Organoids of Human Endometrium: A Powerful In Vitro Model for the Endometrium-Embryo Cross-Talk at the Implantation Site

 , , ,
, , ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Collection
2.2. Isolation and Culture of Endometrial Glandular Organoids
2.3. Human Endometrial Stromal Cells Culture
2.4. Hormonal Treatments
2.5. Transmission Electron Microscopy (TEM)
2.6. Scanning Electron Microscopy (SEM)
2.7. RNA Extraction and Droplet Digital PCR (dd-PCR) Analysis
2.8. Protein Extraction
2.9. SDS-PAGE (Sodium Dodecil Sulphate-Polyacrylammide Gel Electrophoresis)
2.10. Two-Dimensional Electrophoresis (2-D Electrophoresis)
2.11. Western Blotting
2.12. Immunofluorescence
2.13. Statistical Analysis
3. Results
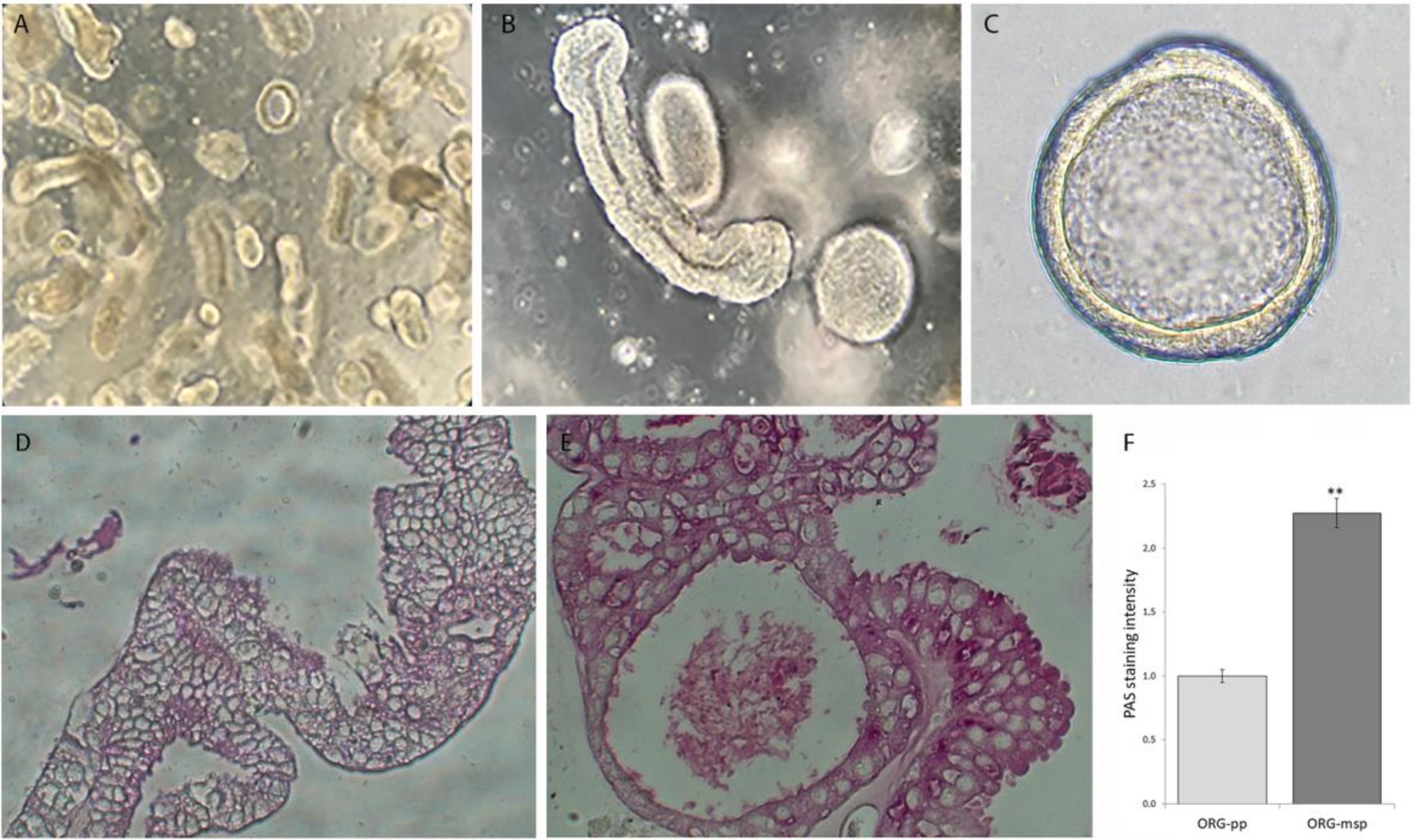
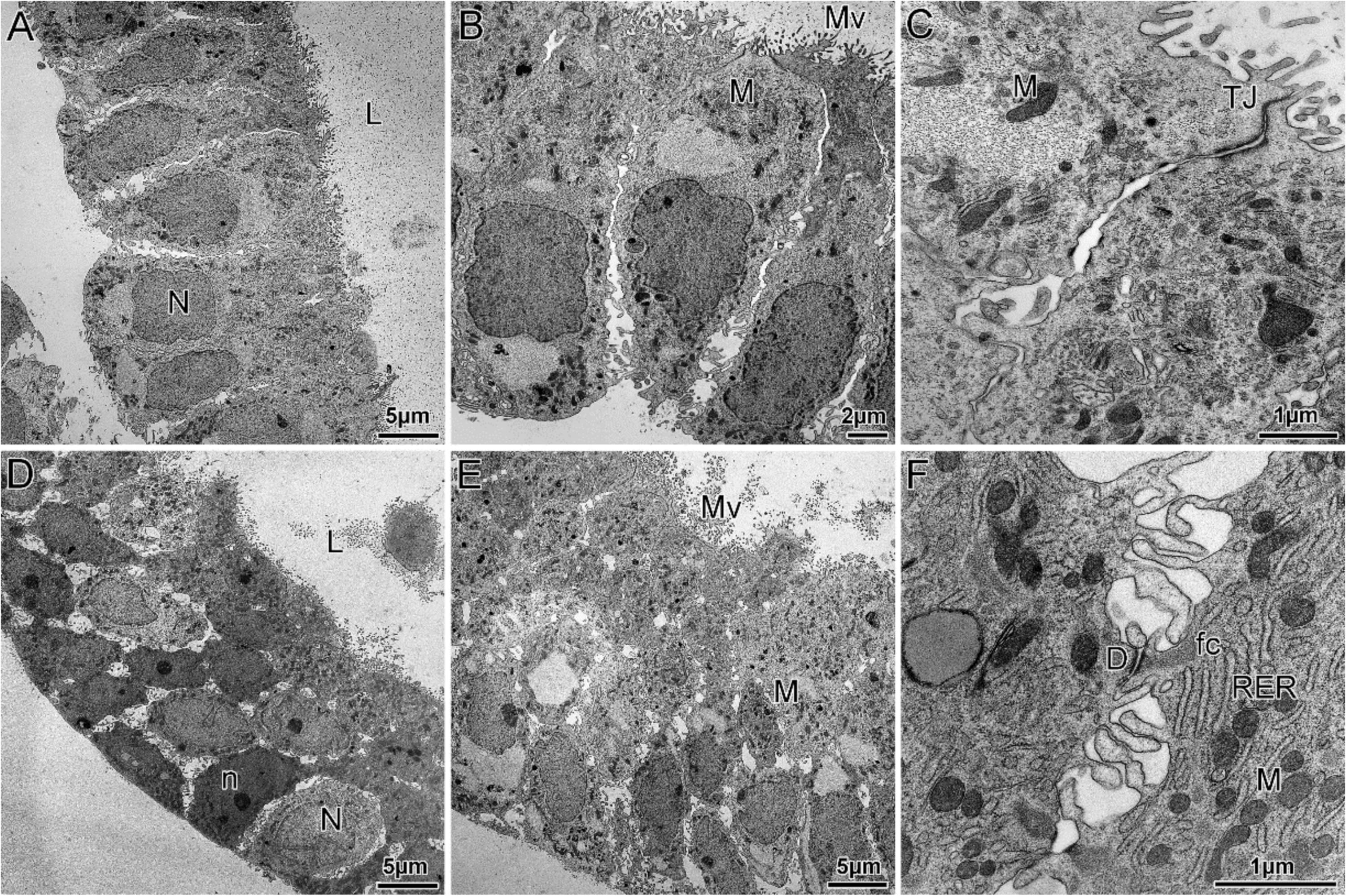
3.1. Established Human Endometrial Epithelial Organoids Respond to Hormones by Recapitulating Morphological and Ultrastructural Features of the Different Phases of the Uterine Cycle
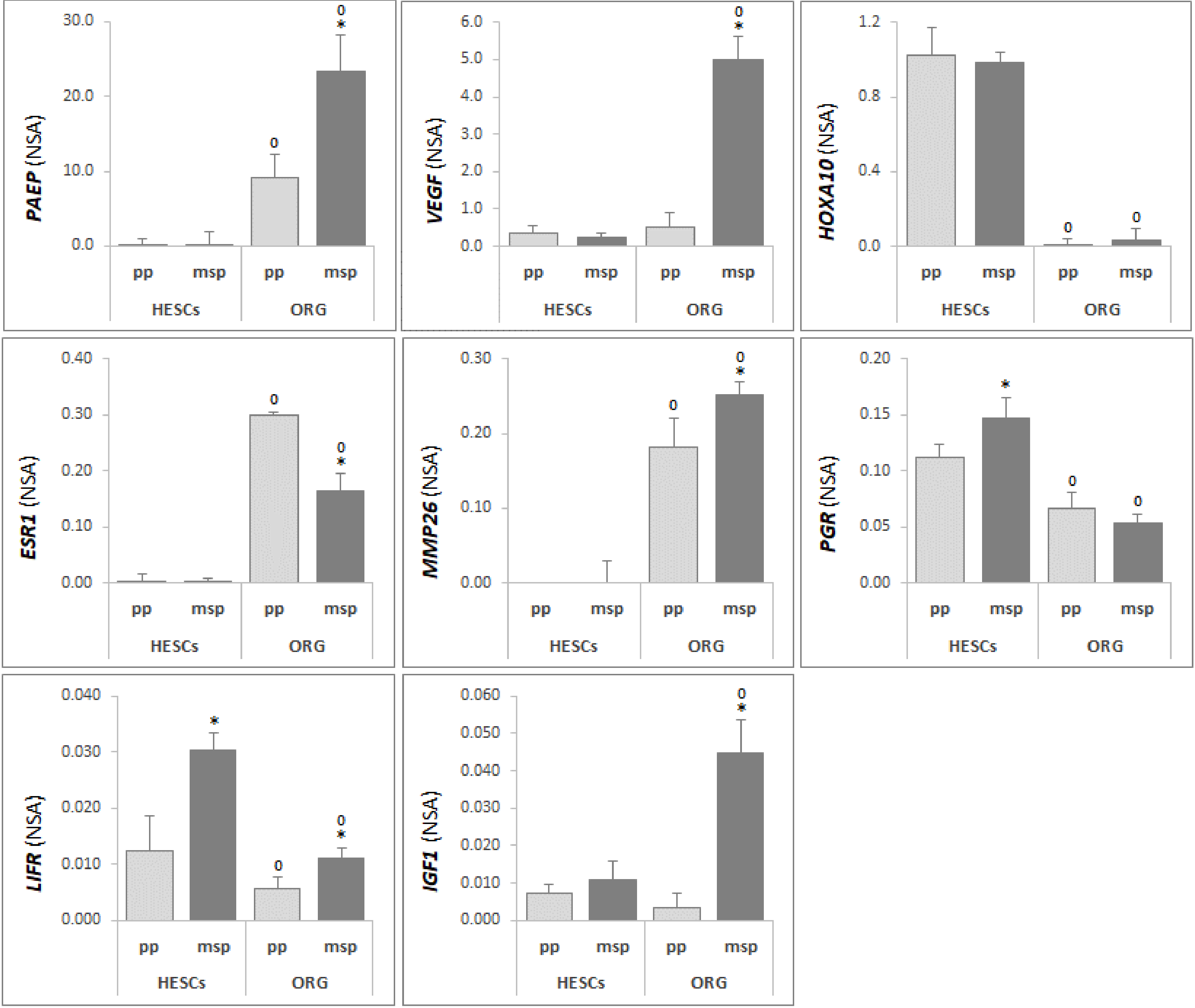
3.2. Differential Gene Expression Profile Between Human Endometrial Epithelial Organoids and Stromal Cells Reflects Tissue Heterogeneity
3.3. Human Endometrial Organoids Recapitulate the Glycodelin (GdA) Molecular Signature In Vivo, a Distinguishing Feature of Different Phases of the Uterine Cycle
3.4. Organoids Isolated from Eutopic Endometrium of Endometriosis-Affected Women, Recapitulate Disease Traits
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Douglas, A.J. Mother-offspring dialogue in early pregnancy: Impact of adverse environment on pregnancy maintenance and neurobiology. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2011, 35, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- Meseguer, M.; Aplin, J.; Caballero-Campo, P.; O’Connor, J.E.; Martín, J.C.; Remohí, J.; Pellicer, A.; Simón, C. Human endometrial mucin MUC1 is up-regulated by progesterone and down-regulated in vitro by the human blastocyst. Biol. Reprod. 2001, 64, 590–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simón, C.; Martin, J.C.; Meseguer, M.; Caballero-Campo, P.; Valbuena, D.; Pellicer, A.; Perilla, D.V. Embryonic regulation of endometrial molecules in human implantation. J. Reprod. Fertil. Suppl. 2000, 55, 43–53. [Google Scholar] [PubMed]
- Boj, S.F.; Hwang, C.-I.; Baker, L.A.; Chio, I.I.C.; Engle, D.D.; Corbo, V.; Jager, M.; Ponz-Sarvise, M.; Tiriac, H.; Spector, M.S.; et al. Organoid models of human and mouse ductal pancreatic cancer. Cell 2014, 160, 324–338. [Google Scholar] [CrossRef] [Green Version]
- Morgan, J.P.; Delnero, P.F.; Zheng, Y.; Verbridge, S.S.; Chen, J.; Craven, M.; Choi, N.; Diaz-Santana, A.; Kermani, P.; Hempstead, B.; et al. Formation of microvascular networks In Vitro. Nat. Protoc. 2013, 8, 1820–1836. [Google Scholar] [CrossRef]
- Quadrato, G.; Nguyen, T.; Macosko, E.Z.; Sherwood, J.; Yang, S.M.; Berger, D.R.; Maria, N.; Scholvin, J.; Goldman, M.; Kinney, J.P.; et al. Cell diversity and network dynamics in photosensitive human brain organoids. Nature 2017, 545, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Vries, R.G.; Snippert, H.J.; Van De Wetering, M.; Barker, N.; Stange, D.; Van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef]
- Takebe, T.; Sekine, K.; Enomura, M.; Koike, H.; Kimura, M.; Ogaeri, T.; Zhang, R.; Ueno, Y.; Zheng, Y.-W.; Koike, N.; et al. Vascularized and functional human liver from an iPSC-derived organ bud transplant. Nature 2013, 499, 481–484. [Google Scholar] [CrossRef]
- Boretto, M.; Cox, B.; Noben, M.; Hendriks, N.; Fassbender, A.; Roose, H.; Amant, F.; Timmerman, D.; Tomassetti, C.; Vanhie, A.; et al. Development of organoids from mouse and human endometrium showing endometrial epithelium physiology and long-term expandability. Development 2017, 144, 1775–1786. [Google Scholar] [CrossRef] [Green Version]
- Turco, M.Y.; Gardner, L.; Hughes, J.; Cindrova-Davies, T.; Gomez, M.J.; Farrell, L.E.; Hollinshead, M.; Marsh, S.G.E.; Brosens, J.J.; Critchley, H.O.; et al. Long-term, hormone-responsive organoid cultures of human endometrium in a chemically defined medium. Nature 2017, 19, 568–577. [Google Scholar] [CrossRef]
- Valentina, P.; Ietta, F.; Luddi, A.; Semplici, B.; Marrocco, C.; Luisi, S.; Piomboni, P. Organoids of human endometrium: Is this a powerful 3D culture model in replicating the endometrial glandular epithelium? Hum. Reprod. 2019, 34, 293. [Google Scholar]
- Fitzgerald, H.C.; Dhakal, P.; Behura, S.K.; Schust, D.J.; Spencer, T.E. Self-renewing endometrial epithelial organoids of the human uterus. Proc. Natl. Acad. Sci. USA 2019, 116, 23132–23142. [Google Scholar] [CrossRef] [PubMed]
- Boretto, M.; Maenhoudt, N.; Luo, X.; Hennes, A.; Boeckx, B.; Bui, B.; Heremans, R.; Perneel, L.; Kobayashi, H.; Van Zundert, I.; et al. Patient-derived organoids from endometrial disease capture clinical heterogeneity and are amenable to drug screening. Nature 2019, 21, 1041–1051. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.; Scioscia, M.; Rademacher, T. Endometrial secretions: Creating a stimulatory microenvironment within the human early placenta and implications for the aetiopathogenesis of preeclampsia. J. Reprod. Immunol. 2011, 89, 118–125. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E.; Charnock-Jones, D.S. Human Early Placental Development: Potential Roles of the Endometrial Glands. Placenta 2007, 28, S64–S69. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.; Sun, X.; Dey, S.K. Mechanisms of implantation: Strategies for successful pregnancy. Nat. Med. 2012, 18, 1754–1767. [Google Scholar] [CrossRef]
- Mylonas, I.; Jeschke, U.; Kunert-Keil, C.; Shabani, N.; Dian, D.; Bauerfeind, I.; Kuhn, C.; Kupka, M.S.; Friese, K. Glycodelin A is expressed differentially in normal human endometrial tissue throughout the menstrual cycle as assessed by immunohistochemistry and in situ hybridization. Fertil. Steril. 2006, 86, 1488–1497. [Google Scholar] [CrossRef]
- Lapidot, T.; Sharon, N. Meet the multifunctional and sexy glycoforms of glycodelin. Glycobiology 2005, 16, 39R–45R. [Google Scholar] [CrossRef] [Green Version]
- Julkunen, M.; Rutanen, E.; Koskimies, A.; Ranta, T.; Bohn, H.; Seppälä, M. Distribution of placental protein 14 in tissues and body fluids during pregnancy. BJOG Int. J. Obstet. Gynaecol. 1985, 92, 1145–1151. [Google Scholar] [CrossRef]
- Seppälä, M.; Koistinen, H.; Chiu, P.C.N.; Yeung, W.S. Glycosylation related actions of glycodelin: Gamete, cumulus cell, immune cell and clinical associations. Hum. Reprod. Updat. 2007, 13, 275–287. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Liu, Y.; Wang, X. The Roles of Glycodelin in Cancer Development and Progression. Front. Immunol. 2017, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.; Lam, K.K.W.; Vijayan, M.; Koistinen, H.; Seppälä, M.; Ng, E.H.; Yeung, W.S.; Chiu, P.C.N. The Pleiotropic Effect of Glycodelin-A in Early Pregnancy. Am. J. Reprod. Immunol. 2016, 75, 290–297. [Google Scholar] [CrossRef] [Green Version]
- Uchida, H.; Maruyama, T.; Nishikawa-Uchida, S.; Miyazaki, K.; Masuda, H.; Yoshimura, Y. Glycodelin in reproduction. Reprod. Med. Biol. 2013, 12, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, U.; Richter, D.U.; Möbius, B.-M.; Briese, V.; Mylonas, I.; Friese, D.M.K. Stimulation of progesterone, estradiol and cortisol in trophoblast tumor bewo cells by glycodelin A N-glycans. Anticancer Res. 2007, 27, 2101–2108. [Google Scholar] [PubMed]
- Lam, K.; Pang, R.; Lee, C.-L.; Koistinen, H.; Seppälä, M.; Ho, P.-C.; Yeung, W.S.; Chiu, P.C.N. Glycodelin-A modulates syncytialization of human BeWo choriocarcinoma cell line. Placenta 2012, 33, 750–752. [Google Scholar] [CrossRef] [PubMed]
- Focarelli, R.; Luddi, A.; De Leo, V.; Capaldo, A.; Stendardi, A.; Pavone, V.; Benincasa, L.; Belmonte, G.; Petraglia, F.; Piomboni, P. Dysregulation of GdA Expression in Endometrium of Women With Endometriosis: Implication for Endometrial Receptivity. Reprod. Sci. 2017, 25, 579–586. [Google Scholar] [CrossRef]
- Viganò, P.; Parazzini, F.; Somigliana, E.; Vercellini, P. Endometriosis: Epidemiology and aetiological factors. Best Pr. Res. Clin. Obstet. Gynaecol. 2004, 18, 177–200. [Google Scholar] [CrossRef]
- Luddi, A.; Marrocco, C.; Governini, L.; Semplici, B.; Pavone, V.; Capaldo, A.; Tosti, C.; Greco, S.; Luisi, S.; Ciarmela, P.; et al. Increased expression of neurogenic factors in uterine fibroids. Hum. Reprod. 2019, 34, 2153–2162. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, S.L. Oestrogen and progesterone action on endometrium: A translational approach to understanding endometrial receptivity. Reprod. Biomed. Online 2013, 27, 497–505. [Google Scholar] [CrossRef] [Green Version]
- Acosta, A.A.; Elberger, L.; Borghi, M.; Calamera, J.C.; Chemes, H.; Doncel, G.F.; Kliman, H.J.; Lema, B.; Lustig, L.; Papier, S. Endometrial dating and determination of the window of implantation in healthy fertile women. Fertil. Steril. 2000, 73, 788–798. [Google Scholar] [CrossRef]
- Rarani, F.Z.; Borhani, F.; Rashidi, B. Endometrial pinopode biomarkers: Molecules and microRNAs. J. Cell. Physiol. 2018, 233, 9145–9158. [Google Scholar] [CrossRef] [PubMed]
- Melkozerova, O.A.; Bashmakova, N.V.; Malgina, G.B.; Bragina, E.E.; Michelson, A.A.; Chistyakova, G.N. Ultrastructural markers of tissue endometrial receptivity in patients with recurrent implantation failure. Gynecol. Endocrinol. 2019, 35, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Suhorutshenko, M.; Kukushkina, V.; Velthut-Meikas, A.; Altmäe, S.; Peters, M.; Mägi, R.; Krjutškov, K.; Koel, M.; Codoñer, F.M.; Martinez-Blanch, J.F.; et al. Endometrial receptivity revisited: Endometrial transcriptome adjusted for tissue cellular heterogeneity. Hum. Reprod. 2018, 33, 2074–2086. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Ma, L.; Ma, W.G.; Maas, R.L.; Dey, S.K. Hoxa-10 regulates uterine stromal cell responsiveness to progesterone during implantation and decidualization in the mouse. Mol. Endocrinol. 1999, 13, 1005–1017. [Google Scholar] [CrossRef]
- Shuya, L.L.; Menkhorst, E.M.; Yap, J.; Li, P.; Lane, N.; Dimitriadis, E. Leukemia Inhibitory Factor Enhances Endometrial Stromal Cell Decidualization in Humans and Mice. PLoS ONE 2011, 6, e25288. [Google Scholar] [CrossRef] [Green Version]
- Altmäe, S.; Reimand, J.; Hovatta, O.; Zhang, P.; Kere, J.; Laisk-Podar, T.; Saare, M.; Peters, M.; Vilo, J.; Stavreus-Evers, A.; et al. Research Resource: Interactome of Human Embryo Implantation: Identification of Gene Expression Pathways, Regulation, and Integrated Regulatory Networks. Mol. Endocrinol. 2011, 26, 203–217. [Google Scholar] [CrossRef]
- Teh, W.T.; McBain, J.; Rogers, P. What is the contribution of embryo-endometrial asynchrony to implantation failure? J. Assist. Reprod. Genet. 2016, 33, 1419–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clevers, H. Modeling Development and Disease with Organoids. Cell 2016, 165, 1586–1597. [Google Scholar] [CrossRef] [Green Version]
- Haider, S.; Gamperl, M.; Burkard, T.R.; Kunihs, V.; Kaindl, U.; Junttila, S.; Fiala, C.; Schmidt, K.; Mendjan, S.; Knöfler, M.; et al. Estrogen Signaling Drives Ciliogenesis in Human Endometrial Organoids. Endocrinology 2019, 160, 2282–2297. [Google Scholar] [CrossRef]
- Bahar, L.; Kahraman, S.; Eras, N.; Pirkevi, C. Comparison of endometrial biopsies of fertile women and women with repeated implantation failure at the ultrastructural level. Turk. J. Med Sci. 2015, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masterton, R.; Armstrong, E.M.; More, I.A.R. The Cyclical Variation in the Percentage of Ciliated Cells in the Normal Human Endometrium. Reproduction 1975, 42, 537–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marquardt, R.; Kim, T.H.; Shin, J.-H.; Jeong, J.-W. Progesterone and Estrogen Signaling in the Endometrium: What Goes Wrong in Endometriosis? Int. J. Mol. Sci. 2019, 20, 3822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilka, R.; Whatling, C.; Domanski, H.; Hansson, S.; Eriksson, P.; Casslén, B. Epithelial expression of matrix metalloproteinase-26 is elevated at mid-cycle in the human endometrium. Mol. Hum. Reprod. 2003, 9, 271–277. [Google Scholar] [CrossRef]
- Kocbek, V.; Vouk, K.; Bersinger, N.A.; Mueller, M.D.; Rižner, T.L. Panels of Cytokines and Other Secretory Proteins as Potential Biomarkers of Ovarian Endometriosis. J. Mol. Diagn. 2015, 17, 325–334. [Google Scholar] [CrossRef]
- Kocbek, V.; Vouk, K.; Mueller, M.D.; Rizner, T.L.; Bersinger, N.A. Elevated glycodelin-A concentrations in serum and peritoneal fluid of women with ovarian endometriosis. Gynecol. Endocrinol. 2013, 29, 455–459. [Google Scholar] [CrossRef]
- Kao, L.C.; Germeyer, A.; Tulac, S.; Lobo, S.; Yang, J.P.; Taylor, R.N.; Osteen, K.; Lessey, B.A.; Giudice, L.C. Expression Profiling of Endometrium from Women with Endometriosis Reveals Candidate Genes for Disease-Based Implantation Failure and Infertility. Endocrinology 2003, 144, 2870–2881. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luddi, A.; Pavone, V.; Semplici, B.; Governini, L.; Criscuoli, M.; Paccagnini, E.; Gentile, M.; Morgante, G.; De Leo, V.; Belmonte, G.; et al. Organoids of Human Endometrium: A Powerful In Vitro Model for the Endometrium-Embryo Cross-Talk at the Implantation Site. Cells 2020, 9, 1121. https://doi.org/10.3390/cells9051121
Luddi A, Pavone V, Semplici B, Governini L, Criscuoli M, Paccagnini E, Gentile M, Morgante G, De Leo V, Belmonte G, et al. Organoids of Human Endometrium: A Powerful In Vitro Model for the Endometrium-Embryo Cross-Talk at the Implantation Site. Cells. 2020; 9(5):1121. https://doi.org/10.3390/cells9051121
Chicago/Turabian StyleLuddi, Alice, Valentina Pavone, Bianca Semplici, Laura Governini, Mattia Criscuoli, Eugenio Paccagnini, Mariangela Gentile, Giuseppe Morgante, Vincenzo De Leo, Giuseppe Belmonte, and et al. 2020. "Organoids of Human Endometrium: A Powerful In Vitro Model for the Endometrium-Embryo Cross-Talk at the Implantation Site" Cells 9, no. 5: 1121. https://doi.org/10.3390/cells9051121
APA StyleLuddi, A., Pavone, V., Semplici, B., Governini, L., Criscuoli, M., Paccagnini, E., Gentile, M., Morgante, G., De Leo, V., Belmonte, G., Zarovni, N., & Piomboni, P. (2020). Organoids of Human Endometrium: A Powerful In Vitro Model for the Endometrium-Embryo Cross-Talk at the Implantation Site. Cells, 9(5), 1121. https://doi.org/10.3390/cells9051121