–2333. [Google Scholar] [CrossRef]
- Green, S.; Walter, P.; Kumar, V.; Krust, A.; Bornert, J.-M.; Argos, P.; Chambon, P. Human oestrogen receptor cDNA: Sequence, expression and homology to v-erb-A. Nat. Cell Biol. 1986, 320, 134–139. [Google Scholar] [CrossRef]
- Greene, G.L.; Gilna, P.; Waterfield, M.; Baker, A.; Hort, Y.; Shine, J. Sequence and expression of human estrogen receptor complementary DNA. Science 1986, 231, 1150–1154. [Google Scholar] [CrossRef]
- Jeltsch, J.M.; Krozowski, Z.; Quirin-Stricker, C.; Gronemeyer, H.; Simpson, R.J.; Garnier, J.M.; Krust, A.; Jacob, F.; Chambon, P. Cloning of the chicken progesterone receptor. Proc. Natl. Acad. Sci. USA 1986, 83, 5424–5428. [Google Scholar] [CrossRef] [Green Version]
- Conneely, O.M.; Sullivan, W.P.; O Toft, D.; Birnbaumer, M.; Cook, R.G.; Maxwell, B.L.; Zarucki-Schulz, T.; Greene, G.L.; Schrader, W.T.; O’Malley, B.W. Molecular cloning of the chicken progesterone receptor. Science 1986, 233, 767–770. [Google Scholar] [CrossRef]
- Chang, C.S.; Kokontis, J.; Liao, S.T. Molecular cloning of human and rat complementary DNA encoding androgen receptors. Science 1988, 240, 324–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lubahn, D.B.; Joseph, D.R.; Sullivan, P.M.; Willard, H.F.; French, F.S.; Wilson, E.M. Cloning of human androgen receptor complementary DNA and localization to the X chromosome. Science 1988, 240, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Arriza, J.L.; Weinberger, C.; Cerelli, G.; Glaser, T.M.; Handelin, B.L.; E Housman, D.; Evans, R.M. Cloning of human mineralocorticoid receptor complementary DNA: Structural and functional kinship with the glucocorticoid receptor. Science 1987, 237, 268–275. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, D.P.; Mangelsdorf, D.J.; Pike, J.W.; Haussler, M.R.; O’Malley, B.W. Molecular cloning of complementary DNA encoding the avian receptor for vitamin D. Science 1987, 235, 1214–1217. [Google Scholar] [CrossRef] [PubMed]
- IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN). The Nomenclature of Steroids. JBIC J. Biol. Inorg. Chem. 2005, 10, 1–19. [Google Scholar]
- Reinking, J.; Lam, M.M.; Pardee, K.; Sampson, H.M.; Liu, S.; Yang, P.; Williams, S.P.; White, W.; Lajoie, G.; Edwards, A.M.; et al. The Drosophila Nuclear Receptor E75 Contains Heme and Is Gas Responsive. Cell 2005, 122, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Wu, N.; Curtin, J.C.; Qatanani, M.; Szwergold, N.R.; Reid, R.A.; Waitt, G.M.; Parks, D.J.; Pearce, K.H.; Wisely, G.B.; et al. Rev-erb, a Heme Sensor That Coordinates Metabolic and Circadian Pathways. Science 2007, 318, 1786–1789. [Google Scholar] [CrossRef]
- Brent, G.A. Mechanisms of thyroid hormone action. J. Clin. Investig. 2012, 122, 3035–3043. [Google Scholar] [CrossRef] [Green Version]
- Eales, J.G. Iodine metabolism and thyroid-related functions in organisms lacking thyroid follicles: Are thyroid hormones also vitamins? Proc. Soc. Exp. Boil. Med. 1997, 214, 302–331. [Google Scholar] [CrossRef]
- Wang, Z.; Benoit, G.; Liu, J.; Prasad, S.; Aarnisalo, P.; Liu, X.; Xu, H.; Walker, N.P.C.; Perlmann, T. Structure and function of Nurr1 identifies a class of ligand-independent nuclear receptors. Nat. Cell Biol. 2003, 423, 555–560. [Google Scholar] [CrossRef]
- Woo, E.J.; Jeong, D.G.; Lim, M.Y.; Jun Kim, S.; Kim, K.J.; Yoon, S.M.; Park, B.C.; Ryu, S.E. Structural insight into the constitutive repression function of the nuclear receptor Rev-erbbeta. J. Mol. Biol. 2007, 373, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Pleasants, J.R.; Johnson, M.H.; Wostmann, B.S. Adequacy of Chemically Defined, Water-Soluble Diet for Germfree BALB/c Mice Through Successive Generations and Litters. J. Nutr. 1986, 116, 1949–1964. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.C.; Micchelli, C.A. Development and characterization of a chemically defined food for Drosophila. PLoS ONE 2013, 8, e67308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hieb, W.F.; Rothstein, M. Sterol Requirement for Reproduction of a Free-Living Nematode. Science 1968, 160, 778–780. [Google Scholar] [CrossRef]
- Crowder, C.M.; Westover, E.J.; Kumar, A.S.; Ostlund, R.E., Jr.; Covey, D.F. Enantiospecificity of cholesterol function in vivo. J. Biol. Chem. 2001, 276, 44369–44372. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Wang, Y.; Hao, L.Y.; Liu, X.; Lesch, C.A.; Sanchez, B.M.; Wendling, J.M.; Morgan, R.W.; Aicher, T.D.; Carter, L.L.; et al. Sterol metabolism controls TH17 differentiation by generating endogenous RORgamma agonists. Nat. Chem. Biol. 2015, 11, 141–147. [Google Scholar] [CrossRef]
- Spann, N.J.; Garmire, L.X.; McDonald, J.G.; Myers, D.S.; Milne, S.B.; Shibata, N.; Reichart, D.; Fox, J.N.; Shaked, I.; Heudobler, D.; et al. Regulated Accumulation of Desmosterol Integrates Macrophage Lipid Metabolism and Inflammatory Responses. Cell 2012, 151, 138–152. [Google Scholar] [CrossRef] [Green Version]
- Byskov, A.G.; Andersen, C.Y.; Nordholm, L.; Thogersen, H.; Guoliang, X.; Wassmann, O.; Andersen, J.V.; Guddal, E.; Roed, T. Chemical structure of sterols that activate oocyte meiosis. Nature 1995, 374, 559–562. [Google Scholar] [CrossRef]
- Tabacik, C.; Aliau, S.; Serrou, B.; Crastes de Paulet, A. Post-HMG CoA reductase regulation of cholesterol biosynthesis in normal human lymphocytes: Lanosten-3 betal-ol-32-al, a natural inhibitor. Biochem. Biophys. Res. Commun. 1981, 101, 1087–1095. [Google Scholar] [CrossRef]
- Trzaskos, J.M.; Favata, M.F.; Fischer, R.T.; Stam, S.H. In situ accumulation of 3 beta-hydroxylanost-8-en-32-aldehyde in hepatocyte cultures. A putative regulator of 3-hydroxy-3-methylglutaryl-coenzyme A reductase activity. J. Biol. Chem. 1987, 262, 12261–12268. [Google Scholar]
- Esaki, K.; Sayano, T.; Sonoda, C.; Akagi, T.; Suzuki, T.; Ogawa, T.; Okamoto, M.; Yoshikawa, T.; Hirabayashi, Y.; Furuya, S. l-Serine Deficiency Elicits Intracellular Accumulation of Cytotoxic Deoxysphingolipids and Lipid Body Formation. J. Biol. Chem. 2015, 290, 14595–14609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lone, M.A.; Santos, T.; Alecu, I.; Silva, L.C.; Hornemann, T. 1-Deoxysphingolipids. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N. Pro-resolving lipid mediators are leads for resolution physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Petasis, N.A. Resolvins and Protectins in Inflammation Resolution. Chem. Rev. 2011, 111, 5922–5943. [Google Scholar] [CrossRef] [Green Version]
- Chakravarthy, M.V.; Lodhi, I.J.; Yin, L.; Malapaka, R.R.; Xu, H.E.; Turk, J.; Semenkovich, C.F. Identification of a physiologically relevant endogenous ligand for PPARalpha in liver. Cell 2009, 138, 476–488. [Google Scholar] [CrossRef] [Green Version]
- Tiefenbach, J.; Magomedova, L.; Liu, J.; Reunov, A.A.; Tsai, R.; Eappen, N.S.; Jockusch, R.A.; Nislow, C.; Cummins, C.L.; Krause, H.M. Idebenone and coenzyme Q10 are novel PPARalpha/gamma ligands, with potential for treatment of fatty liver diseases. Dis. Model. Mech. 2018, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Gaetani, S.; Oveisi, F.; Verme, J.L.; Serrano, A.; De Fonseca, F.R.; Rosengarth, A.; Luecke, H.; Di Giacomo, B.; Tarzia, G.; et al. Oleylethanolamide regulates feeding and body weight through activation of the nuclear receptor PPAR-α. Nature 2003, 425, 90–93. [Google Scholar] [CrossRef]
- Zhang, M.; Sayyad, A.; Dhesi, A.; Orellana, A. (Enantioselective Synthesis of 7(S)-Hydroxydocosahexaenoic Acid, a Brain-Specific Endogenous Ligand for PPARalpha. J. Org. Chem. 2020. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, L.J.; Seo, D.E.; Jackson, B.; Ivanova, N.B.; Santori, F.R. Nuclear Hormone Receptors and Their Ligands: Metabolites in Control of Transcription. Cells 2020, 9, 2606. https://doi.org/10.3390/cells9122606
Tao LJ, Seo DE, Jackson B, Ivanova NB, Santori FR. Nuclear Hormone Receptors and Their Ligands: Metabolites in Control of Transcription. Cells. 2020; 9(12):2606. https://doi.org/10.3390/cells9122606
Chicago/Turabian StyleTao, Lian Jing, Dong Eun Seo, Benjamin Jackson, Natalia B. Ivanova, and Fabio Rinaldo Santori. 2020. "Nuclear Hormone Receptors and Their Ligands: Metabolites in Control of Transcription" Cells 9, no. 12: 2606. https://doi.org/10.3390/cells9122606
APA StyleTao, L. J., Seo, D. E., Jackson, B., Ivanova, N. B., & Santori, F. R. (2020). Nuclear Hormone Receptors and Their Ligands: Metabolites in Control of Transcription. Cells, 9(12), 2606. https://doi.org/10.3390/cells9122606