The Actin-Family Protein Arp4 Is a Novel Suppressor for the Formation and Functions of Nuclear F-Actin

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Plasmid Constructs
2.2. Introduction of Constructs into Cells and Fluorescence Microscopy
2.3. Antibodies and Immunofluorescence Staining
2.4. Quantitative RT-PCR Analysis
2.5. Dual-Luciferase Reporter Assay
2.6. Chromatin Immunoprecipitation (ChIP) Assay
2.7. Nuclear Transfer and Injection of Oocytes
2.8. Statistical Analyses
3. Results and Discussion
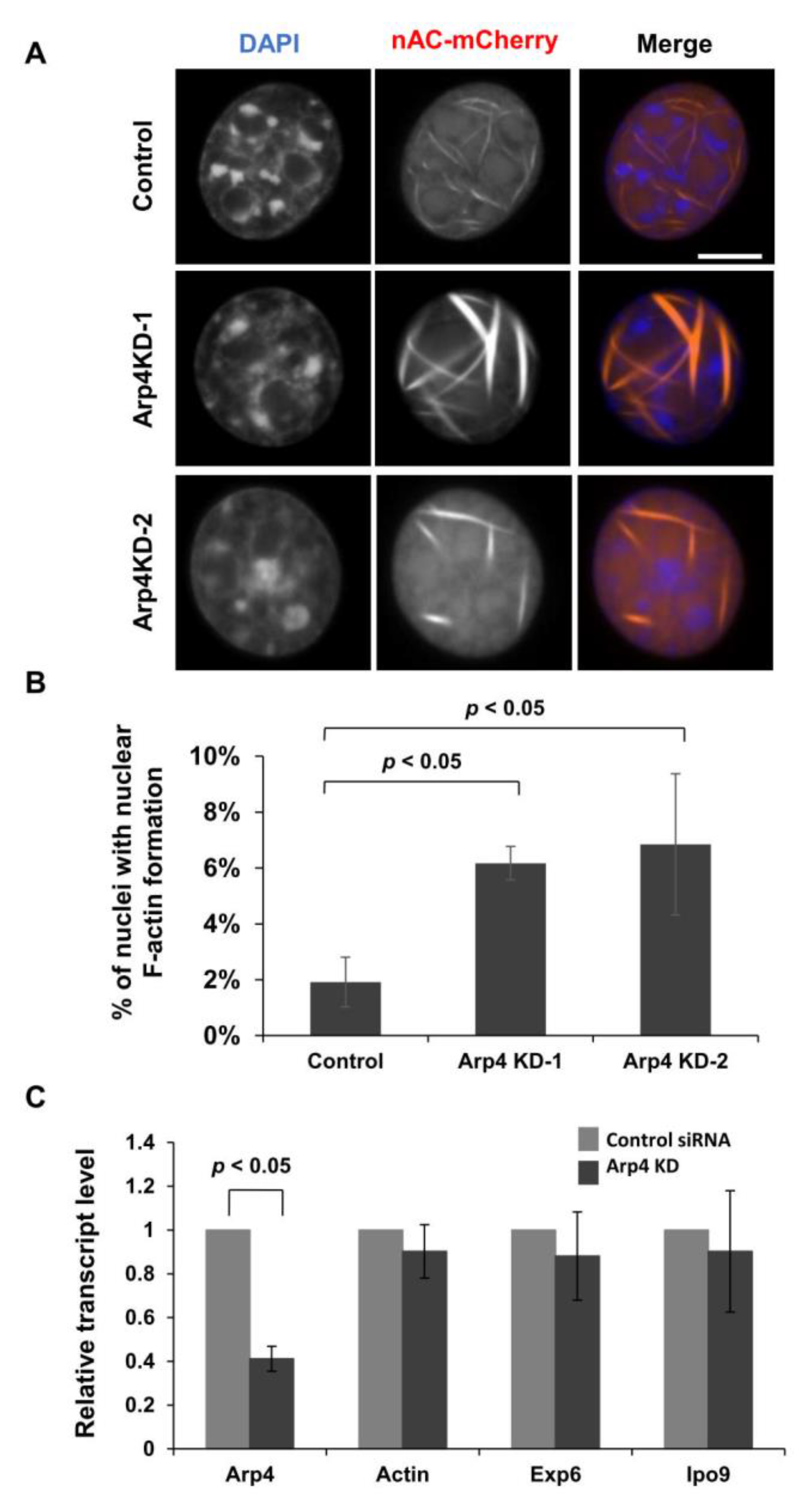
3.1. Depletion of Arp4 Induces Aberrant Endogenous Actin Structures in the Nucleus
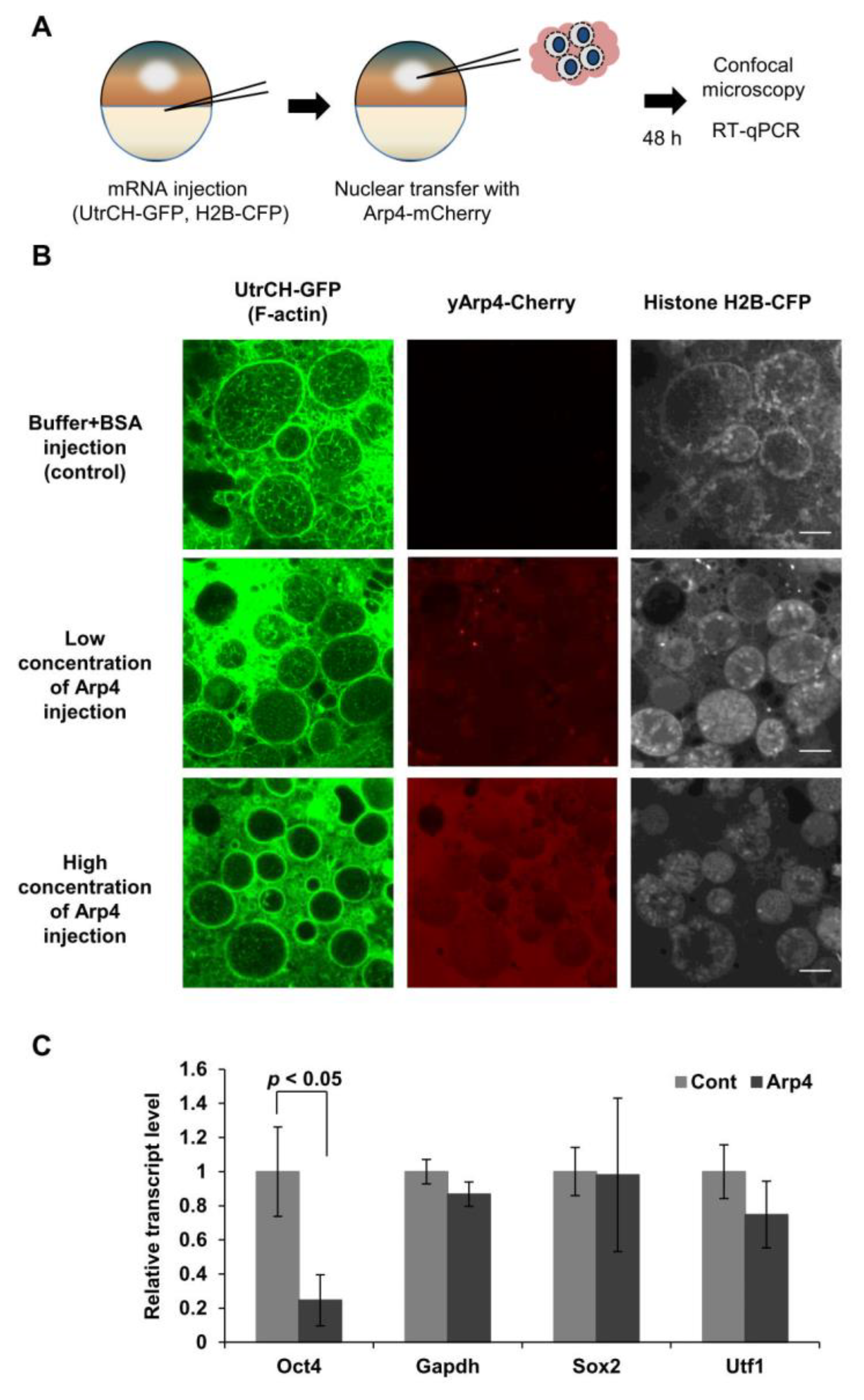
3.2. Arp4 Suppresses Nuclear F-Actin Formation in Xenopus Oocytes
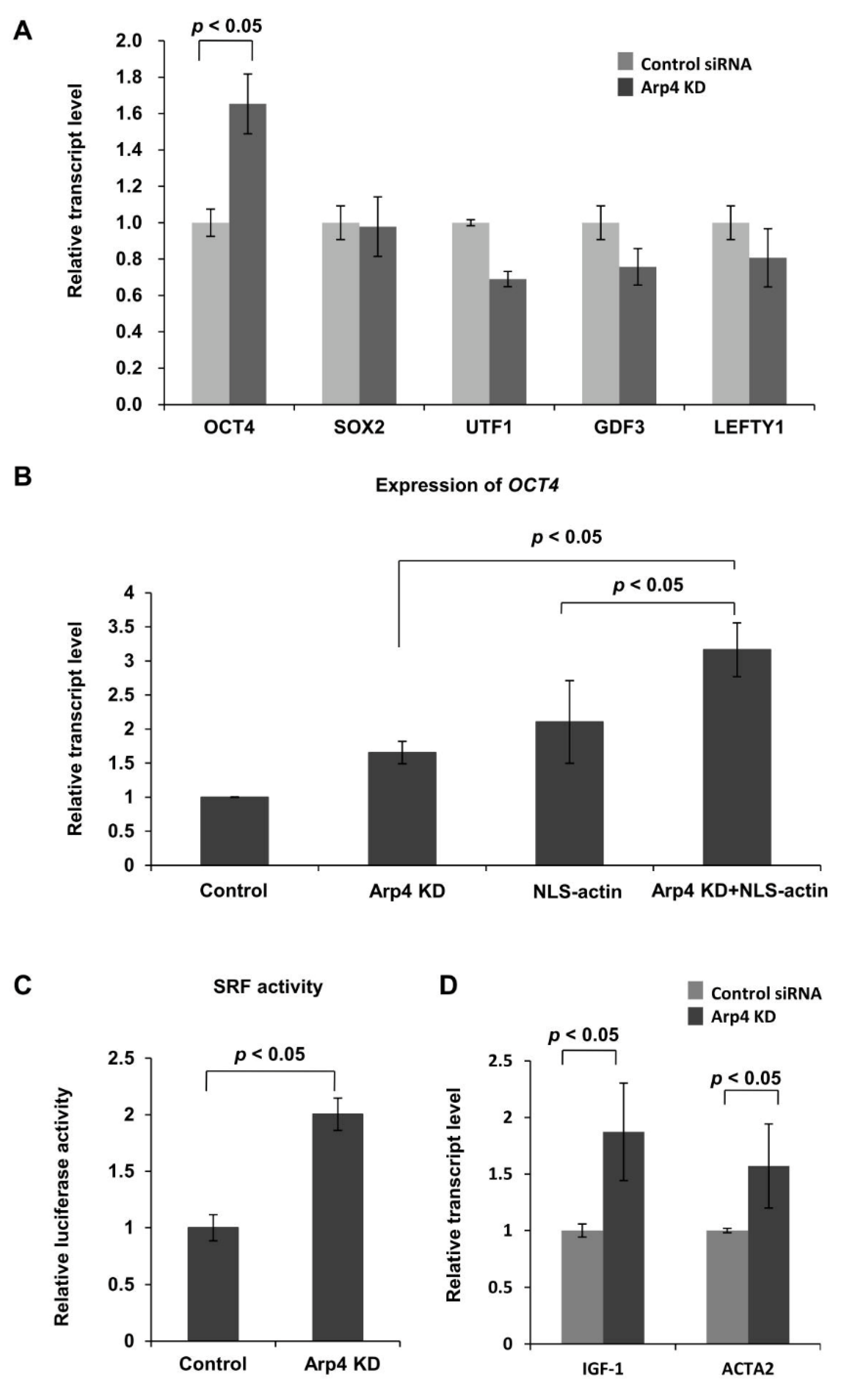
3.3. Effects of Arp4 Knockdown on F-Actin-Inducible Transcription
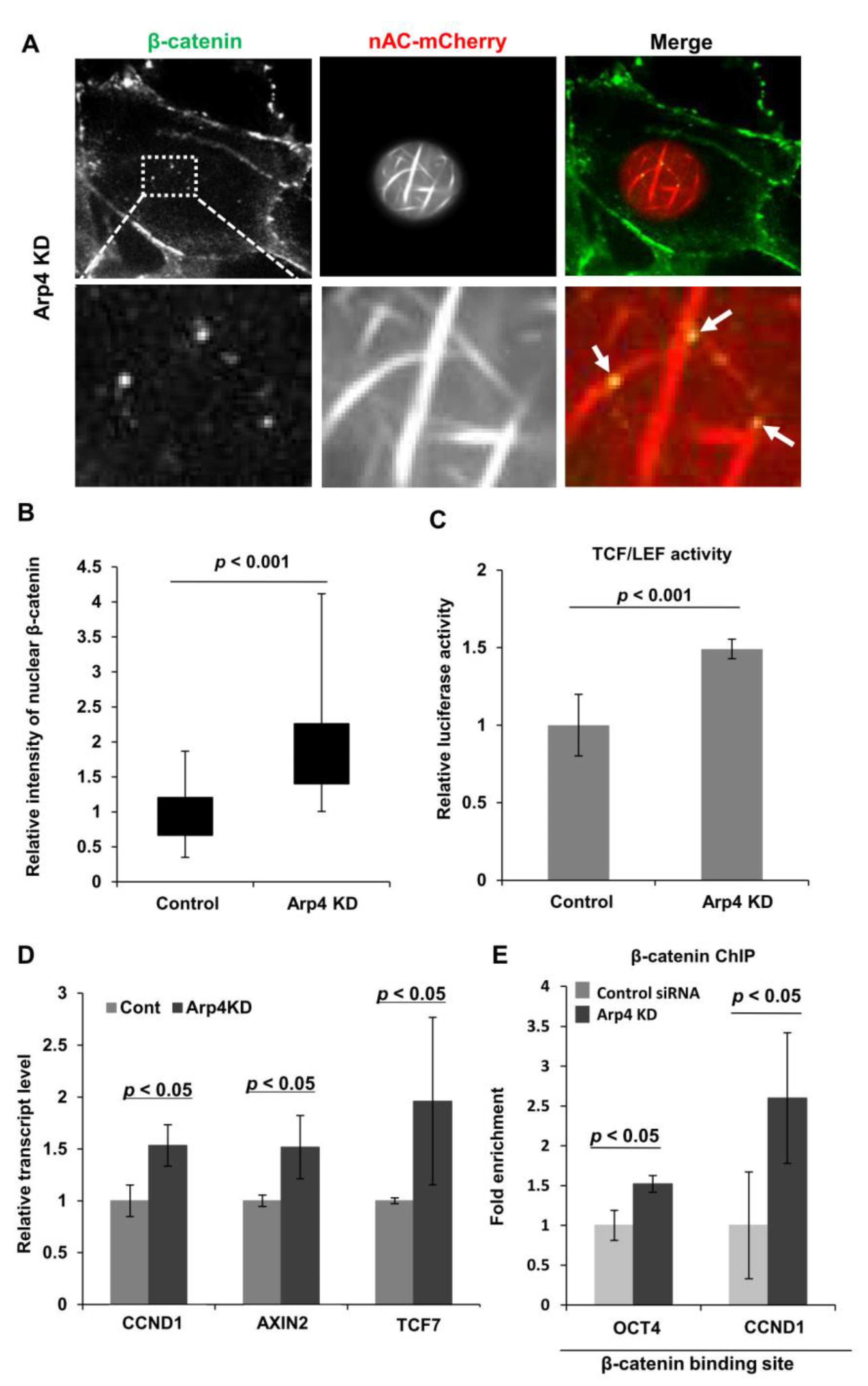
3.4. Depletion of Arp4 Activates Wnt/β-Catenin-Targeting Genes
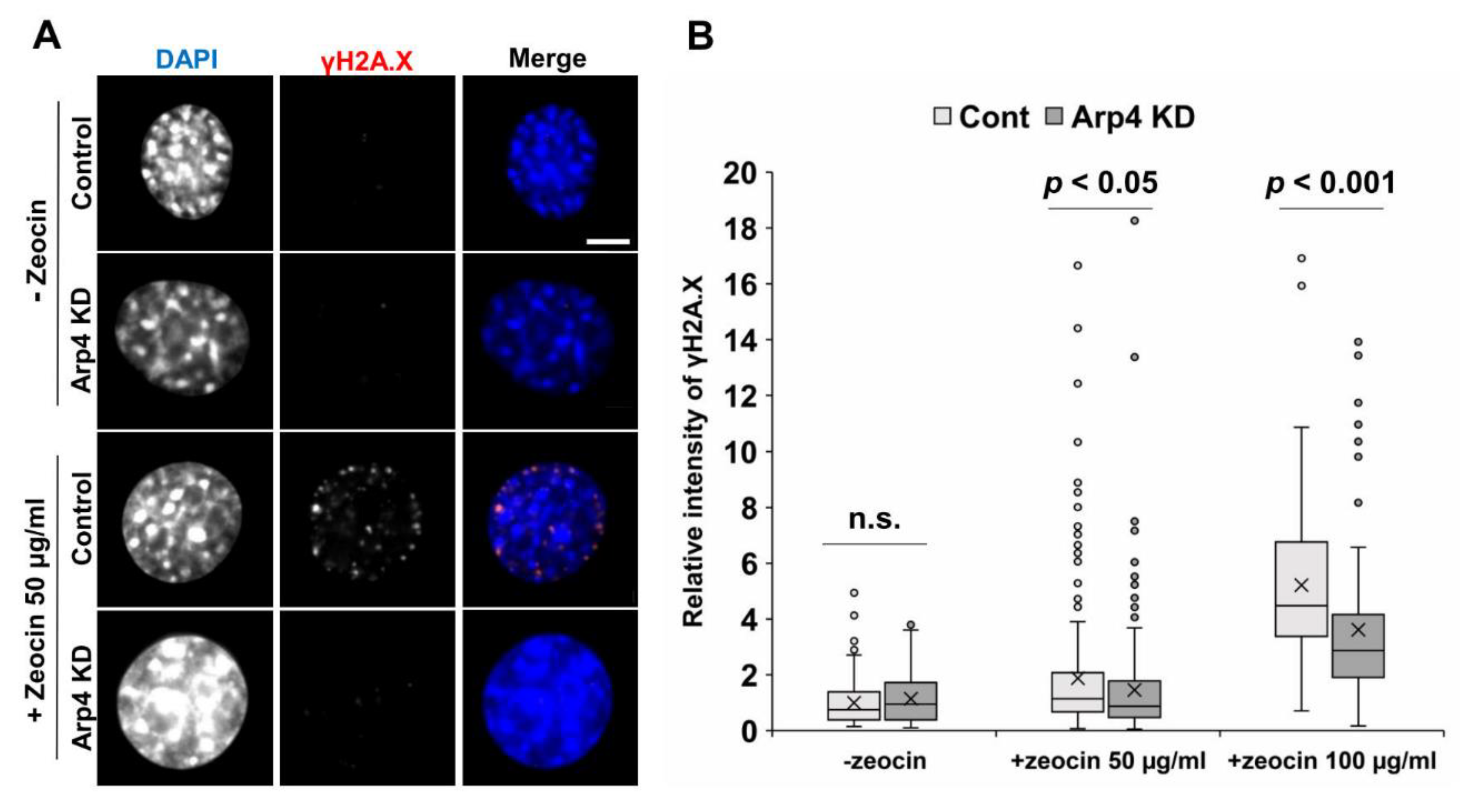
3.5. Arp4 Knockdown Affects DNA Double-Strand Break Repair
3.6. Arp4 is Involved in Genome Functions as a Novel Suppressor of F-Actin Formation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dopie, J.; Skarp, K.P.; Rajakylä, E.K.; Tanhuanpää, K.; Vartiainen, M.K. Active maintenance of nuclear actin by importin 9 supports transcription. Proc. Natl. Acad. Sci. USA 2012, 109, E544–E552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stüven, T.; Hartmann, E.; Görlich, D. Exportin 6: A novel nuclear export receptor that is specific for profilin· actin complexes. EMBO J. 2003, 22, 5928–5940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oma, Y.; Harata, M. Actin-related proteins localized in the nucleus: From discovery to novel roles in nuclear organization. Nucleus 2011, 2, 38–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mal, C.; Baarlink, C.; Wang, H.; Grosse, R. Nuclear Actin Network Assembly by Formins Regulates the SRF. Science 2013, 340, 864–868. [Google Scholar]
- Vartiainen, M.K.; Guettler, S.; Larijani, B.; Treisman, R. Nuclear actin regulates dynamic subcellular localization and activity of the SRF cofactor MAL. Science 2007, 316, 1749–1752. [Google Scholar] [CrossRef] [Green Version]
- Fenn, S.; Gerhold, C.B.; Hopfner, K.P. Nuclear actin-related proteins take shape. Bioarchitecture 2011, 1, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, K.; Pasque, V.; Jullien, J.; Gurdon, J.B. Nuclear actin polymerization is required for transcriptional reprogramming of Oct4 by oocytes. Genes Dev. 2011, 25, 946–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamazaki, S.; Yamamoto, K.; de Lanerolle, P.; Harata, M. Nuclear F-actin enhances the transcriptional activity of β-catenin by increasing its nuclear localization and binding to chromatin. Histochem. Cell Biol. 2016, 145, 389–399. [Google Scholar] [CrossRef]
- Belin, B.J.; Lee, T.; Mullins, R.D. DNA damage induces nuclear actin filament assembly by Formin-2 and Spire-1 / 2 that promotes efficient DNA repair. Elife 2015, 4, e07715. [Google Scholar]
- Schrank, B.R.; Aparicio, T.; Li, Y.; Chang, W.; Chait, B.T.; Gundersen, G.G.; Gottesman, M.E.; Gautier, J. Nuclear ARP2/3 drives DNA break clustering for homology-directed repair. Nature 2018, 559, 61–66. [Google Scholar] [CrossRef]
- Agostino, C.D.; Ryu, T.; Zapotoczny, G.; Delabaere, L.; Li, X.; Khodaverdian, V.Y.; Amaral, N.; Lin, E.; Rau, A.R.; Chiolo, I. Nuclear F-actin and myosins drive relocalization of heterochromatic breaks. Nature 2018, 559, 54–60. [Google Scholar]
- Plessner, M.; Melak, M.; Chinchilla, P.; Baarlink, C.; Grosse, R. Nuclear F-actin formation and reorganization upon cell spreading. J. Biol. Chem. 2015, 290, 11209–11216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Sherrard, A.; Zhao, B.; Melak, M.; Trautwein, J.; Kleinschnitz, E.-M.; Tsopoulidis, N.; Fackler, O.T.; Schwan, C.; Grosse, R. GPCR-induced calcium transients trigger nuclear actin assembly for chromatin dynamics. Nat. Commun. 2019, 10, 5271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baarlink, C.; Plessner, M.; Sherrard, A.; Morita, K.; Misu, S.; Virant, D.; Kleinschnitz, E.M.; Harniman, R.; Alibhai, D.; Baumeister, S.; et al. A transient pool of nuclear F-actin at mitotic exit controls chromatin organization. Nat. Cell Biol. 2017, 19, 1389–1399. [Google Scholar] [CrossRef]
- McDonald, D.; Carrero, G.; Andrin, C.; de Vries, G.; Hendzel, M.J. Nucleoplasmic β-actin exists in a dynamic equilibrium between low-mobility polymeric species and rapidly diffusing populations. J. Cell Biol. 2006, 172, 541–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plessner, M.; Grosse, R. Dynamizing nuclear actin filaments. Curr. Opin. Cell Biol. 2019, 56, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Caridi, C.P.; Plessner, M.; Grosse, R.; Chiolo, I. Nuclear actin filaments in DNA repair dynamics. Nat. Cell Biol. 2019, 21, 1068–1077. [Google Scholar] [CrossRef] [PubMed]
- Dopie, J.; Rajakyla, E.K.; Joensuu, M.S.; Huet, G.; Ferrantelli, E.; Xie, T.; Ja, H. Genome-wide RNAi screen for nuclear actin reveals a network of cofilin regulators. J. Cell Sci. 2015, 128, 2388–2400. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, K.; Teperek, M.; Yusa, K.; Allen, G.E.; Bradshaw, C.R.; Gurdon, J.B. Nuclear Wave1 is required for reprogramming transcription in oocytes and for normal development. Science 2013, 341, 1002–1005. [Google Scholar] [CrossRef] [Green Version]
- Ho, C.Y.; Jaalouk, D.E.; Vartiainen, M.K.; Lammerding, J. Lamin A/C and emerin regulate MKL1–SRF activity by modulating actin dynamics. Nature 2013, 497, 507. [Google Scholar] [CrossRef] [Green Version]
- Saravanan, M.; Wuerges, J.; Bose, D.; McCormack, E.A.; Cook, N.J.; Zhang, X.; Wigley, D.B. Interactions between the nucleosome histone core and Arp8 in the INO80 chromatin remodeling complex. Proc. Natl. Acad. Sci. USA 2012, 109, 20883–20888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenn, S.; Breitsprecher, D.; Gerhold, C.B.; Witte, G.; Faix, J.; Hopfner, K. proteins 4 and 8 reveals their interaction with actin. EMBO J. 2011, 30, 2153–2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerhold, C.B.; Winkler, D.D.; Lakomek, K.; Seifert, F.U.; Fenn, S.; Kessler, B.; Witte, G.; Luger, K. Structure of Actin-related protein 8 and its contribution to nucleosome binding. Nucleic Acids Res. 2012, 40, 11036–11046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, R.C.; Turbedsky, K.; Kaiser, D.A.; Marchand, J.-B.; Higgs, H.N.; Choe, S.; Pollard, T.D. Crystal structure of Arp2/3 complex. Science 2001, 294, 1679–1684. [Google Scholar] [CrossRef] [Green Version]
- Mullins, R.D.; Heuser, J.A.; Pollard, T.D. The interaction of Arp2/3 complex with actin: Nucleation, high affinity pointed end capping, and formation of branching networks of filaments. Proc. Natl. Acad. Sci. USA 1998, 95, 6181–6186. [Google Scholar] [CrossRef] [Green Version]
- Tsopoulidis, N.; Kaw, S.; Laketa, V.; Kutscheidt, S.; Baarlink, C.; Stolp, B.; Grosse, R.; Fackler, O.T. T cell receptor–triggered nuclear actin network formation drives CD4+ T cell effector functions. Sci. Immunol. 2019, 4, eaav1987. [Google Scholar] [CrossRef] [Green Version]
- Harata, M.; Oma, Y.; Tabuchi, T.; Zhang, Y.; Stillman, D.J.; Mizuno, S. Multiple actin-related proteins of Saccharomyces cerevisiae are present in the nucleus. J. Biochem. 2000, 128, 665–671. [Google Scholar] [CrossRef]
- Nishimoto, N.; Watanabe, M.; Watanabe, S.; Sugimoto, N.; Yugawa, T.; Ikura, T.; Koiwai, O.; Kiyono, T.; Fujita, M. Heterocomplex formation by Arp4 and beta-actin is involved in the integrity of the Brg1 chromatin remodeling complex. J. Cell Sci. 2012, 125, 3870–3882. [Google Scholar] [CrossRef] [Green Version]
- Tosi, A.; Haas, C.; Herzog, F.; Gilmozzi, A.; Berninghausen, O.; Ungewickell, C.; Gerhold, C.B.; Lakomek, K.; Aebersold, R.; Beckmann, R. Structure and subunit topology of the INO80 chromatin remodeler and its nucleosome complex. Cell 2013, 154, 1207–1219. [Google Scholar] [CrossRef] [Green Version]
- Dion, V.; Shimada, K.; Gasser, S.M. Actin-related proteins in the nucleus: Life beyond chromatin remodelers. Curr. Opin. Cell Biol. 2010, 22, 383–391. [Google Scholar] [CrossRef]
- Cao, T.; Sun, L.; Jiang, Y.; Huang, S.; Wang, J.; Chen, Z. Crystal structure of a nuclear actin ternary complex. Proc. Natl. Acad. Sci. USA 2016, 113, 8985–8990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurdon, J.B.; Melton, D.A. Nuclear Reprogramming in Cells. Science 2008, 322, 1811–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrne, J.A.; Simonsson, S.; Western, P.S.; Gurdon, J.B. Nuclei of Adult Mammalian Somatic Cells Are Directly Reprogrammed to oct-4 Stem Cell Gene Expression by Amphibian Oocytes. Curr. Biol. 2003, 13, 1206–1213. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, S.; Yamamoto, K.; Tokunaga, M.; Sakata-Sogawa, K.; Harata, M. Nuclear actin activates human transcription factor genes including the OCT4 Gene. Biosci. Biotechnol. Biochem. 2015, 79, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Hu, W.; Chernaya, O.; Antic, N.; Gu, L.; Gupta, M.; Piano, M.; Lanerolle, P. De Increased Myosin Light Chain Kinase Expression in Hypertension: Regulation by Serum Response Factor via an Insertion Mutation in the Promoter. Mol. Biol. Cell 2006, 17, 4039–4050. [Google Scholar] [CrossRef] [Green Version]
- Harata, M.; Mochizuki, R.; Mizuno, S. Two isoforms of a human actin-related protein show nuclear localization and mutually selective expression between brain and other tissues. Biosci. Biotechnol. Biochem. 1999, 63, 917–923. [Google Scholar] [CrossRef]
- Clapier, C.R.; Cairns, B.R. The Biology of Chromatin Remodeling Complexes. Annu. Rev. Biochem. 2009, 78, 273–304. [Google Scholar] [CrossRef]
- Kapoor, P.; Chen, M.; Winkler, D.D.; Luger, K.; Shen, X. Evidence for monomeric actin function in INO80 chromatin remodeling. Nat. Struct. Mol. Biol. 2013, 20, 426. [Google Scholar] [CrossRef] [Green Version]
- Harata, M.; Oma, Y.; Mizuno, S.; Jiang, Y.W.; Stillman, D.J.; Wintersberger, U. The nuclear actin-related protein of Saccharomyces cerevisiae, Act3p/Arp4, interacts with core histones. Mol. Biol. Cell 1999, 10, 2595–2605. [Google Scholar] [CrossRef] [Green Version]
- Sunada, R.; Görzer, I.; Oma, Y.; Yoshida, T.; Suka, N.; Wintersberger, U.; Harata, M. The nuclear actin-related protein Act3p/Arp4p is involved in the dynamics of chromatin-modulating complexes. Yeast 2005, 22, 753–768. [Google Scholar] [CrossRef] [Green Version]
- Kast, D.J.; Dominguez, R. Arp you ready for actin in the nucleus? EMBO J. 2011, 30, 2097–2098. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Tang, J.; Lopez-pajares, V.; Tao, S.; Qu, K.; Crabtree, G.R.; Khavari, P.A. Article ACTL6a Enforces the Epidermal Progenitor State by Suppressing SWI/SNF-Dependent Induction of KLF4. Stem Cell 2013, 12, 193–203. [Google Scholar]
- Lu, W.; Fang, L.; Ouyang, B.; Zhang, X.; Zhan, S.; Feng, X.; Bai, Y.; Han, X.; Kim, H.; He, Q. Actl6a protects embryonic stem cells from differentiating into primitive endoderm. Stem Cells 2015, 33, 1782–1793. [Google Scholar] [CrossRef] [PubMed]
- Prasad, P.; Rönnerblad, M.; Arner, E.; Itoh, M.; Kawaji, H.; Lassmann, T.; Daub, C.O.; Forrest, A.R.R.; Lennartsson, A.; Ekwall, K. High-throughput transcription profiling identifies putative epigenetic regulators of hematopoiesis. Blood 2014, 123, e46–e57. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, J.; Wei, Y.; Li, Q.; Wang, Q. ACTL6A regulates follicle-stimulating hormone-driven glycolysis in ovarian cancer cells via PGK1. Cell Death Dis. 2019, 10, 811. [Google Scholar] [CrossRef] [Green Version]
- Saladi, S.V.; Ross, K.; Karaayvaz, M.; Tata, P.R.; Mou, H.; Rajagopal, J.; Ramaswamy, S.; Ellisen, L.W. ACTL6A Is Co-Amplified with p63 in Squamous Cell Carcinoma to Drive YAP Activation, Regenerative Proliferation, and Poor Prognosis. Cancer Cell 2017, 31, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Yang, H.; Xiao, S. ACTL6A expression promotes invasion, metastasis and epithelial mesenchymal transition of colon cancer. BMC Cancer 2018, 18, 1020. [Google Scholar] [CrossRef]
- Sun, W.; Wang, W.; Lei, J.; Li, H.; Wu, Y. Actin-like protein 6A is a novel prognostic indicator promoting invasion and metastasis in osteosarcoma. Oncol. Rep. 2017, 37, 2405–2417. [Google Scholar] [CrossRef]
- Fiore, A.P.Z.P.; Spencer, V.A.; Mori, H.; Carvalho, H.F.; Bissell, M.J.; Bruni-Cardoso, A. Laminin-111 and the Level of Nuclear Actin Regulate Epithelial Quiescence via Exportin-6. Cell Rep. 2017, 19, 2102–2115. [Google Scholar] [CrossRef] [Green Version]
- Kelpsch, D.J.; Tootle, T.L. Nuclear Actin: From Discovery to Function. Anat. Rec. (Hoboken) 2018, 301, 1999–2013. [Google Scholar] [CrossRef] [Green Version]
- Okorokov, A.L.; Rubbi, C.P.; Metcalfe, S.; Milner, J. The interaction of p53 with the nuclear matrix is mediated by F-actin and modulated by DNA damage. Oncogene 2002, 21, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Serebryannyy, L.A.; Cruz, C.M.; De Lanerolle, P. A role for nuclear actin in HDAC 1 and 2 regulation. Sci. Rep. 2016, 6, 28460. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.Z.; Thuraisingam, T.; de Lima Morais, D.A.; Rola-Pleszczynski, M.; Radzioch, D. Nuclear translocation of β-actin is involved in transcriptional regulation during macrophage differentiation of HL-60 cells. Mol. Biol. Cell 2010, 21, 811–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamazaki, S.; Gerhold, C.; Yamamoto, K.; Ueno, Y.; Grosse, R.; Miyamoto, K.; Harata, M. The Actin-Family Protein Arp4 Is a Novel Suppressor for the Formation and Functions of Nuclear F-Actin. Cells 2020, 9, 758. https://doi.org/10.3390/cells9030758
Yamazaki S, Gerhold C, Yamamoto K, Ueno Y, Grosse R, Miyamoto K, Harata M. The Actin-Family Protein Arp4 Is a Novel Suppressor for the Formation and Functions of Nuclear F-Actin. Cells. 2020; 9(3):758. https://doi.org/10.3390/cells9030758
Chicago/Turabian StyleYamazaki, Shota, Christian Gerhold, Koji Yamamoto, Yuya Ueno, Robert Grosse, Kei Miyamoto, and Masahiko Harata. 2020. "The Actin-Family Protein Arp4 Is a Novel Suppressor for the Formation and Functions of Nuclear F-Actin" Cells 9, no. 3: 758. https://doi.org/10.3390/cells9030758
APA StyleYamazaki, S., Gerhold, C., Yamamoto, K., Ueno, Y., Grosse, R., Miyamoto, K., & Harata, M. (2020). The Actin-Family Protein Arp4 Is a Novel Suppressor for the Formation and Functions of Nuclear F-Actin. Cells, 9(3), 758. https://doi.org/10.3390/cells9030758