Chondrogenic Differentiation from Induced Pluripotent Stem Cells Using Non-Viral Minicircle Vectors

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Minicircle Production
2.2. hiPSC Culture
2.3. EB Generation and OG Induction
2.4. Minicircle Transfection
2.5. Enzyme-Linked Immunosorbent Assay
2.6. Osteogenic Differentiation
2.7. Adipogenic Differentiation
2.8. Chondrogenic Differentiation Using Pellet Culture
2.9. Polymerase Chain Reaction
2.10. Ethics
2.11. Osteochondral Defect Model
2.12. Histological Analysis
2.13. Immunohistochemistry
2.14. Flow Cytometry
2.15. Statistical Analysis
2.16. Ethics
3. Results
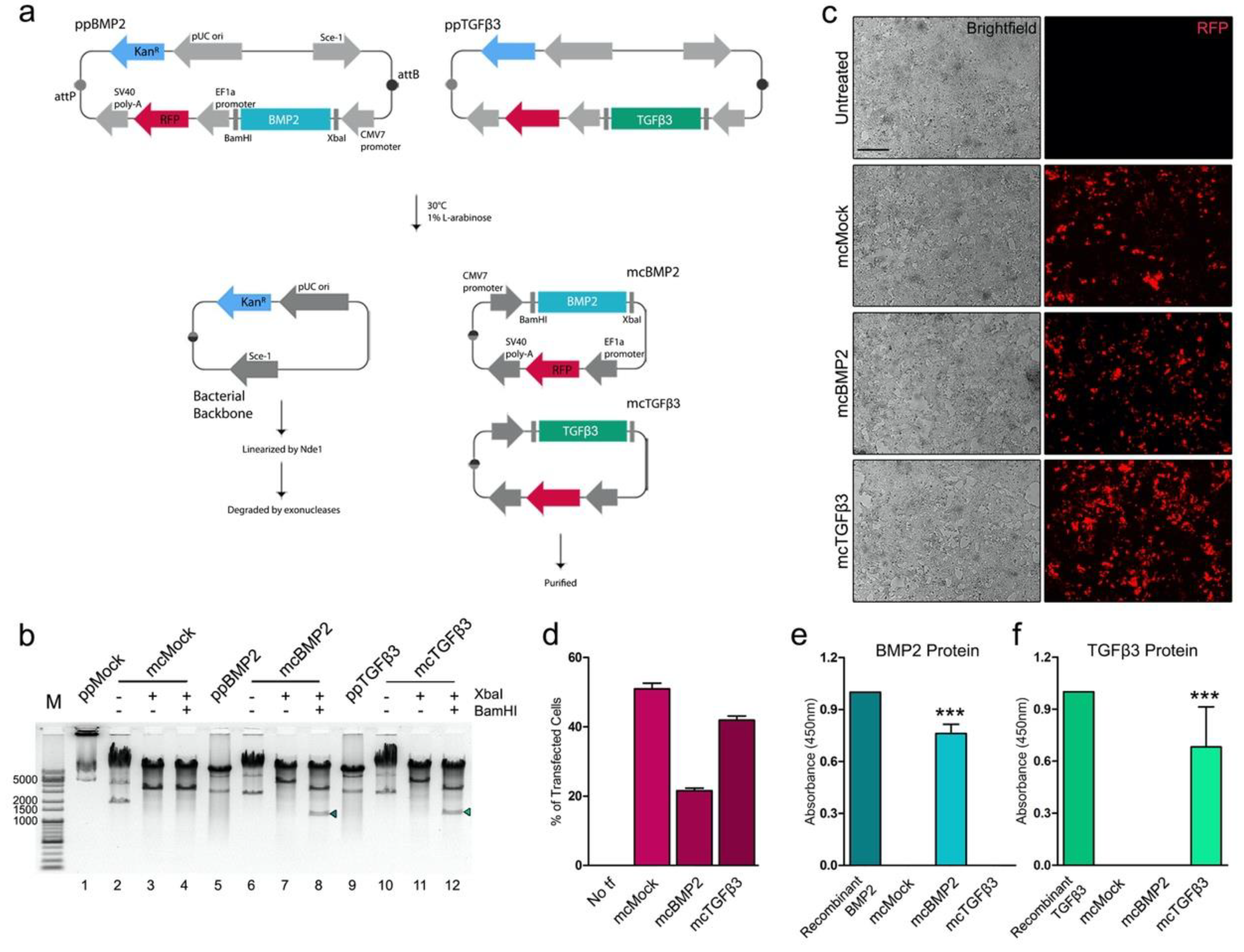
3.1. Generation of Minicircles Encoding Human Growth Factors
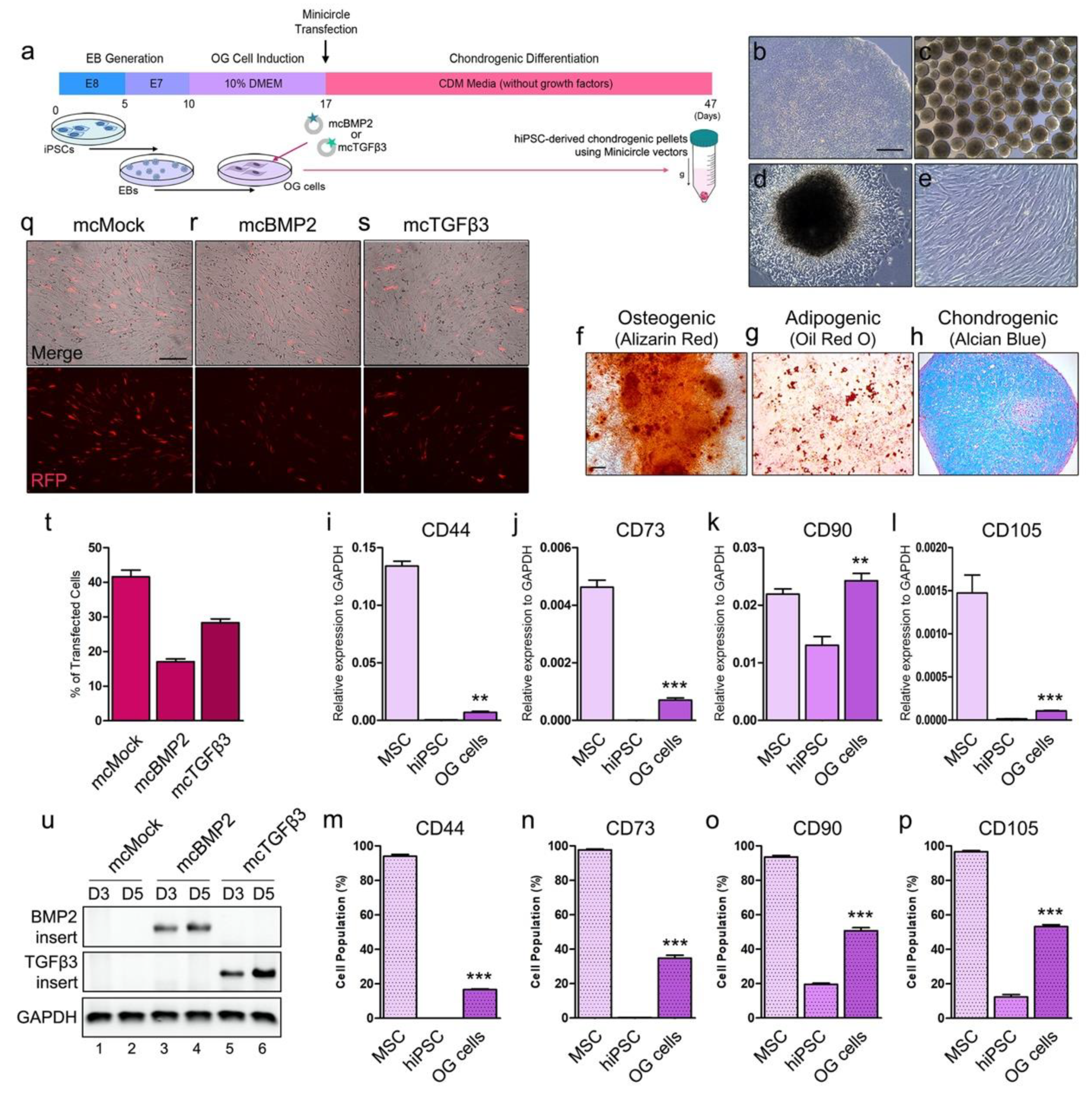
3.2. Chondrogenesis with Minicircles Using Human iPSC-Derived OG Cells
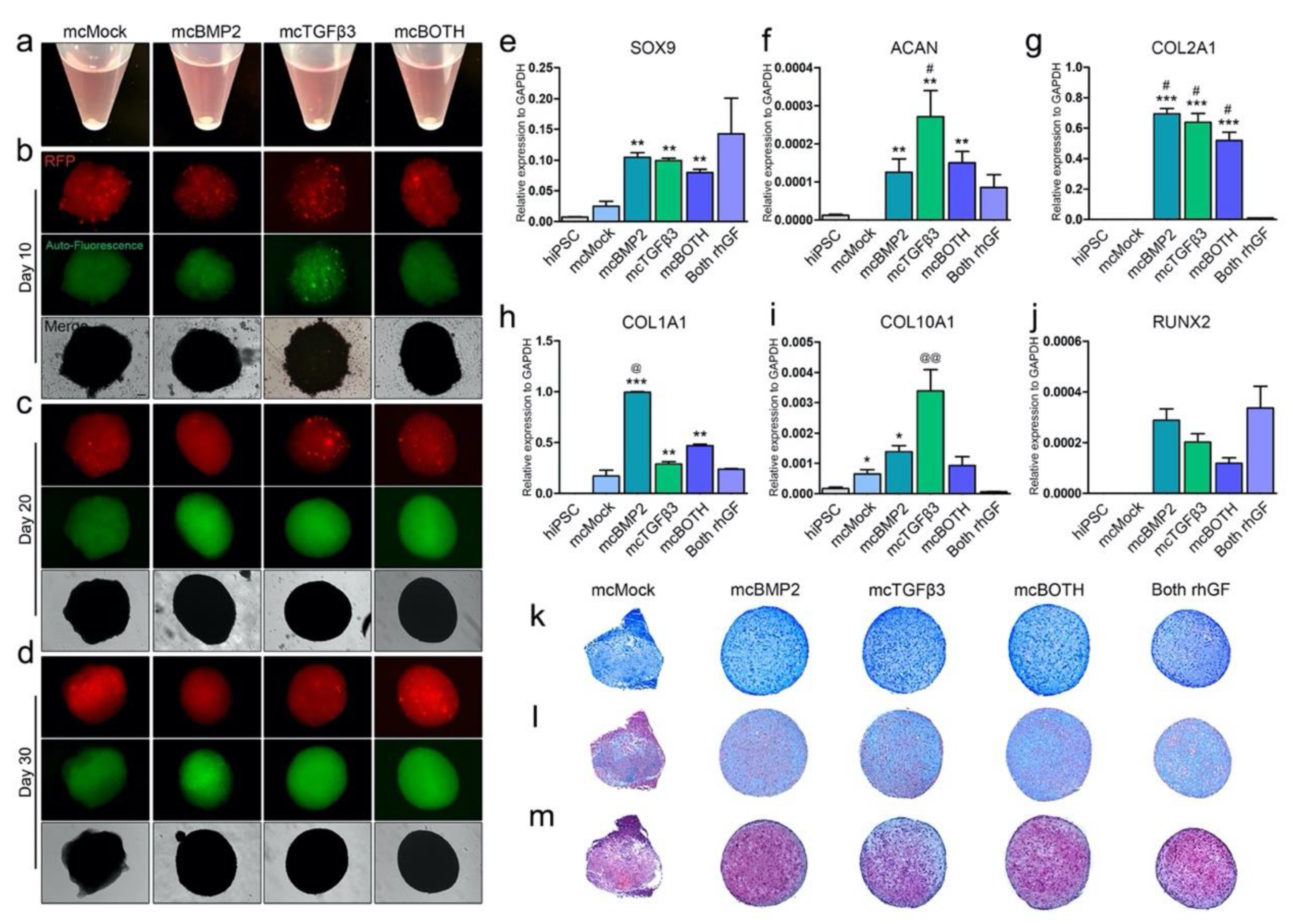
3.3. Characterization of Minicircle-Based Chondrogenic Pellets
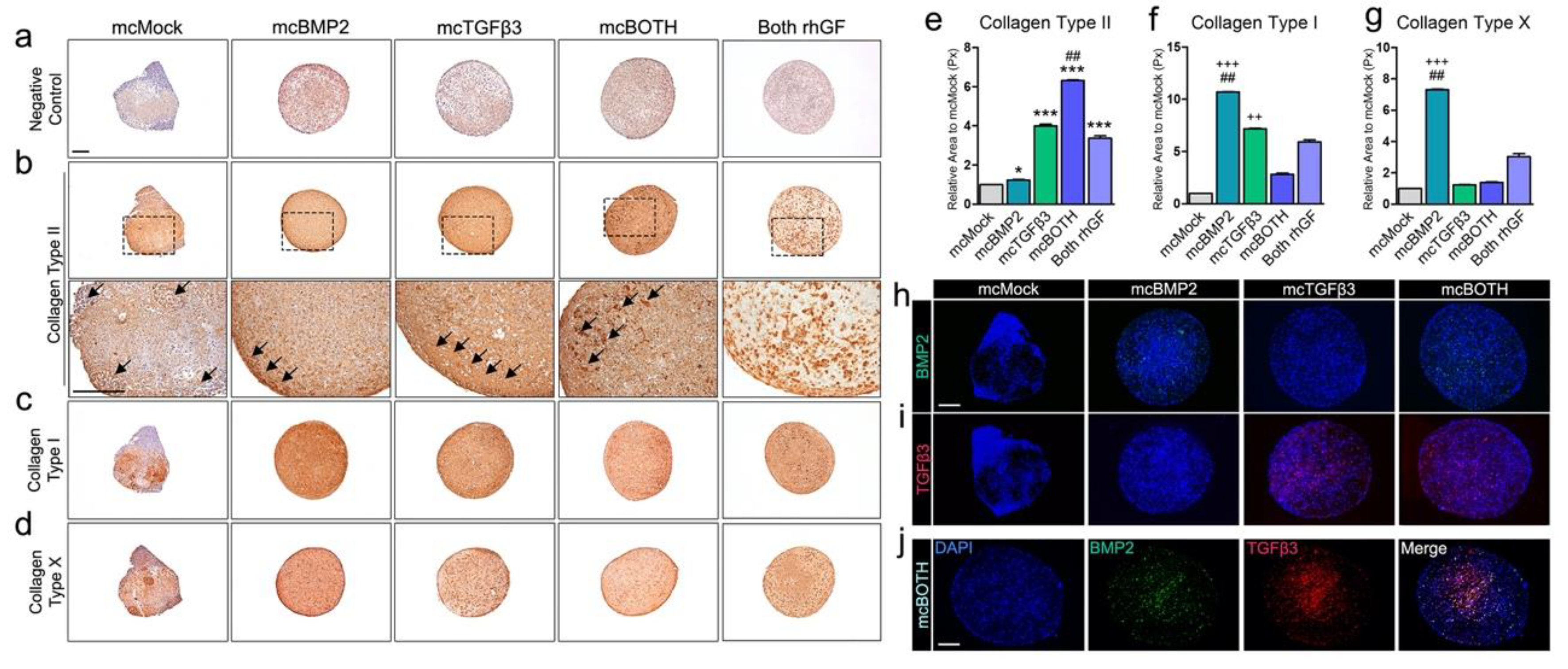
3.4. Further Analysis of Minicircle-Based Chondrogenic Pellets
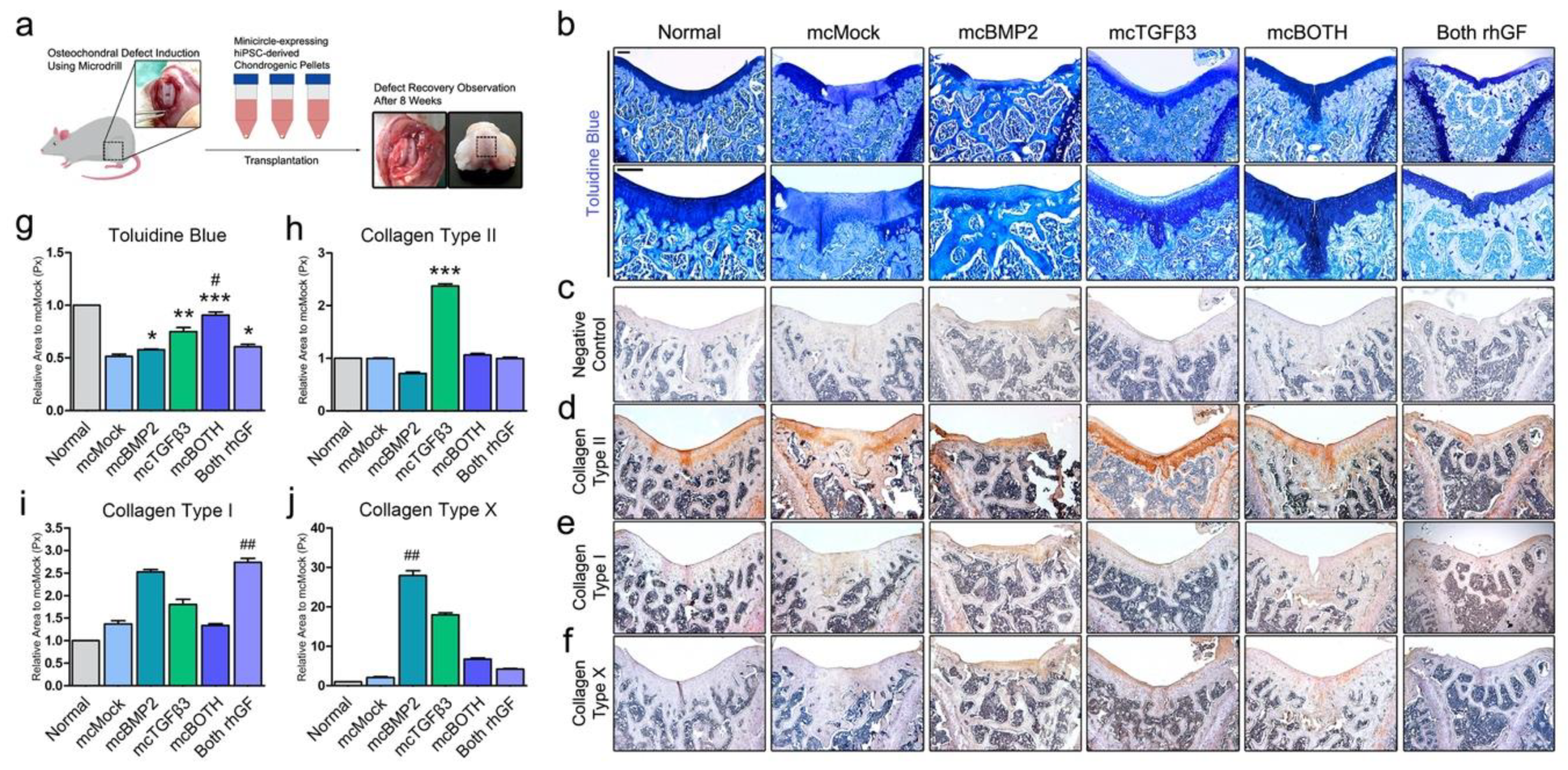
3.5. In Vivo Transplantation of Chondrogenic Pellets in Osteochondral Rat Model
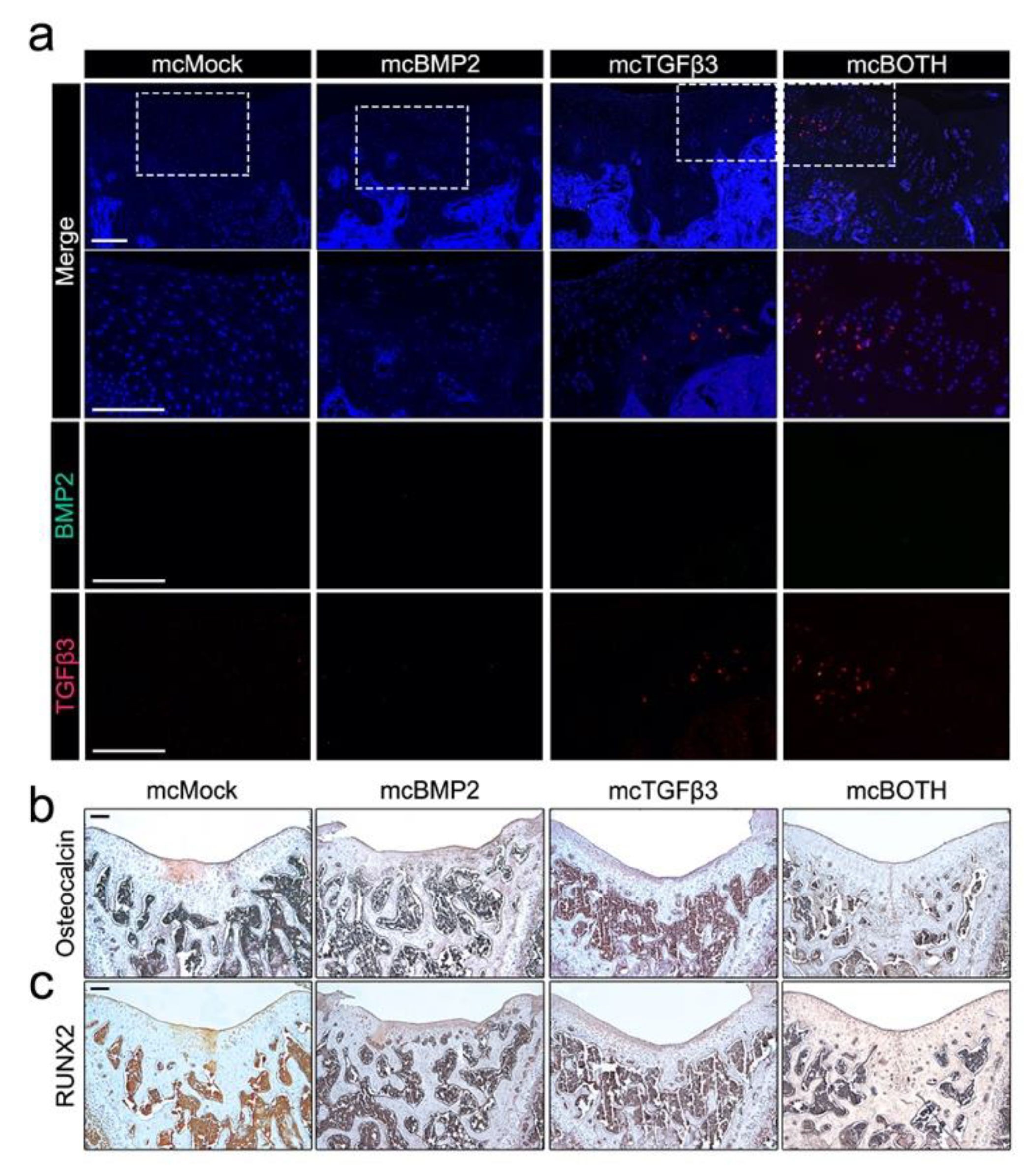
3.6. Further Analysis of Implanted Chondrogenic Pellets
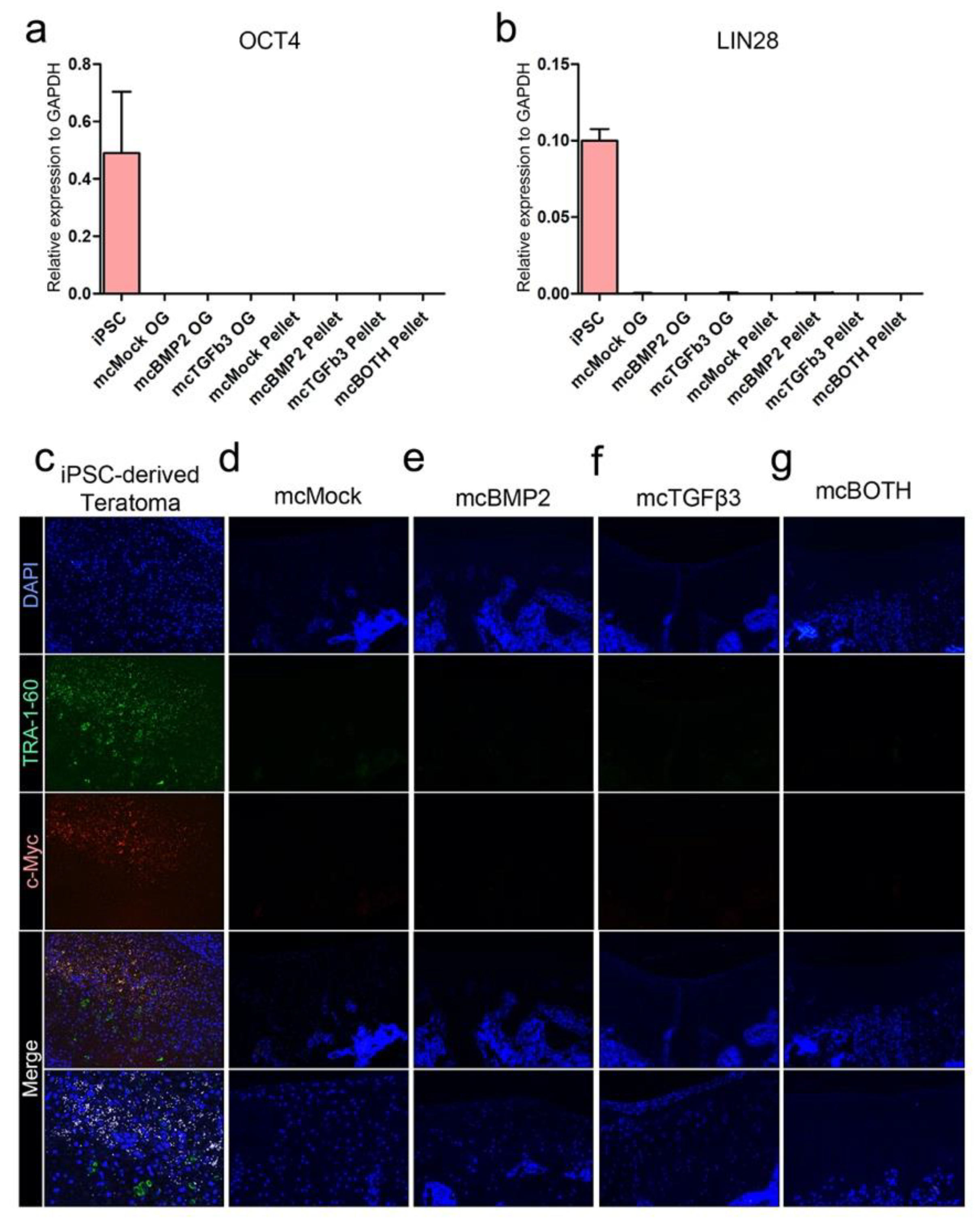
3.7. Further Analysis of Pluripotency and Tumorigenicity in Chondrogenic Pellets In Vitro and In Vivo
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Diekman, B.O.; Christoforou, N.; Willard, V.P.; Sun, H.; Sanchez-Adams, J.; Leong, K.W.; Guilak, F. Cartilage tissue engineering using differentiated and purified induced pluripotent stem cells. Proc. Natl. Acad. Sci. USA 2012, 109, 19172–19177. [Google Scholar] [CrossRef] [Green Version]
- Pittenger, M.F. Multilineage Potential of Adult Human Mesenchymal Stem Cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Diekman, B.O.; Rowland, C.; Lennon, N.P.; Caplan, A.; Guilak, F. Chondrogenesis of Adult Stem Cells from Adipose Tissue and Bone Marrow: Induction by Growth Factors and Cartilage-Derived Matrix. Tissue Eng. Part A 2009, 16, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Rim, Y.A.; Yi, H.; Park, N.; Park, S.-H.; Ju, J.H. The Generation of Human Induced Pluripotent Stem Cells from Blood Cells: An Efficient Protocol Using Serial Plating of Reprogrammed Cells by Centrifugation. Stem Cells Int. 2016, 2016, 1329459. [Google Scholar] [CrossRef] [PubMed]
- Nam, Y.; Rim, Y.A.; Jung, S.M.; Ju, J.H. Cord blood cell-derived iPSCs as a new candidate for chondrogenic differentiation and cartilage regeneration. Stem Cell Res. Ther. 2017, 8, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elizabeth, A.; Karin, P.; Ganna, B.; Bryant, S.J. Chondrogenic differentiation of human induced pluripotent stem cells in a photoclickable biomimetic PEG hydrogel. Front. Bioeng. Biotechnol. 2016, 4. [Google Scholar] [CrossRef]
- Majumdar, M.K.; Wang, E.; Morris, E.A. BMP-2 and BMP-9 promotes chondrogenic differentiation of human multipotential mesenchymal cells and overcomes the inhibitory effect of IL-1. J. Cell. Physiol. 2001, 189, 275–284. [Google Scholar] [CrossRef]
- Oshin, A.O.; Stewart, M. The role of bone morphogenetic proteins in articular cartilage development, homeostasis and repair. Veter- Comp. Orthop. Traumatol. 2007, 20, 151–158. [Google Scholar] [CrossRef]
- Shu, B.; Zhang, M.; Xie, R.; Wang, M.; Jin, H.; Hou, W.; Tang, D.; Harris, S.E.; Mishina, Y.; O’Keefe, R.J.; et al. BMP2, but not BMP4, is crucial for chondrocyte proliferation and maturation during endochondral bone development. J. Cell Sci. 2011, 124, 3428–3440. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, B. In VitroChondrogenesis of Bone Marrow-Derived Mesenchymal Progenitor Cells. Exp. Cell Res. 1998, 238, 265–272. [Google Scholar] [CrossRef]
- Mackay, A.M.; Beck, S.C.; Murphy, M.; Barry, F.P.; Chichester, C.O.; Pittenger, M.F. Chondrogenic Differentiation of Cultured Human Mesenchymal Stem Cells from Marrow. Tissue Eng. 1998, 4, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Tuli, R.; Tuli, S.; Nandi, S.; Huang, X.; Manner, P.A.; Hozack, W.J.; Danielson, K.G.; Hall, D.J.; Tuan, R.S. Transforming Growth Factor-β-mediated Chondrogenesis of Human Mesenchymal Progenitor Cells Involves N-cadherin and Mitogen-activated Protein Kinase and Wnt Signaling Cross-talk. J. Boil. Chem. 2003, 278, 41227–41236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelttari, K.; Winter, A.; Steck, E.; Goetzke, K.; Hennig, T.; Ochs, B.G.; Aigner, T.; Richter, W. Premature induction of hypertrophy during in vitro chondrogenesis of human mesenchymal stem cells correlates with calcification and vascular invasion after ectopic transplantation in SCID mice. Arthritis Rheum. 2006, 54, 3254–3266. [Google Scholar] [CrossRef] [PubMed]
- Mueller, M.B.; Tuan, R.S. Functional characterization of hypertrophy in chondrogenesis of human mesenchymal stem cells. Arthritis Rheum. 2008, 58, 1377–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, M.B.; Fischer, M.; Zellner, J.; Berner, A.; Dienstknecht, T.; Prantl, L.; Kujat, R.; Nerlich, M.; Tuan, R.S.; Angele, P. Hypertrophy in mesenchymal stem cell chondrogenesis: Effect of TGF-beta isoforms and chondrogenic conditioning. Cells Tissues Organs 2010, 192, 158–166. [Google Scholar] [CrossRef] [Green Version]
- Barry, F.P.; Boynton, R.E.; Liu, B.; Murphy, M. Chondrogenic Differentiation of Mesenchymal Stem Cells from Bone Marrow: Differentiation-Dependent Gene Expression of Matrix Components. Exp. Cell Res. 2001, 268, 189–200. [Google Scholar] [CrossRef]
- Shen, B.; Wei, A.; Tao, H.; Diwan, A.; Ma, D. BMP-2 Enhances TGF-β3–Mediated Chondrogenic Differentiation of Human Bone Marrow Multipotent Mesenchymal Stromal Cells in Alginate Bead Culture. Tissue Eng. Part A 2009, 15, 1311–1320. [Google Scholar] [CrossRef]
- Kim, Y.I.; Ryu, J.-S.; Yeo, J.E.; Choi, Y.-J.; Kim, Y.S.; Ko, K.; Koh, Y.-G. Overexpression of TGF-β1 enhances chondrogenic differentiation and proliferation of human synovium-derived stem cells. Biochem. Biophys. Res. Commun. 2014, 450, 1593–1599. [Google Scholar] [CrossRef]
- Tsuchiya, H.; Kitoh, H.; Sugiura, F.; Ishiguro, N. Chondrogenesis enhanced by overexpression of sox9 gene in mouse bone marrow-derived mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2003, 301, 338–343. [Google Scholar] [CrossRef]
- Wang, Z.; Li, X.; He, X.; Wu, B.; Xu, M.; Chang, H.; Zhang, X.; Xing, Z.; Jing, X.; Kong, D.; et al. Delivery of the Sox9 gene promotes chondrogenic differentiation of human umbilical cord blood-derived mesenchymal stem cells in an in vitro model. Braz. J. Med Boil. Res. 2014, 47, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Do, H.-J.; Yang, H.-M.; Oh, J.-H.; Choi, S.-J.; Kim, D.-K.; Cha, K.-Y.; Chung, H.-M. Overexpression of SOX9 in mouse embryonic stem cells directs the immediate chondrogenic commitment. Exp. Mol. Med. 2005, 37, 261–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, M.; Chen, Z.; Hu, S.; Jia, F.; Li, Z.; Hoyt, G.; Robbins, R.C.; Kay, M.A.; Wu, J.C. Novel minicircle vector for gene therapy in murine myocardial infarction. Circulation 2009, 120, S230–S237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jechlinger, W. Optimization and delivery of plasmid DNA for vaccination. Expert Rev. Vaccines 2006, 5, 803–825. [Google Scholar] [CrossRef] [PubMed]
- Gill, D.; A Pringle, I.; Hyde, S. Progress and Prospects: The design and production of plasmid vectors. Gene Ther. 2009, 16, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Bandara, N.; Gurusinghe, S.; Chen, S.; Chen, H.; Wang, L.; Lim, S.Y.; Strappe, P. Minicircle DNA-mediated endothelial nitric oxide synthase gene transfer enhances angiogenic responses of bone marrow-derived mesenchymal stem cells. Stem Cell Res. Ther. 2016, 7, 48. [Google Scholar] [CrossRef] [Green Version]
- Park, N.; Rim, Y.A.; Jung, H.; Kim, J.; Yi, H.; Kim, Y.; Jang, Y.; Jung, S.M.; Lee, J.; Kwok, S.-K.; et al. Etanercept-Synthesising Mesenchymal Stem Cells Efficiently Ameliorate Collagen-Induced Arthritis. Sci. Rep. 2017, 7, 39593. [Google Scholar] [CrossRef] [Green Version]
- Rim, Y.A.; Yi, H.; Kim, Y.; Park, N.; Jung, H.; Kim, J.; Jung, S.M.; Park, S.-H.; Ju, J.H. Self in vivo production of a synthetic biological drug CTLA4Ig using a minicircle vector. Sci. Rep. 2014, 4, 6935. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.; Kim, Y.; Kim, J.; Jung, H.; Rim, Y.A.; Jung, S.M.; Park, S.-H.; Ju, J.H. A New Strategy to Deliver Synthetic Protein Drugs: Self-reproducible Biologics Using Minicircles. Sci. Rep. 2014, 4, 5961. [Google Scholar] [CrossRef] [Green Version]
- Solchaga, L.A.; Penick, K.; Goldberg, V.M.; Caplan, A.; Welter, J.F. Fibroblast Growth Factor-2 Enhances Proliferation and Delays Loss of Chondrogenic Potential in Human Adult Bone-Marrow-Derived Mesenchymal Stem Cells. Tissue Eng. Part A 2009, 16, 1009–1019. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Tian, X.; Yuan, Y.; Song, Z.; Zhang, L.; Wang, X.; Li, T. Effect of cell culture using chitosan membranes on stemness marker genes in mesenchymal stem cells. Mol. Med. Rep. 2013, 7, 1945–1949. [Google Scholar] [CrossRef]
- Jia, F.; Wilson, K.D.; Sun, N.; Gupta, D.M.; Huang, M.; Li, Z.; Panetta, N.J.; Chen, Z.Y.; Robbins, R.C.; Kay, M.A.; et al. A nonviral minicircle vector for deriving human iPS cells. Nat. Methods 2010, 7, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.-Y.; He, C.-Y.; Ehrhardt, A.; A Kay, M. Minicircle DNA vectors devoid of bacterial DNA result in persistent and high-level transgene expression in vivo. Mol. Ther. 2003, 8, 495–500. [Google Scholar] [CrossRef]
- Stenler, S.; Blomberg, P.; Smith, C.E. Safety and efficacy of DNA vaccines: Plasmids vs. minicircles. Hum. Vaccines Immunother. 2014, 10, 1306–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hautier, A.; Salentey, V.; Aubert-Foucher, E.; Bougault, C.; Beauchef, G.; Ronziere, M.C.; De Sobarnitsky, S.; Paumier, A.; Galera, P.; Piperno, M.; et al. Bone morphogenetic protein-2 stimulates chondrogenic expression in human nasal chondrocytes expanded in vitro. Growth Factors 2008, 26, 201–211. [Google Scholar] [CrossRef]
- James, A.W.; Xu, Y.; Lee, J.K.; Wang, R.; Longaker, M.T. Differential Effects of TGF-β1 and TGF-β3 on Chondrogenesis in Posterofrontal Cranial Suture–Derived Mesenchymal Cells In Vitro. Plast. Reconstr. Surg. 2009, 123, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Teramura, T.; Onodera, Y.; Mihara, T.; Hosoi, Y.; Hamanishi, C.; Fukuda, K. Induction of Mesenchymal Progenitor Cells with Chondrogenic Property from Mouse-Induced Pluripotent Stem Cells. Cell. Reprogramming 2010, 12, 249–261. [Google Scholar] [CrossRef]
- Koyama, N.; Miura, M.; Nakao, K.; Kondo, E.; Fujii, T.; Taura, D.; Kanamoto, N.; Sone, M.; Yasoda, A.; Arai, H.; et al. Human Induced Pluripotent Stem Cells Differentiated into Chondrogenic LineageViaGeneration of Mesenchymal Progenitor Cells. Stem Cells Dev. 2013, 22, 102–113. [Google Scholar] [CrossRef]
- Kim, K.; Doi, A.; Wen, B.; Ng, K.; Zhao, R.; Cahan, P.; Kim, J.; Aryee, M.J.; Ji, H.; Ehrlich, L.I.R.; et al. Epigenetic memory in induced pluripotent stem cells. Nature 2010, 467, 285–290. [Google Scholar] [CrossRef] [Green Version]
- Phetfong, J.; Supokawej, A.; Wattanapanitch, M.; Kheolamai, P.; U-Pratya, Y.; Issaragrisil, S. Cell type of origin influences iPSC generation and differentiation to cells of the hematoendothelial lineage. Cell and Tissue Res. 2016, 365, 101–112. [Google Scholar] [CrossRef]
- Kang, R.; Zhou, Y.; Tan, S.; Zhou, G.; Aagaard, L.; Xie, L.; Bünger, C.; Bolund, L.; Luo, Y. Mesenchymal stem cells derived from human induced pluripotent stem cells retain adequate osteogenicity and chondrogenicity but less adipogenicity. Stem Cell Res. Ther. 2015, 6, 144. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.S.; Pelekanos, R.A.; Ellis, R.L.; Horne, R.; Wolvetang, E.J.; Fisk, N.M. Small Molecule Mesengenic Induction of Human Induced Pluripotent Stem Cells to Generate Mesenchymal Stem/Stromal Cells. Stem Cells Transl. Med. 2012, 1, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Gregory, C.A.; Lee, R.H.; Reger, R.L.; Qin, L.; Hai, B.; Park, M.S.; Yoon, N.; Clough, B.; McNeill, E.; et al. MSCs derived from iPSCs with a modified protocol are tumor-tropic but have much less potential to promote tumors than bone marrow MSCs. Proc. Natl. Acad. Sci. 2014, 112, 530–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyd, N.L.; Robbins, K.; Dhara, S.K.; West, F.D.; Stice, S.L. Human Embryonic Stem Cell–Derived Mesoderm-like Epithelium Transitions to Mesenchymal Progenitor Cells. Tissue Eng. Part A 2009, 15, 1897–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aluigi, M.; Fogli, M.; Curti, A.; Isidori, A.; Gruppioni, E.; Chiodoni, C.; Colombo, M.P.; Versura, P.; D’Errico-Grigioni, A.; Ferri, E.; et al. Nucleofection Is an Efficient Nonviral Transfection Technique for Human Bone Marrow-Derived Mesenchymal Stem Cells. Stem Cells 2006, 24, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Zaragosi, L.-E.; Billon, N.; Ailhaud, G.; Dani, C. Nucleofection Is a Valuable Transfection Method for Transient and Stable Transgene Expression in Adipose Tissue-Derived Stem Cells. Stem Cells 2006, 25, 790–797. [Google Scholar] [CrossRef]
- Pellaud, J.; Schote, U.; Arvinte, T.; Seelig, J. Conformation and self-association of human recombinant transforming growth factor-beta3 in aqueous solutions. J. Boil. Chem. 1999, 274, 274. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Human BMP2 |
| CTCGTTCCCGAGCTTGGTCGGAGGAAGTTTGCGGCCGCGTCAAGCGGAAGGCCCAGTAGTCAGCCT AGCGACGAGGTCCTTTCTGAATTCGAGCTTCGGCTCCTGTCCATGTTTGGACTTAAACAGCGACCTA CGCCCAGCCGGGATGCCGTTGTACCGCCCTATATGCTCGATCTTTATCGAAGACATTCCGGTCAGCCA GGATCACCGGCTCCAGATCATAGACTTGAGCGCGCTGCCTCCCGGGCAAACACTGTGCGATCCTTTC ACCATGAGGAATCACTGGAAGAATTGCCAGAAACTTCAGGTAAGACTACGAGACGATTCTTTTTTAA TCTCTCATCCATTCCTACAGAAGAATTCATTACGTCTGCCGAGCTTCAGGTATTCAGAGAACAGATGC AAGATGCTTTGGGGAATAACAGCAGCTTTCACCATCGCATCAACATATACGAGATAATCAAACCCGC AACAGCCAACAGCAAATTTCCCGTAACGCGATTGCTGGATACGCGACTTGTGAACCAAAACGCTAG CAGATGGGAATCATTCGATGTGACGCCCGCGGTCATGAGATGGACCGCTCAGGGCCACGCGAATCA CGGCTTTGTTGTAGAGGTGGCACATCTTGAAGAGAAGCAAGGTGTCAGCAAAAGACATGTACGAAT AAGTCGATCACTCCATCAAGATGAACACTCATGGAGCCAAATAAGACCTCTCCTTGTGACATTCGGG CATGACGGAAAGGGTCACCCTCTTCACAAAAGGGAGAAGCGCCAGGCGAAGCATAAACAGCGGA AACGCCTTAAGTCAAGTTGCAAACGCCATCCTTTGTACGTCGATTTCTCCGATGTTGGATGGAAT GATTGGATCGTAGCTCCTCCTGGATACCATGCCTTCTATTGCCATGGCGAGTGCCCGTTCCCTCTT GCGGATCATCTCAACAGTACCAATCATGCAATCGTGCAAACCCTTGTAAACAGCGTCAACTCCAA AATTCCCAAGGCTTGTTGCGTTCCTACTGAGCTGAGCGCCATAAGTATGCTGTACCTCGATGAAA ATGAAAAAGTTGTCCTGAAGAATTATCAAGATATGGTGGTAGAAGGTTGTGGATGTAGG |
| Human TGFβ3 |
| TTGTCCACCTGTACTACTTTGGATTTTGGTCACATAAAAAAAAAACGGGTCGAGGCAATCCGAGGGC AAATTCTCAGCAAACTGAGGCTTACATCACCCCCCGAACCGACCGTTATGACCCACGTACCATATCA GGTCTTGGCTCTGTATAACTCTACTCGCGAACTGCTTGAGGAGATGCATGGGGAAAGAGAGGAGGG TTGTACCCAAGAGAATACCGAAAGCGAGTACTATGCTAAGGAGATTCATAAATTCGATATGATTCAG GGTCTGGCAGAGCACAACGAGCTGGCAGTGTGTCCAAAAGGAATCACCTCAAAGGTGTTTCGCTTC AATGTATCCAGCGTCGAAAAGAATCGCACCAACCTCTTCCGAGCGGAGTTTAGGGTTCTTCGGGTAC CAAACCCTAGCTCAAAGCGAAATGAGCAACGCATTGAGTTGTTCCAGATACTTAGGCCGGATGAAC ACATTGCGAAGCAGAGGTATATAGGTGGTAAAAACCTCCCGACTCGGGGTACTGCGGAGTGGCTCTC ATTTGATGTCACCGACACAGTACGCGAATGGCTTCTGCGAAGAGAGAGCAATCTTGGACTTGAAATC AGTATCCACTGTCCTTGTCATACCTTCCAACCGAATGGAGATATACTGGAGAACATCCACGAGGTAAT GGAAATTAAGTTTAAAGGCGTGGACAACGAAGATGATCACGGTCGGGGTGATCTGGGACGACTGAA GAAACAAAAAGACCACCATAACCCGCATCTGATCCTTATGATGATCCCCCCGCATAGACTCGACAAC CCAGGTCAAGGCGGGCAGAGAAAGAAAAGAGCTCTGGATACTAACTACTGTTTTAGGAATCTGG AAGAAAACTGCTGCGTACGACCCTTGTATATTGATTTTAGACAAGACCTCGGTTGGAAATGGGT CCACGAACCAAAGGGATACTATGCCAATTTCTGTAGCGGCCCTTGTCCCTACTTGAGGAGTGCC GACACTACACATTCTACTGTGCTCGGTTTGTATAACACCTTGAACCCAGAAGCTAGTGCATCTCC CTGCTGCGTTCCCCAGGATCTCGAACCCCTCACTATTTTGTATTACGTTGGTCGGACACCAAAAG TCGAACAACTTTCAAACATGGTCGTGAAGTCCTGTAAGTGCAGC |
| Gene Name | Sequences (5′ → 3′) | Length (bp) |
|---|---|---|
| CD44 | F: AAGGTGGAGCAAACACAACC | 151 |
| R: AGCTTTTTCTTCTGCCCACA | ||
| CD73 | F: CCAATTCTGAGTGCAAACAT | 315 |
| R: CCTCCCACCACGACGTCCAC | ||
| CD90 | F: CTAGTGGACCAGAGCCTTCG | 236 |
| R: TGGAGTGCACACGTGTAGGT | ||
| CD105 | F: CACTAGCCAGGTCTCGAAGG | 165 |
| R: CTGAGGACCAGAAGCACCTC | ||
| BMP2 insert (Active domain) | F: CAGGCGAAGCATAAACAGCG | 342 |
| R: CCTACATCCACAACCTTCTACC | ||
| TGFβ3 insert (Active domain) | F: CTGCTGCGTACGACCCTTG | 295 |
| R: GCTGCACTTACAGGACTTCACG | ||
| SOX9 | F: GACTTCCGCGACGTGGAC | 99 |
| R: GTTGGGCGGCAGGTACTG | ||
| ACAN | F: TCGAGGACAGCGAGGCC | 85 |
| R: TCGAGGGTGTAGCGTGTAGAGA | ||
| COL2A1 | F: GGCAATAGCAGGTTCACGTACA | 79 |
| R: CGATAACAGTCTTGCCCCACTTA | ||
| COL1A1 | F: TCTGCGACAACGGCAAGGTG | 146 |
| R: GACGCCGGTGGTTTCTTGGT | ||
| COL10A1 | F: CAGGCATAAAAGGCCCAC | 108 |
| R: GTGGACCAGGAGTACCTTGC | ||
| RUNX2 | F: CTCTACCACCCCGCTGTCTT | 143 |
| R: CACCTGCCTGGCTCTTCTTAC | ||
| OCT4 | F: GACAGGGGGAGGGGAGGAGCTAGG | 144 |
| R: CTTCCCTCCAACCAGTTGCCCCAAAC | ||
| LIN28 | F: GTTCGGCTTCCTGTCCAT | 121 |
| R: CTGCCTCACCCTCCTTCA | ||
| GAPDH | F: ACCCACTCCTCCACCTTTGA | 101 |
| R: CTGTTGCTGTAGCCAAATTCGT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rim, Y.A.; Nam, Y.; Park, N.; Jung, H.; Lee, K.; Lee, J.; Ju, J.H. Chondrogenic Differentiation from Induced Pluripotent Stem Cells Using Non-Viral Minicircle Vectors. Cells 2020, 9, 582. https://doi.org/10.3390/cells9030582
Rim YA, Nam Y, Park N, Jung H, Lee K, Lee J, Ju JH. Chondrogenic Differentiation from Induced Pluripotent Stem Cells Using Non-Viral Minicircle Vectors. Cells. 2020; 9(3):582. https://doi.org/10.3390/cells9030582
Chicago/Turabian StyleRim, Yeri Alice, Yoojun Nam, Narae Park, Hyerin Jung, Kijun Lee, Jennifer Lee, and Ji Hyeon Ju. 2020. "Chondrogenic Differentiation from Induced Pluripotent Stem Cells Using Non-Viral Minicircle Vectors" Cells 9, no. 3: 582. https://doi.org/10.3390/cells9030582
APA StyleRim, Y. A., Nam, Y., Park, N., Jung, H., Lee, K., Lee, J., & Ju, J. H. (2020). Chondrogenic Differentiation from Induced Pluripotent Stem Cells Using Non-Viral Minicircle Vectors. Cells, 9(3), 582. https://doi.org/10.3390/cells9030582