Transcriptional Plasticity of Autophagy-Related Genes Correlates with the Genetic Response to Nitrate Starvation in Arabidopsis Thaliana



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Plant Growth Condition
2.3. RNA Extraction and Gene Expression Analysis
2.4. Statistical Analysis
3. Results
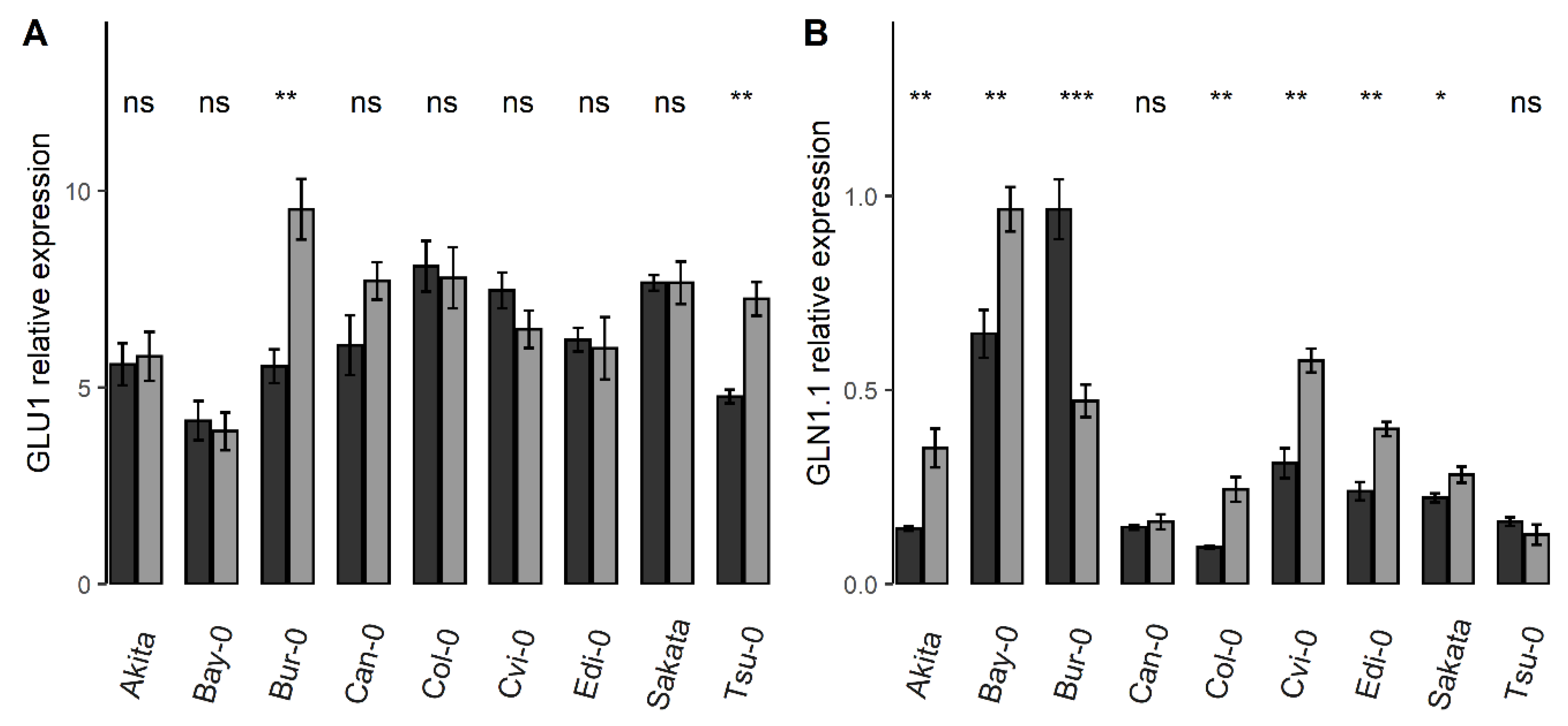
3.1. Variatons in GLN1.1, Nii, GDH, GLU1, and GLT Gene Expressions Are Good Markers to Characterize the Genotype-Response to N Starvation
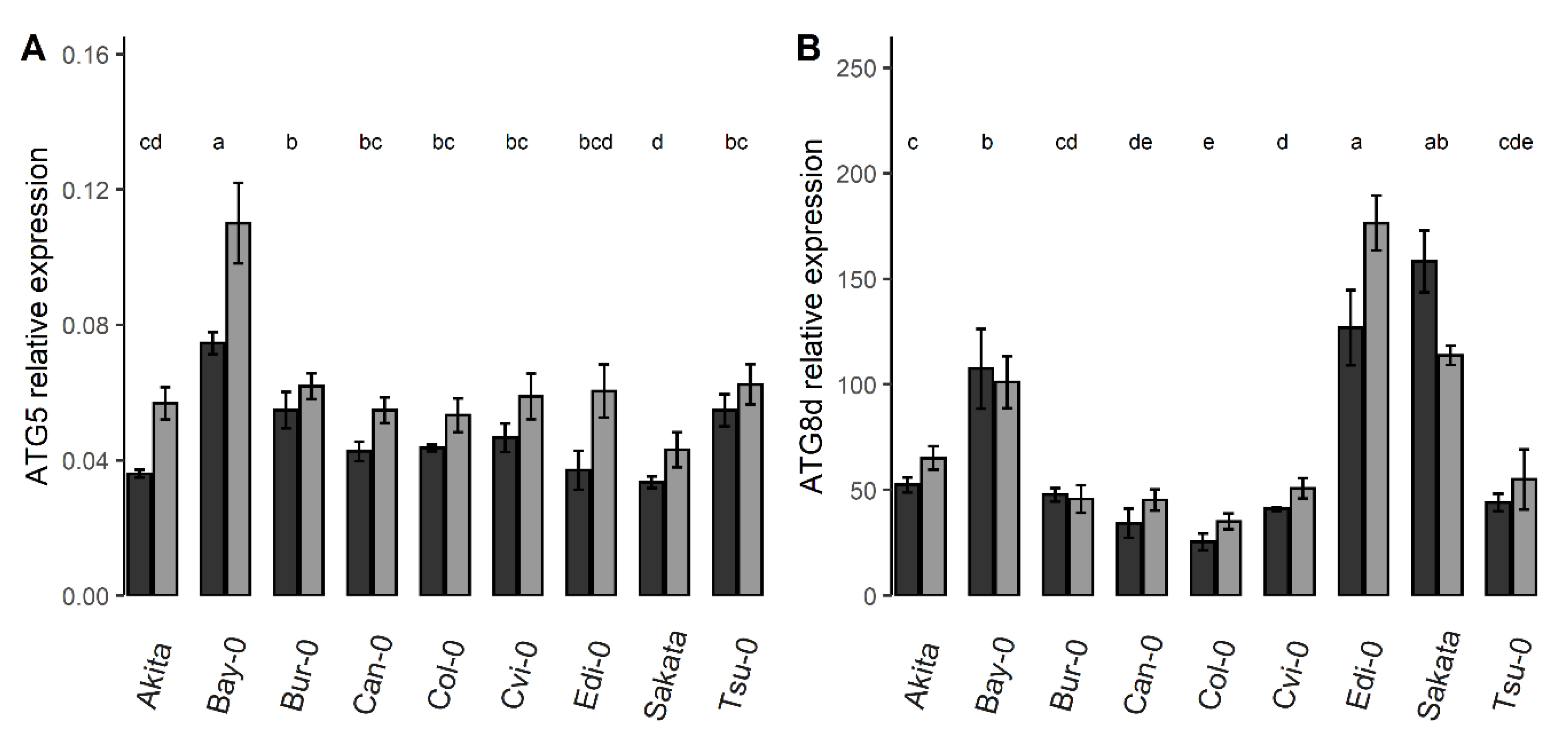
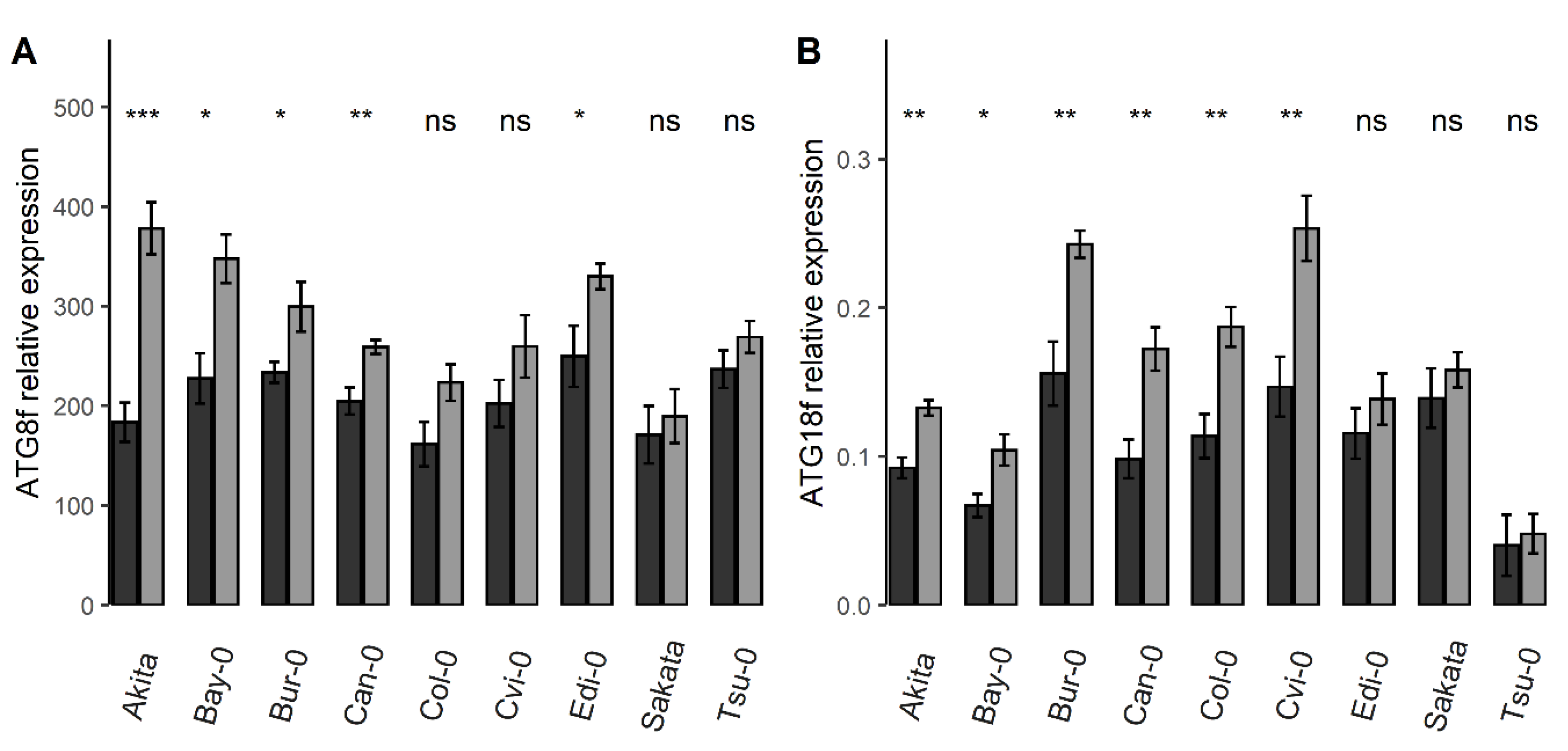
3.2. The Relative Expressions of 18 Autophagy Genes Were Higher under Nitrate Starvation Than in the Control Condition
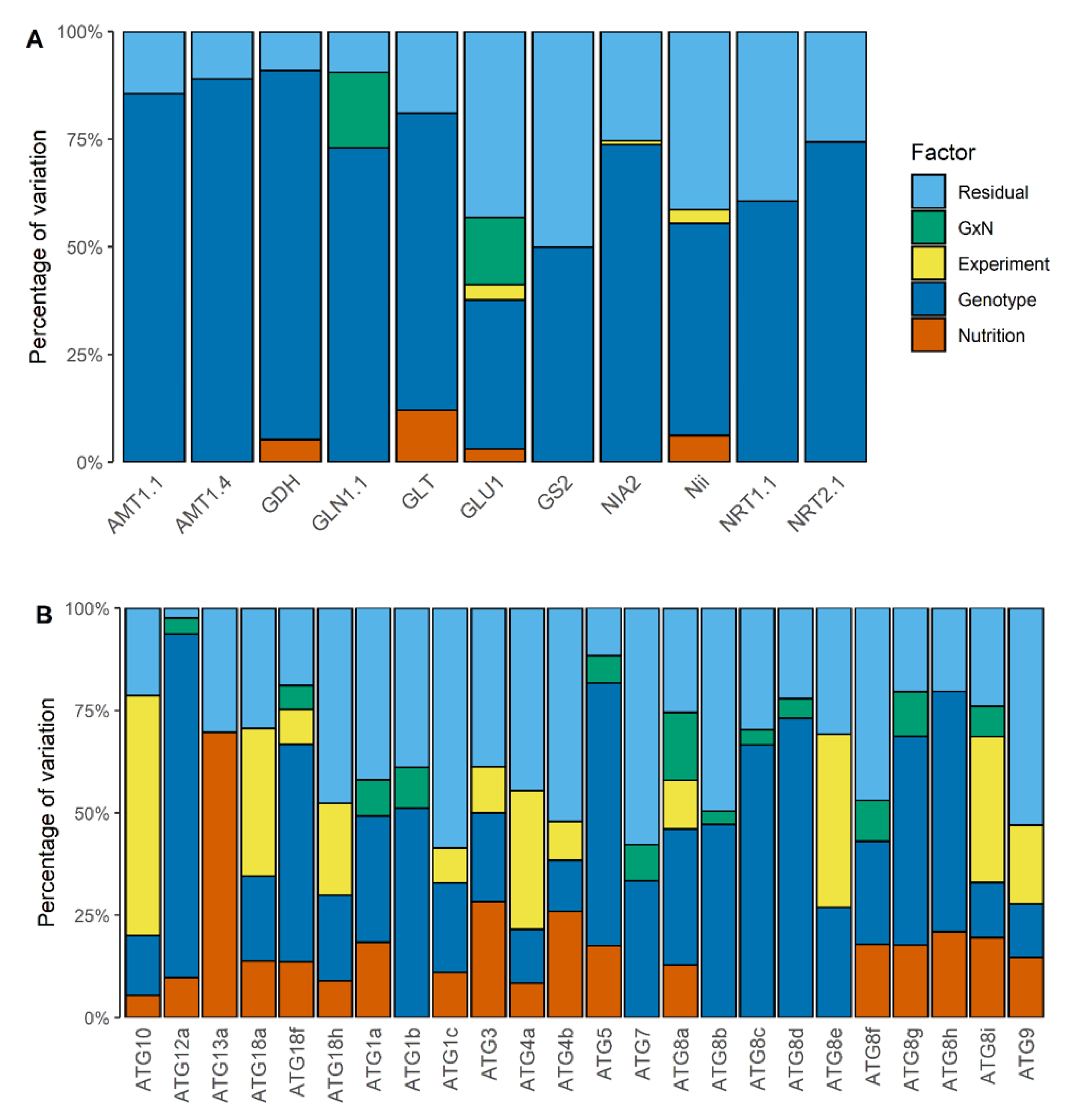
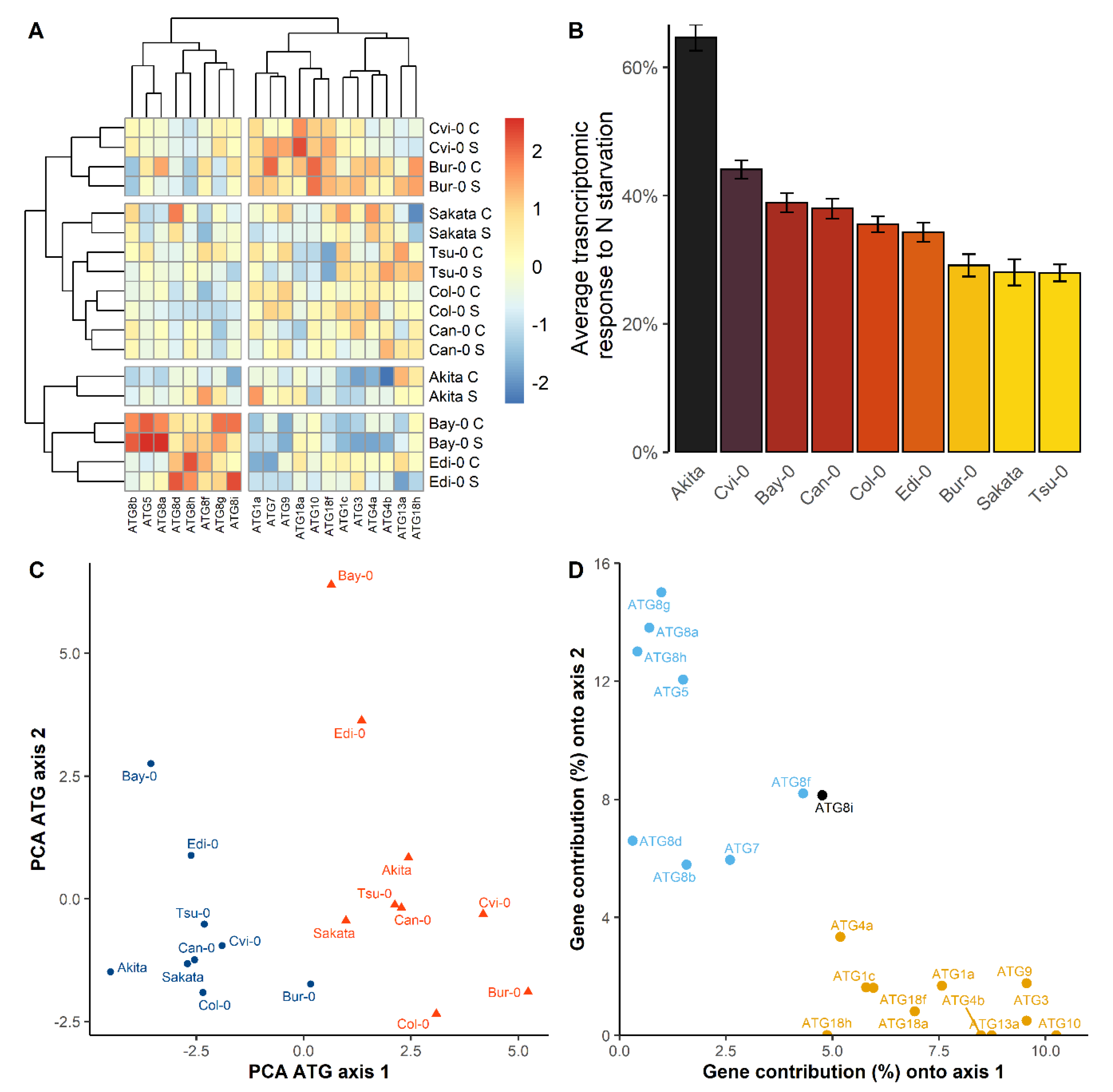
3.3. The Collection of Natural Accessions Revealed Genetic Variation of the Relative ATG Gene Expressions
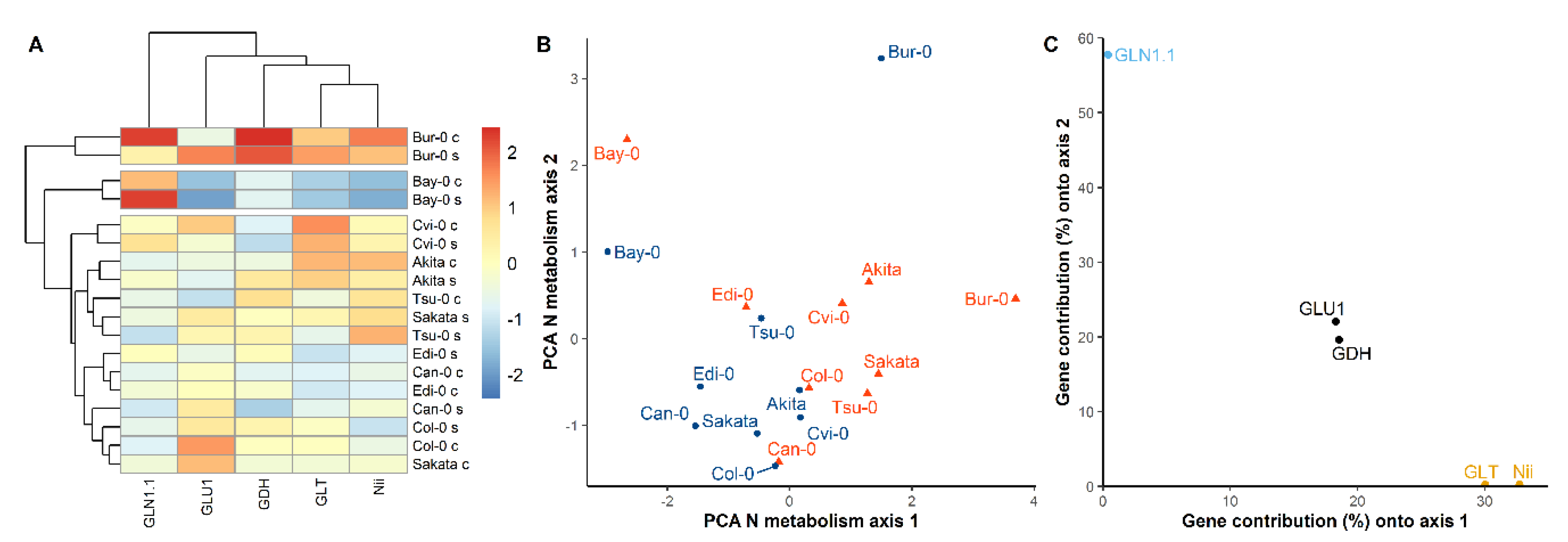
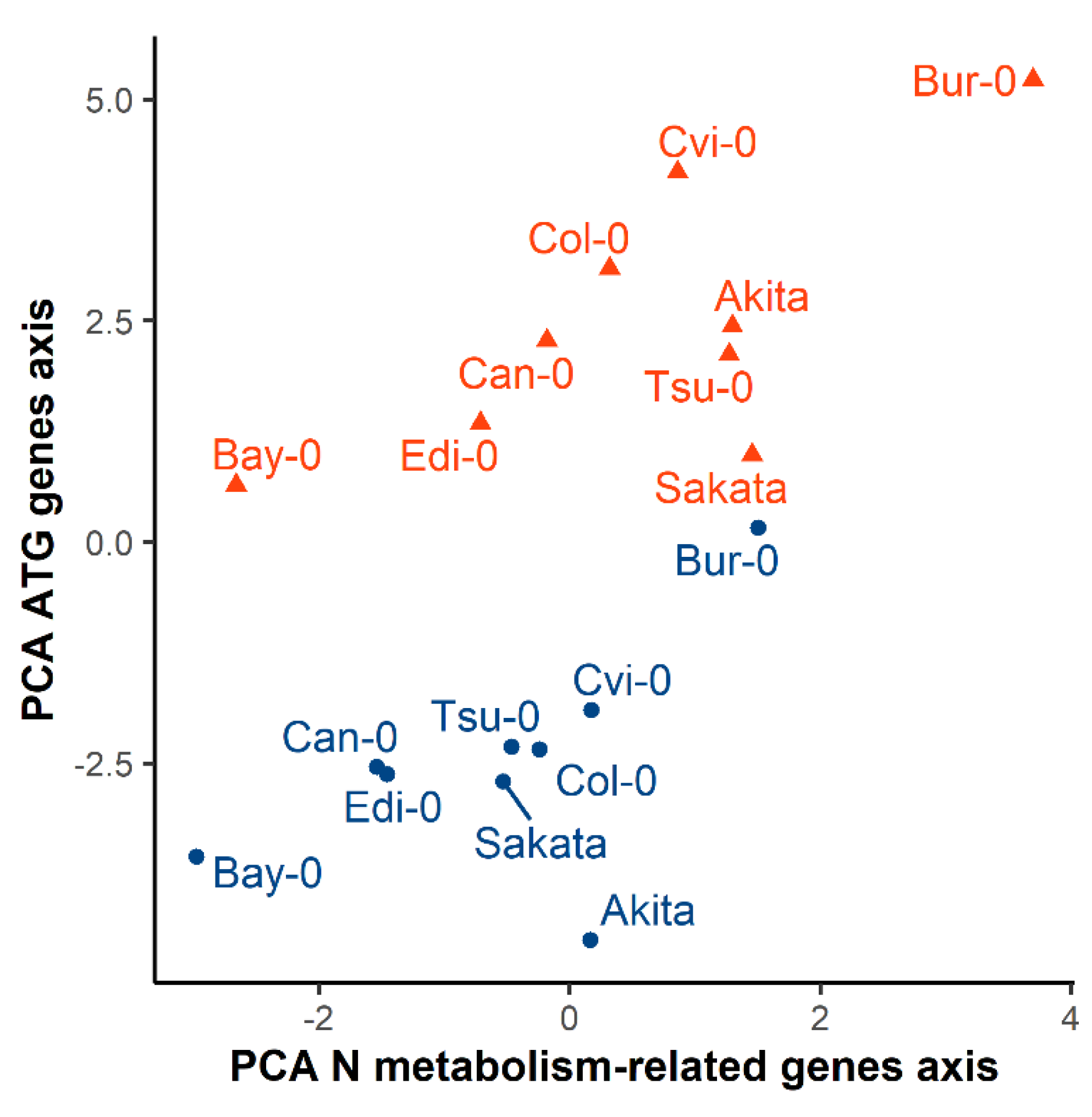
3.4. The Diversity of N Metabolism Gene Expressions and ATG Gene Expressions Are Correlated in the Two Nutritive Conditions
4. Discussion
4.1. Coordinated Regulation of ATG Genes in Response to N Stress Condition
4.2. Transcriptinal Diversity of ATG Genes among Arabidopsis Accessions
4.3. Autophagy Is Part of the Integral Response of the N Metabolism to N Availability
- (i)
- the results of our study are consistent with the other experiments,
- (ii)
- the average on the nine accessions allowed us to obtain more congruent results than with a single accession, supporting then the relevance of our approach based on the natural diversity,
- (iii)
- our study provides new results, in particular for ATG genes, that remain poorly documented in the published transcriptomic data cited in Table S4.
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef] [Green Version]
- Lassaletta, L.; Billen, G.; Grizzetti, B.; Anglade, J.; Garnier, J. 50 year trends in nitrogen use efficiency of world cropping systems: The relationship between yield and nitrogen input to cropland. Environ. Res. Lett. 2014, 9, 105011. [Google Scholar] [CrossRef]
- Strahm, B.D.; Harrison, R.B. Nitrate Sorption in a Variable-Charge Forest Soil of the Pacific Northwest. Soil Sci. 2006, 171, 313–321. [Google Scholar] [CrossRef]
- Tschoep, H.; Gibon, Y.; Carillo, P.; Armengaud, P.; Szecowka, M.; Nunes-Nesi, A.; Fernie, A.R.; Koehl, K.; Stitt, M. Adjustment of growth and central metabolism to a mild but sustained nitrogen-limitation in Arabidopsis. Plant Cell Environ. 2009, 32, 300–318. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.M.; Wang, R.L.; Zhu, T.; Rothstein, S.J. Global transcription profiling reveals differential responses to chronic nitrogen stress and putative nitrogen regulatory components in Arabidopsis. BMC Genom. 2007, 8, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krapp, A.; Berthomé, R.; Orsel, M.; Mercey-Boutet, S.; Yu, A.; Castaings, L.; Elftieh, S.; Major, H.; Renou, J.P.; Daniel-Vedele, F. Arabidopsis roots and shoots show distinct temporal adaptation patterns toward nitrogen starvation. Plant Physiol. 2011, 157, 1255–1282. [Google Scholar] [CrossRef] [Green Version]
- Menz, J.; Li, Z.; Schulze, W.X.; Ludewig, U. Early nitrogen-deprivation responses in Arabidopsis roots reveal distinct differences on transcriptome and (phospho-) proteome levels between nitrate and ammonium nutrition. Plant J. 2016, 88, 717–734. [Google Scholar] [CrossRef]
- Scheible, W.R.; Morcuende, R.; Czechowski, T.; Fritz, C.; Osuna, D.; Palacios-Rojas, N.; Schindelasch, D.; Thimm, O.; Udvardi, M.K.; Stitt, M. Genome-wide reprogramming of primary and secondary metabolism, protein synthesis, cellular growth processes, and the regulatory infrastructure of arabidopsis in response to nitrogen. Plant Physiol. 2004, 136, 2483–2499. [Google Scholar] [CrossRef] [Green Version]
- Kiba, T.; Krapp, A. Plant Nitrogen Acquisition Under Low Availability: Regulation of Uptake and Root Architecture. Plant Cell Physiol. 2016, 57, 707–714. [Google Scholar] [CrossRef] [Green Version]
- Krapp, A.; David, L.C.; Chardin, C.; Girin, T.; Marmagne, A.; Leprince, A.-S.; Chaillou, S.; Ferrario-Méry, S.; Meyer, C.; Daniel-Vedele, F. Nitrate transport and signalling in Arabidopsis. J. Exp. Bot. 2014, 65, 789–798. [Google Scholar] [CrossRef]
- Nacry, P.; Bouguyon, E.; Gojon, A. Nitrogen acquisition by roots: Physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource. Plant Soil 2013, 370, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Loqué, D.; Yuan, L.; Kojima, S.; Gojon, A.; Wirth, J.; Gazzarrini, S.; Ishiyama, K.; Takahashi, H.; von Wirén, N. Additive contribution of AMT1;1 and AMT1;3 to high-affinity ammonium uptake across the plasma membrane of nitrogen-deficient Arabidopsis roots. Plant J. 2006, 48, 522–534. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Loqué, D.; Kojima, S.; Rauch, S.; Ishiyama, K.; Inoue, E.; Takahashi, H.; Von Wiréna, N. The organization of high-affinity ammonium uptake in Arabidopsis roots depends on the spatial arrangement and biochemical properties of AMT1-type transporters. Plant Cell 2007, 19, 2636–2652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takabayashi, A.; Niwata, A.; Tanaka, A. Direct interaction with ACR11 is necessary for post-transcriptional control of GLU1-encoded ferredoxin-dependent glutamate synthase in leaves. Sci. Rep. 2016, 6, 29668. [Google Scholar] [CrossRef] [Green Version]
- Krapp, A. Plant nitrogen assimilation and its regulation: A complex puzzle with missing pieces. Curr. Opin. Plant Biol. 2015, 25, 115–122. [Google Scholar] [CrossRef]
- Alonso-Blanco, C.; Aarts, M.G.M.; Bentsink, L.; Keurentjes, J.J.B.; Reymond, M.; Vreugdenhil, D.; Koornneef, M. What has natural variation taught us about plant development, physiology, and adaptation? Plant Cell 2009, 21, 1877–1896. [Google Scholar] [CrossRef] [Green Version]
- Ikram, S.; Bedu, M.; Daniel-Vedele, F.; Chaillou, S.; Chardon, F. Natural variation of Arabidopsis response to nitrogen availability. J. Exp. Bot. 2012, 63, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Chietera, G.; Chaillou, S.; Bedu, M.; Marmagne, A.; Masclaux-Daubresse, C.; Chardon, F. Impact of the genetic–environment interaction on the dynamic of nitrogen pools in arabidopsis. Agriculture 2018, 8, 28. [Google Scholar] [CrossRef] [Green Version]
- North, K.A.; Ehlting, B.; Koprivova, A.; Rennenberg, H.; Kopriva, S. Natural variation in Arabidopsis adaptation to growth at low nitrogen conditions. Plant Physiol. Biochem. 2009, 47, 912–918. [Google Scholar] [CrossRef]
- Menz, J.; Range, T.; Trini, J.; Ludewig, U.; Neuhäuser, B. Molecular basis of differential nitrogen use efficiencies and nitrogen source preferences in contrasting Arabidopsis accessions. Sci. Rep. 2018, 8, 3373. [Google Scholar] [CrossRef] [Green Version]
- Chardon, F.; Barthélémy, J.; Daniel-Vedele, F.; Masclaux-Daubresse, C. Natural variation of nitrate uptake and nitrogen use efficiency in arabidopsis thaliana cultivated with limiting and ample nitrogen supply. J. Exp. Bot. 2010, 61, 2293–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masclaux-Daubresse, C.; Chardon, F. Exploring nitrogen remobilization for seed filling using natural variation in Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 2131–2142. [Google Scholar] [CrossRef] [Green Version]
- Thompson, A.R.; Vierstra, R.D. Autophagic recycling: Lessons from yeast help define the process in plants. Curr. Opin. Plant Biol. 2005, 8, 165–173. [Google Scholar] [CrossRef]
- Li, F.; Vierstra, R.D. Autophagy: A multifaceted intracellular system for bulk and selective recycling. Trends Plant Sci. 2012, 17, 526–537. [Google Scholar] [CrossRef] [PubMed]
- Havé, M.; Marmagne, A.; Chardon, F.; Masclaux-Daubresse, C. Nitrogen remobilization during leaf senescence: Lessons from Arabidopsis to crops. J. Exp. Bot. 2017, 68, 2513–2529. [Google Scholar]
- Slavikova, S.; Ufaz, S.; Avin-Wittenberg, T.; Levanony, H.; Galili, G. An autophagy-associated Atg8 protein is involved in the responses of Arabidopsis seedlings to hormonal controls and abiotic stresses. J. Exp. Bot. 2008, 59, 4029–4043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Xiong, Y.; Bassham, D.C. Autophagy is required for tolerance of drought and salt stress in plants. Autophagy 2009, 5, 954–963. [Google Scholar] [CrossRef] [Green Version]
- Fagard, M.; Launay, A.; Clément, G.; Courtial, J.; Dellagi, A.; Farjad, M.; Krapp, A.; Soulié, M.C.; Masclaux-Daubresse, C.; Clement, G.; et al. Nitrogen metabolism meets phytopathology. J. Exp. Bot. 2014, 65, 5643–5656. [Google Scholar] [CrossRef]
- Mizushima, N.; Yoshimori, T.; Ohsumi, Y. The Role of Atg Proteins in Autophagosome Formation. Annu. Rev. Cell Dev. Biol. 2011, 27, 107–132. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, K.; Hanaoka, H.; Sato, S.; Kato, T.; Tabata, S.; Noda, T.; Ohsumi, Y. Processing of ATG8s, ubiquitin-like proteins, and their deconjugation by ATG4s are essential for plant autophagy. Plant Cell 2004, 16, 2967–2983. [Google Scholar] [CrossRef]
- Hanaoka, H.; Noda, T.; Shirano, Y.; Kato, T.; Hayashi, H.; Shibata, D.; Tabata, S.; Ohsumi, Y. Leaf senescence and starvation-induced chlorosis are accelerated by the disruption of an Arabidopsis autophagy gene. Plant Physiol. 2002, 129, 1181–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doelling, J.H.; Walker, J.M.; Friedman, E.M.; Thompson, A.R.; Vierstra, R.D. The APG8/12-activating enzyme APG7 is required for proper nutrient recycling and senescence in Arabidopsis thaliana. J. Biol. Chem. 2002, 277, 33105–33114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guiboileau, A.; Yoshimoto, K.; Soulay, F.; Bataillé, M.-P.; Avice, J.-C.; Masclaux-Daubresse, C. Autophagy machinery controls nitrogen remobilization at the whole-plant level under both limiting and ample nitrate conditions in Arabidopsis. New Phytol. 2012, 194, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Soulay, F.; Saudemont, B.; Elmayan, T.; Marmagne, A.; Masclaux-Daubresse, C. Overexpression of ATG8 in Arabidopsis Stimulates Autophagic Activity and Increases Nitrogen Remobilization Efficiency and Grain Filling. Plant Cell Physiol. 2019, 60, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Bassham, D.C. TOR Is a Negative Regulator of Autophagy in Arabidopsis thaliana. PLoS ONE 2010, 5, e11883. [Google Scholar] [CrossRef] [Green Version]
- Thompson, A.R.; Doelling, J.H.; Suttangkakul, A.; Vierstra, R.D. Autophagic nutrient recycling in Arabidopsis directed by the ATG8 and ATG12 conjugation pathways. Plant Physiol. 2005, 138, 2097–2110. [Google Scholar] [CrossRef] [Green Version]
- Buchanan-Wollaston, V.; Page, T.; Harrison, E.; Breeze, E.; Lim, P.O.; Nam, H.G.; Lin, J.-F.; Wu, S.-H.; Swidzinski, J.; Ishizaki, K.; et al. Comparative transcriptome analysis reveals significant differences in gene expression and signalling pathways between developmental and dark/starvation-induced senescence in Arabidopsis. Plant J. 2005, 42, 567–585. [Google Scholar] [CrossRef]
- Wang, P.; Nolan, T.M.; Yin, Y.; Bassham, D.C. Identification of transcription factors that regulate ATG8 expression and autophagy in Arabidopsis. Autophagy 2020, 16, 123–139. [Google Scholar] [CrossRef]
- O’Malley, R.C.; Huang, S.S.C.; Song, L.; Lewsey, M.G.; Bartlett, A.; Nery, J.R.; Galli, M.; Gallavotti, A.; Ecker, J.R. Cistrome and Epicistrome Features Shape the Regulatory DNA Landscape. Cell 2016, 165, 1280–1292. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, J.M.; Riveras, E.; Vidal, E.A.; Gras, D.E.; Contreras-López, O.; Tamayo, K.P.; Aceituno, F.; Gómez, I.; Ruffel, S.; Lejay, L.; et al. Systems approach identifies TGA1 and TGA4 transcription factors as important regulatory components of the nitrate response of Arabidopsis thaliana roots. Plant J. 2014, 80, 1–13. [Google Scholar] [CrossRef]
- McKhann, H.I.; Camilleri, C.; Bérard, A.; Bataillon, T.; David, J.L.; Reboud, X.; Le Corre, V.; Caloustian, C.; Gut, I.G.; Brunel, D. Nested core collections maximizing genetic diversity in Arabidopsis thaliana. Plant J. 2004, 38, 193–202. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Zeng, Z.; Liu, C.; Yuan, W.; Hou, N.; Bian, H.; Zhu, M.; Han, N. A barley homolog of yeast ATG6 is involved in multiple abiotic stress responses and stress resistance regulation. Plant Physiol. Biochem. 2017, 115, 97–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avila-Ospina, L.; Marmagne, A.; Soulay, F.; Masclaux-Daubresse, C. Identification of Barley (Hordeum vulgare L.) Autophagy Genes and Their Expression Levels during Leaf Senescence, Chronic Nitrogen Limitation and in Response to Dark Exposure. Agronomy 2016, 6, 15. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Chen, M.; Wang, E.; Hu, L.; Hawkesford, M.J.; Zhong, L.; Chen, Z.; Xu, Z.; Li, L.; Zhou, Y.; et al. Genome-wide analysis of autophagy-associated genes in foxtail millet (Setaria italica L.) and characterization of the function of SiATG8a in conferring tolerance to nitrogen starvation in rice. BMC Genom. 2016, 17, 797. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Chung, T.; Pennington, J.G.; Federico, M.L.; Kaeppler, H.F.; Kaeppler, S.M.; Otegui, M.S. Autophagic recycling plays a central role in maize nitrogen remobilization. Plant Cell 2015, 27, 1389–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Jia, X.; Huo, L.; Che, R.; Gong, X.; Wang, P.; Ma, F. MdATG18a overexpression improves tolerance to nitrogen deficiency and regulates anthocyanin accumulation through increased autophagy in transgenic apple. Plant. Cell Environ. 2018, 41, 469–480. [Google Scholar] [CrossRef]
- Wang, P.; Sun, X.; Jia, X.; Ma, F. Apple autophagy-related protein MdATG3s afford tolerance to multiple abiotic stresses. Plant Sci. 2017, 256, 53–64. [Google Scholar] [CrossRef]
- Wang, P.; Sun, X.; Jia, X.; Wang, N.; Gong, X.; Ma, F. Characterization of an Autophagy-Related Gene MdATG8i from Apple. Front. Plant Sci. 2016, 7, 720. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Sun, X.; Wang, N.; Jia, X.; Ma, F. Ectopic expression of an autophagy-associated MdATG7b gene from apple alters growth and tolerance to nutrient stress in Arabidopsis thaliana. Plant Cell. Tissue Organ Cult. 2017, 128, 9–23. [Google Scholar] [CrossRef]
- Van Janse Rensburg, H.C.; Van den Ende, W.; Signorelli, S. Autophagy in plants: Both a puppet and a puppet master of sugars. Front. Plant Sci. 2019, 10, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Sheen, J. Novel links in the plant TOR kinase signaling network. Curr. Opin. Plant Biol. 2015, 28, 83–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Leene, J.; Han, C.; Gadeyne, A.; Eeckhout, D.; Matthijs, C.; Cannoot, B.; De Winne, N.; Persiau, G.; Van De Slijke, E.; Van de Cotte, B.; et al. Capturing the phosphorylation and protein interaction landscape of the plant TOR kinase. Nat. Plants 2019, 5, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Ueda, Y.; Konishi, M.; Yanagisawa, S. Molecular basis of the nitrogen response in plants. Soil Sci. Plant Nutr. 2017, 63, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Avin-Wittenberg, T.; Michaeli, S.; Honig, A.; Galili, G. ATI1, a newly identified atg8-interacting protein, binds two different Atg8 homologs. Plant Signal. Behav. 2012, 7, 685–687. [Google Scholar] [CrossRef] [Green Version]
- Jia, M.; Liu, X.; Xue, H.; Wu, Y.; Shi, L.; Wang, R.; Chen, Y.; Xu, N.; Zhao, J.; Shao, J.; et al. Noncanonical ATG8–ABS3 interaction controls senescence in plants. Nat. Plants 2019, 5, 212–224. [Google Scholar] [CrossRef]
- Honig, A.; Avin-Wittenberg, T.; Ufaz, S.; Galili, G. A new type of compartment, defined by plant-specific Atg8-interacting proteins, is induced upon exposure of Arabidopsis plants to carbon starvation. Plant Cell 2012, 24, 288–303. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Tian, Z.; Li, Y.; Hua, X.; Zhang, D.; Li, J.; Jin, H.; Xu, J.; Chen, W.; Niu, B.; et al. ATG7 Promotes Bladder Cancer Invasion via Autophagy-Mediated Increased ARHGDIB mRNA Stability. Adv. Sci. 2019, 6, 1801927. [Google Scholar] [CrossRef] [Green Version]
- Marshall, R.S.; Vierstra, R.D. Autophagy: The Master of Bulk and Selective Recycling. Annu. Rev. Plant Biol. 2018, 69, 173–208. [Google Scholar] [CrossRef]
- Devarenne, T.P. The plant cell death suppressor Adi3 interacts with the autophagic protein Atg8h. Biochem. Biophys. Res. Commun. 2011, 412, 699–703. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Clément, G.; Anne, P.; Routaboul, J.M.; Guiboileau, A.; Soulay, F.; Shirasu, K.; Yoshimoto, K. Stitching together the multiple dimensions of autophagy using metabolomics and transcriptomics reveals impacts on metabolism, development, and plant responses to the environment in Arabidopsis. Plant Cell 2014, 26, 1857–1877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, A.R.; Suttangkakul, A.; Vierstra, R.D. The ATG12-conjugating enzyme ATG10 is essential for autophagic vesicle formation in Arabidopsis thaliana. Genetics 2008, 178, 1339–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rioux, J.D.; Xavier, R.J.; Taylor, K.D.; Silverberg, M.S.; Goyette, P.; Huett, A.; Green, T.; Kuballa, P.; Barmada, M.M.; Datta, L.W.; et al. Genome-wide association study identifies new susceptibility loci for Crohn disease and implicates autophagy in disease pathogenesis. Nat. Genet. 2007, 39, 596–604. [Google Scholar] [CrossRef]
- Balla, K.M.; Lažetić, V.; Troemel, E.R. Natural variation in the roles of C. elegans autophagy components during microsporidia infection. PLoS ONE 2019, 14, e0216011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Blanco, C.; Andrade, J.; Becker, C.; Bemm, F.; Bergelson, J.; Borgwardt, K.M.M.; Cao, J.; Chae, E.; Dezwaan, T.M.M.; Ding, W.; et al. 1,135 Genomes Reveal the Global Pattern of Polymorphism in Arabidopsis thaliana. Cell 2016, 166, 481–491. [Google Scholar] [CrossRef] [Green Version]
- Meyer, R.C.; Gryczka, C.; Neitsch, C.; Müller, M.; Bräutigam, A.; Schlereth, A.; Schön, H.; Weigelt-Fischer, K.; Altmann, T. Genetic diversity for nitrogen use efficiency in Arabidopsis thaliana. Planta 2019, 250, 41–57. [Google Scholar] [CrossRef] [Green Version]
- Canales, J.; Moyano, T.C.; Villarroel, E.; Gutiérrez, R.A. Systems analysis of transcriptome data provides new hypotheses about Arabidopsis root response to nitrate treatments. Front. Plant Sci. 2014, 5, 22. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | AGI Code | Control | Starvation | Differential Variation | p-Value | Significant Difference | ||
|---|---|---|---|---|---|---|---|---|
| Mean | Sd | Mean | Sd | |||||
| NRT2.1 | At1g08090 | 0.011 | 0.007 | 0.009 | 0.005 | −19.0% | 0.050 | ns |
| NRT1.1 | At1g12110 | 0.119 | 0.038 | 0.117 | 0.044 | −1.0% | 0.861 | ns |
| NIA2 | At1g37130 | 1.651 | 0.789 | 1.541 | 0.743 | −6.7% | 0.398 | ns |
| Nii | At2g15620 | 1.817 | 0.560 | 2.055 | 0.541 | 13.1% | 0.013 | * |
| GS2 | At5g35630 | 4.844 | 1.492 | 5.162 | 1.443 | 6.6% | 0.204 | ns |
| GLN1.1 | At5g37600 | 0.335 | 0.318 | 0.409 | 0.272 | 21.9% | 0.146 | ns |
| AMT1.1 | At4g13510 | 0.156 | 0.100 | 0.165 | 0.101 | 5.9% | 0.591 | ns |
| AMT1.4 | At4g28700 | 0.003 | 0.002 | 0.004 | 0.002 | 19.5% | 0.104 | ns |
| GDH1 | At5g18170 | 0.066 | 0.030 | 0.079 | 0.022 | 19.4% | 0.005 | ** |
| GLU1 | At5g04140 | 6.208 | 1.835 | 6.972 | 2.141 | 12.3% | 0.026 | * |
| GLT | At5g16150 | 0.284 | 0.110 | 0.372 | 0.139 | 31.0% | 0.000 | *** |
| Gene Name | AGI Code | Control | Starvation | Differential Variation | p-Value | Significant Difference | ||
|---|---|---|---|---|---|---|---|---|
| Mean | Sd | Mean | Sd | |||||
| ATG1a | At3g61960 | 5.727 | 1.943 | 7.841 | 2.455 | 36.9% | 0.000 | *** |
| ATG1b | At3g53930 | 0.034 | 0.008 | 0.033 | 0.010 | −3.7% | 0.395 | ns |
| ATG1c | At2g37840 | 0.157 | 0.088 | 0.215 | 0.083 | 36.7% | 0.000 | *** |
| ATG3 | At5g61500 | 0.368 | 0.125 | 0.509 | 0.117 | 38.4% | 0.000 | *** |
| ATG4a | At2g44140 | 0.053 | 0.019 | 0.063 | 0.016 | 18.9% | 0.002 | ** |
| ATG4b | At3g59950 | 0.117 | 0.046 | 0.164 | 0.036 | 40.0% | 0.000 | *** |
| ATG5 | At5g17290 | 0.047 | 0.016 | 0.063 | 0.019 | 33.9% | 0.000 | *** |
| ATG7 | At5g45900 | 8.386 | 2.284 | 8.830 | 2.523 | 5.3% | 0.280 | ns |
| ATG8a | At4g21980 | 0.500 | 0.222 | 0.702 | 0.244 | 40.3% | 0.000 | *** |
| ATG8b | At4g04620 | 22.159 | 10.227 | 22.614 | 13.182 | 2.1% | 0.820 | ns |
| ATG8c | At1g62040 | 65.795 | 34.432 | 73.977 | 39.545 | 12.4% | 0.196 | ns |
| ATG8d | At2g05630 | 66.023 | 51.705 | 71.989 | 46.136 | 9.0% | 0.474 | ns |
| ATG8e | At2g45170 | 0.060 | 0.044 | 0.069 | 0.040 | 15.3% | 0.199 | ns |
| ATG8f | At4g16520 | 214.773 | 63.636 | 280.682 | 79.545 | 30.7% | 0.000 | *** |
| ATG8g | At3g60640 | 24.148 | 8.381 | 32.386 | 11.648 | 34.1% | 0.000 | *** |
| ATG8h | At3g06420 | 15.341 | 7.273 | 24.545 | 9.659 | 60.0% | 0.000 | *** |
| ATG8i | At3g15580 | 0.733 | 0.284 | 1.097 | 0.386 | 49.6% | 0.000 | *** |
| ATG9 | At2g31260 | 0.065 | 0.036 | 0.106 | 0.043 | 62.6% | 0.000 | *** |
| ATG10 | At3g07525 | 0.035 | 0.013 | 0.043 | 0.025 | 22.6% | 0.020 | * |
| ATG12a | At1g54210 | 0.189 | 0.136 | 0.297 | 0.184 | 56.8% | 0.000 | *** |
| ATG13a | At3g49590 | 0.247 | 0.044 | 0.374 | 0.053 | 51.1% | 0.000 | *** |
| ATG18a | At3g62770 | 0.203 | 0.099 | 0.277 | 0.108 | 36.4% | 0.000 | *** |
| ATG18f | At5g54730 | 0.107 | 0.056 | 0.162 | 0.069 | 52.0% | 0.000 | *** |
| ATG18h | At1g54710 | 0.188 | 0.090 | 0.238 | 0.093 | 26.7% | 0.002 | ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bedu, M.; Marmagne, A.; Masclaux-Daubresse, C.; Chardon, F. Transcriptional Plasticity of Autophagy-Related Genes Correlates with the Genetic Response to Nitrate Starvation in Arabidopsis Thaliana. Cells 2020, 9, 1021. https://doi.org/10.3390/cells9041021
Bedu M, Marmagne A, Masclaux-Daubresse C, Chardon F. Transcriptional Plasticity of Autophagy-Related Genes Correlates with the Genetic Response to Nitrate Starvation in Arabidopsis Thaliana. Cells. 2020; 9(4):1021. https://doi.org/10.3390/cells9041021
Chicago/Turabian StyleBedu, Magali, Anne Marmagne, Céline Masclaux-Daubresse, and Fabien Chardon. 2020. "Transcriptional Plasticity of Autophagy-Related Genes Correlates with the Genetic Response to Nitrate Starvation in Arabidopsis Thaliana" Cells 9, no. 4: 1021. https://doi.org/10.3390/cells9041021
APA StyleBedu, M., Marmagne, A., Masclaux-Daubresse, C., & Chardon, F. (2020). Transcriptional Plasticity of Autophagy-Related Genes Correlates with the Genetic Response to Nitrate Starvation in Arabidopsis Thaliana. Cells, 9(4), 1021. https://doi.org/10.3390/cells9041021