1. Introduction
The nuclear envelope (NE) separates the genetic material in the nucleus from the cytoplasm and all its organelles. It comprises the outer nuclear membrane (ONM), which is continuous with the membrane system of the endoplasmic reticulum (ER), the inner nuclear membrane (INM) and the nuclear pore complexes (NPC), where INM and ONM are connected. In metazoans, a nuclear lamina is associated with the nuclear face of the NE, with characteristic intermediate filament proteins (lamins) tightly binding to INM-proteins. The protein composition of the INM is clearly distinct from that of the ONM and in recent years a large number of INM-specific proteins have been identified [
1,
2,
3]. Emerin, loss of which causes X-linked Emery-Dreifuss muscular dystrophy [
4,
5,
6,
7,
8], is one of the best-characterized proteins of the INM [
9,
10,
11], although it was also described as a component of the ONM and the peripheral ER [
12]. It is a tail-anchored protein with a single transmembrane domain close to its C-terminal end and we recently showed that emerin is inserted post-translationally into ER-membranes via the GET/TRC40 pathway [
13]. Emerin has an N-terminal LEM-domain, which received its name from three proteins containing this characteristic feature (Lap2, emerin and Man1; [
14]). Major binding partners of emerin are A- and also B-type lamins [
6]. Indeed, the localization of emerin at the nuclear envelope was shown to depend on lamin A [
15]. Hence, retention of emerin upon binding to lamins plays an important role in INM-targeting. Besides lamins, a number of emerin interaction partners have been described, for example BAF (barrier-to-autointegration factor (BANF1) [
16], nesprin-1α [
17] and HDAC3 [
18]; for review see [
9]).
For proteins of the INM, the identification of interacting proteins is particularly challenging, because conditions that are typically used in, for example, co-immunoprecipitation approaches, may not lead to complete solubilization of NE-structures. Genetic approaches like the yeast-two-hybrid method, on the other hand, detect interactions under rather non-physiological conditions. Neither approach addresses the native subcellular localization of a protein of interest. To overcome these problems, several methods have been developed that use proximity-based labeling, typically introducing biotin into unknown proteins, allowing subsequent affinity capture with immobilized streptavidin and analysis by mass-spectrometry (for review see [
19]). One approach termed BioID uses a biotin ligase from
E. coli for biotinylation of proteins [
20]. As an alternative, ascorbate peroxidase (APEX), an enzyme that generates radicals from biotin phenol in the presence of H
2O
2 was introduced [
21]. Modification of proteins occurs within a range of ~20 nm of the enzyme, which can be genetically fused to the N- or C-terminus of proteins of interest for probing their direct environment and identifying potential interaction partners [
22,
23,
24,
25]. An improved version of APEX, APEX2, is far more active than the original enzyme [
26]. Very recently, we devised a method where APEX2 is not directly fused, but instead targeted to a protein of interest, in a rapamycin-dependent manner [
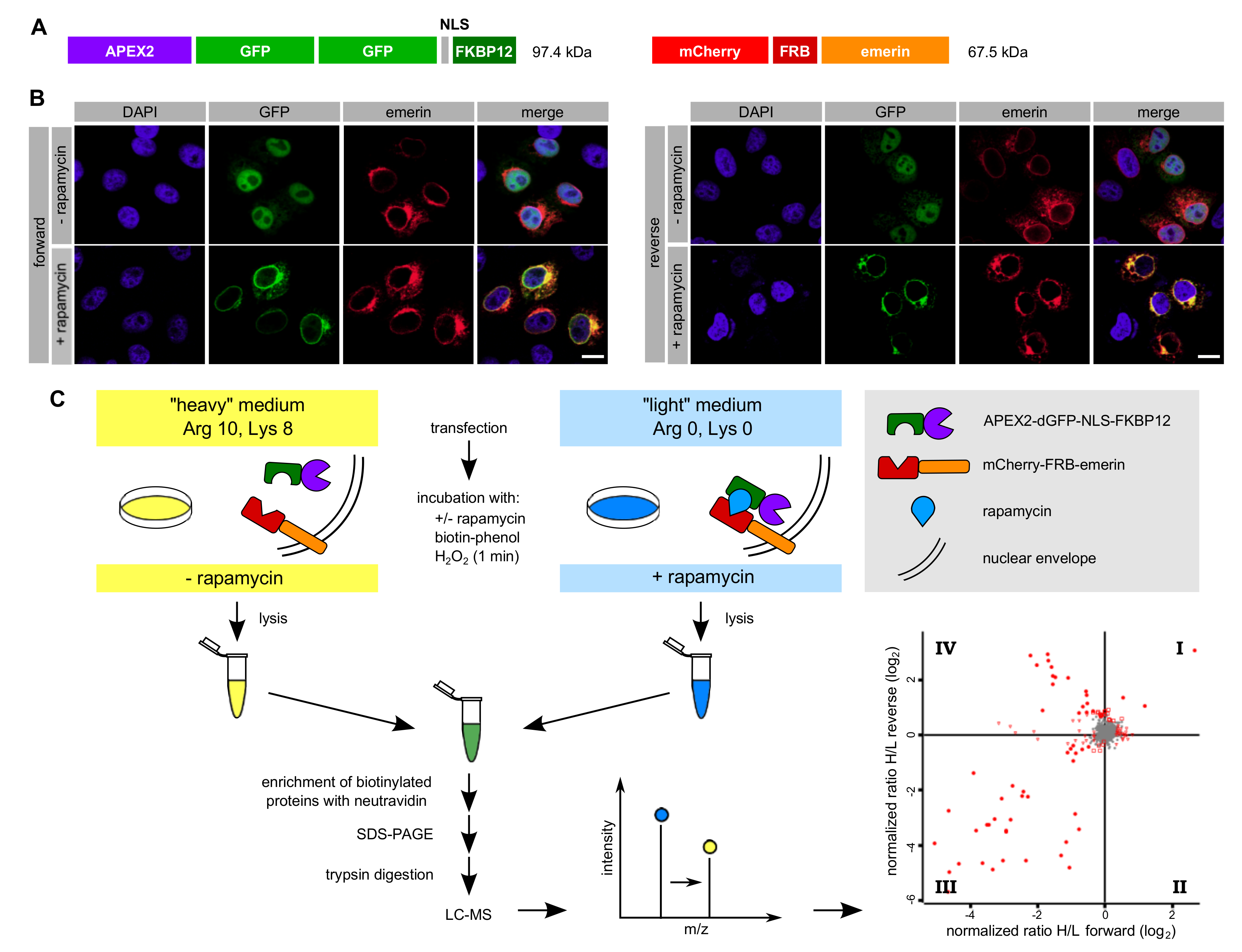
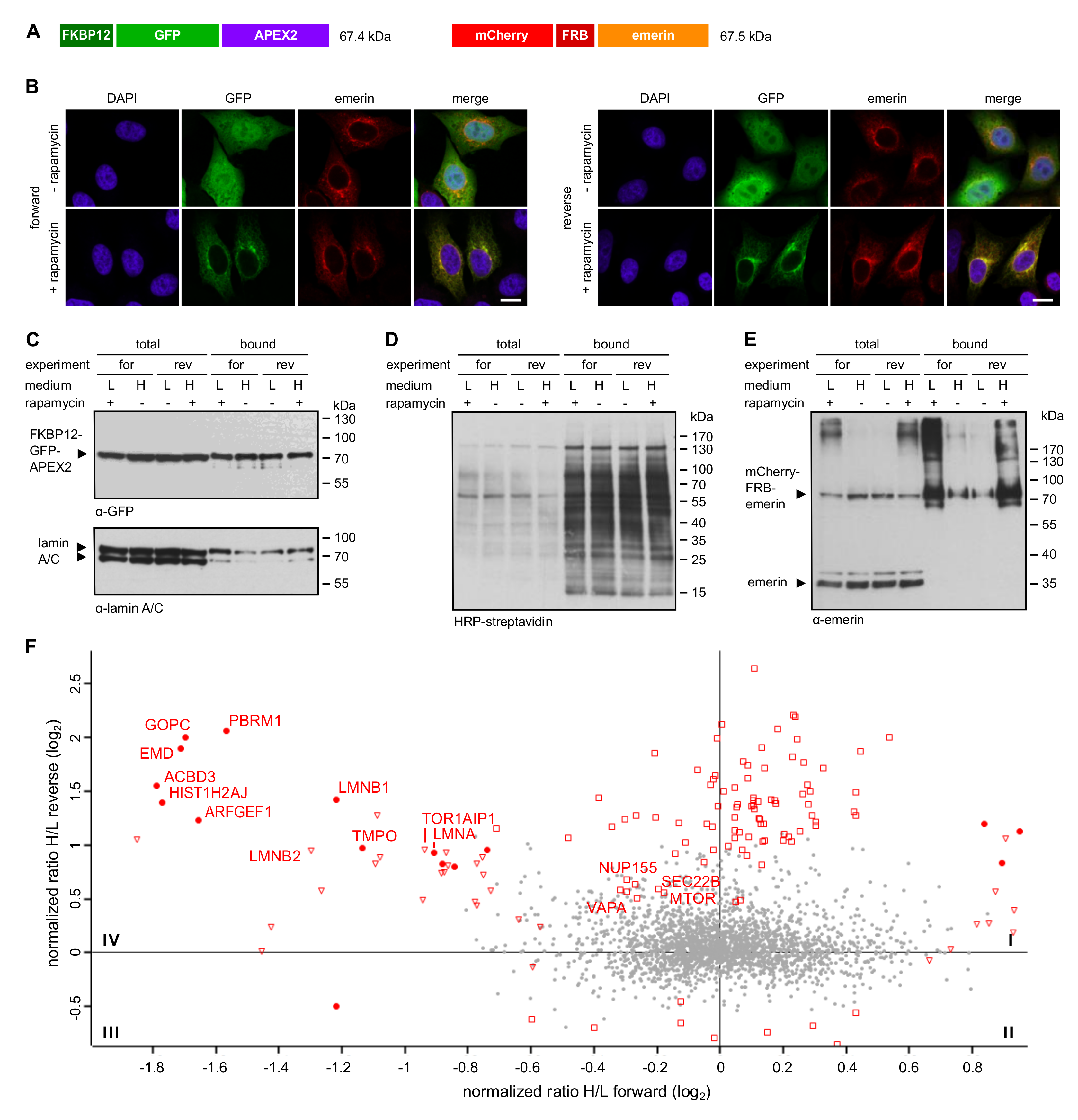
27]. The method utilizes FKBP12-tagged APEX2, which interacts with an FRB-tagged protein of interest upon addition of rapamycin to intact HeLa cells for a short period of time. SILAC, followed by quantitative mass spectrometry, then allows the comparison of proteins that get biotinylated by APEX2 in either the absence or the presence of rapamycin. Using this method (Rapamycin- and APEX-dependent identification of proteins by SILAC; RAPIDS), we identified lamin A and several other proteins that directly interact with or are in close proximity to emerin.
2. Materials and Methods
2.1. Plasmids
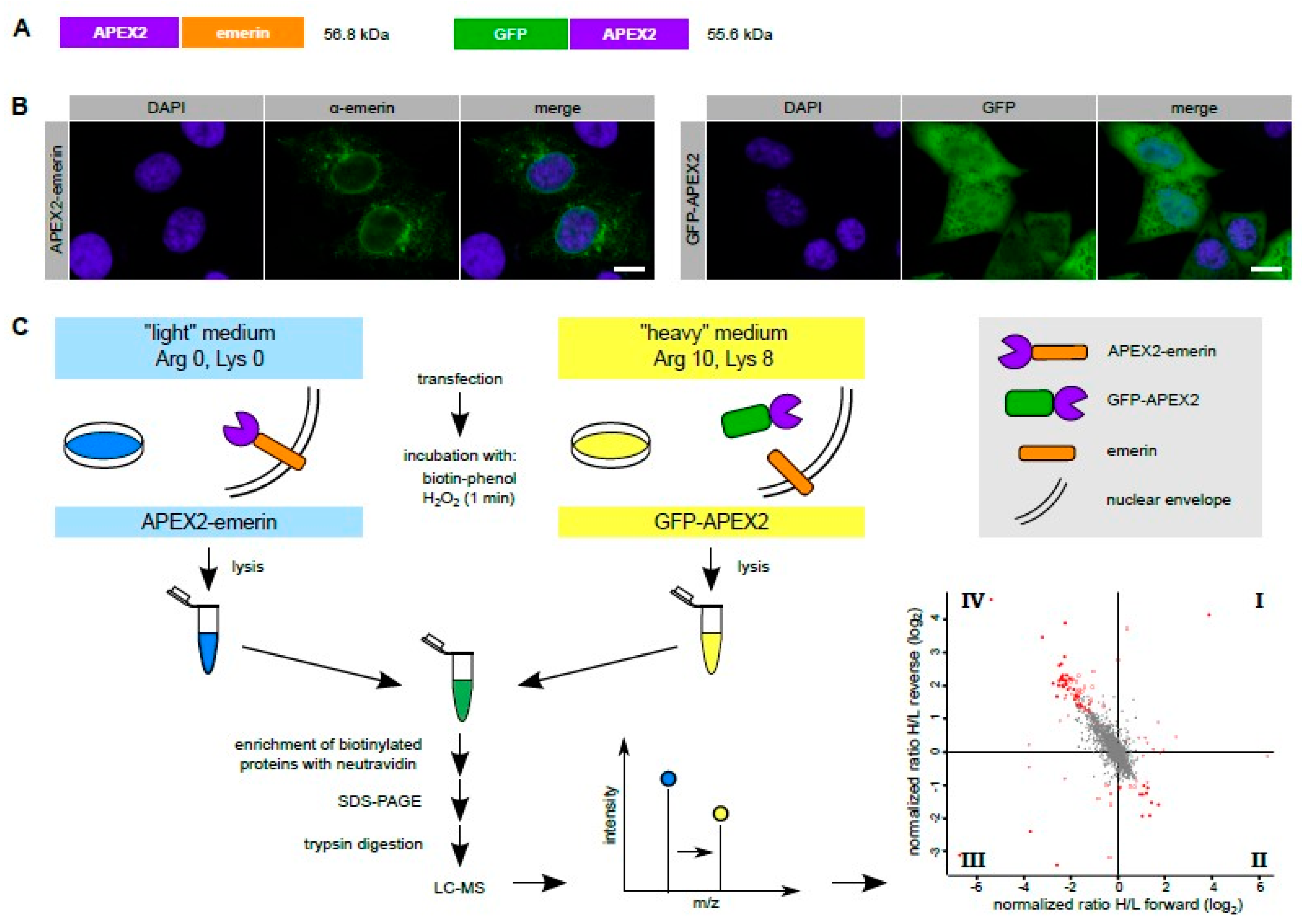
All plasmids were cloned using standard procedures and were verified by sequencing. Plasmids coding for APEX2-dGFP-NLS-FKBP12 and FKBP12-GFP-APEX2 were described before [
27]. pcDNA3-FKBP12-GFP-APEX2 was used as a template to amplify the APEX2 coding sequence by using primers G1852 (5′-GCGCTAGCGTCGCCACCATGGGAAAGTCTTACCCAACTGTGAG) and G1853 (5′-GCCTCGAGAACCAGAAGCTCCTGAGGCATCAGCAAACCCAAGCTCG) or G1854 (5′-GCGAATTCTGGAAAGTCTTACCCAACTGTGAG) and G1855 (5′-GCGGATCCGGCATCAGCAAACCCAAGCTC). The PCR products were cloned into a pmCherry-C1 derivative, encoding mCherry-emerin and into pEGFP-C1 (Clontech Laboratories, Mountain View, CA, USA), generating pAPEX2-emerin and pGFP-APEX2, respectively. Cloning of pmCherry-FRB-emerin was described previously [
13]. All fusion proteins contained the full-length sequence of emerin or APEX2, respectively, with short linker sequences as determined by the restriction sites.
2.2. Cell Culture and Transfection
HeLa P4 cells [
28] were obtained from the NIH AIDS Reagent Program and grown in DMEM (Life technologies, Carlsbad, CA, USA) supplemented with 10% (v/v) FBS (Life technologies, Carlsbad, CA, USA), 100 U mL
−1 penicillin, 100 µg mL
−1 streptomycin and 2 mM L-glutamine (Life technologies, Carlsbad, CA, USA) under 5% CO
2 at 37°C. Cells were tested for mycoplasma contamination on a regular basis.
For SILAC, HeLa cells were labeled with heavy or light isotopes of arginine and lysine. For this purpose, DMEM (high glucose) lacking glutamine, lysine and arginine (Thermo Fisher Scientific, Waltham, MA, USA) was supplemented with 10% (v/v) dialyzed FBS (Life technologies, Carlsbad, CA, USA), 6 mM L-glutamine (Life technologies), 100 U mL
−1 penicillin and 100 µg mL
−1 streptomycin. SILAC media was prepared as described [
27], using
13C
615N
2-L-lysine (Silantes, Munich, Germany) and
13C
615N
4-L-arginine (Silantes, Munich, Germany) or
12C
614N
2-L-lysine (Sigma-Aldrich, St. Louis, MO, USA) and
12C
614N
4-L-arginine (Sigma-Aldrich, St. Louis, MO, USA) as heavy or light amino acids, respectively. To ensure sufficient incorporation of heavy amino acids, cells were passaged five to seven times in SILAC medium before the biotinylation experiment. The incorporation rate was confirmed to be ≥97% by mass spectrometry.
Cells were transfected using the calcium phosphate method [
27,
29].
2.3. Rapamycin-Dependent Biotinylation Assay
Cells grown in SILAC medium were subjected to biotinylation reactions as described [
27]. The biotinylation experiment using the FKBP12-GFP-APEX2 construct was performed in 10 cm dishes. The cells were transfected with pmCherry-FRB-emerin and pcDNA3-FKBP12-GFP-APEX2 and grown to confluency. Cells were then incubated for 30 min with 500 µM biotin-phenol (Iris Biotech, Marktredwitz, Germany), in the presence or absence of 200 nM rapamycin (Sigma Aldrich, St. Louis, MO, USA). Experiments were performed in both forward and reverse conditions. For forward reactions, cells grown in “light” SILAC medium were treated with rapamycin and cells grown in “heavy” SILAC medium were not. For reverse reactions, this labeling scheme was switched. After incubation with biotin-phenol and rapamycin, 1 mM H
2O
2 was added to initiate the biotinylation reaction at room temperature. After 1 min, the medium was aspirated and cells were washed twice with quenching buffer (5 mM Trolox, 10 mM NaN
3, 10 mM sodium ascorbate in PBS) and once with PBS. Cells used for fluorescence microscopy were fixed immediately.
For Western blot and SILAC analyses, cells from each dish were lysed with 1.4 mL RIPA buffer (50 mM Tris, pH 7.4, 5 mM Trolox, 0.5% (w/v) sodium deoxycholate, 150 mM NaCl, 0.1% (w/v) sodium dodecyl sulfate (SDS), 1% (v/v) Triton X-100, 1 mM phenylmethane sulfonyl fluoride (PMSF), 10 mM NaN3, 10 mM sodium ascorbate, 1 µg mL−1 aprotinin, 1 µg mL−1 leupeptin and 1 µg mL−1 pepstatin). The cell lysate was incubated for 5 min on ice and centrifuged for 10 min at 16,000 g and 4°C. The cleared cell lysate was used to enrich biotinylated proteins with Neutravidin beads (Thermo Fisher Scientific, Waltham, MA, USA). For mass spectrometry, cell lysates derived from three 10 cm dishes were pooled, the protein concentration of the cell lysates was determined using the Pierce 660 nm Protein Assay (Thermo Fisher Scientific, Waltham, MA, USA) and equal protein amounts of samples treated with or without rapamycin were mixed prior to addition to Neutravidin beads. For Western blot analyses, the samples were kept separately. For each forward or reverse experiment, six batches of 140 µl Neutravidin beads were incubated with 1.4 mL cell lysate overnight at 4 °C on a rotor. The beads were washed once with washing buffer 1 (50 mM HEPES (pH 7.4), 0.1% (w/v) sodium deoxycholate, 1% (v/v) Triton X-100, 500 mM NaCl, 1 mM ethylenediaminetetraacetic acid (EDTA), once with washing buffer 2 (50 mM Tris (pH 8.0), 250 mM LiCl, 0.5% (v/v) Nonidet P-40, 0.5% (w/v) sodium deoxycholate, 1 mM EDTA) and twice with washing buffer 3 (50 mM Tris (pH 7.4) and 50 mM NaCl). For each washing step, the beads were incubated for 8 min at 4 °C on a rotor. After the last washing step, the buffer was removed carefully and bound proteins were eluted from the beads by incubation for 5 min at 95 °C with 90 µl SDS sample buffer (4% (w/v) SDS, 125 mM Tris pH 6.8, 10% (v/v) glycerol, 0.02% (v/v) bromophenol blue, 10% (v/v) β-mercaptoethanol) supplemented with 5 mM desthiobiotin (Sigma-Aldrich, St. Louis, MO, USA). To increase the protein concentration, three batches of beads were consecutively eluted in the same buffer.
The biotinylation experiment using APEX2-dGFP-NLS-FKBP12 was performed as described above with the following changes: Cells were seeded in six-well plates and transfected with pmCherry-FRB-emerin and pAPEX2-dGFP-NLS-FKBP12. Cells of two wells were lysed in 0.5 mL RIPA buffer. In total, 6 mL of rapamycin-treated cell lysate and 6 mL of lysate derived from cells that had not been treated with rapamycin were obtained and the same protein amounts were mixed. 24 batches of 40 µL Neutravidin beads were incubated with 0.5 mL cell lysate each. For elution, three batches of beads were consecutively incubated in 40 µL SDS sample buffer.
2.4. Standard APEX-Biotinylation Assay
The biotinylation experiment using APEX2 directly fused to emerin was performed as described above for the FKBP12-GFP-APEX2 construct with the following changes: instead of comparing biotinylated proteins with or without incubation of cells with rapamycin, the biotinylation of proteins in cells that had been transfected with either pAPEX2-emerin or with pGFP-APEX2 was compared by SILAC.
2.5. Mass Spectrometric Analyses
Samples were separated on 4–12% NuPAGE Novex Bis-Tris Minigels (Invitrogen, Carlsbad, California). Gels were stained with Coomassie Blue, and each lane sliced into 11–12 equidistant bands. After washing, gel slices were reduced with dithiothreitol (DTT), alkylated with 2-iodoacetamide and digested with trypsin (sequencing grade, Promega, Madison Wisconsin) overnight. The resulting peptide mixtures were then extracted, dried in a SpeedVac, reconstituted in 2% acetonitrile/0.1% formic acid (v/v) and analyzed by nano-LC-MS/MS on a hybrid quadrupole/orbitrap mass spectrometer (Q Exactive, Thermo Fisher Scientific, Dreieich, Germany) as described previously [
30]. Raw data processing and analysis was performed as described [
27], using MaxQuant Software version 1.5.7.4 (Max Planck Institute for Biochemistry, Martinsried, Germany). Perseus Software version 1.5.6.0 (Max Planck Institute for Biochemistry, Martinsried, Germany) was used for statistical evaluation of relative protein quantitation values and a two-sided Significance B test [
31] was performed using normalized log
2 ratios. For the analysis, a Benjamini-Hochberg correction was applied and a threshold value of 0.05 was chosen. For the rapamycin approach, experiments were performed twice, each with two biological and two technical replicates. For the standard APEX-approach, only one experiment was performed, again with two biological and two technical replicates.
2.6. Data Availability
The MS proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE [
32] partner repository with the dataset identifier PXD009783.
2.7. Western Blot Analyses
Western blotting was performed according to standard methods using HRP-coupled secondary antibodies. To detect biotinylated proteins, they were separated by SDS-PAGE using Mini Protean TGX Precast gels (4–20%; Biorad, Hercules, CA, USA). After transfer to nitrocellulose, the membranes were incubated in blocking buffer (3% BSA in TBS-T (24.8 mM Tris, pH 7.4, 137 mM NaCl, 2.7 mM KCl, 1% (v/v) Tween 20)) overnight at 4 °C. Incubation with streptavidin-HRP (Jackson ImmunoResearch Laboratories, West Grove, PA, USA; diluted 1:5,000–1:40,000 in blocking buffer) for 1 h at room temperature was followed by four washing steps with TBS-T. For detection of proteins, Immobilon Western Chemiluminescent HRP Substrate (Millipore, Burlington, MA, USA) was used.
2.8. Immunofluorescence and Fluorescence Microscopy
For fluorescence microscopy, cells were grown on coverslips and fixed with 3.7% (v/v) formaldehyde. Cells expressing fluorescently labeled proteins were mounted directly with MOWIOL supplemented with 1 µg mL
−1 DAPI. For immunofluorescence, fixed cells were permeabilized with 0.5% (v/v) Triton X-100 in PBS for 5 min at room temperature and blocked with 3% (w/v) BSA in PBS for 15 min at room temperature. Staining was performed for 1 h at room temperature using appropriate primary antibodies and fluorescently labeled secondary antibodies (
Table S5), which were diluted in 3% BSA in PBS. Afterwards, cells were embedded in MOWIOL-DAPI.
Microscopic analysis was performed using an LSM-510 confocal laser scanning microscope (Zeiss, Oberkochen, Germany) with a 63x LCI Plan-Neofluar 1.3 NA oil corrected objective.
2.9. Cross-Linking and Coimmunoprecipitation
HeLa P4 cells were washed twice with cold PBS containing 0.1 mM CaCl2 and 1 mM MgCl2 and incubated with dithiobis(succinimidyl propionate); (DSP; Thermo Scientific) at a final concentration of 1 mM in DMSO for 2 h on ice. For control reactions, DMSO alone was used. DSP was quenched by the addition of 20 mM Tris-HCl, pH 7.4, for 15 min. The cells were then washed twice with cold PBS and lysed with 1 mL of lysis buffer (0.5% sodium deoxycholate, 50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 0.25% SDS, and 0.5% Triton X-100 with Complete protease inhibitor mixture (Roche Applied Science)) for 30 min on ice. To reduce viscosity, the lysate was passed through a 27-gauge×3/4-inch needle and then centrifuged at 15,000 g for 20 min at 4 °C. For immunoprecipitation of endogenous protein complexes, 3 µg of mouse anti-emerin, or IgG as a control were immobilized on 50 µL of Protein A-Sepharose 4 Fast Flow beads (GE Healthcare) for 3 h and incubated with lysates from 24 × 106 cells that had or had not been subjected to cross-linking as described above. The beads were then washed four times with washing buffer (10 mM HEPES, 150 mM NaCl, 1 mM EGTA, 0.1 mM MgCl2, 0.1% Triton X-100, and Complete protease inhibitor mixture), and proteins were eluted with sample buffer containing 50 mM DTT.
2.10. Gene Ontology Analysis
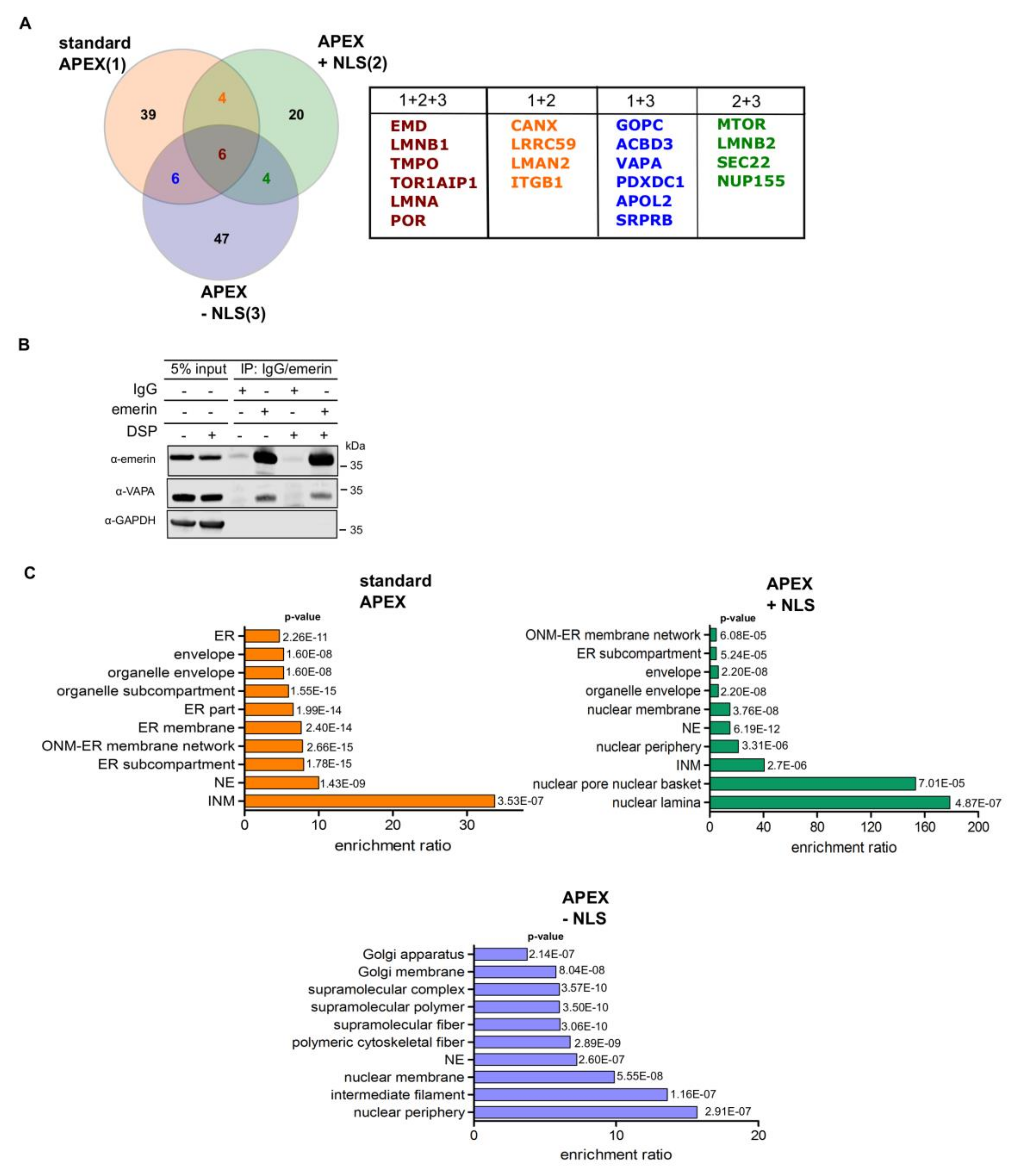
For gene ontology cellular compartment (GOCC) analysis of all the significant proteins found in the three proteomic approaches, the H. sapiens proteome database was used as reference. Proteins were analyzed using WebGestalt (WEB-based Gene SeT AnaLysis Toolkit) with Benjamini-Hochberg multiple test adjustment with a significance level of the false discovery rate (FDR) of p < 0.05. The GO terms were selected for the classification of proteins only if there were more than five and less than 2000 proteins in the reference set.
4. Discussion
Proximity-dependent biotinylation approaches have been used very successfully for the analysis of the interactome of many proteins [
20,
21]. The described methods have in common that they all identify protein neighbors and physical interaction has to be investigated by other approaches. Here, we used different versions of APEX2-dependent biotinylation to probe the environment of emerin at the INM and the ER. First, we used the “standard” approach, where APEX2 was directly fused to emerin. Second, we performed two types of RAPIDS, one with a nuclear version of APEX2 and one with a version that resides in the nucleus and the cytoplasm alike. In both cases, the FKBP12-APEX2 interacts with FRB-tagged mCherry-emerin only upon addition of rapamycin to the cells. Quantitative proteomics then allows a faithful identification of real binding partners or neighboring proteins. This becomes obvious when one compares the results of the “standard” APEX-experiment (
Figure 2D) and RAPIDS (
Figure 4D and
Figure 5F). Clearly, the latter yielded a more defined cloud of non-specifically biotinylated proteins around the intersection point of the x- and y-axis in the scatter plot as compared to the more elongated shape of this population in
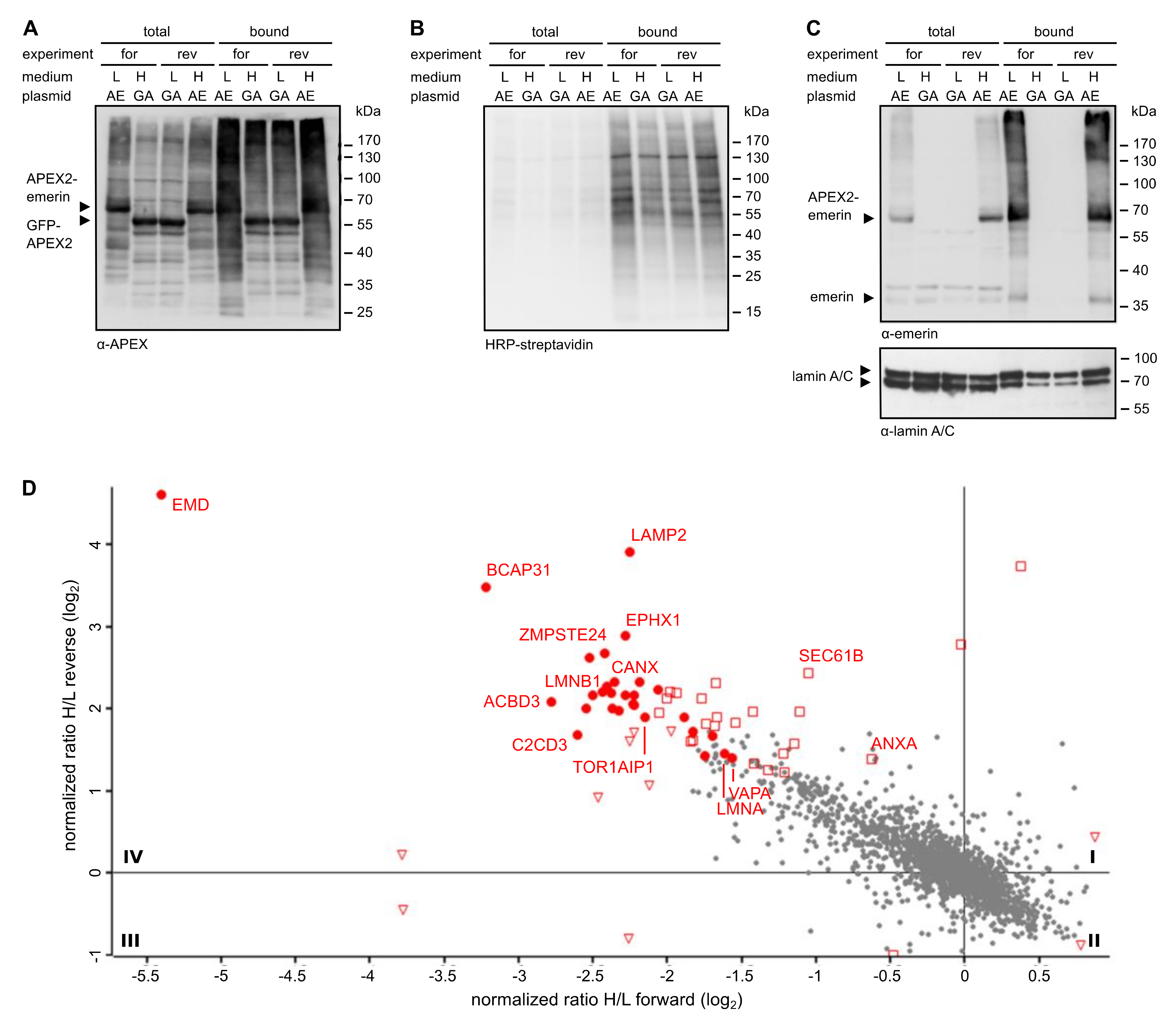
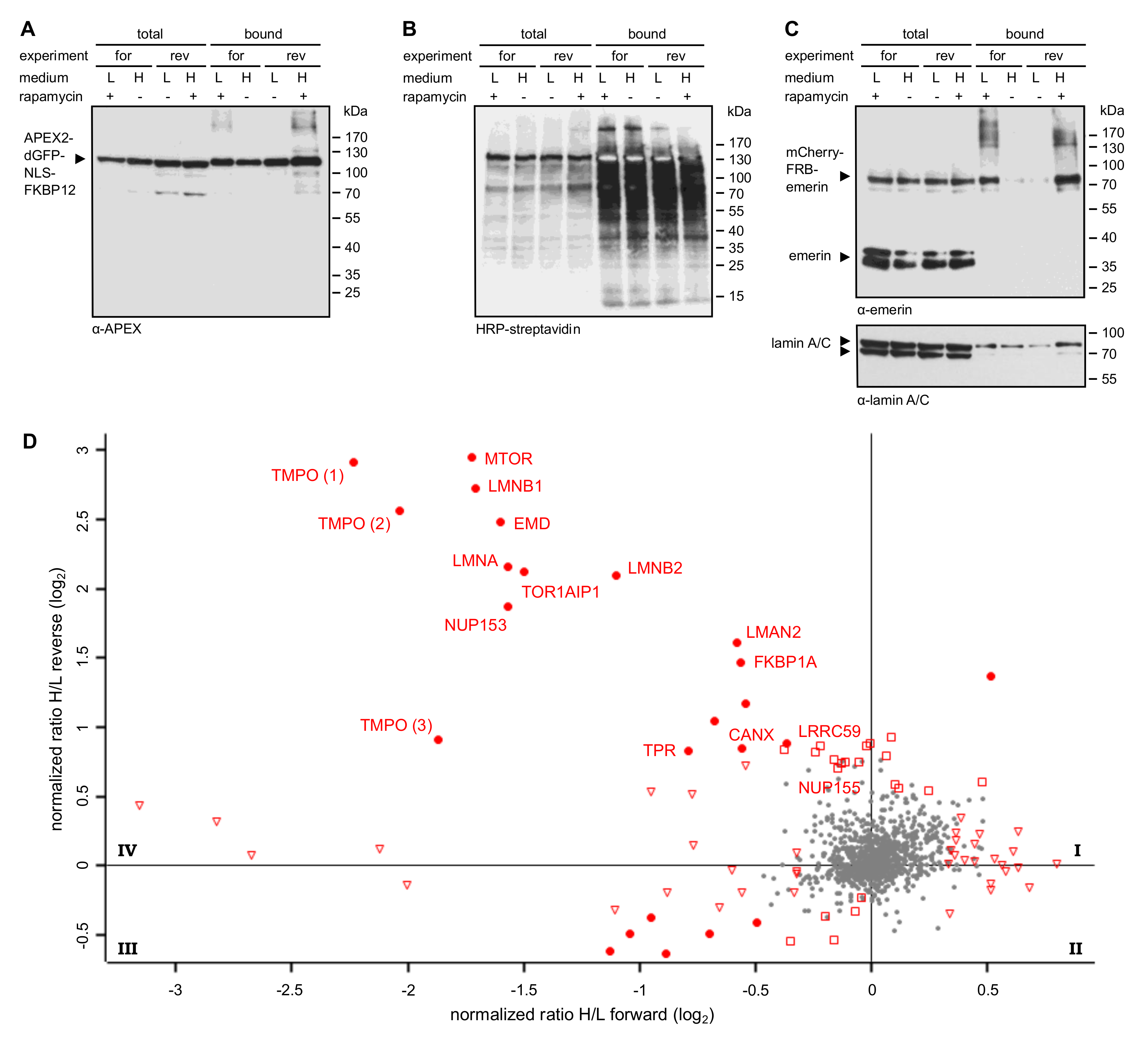
Figure 2D. Thus, a careful design of the experimental conditions in RAPIDS should allow a better discrimination of specific versus unspecific neighbors, compared to other proximity-based methods.
The identification of proteins that interact directly with our protein of interest, emerin, is not trivial, since for affinity approaches, the conditions that are needed to release proteins from the nuclear lamina may also lead to the disruption of specific protein-protein interactions. Indeed, several reported binding partners of emerin were not identified in a large scale affinity approach [
34], including certain nesprins and β-catenin. Using RAPIDS, we identified several known interaction partners of emerin, including LAP1 (TOR1AIP; [
35]) and A- and B-type lamins. Lamins are rather abundant proteins with a few million copies per HeLa cell for lamin A and lamin B1 [
36]. Interestingly, lamin B2, which is far less abundant than lamin A, was also identified with similar significance levels as lamin A, suggesting a specific interaction. Furthermore, several proteins of the INM were found, e.g. LAP2β (thymopoietin, TMPO). Interestingly, emerin was identified as a proximity partner of LAP2β using BioID-approaches [
37]. Emerin itself was also identified in our screen. This could result from biotinylation of mCherry-FRB-emerin and/or endogenous emerin, which is known to interact with itself, at least in vitro [
33]. We also found the nucleoporins Tpr, Nup153 and Nup155 in our screen. These seem to be specific proximity partners, because other nucleoporins, whose abundance is expected to be very similar, were not identified. Tpr and Nup153 localize to the nuclear basket of the NPC, that is, in close proximity to the nuclear lamina and emerin. Nup 155 is a component of the inner ring of the NPC. Whether this reflects a direct interaction or a close proximity remains to be investigated in future studies. Interestingly, Tpr and Nup155 were previously identified in emerin-containing complexes [
34]. Another potential interaction partner of emerin is VAPA, which was found to coprecipitate with emerin (
Figure 6B). Importantly, only endogenous proteins were analyzed in this experiment. VAPA has been described as a protein of the ER [
38]. Emerin was previously detected at the level of the ER as well [
12] and also suggested to exchange between ER and NE [
39]. Furthermore, emerin is initially inserted into the ER-membrane system in a post-translational manner [
13]. Hence, it is possible that the two proteins interact at the level of the ER. It remains to be investigated whether VAPA can also reach the INM and whether it is a direct or an indirect interaction partner of emerin.
What is the difference between our three types of SILAC experiments (
Figure 6), particularly between RAPIDS using APEX2-constructs containing or lacking an NLS? In principle, the addition of rapamycin to cells is expected to sequester the diffusible FKBP12-containing proteins to all sites where mCherry-FRB-emerin is present. Hence, also the large APEX2-dGFP-NLS-FKBP12 could leave the nucleus and bind to targets at ER-membranes (
Figure 3B). The smaller APEX-version, FKBP12-GFP-APEX2, initially localized all over the cell and was then recruited to the ER upon addition of rapamycin in cells co-expressing mCherry-FRB-emerin (
Figure 5B). Thus, both proteins should be able to induce biotinylation of INM-proteins as well as ER-membrane proteins facing the cytoplasm. Nevertheless, the spectrum of identified proteins is somewhat different for the two types of APEX2-constructs. Major partners of emerin were identified with both versions (e.g., lamins and LAP1; these were also found in our “standard” APEX-approach,
Figure 1 and
Figure 2), whereas Tpr and Nup153 were only found with the nuclear version of APEX. APEX2 lacking the NLS, on the other hand, identified a number of additional ER-proteins like ARFGEF1 and VAPA (for comparison see
Figure 6 and
Table S4). The observed differences may result from the different sizes of the APEX constructs (97.4 vs. 67.4 kDa) and/or their enzymatic domains residing at opposite ends (N-terminal vs. C-terminal) of the proteins. Compared to RAPIDS, the “standard” APEX-approach offers less control over the experimental conditions, since the differentiation between specific and non-specific hits relies on the expression of different proteins (APEX2-emerin versus GFP-APEX2;
Figure 1) and subsequent comparison of biotinylated proteins by SILAC. Nevertheless, the method identified several proteins, which were also found in RAPIDS (
Figure 6A). Note, however, that the “standard” APEX experiment was only performed once (yet with biological and technical replicates).
Several reasons could account for the fact that some established binding partners of emerin were not identified in our screens. First, the protein must be expressed in the cell line used for RAPIDS at a sufficiently high level, so that it binds not only to endogenous but also to exogenous emerin. Second, the affinity may not be high enough to allow a faithful detection. The histone deacetylase HDAC3, for example, has a rather low affinity for emerin (7.3 µM; [
18]) and may thus escape biotinylation. Third, appropriate sites for biotinylation must be available on the surface of potential proximity partners. Small proteins like BAF (89 amino acids), a protein that binds to the N-terminal region of emerin [
9], may lack such sites. They could also be protected by other proteins, impeding modification by the biotin-phenoxyl radicals. Fourth, the tag of our overexpressed version of emerin could affect protein-protein interactions. mCherry-FRB-emerin and APEX2-emerin retain the natural C-terminal end of emerin to allow post-translational ER-insertion as a tail-anchored protein. They could, however, interfere with binding of proteins like BAF to the N-terminal region. Furthermore, the size of the tag could affect the efficiency of targeting of the fusion proteins to the INM [
40]. In fact, mCherry-FRB-emerin is about 10 kD larger than APEX2-emerin. Although mCherry-FRB-emerin clearly reached the INM, a substantial portion was observed at the level of the ER (
Figure 3B and
Figure 5B).
In summary, the choice of tags and their position in the fusion proteins could affect the results of biotinylation-based approaches for the identification of proximity partners. A direct fusion of APEX2 to emerin is straight-forward and yielded meaningful results. Depending on the protein of interest, however, such direct fusions may hamper the identification of certain partner proteins, for example as a result of inefficient targeting to the final destination [
27]. Compared to this standard approach, RAPIDS offers the advantage of a physical separation of the radical-generating enzyme and the protein of interest. This allows focusing on specific cellular compartments, for example the nucleus, simply by targeting APEX2 to this organelle, prior to its rapamycin-mediated dimerization with a protein of interest (see
Figure 6B). Notably, a similar approach was used very recently in the context of BioID [
37]. Furthermore, a simple, single-parameter-change (+/- rapamycin) allows a faithful evaluation of specific versus unspecific hits. One caveat of the method as described here results from possible effects of overexpression of the protein of interest. This could be avoided by (i.) the generation of stable cell lines with options to control the expression level of, for example, mCherry-FRB-emerin and/or (ii.) by depleting endogenous proteins with specific siRNAs. On the other hand, overexpression may allow the identification of proteins, which interact (or are in close proximity) only transiently. One example here could be VAPA, an ER-protein that as such is not expected to interact with the INM-protein emerin. The biological significance of the emerin-VAPA interaction remains to be investigated.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





