Adenosine A2AReceptors in Substance Use Disorders: A Focus on Cocaine

 , ,
, ,  , and
, and 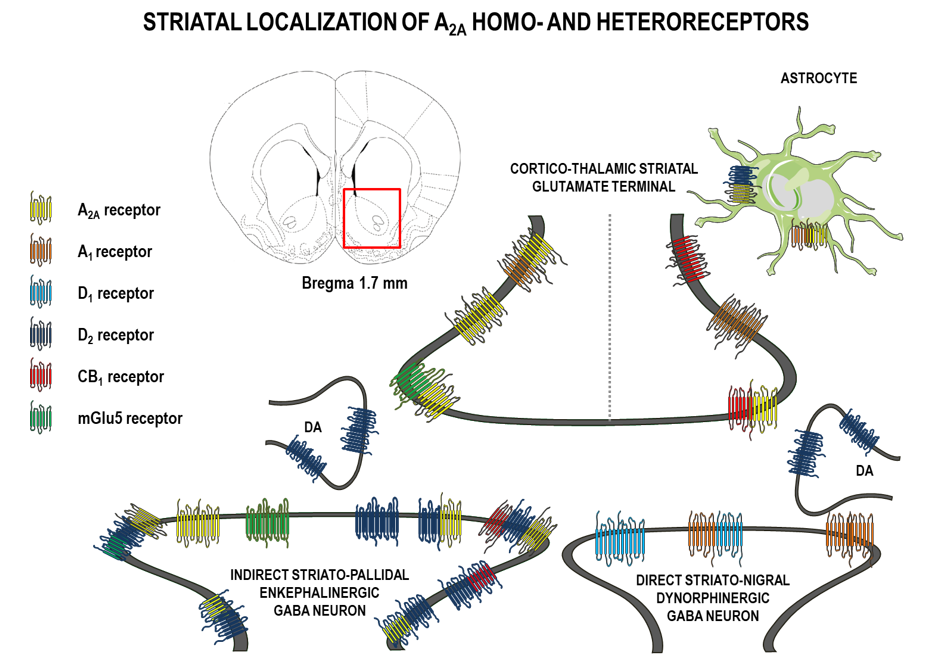{kind=link}
{kind=link}
Abstract
:1. Introduction
2. A2ARs and Their Localization in the Brain
- homodimeric A2AR-A2AR complexes on the cell surface, as shown using BRETassays [31];
- The first recognized and best known heteromeric interaction was found between the A2ARs and D2 receptors, by which the activation of A2A (Gs/olf-coupled) receptors interfered with coupling of D2 receptors to Gi isoforms of G proteins [54,55,56]. A2A-D2 heteroreceptor complexes with antagonistic receptor–receptor interaction were found to exist on:
- the cortico–striatal glutamate nerve terminals, where the A2AR protomer inhibited the D2 receptor protomer-induced inhibition of glutamate release [47];
- striatal cholinergic interneurons [57];and
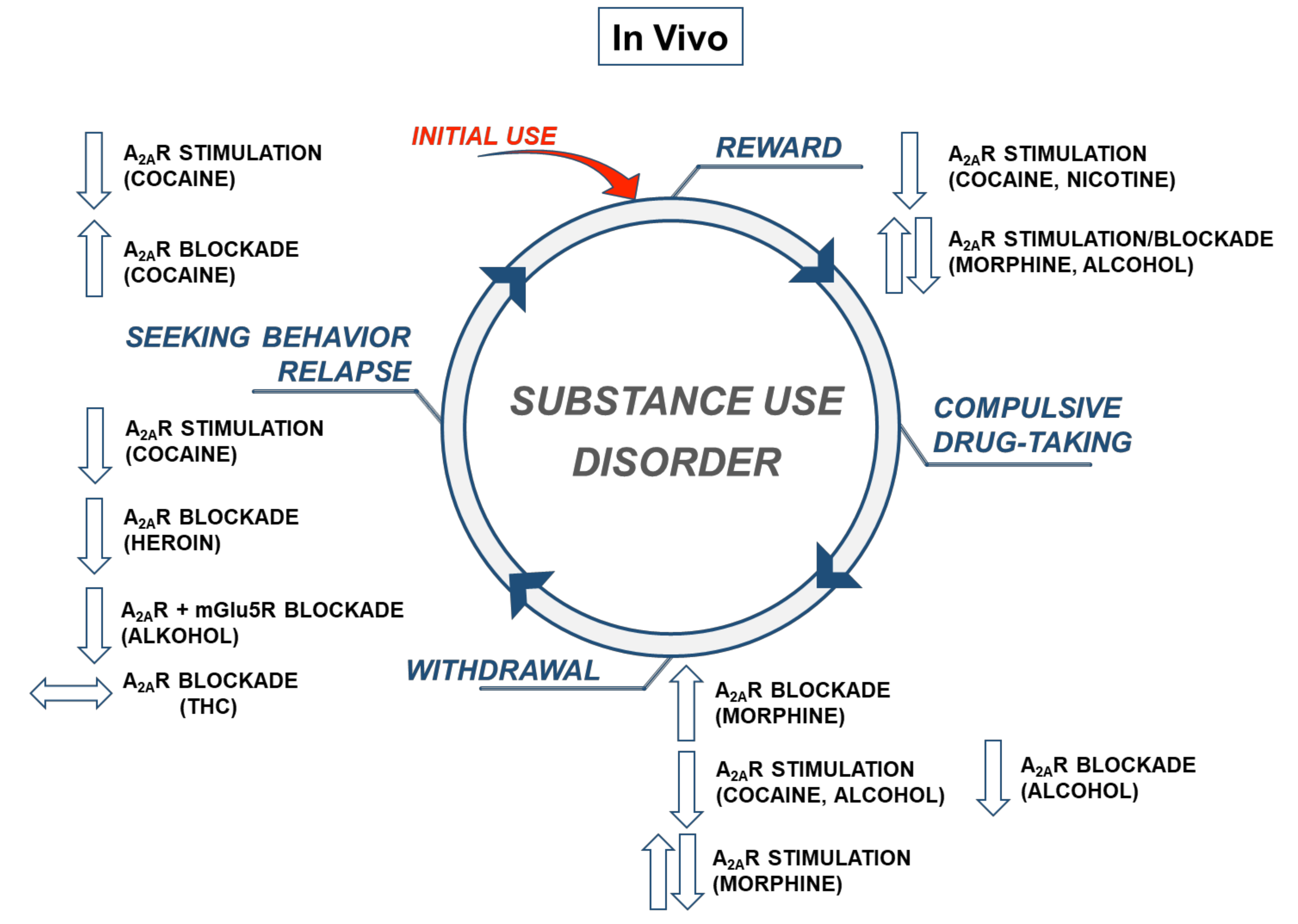
3. A2AR and Behavioral Actions to Cocaine and Other Drugs of Abuse—In Vivo Evidence
3.1. Locomotor Responses to Drugs of Abuse
3.2. Discrimination
3.3. Reward
3.4. Seeking Behavior
3.5. Withdrawal
4. A2AR and Behavioral Actions to Cocaine and Other Drugs of Abuse—Ex Vivo Evidence
5. Neurochemical Correlates to Behavioral Findings in Rat Models of Substance Use Disorder. A Focus on Cocaine
5.1. New Mechanisms of Acute Cocaine
5.2. Cocaine Self-Administration
5.3. Cocaine-Seeking
5.4. The Role of the A2AR-D2Receptor Heterocomplexes in Cocaine Use and Addiction
6. Final Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- McLellan, A.T. Substance Misuse and Substance use Disorders: Why do they Matter in Healthcare? Trans. Am. Clin. Climatol. Assoc. 2017, 128, 112–130. [Google Scholar] [PubMed]
- Bennett, S.; Petrash, P. The Neurobiology of Substance Use Disorders: Information for Assessment and Clinical Treatment. Smith Coll. Stud. Soc. 2014, 84, 273–291. [Google Scholar] [CrossRef]
- Hasin, D.S.; O’Brien, C.P.; Auriacombe, M.; Borges, G.; Bucholz, K.; Budney, A.; Compton, W.M.; Crowley, T.; Ling, W.; Petry, N.M.; et al. DSM-5 Criteria for Substance Use Disorders: Recommendations and Rationale. Am. J. Psychiatry 2013, 170, 834–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha, R.A.; Johansson, B.; van der Ploeg, I.; Sebastiao, A.M.; Ribeiro, J.A.; Fredholm, B.B. Evidence for functionally important adenosine A2a receptors in the rat hippocampus. Brain Res. 1994, 649, 208–216. [Google Scholar] [CrossRef]
- Dixon, A.K.; Gubitz, A.K.; Sirinathsinghji, D.J.; Richardson, P.J.; Freeman, T.C. Tissue distribution of adenosine receptor mRNAs in the rat. Br. J. Pharmacol. 1996, 118, 1461–1468. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, M.F.; Williams, M. Direct autoradiographic localization of adenosine A2 receptors in the rat brain using the A2-selective agonist, [3H]CGS 21680. Eur. J. Pharmacol. 1989, 168, 243–246. [Google Scholar] [CrossRef]
- Rebola, N.; Canas, P.M.; Oliveira, C.R.; Cunha, R.A. Different synaptic and subsynaptic localization of adenosine A2A receptors in the hippocampus and striatum of the rat. Neuroscience 2005, 132, 893–903. [Google Scholar] [CrossRef]
- Rosin, D.L.; Robeva, A.; Woodard, R.L.; Guyenet, P.G.; Linden, J. Immunohistochemical localization of adenosine A2A receptors in the rat central nervous system. J. Comp. Neurol. 1998, 401, 163–186. [Google Scholar] [CrossRef]
- Rosin, D.L.; Hettinger, B.D.; Lee, A.; Linden, J. Anatomy of adenosine A2A receptors in brain: Morphological substrates for integration of striatal function. Neurology 2003, 61, S12–S18. [Google Scholar] [CrossRef]
- Svenningsson, P.; Le Moine, C.; Aubert, I.; Burbaud, P.; Fredholm, B.B.; Bloch, B. Cellular distribution of adenosine A2A receptor mRNA in the primate striatum. J. Comp. Neurol. 1998, 399, 229–240. [Google Scholar] [CrossRef]
- Svenningsson, P.; Le Moine, C.; Fisone, G.; Fredholm, B.B. Distribution, biochemistry and function of striatal adenosine A2A receptors. Prog. Neurobiol. 1999, 59, 355–396. [Google Scholar] [CrossRef]
- Peterfreund, R.A.; MacCollin, M.; Gusella, J.; Fink, J.S. Characterization and expression of the human A2a adenosine receptor gene. J. Neurochem. 1996, 66, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Schiffmann, S.N.; Jacobs, O.; Vanderhaeghen, J.J. Striatal restricted adenosine A2 receptor (RDC8) is expressed by enkephalin but not by substance P neurons: an in situ hybridization histochemistry study. J. Neurochem. 1991, 57, 1062–1067. [Google Scholar] [CrossRef] [PubMed]
- Schiffmann, S.N.; Fisone, G.; Moresco, R.; Cunha, R.A.; Ferre, S. Adenosine A2A receptors and basal ganglia physiology. Prog. Neurobiol. 2007, 83, 277–292. [Google Scholar] [CrossRef] [Green Version]
- Brooks, D.J.; Doder, M.; Osman, S.; Luthra, S.K.; Hirani, E.; Hume, S.; Kase, H.; Kilborn, J.; Martindill, S.; Mori, A. Positron emission tomography analysis of [11C]KW-6002 binding to human and rat adenosine A2A receptors in the brain. Synapse 2008, 62, 671–681. [Google Scholar] [CrossRef]
- Lee, Y.C.; Chien, C.L.; Sun, C.N.; Huang, C.L.; Huang, N.K.; Chiang, M.C.; Lai, H.L.; Lin, Y.S.; Chou, S.Y.; Wang, C.K.; et al. Characterization of the rat A2A adenosine receptor gene: a 4.8-kb promoter-proximal DNA fragment confers selective expression in the central nervous system. Eur. J. Neurosci. 2003, 18, 1786–1796. [Google Scholar] [CrossRef]
- Nishizaki, T.; Nagai, K.; Nomura, T.; Tada, H.; Kanno, T.; Tozaki, H.; Li, X.X.; Kondoh, T.; Kodama, N.; Takahashi, E.; et al. A new neuromodulatory pathway with a glial contribution mediated via A(2a) adenosine receptors. Glia 2002, 39, 133–147. [Google Scholar] [CrossRef]
- Sebastião, A.M.; Ribeiro, J.A. Tuning and fine-tuning of synapses with adenosine. Curr. Neuropharmacol. 2009, 7, 180–194. [Google Scholar] [CrossRef] [Green Version]
- Quiroz, C.; Luján, R.; Uchigashima, M.; Simoes, A.P.; Lerner, T.N.; Borycz, J.; Kachroo, A.; Canas, P.M.; Orru, M.; Schwarzschild, M.A.; et al. Key modulatory role of presynaptic adenosine A2A receptors in cortical neurotransmission to the striatal direct pathway. ScientificWorldJournal 2009, 9, 1321–1344. [Google Scholar] [CrossRef] [Green Version]
- Daré, E.; Schulte, G.; Karovic, O.; Hammarberg, C.; Fredholm, B.B. Modulation of glial cell functions by adenosine receptors. Physiol. Behav. 2007, 92, 15–20. [Google Scholar] [CrossRef]
- Hettinger, B.D.; Lee, A.; Linden, J.; Rosin, D.L. Ultrastructural localization of adenosine A2A receptors suggests multiple cellular sites for modulation of GABAergic neurons in rat striatum. J. Comp. Neurol. 2001, 431, 331–346. [Google Scholar] [CrossRef]
- Matos, M.; Augusto, E.; Santos-Rodrigues, A.D.; Schwarzschild, M.A.; Chen, J.F.; Cunha, R.A.; Agostinho, P. Adenosine A2A receptors modulate glutamate uptake in cultured astrocytes and gliosomes. Glia 2012, 60, 702–716. [Google Scholar] [CrossRef] [PubMed]
- Matos, M.; Augusto, E.; Agostinho, P.; Cunha, R.A.; Chen, J.F. Antagonistic interaction between adenosine A2A receptors and Na+/K+-ATPase-α2 controlling glutamate uptake in astrocytes. J. Neurosci. 2013, 33, 18492–18502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augood, S.J.; Emson, P.C. Adenosine A2a receptor mRNA is expressed by enkephalin cells but not by somatostatin cells in rat striatum: a co-expression study. Brain Res. Mol. Brain Res. 1994, 22, 204–210. [Google Scholar] [CrossRef]
- Fink, J.S.; Weaver, D.R.; Rivkees, S.A.; Peterfreund, R.A.; Pollack, A.E.; Adler, E.M.; Reppert, S.M. Molecular cloning of the rat A2 adenosine receptor: selective co-expression with D2 dopamine receptors in rat striatum. Brain Res. Mol. Brain Res. 1992, 14, 186–195. [Google Scholar] [CrossRef]
- Rodrigues, R.J.; Alfaro, T.M.; Rebola, N.; Oliveira, C.R.; Cunha, R.A. Co-localization and functional interaction between adenosine A(2A) and metabotropic group 5 receptors in glutamatergic nerve terminals of the rat striatum. J. Neurochem. 2005, 92, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.J.; James, S.; Reddington, M.; Richardson, P.J. Both A1 and A2a purine receptors regulate striatal acetylcholine release. J. Neurochem. 1990, 55, 31–38. [Google Scholar] [CrossRef]
- Kurokawa, M.; Kirk, I.P.; Kirkpatrick, K.A.; Kase, H.; Richardson, P.J. Inhibition by KF17837 of adenosine A2A receptor-mediated modulation of striatal GABA and ACh release. Br. J. Pharmacol. 1994, 113, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Kurokawa, M.; Koga, K.; Kase, H.; Nakamura, J.; Kuwana, Y. Adenosine A2a receptor-mediated modulation of striatal acetylcholine release in vivo. J. Neurochem. 1996, 66, 1882–1888. [Google Scholar] [CrossRef]
- Canals, M.; Marcellino, D.; Fanelli, F.; Ciruela, F.; de Benedetti, P.; Goldberg, S.R.; Neve, K.; Fuxe, K.; Agnati, L.F.; Woods, A.S.; et al. Adenosine A2A-dopamine D2 receptor-receptor heteromerization: qualitative and quantitative assessment by fluorescence and bioluminescence energy transfer. J. Biol. Chem. 2003, 278, 46741–46749. [Google Scholar] [CrossRef] [Green Version]
- Canals, M.; Burgueño, J.; Marcellino, D.; Cabello, N.; Canela, E.I.; Mallol, J.; Agnati, L.; Ferré, S.; Bouvier, M.; Fuxe, K.; et al. Homodimerization of adenosine A2A receptors: qualitative and quantitative assessment by fluorescence and bioluminescence energy transfer. J. Neurochem. 2004, 88, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Brito, I.; Romero-Fernandez, W.; Di Palma, M.; Oflijan, J.; Skieterska, K.; Duchou, J.; Van Craenenbroeck, K.; Suarez-Boomgaard, D.; Rivera, A.; et al. The G protein-coupled receptor heterodimer network (GPCR-HetNet) and its hub components. Int. J. Mol. Sci. 2014, 15, 8570–8590. [Google Scholar] [CrossRef] [PubMed]
- Borroto Escuela, D.; Hagman, B.; Woolfenden, M.; Pinton, L.; Jiménez-Beristain, A.; Oflijan, J.; Narváez Peláez, M.; Palma, M.; Feltmann, K.; Sartini, S.; et al. In Situ Proximity Ligation Assay to Study and Understand the Distribution and Balance of GPCR Homo- and Heteroreceptor Complexes in the Brain. In Receptor and Ion Channel Detection in the Brain; Lujan, R., Ciruela, F., Eds.; Humana Press: New York, NY, USA, 2016; pp. 109–124. [Google Scholar]
- Borroto Escuela, D.; Brito, I.; Palma, M.; Jiménez-Beristain, A.; Narváez Peláez, M.; Corrales, F.; Pita, M.; Sartini, S.; Ambrogini, P.; Lattanzi, D.; et al. On the Role of the Balance of GPCR Homo/ Heteroreceptor Complexes in the Brain. J. Adv. Neurosci. Res. 2015, 2, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Hinz, S.; Navarro, G.; Franco, R.; Müller, C.E.; Fuxe, K. Understanding the Role of Adenosine A2AR Heteroreceptor Complexes in Neurodegeneration and Neuroinflammation. Front. Neurosci. 2018, 12, 43. [Google Scholar] [CrossRef] [Green Version]
- Ciruela, F.; Casadó, V.; Rodrigues, R.J.; Luján, R.; Burgueño, J.; Canals, M.; Borycz, J.; Rebola, N.; Goldberg, S.R.; Mallol, J.; et al. Presynaptic control of striatal glutamatergic neurotransmission by adenosine A1-A2A receptor heteromers. J. Neurosci. 2006, 26, 2080–2087. [Google Scholar] [CrossRef] [Green Version]
- Ferre, S.; Karcz-Kubicha, M.; Hope, B.T.; Popoli, P.; Burgueno, J.; Gutierrez, M.A.; Casado, V.; Fuxe, K.; Goldberg, S.R.; Lluis, C.; et al. Synergistic interaction between adenosine A2A and glutamate mGlu5 receptors: implications for striatal neuronal function. Proc. Natl. Acad. Sci. USA 2002, 99, 11940–11945. [Google Scholar] [CrossRef] [Green Version]
- Ferre, S.; Popoli, P.; Rimondini, R.; Reggio, R.; Kehr, J.; Fuxe, K. Adenosine A2A and group I metabotropic glutamate receptors synergistically modulate the binding characteristics of dopamine D2 receptors in the rat striatum. Neuropharmacology 1999, 38, 129–140. [Google Scholar] [CrossRef]
- Nishi, A.; Liu, F.; Matsuyama, S.; Hamada, M.; Higashi, H.; Nairn, A.C.; Greengard, P. Metabotropic mGlu5 receptors regulate adenosine A2A receptor signaling. Proc. Natl. Acad. Sci. USA 2003, 100, 1322–1327. [Google Scholar] [CrossRef] [Green Version]
- Carriba, P.; Ortiz, O.; Patkar, K.; Justinova, Z.; Stroik, J.; Themann, A.; Muller, C.; Woods, A.S.; Hope, B.T.; Ciruela, F.; et al. Striatal adenosine A2A and cannabinoid CB1 receptors form functional heteromeric complexes that mediate the motor effects of cannabinoids. Neuropsychopharmacology 2007, 32, 2249–2259. [Google Scholar] [CrossRef] [Green Version]
- Ferré, S.; Lluís, C.; Justinova, Z.; Quiroz, C.; Orru, M.; Navarro, G.; Canela, E.I.; Franco, R.; Goldberg, S.R. Adenosine-cannabinoid receptor interactions. Implications for striatal function. Br. J. Pharmacol. 2010, 160, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Cabello, N.; Gandia, J.; Bertarelli, D.C.; Watanabe, M.; Lluis, C.; Franco, R.; Ferre, S.; Lujan, R.; Ciruela, F. Metabotropic glutamate type 5, dopamine D2 and adenosine A2a receptors form higher-order oligomers in living cells. J. Neurochem. 2009, 109, 1497–1507. [Google Scholar] [CrossRef] [PubMed]
- Carriba, P.; Navarro, G.; Ciruela, F.; Ferre, S.; Casado, V.; Agnati, L.; Cortes, A.; Mallol, J.; Fuxe, K.; Canela, E.I.; et al. Detection of heteromerization of more than two proteins by sequential BRET-FRET. Nat. Methods 2008, 5, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Agnati, L.F.; Jacobsen, K.; Hillion, J.; Canals, M.; Torvinen, M.; Tinner-Staines, B.; Staines, W.; Rosin, D.; Terasmaa, A.; et al. Receptor heteromerization in adenosine A2A receptor signaling: relevance for striatal function and Parkinson’s disease. Neurology 2003, 61, S19–S23. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Dahlstrom, A.B.; Jonsson, G.; Marcellino, D.; Guescini, M.; Dam, M.; Manger, P.; Agnati, L. The discovery of central monoamine neurons gave volume transmission to the wired brain. Prog. Neurobiol. 2010, 90, 82–100. [Google Scholar] [CrossRef] [PubMed]
- Beggiato, S.; Antonelli, T.; Tomasini, M.C.; Borelli, A.C.; Agnati, L.F.; Tanganelli, S.; Fuxe, K.; Ferraro, L. Adenosine A2A-D2 receptor-receptor interactions in putative heteromers in the regulation of the striato–pallidal gaba pathway: possible relevance for parkinson’s disease and its treatment. Curr. Protein Pept. Sci. 2014, 15, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Tozzi, A.; Tscherter, A.; Belcastro, V.; Tantucci, M.; Costa, C.; Picconi, B.; Centonze, D.; Calabresi, P.; Borsini, F. Interaction of A2A adenosine and D2 dopamine receptors modulates corticostriatal glutamatergic transmission. Neuropharmacology 2007, 53, 783–789. [Google Scholar] [CrossRef]
- Flajolet, M.; Wang, Z.; Futter, M.; Shen, W.; Nuangchamnong, N.; Bendor, J.; Wallach, I.; Nairn, A.C.; Surmeier, D.J.; Greengard, P. FGF acts as a co-transmitter through adenosine A(2A) receptor to regulate synaptic plasticity. Nat. Neurosci. 2008, 11, 1402–1409. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Ravani, A.; Tarakanov, A.O.; Brito, I.; Narvaez, M.; Romero-Fernandez, W.; Corrales, F.; Agnati, L.F.; Tanganelli, S.; Ferraro, L.; et al. Dopamine D2 receptor signaling dynamics of dopamine D2-neurotensin 1 receptor heteromers. Biochem. Biophys. Res. Commun. 2013, 435, 140–146. [Google Scholar] [CrossRef]
- Fuxe, K.; Borroto-Escuela, D.; Fisone, G.; Agnati, L.F.; Tanganelli, S. Understanding the role of heteroreceptor complexes in the central nervous system. Curr. Protein Pept. Sci. 2014, 15, 647. [Google Scholar] [CrossRef]
- Fuxe, K.; Ferré, S.; Canals, M.; Torvinen, M.; Terasmaa, A.; Marcellino, D.; Goldberg, S.R.; Staines, W.; Jacobsen, K.X.; Lluis, C.; et al. Adenosine A2A and dopamine D2 heteromeric receptor complexes and their function. J. Mol. Neurosci. 2005, 26, 209–220. [Google Scholar] [CrossRef]
- Torvinen, M.; Marcellino, D.; Canals, M.; Agnati, L.F.; Lluis, C.; Franco, R.; Fuxe, K. Adenosine A2A receptor and dopamine D3 receptor interactions: Evidence of functional A2A/D3 heteromeric complexes. Mol. Pharmacol. 2005, 67, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Rivera, A.; Cuellar, B.; Giron, F.J.; Grandy, D.K.; de la Calle, A.; Moratalla, R. Dopamine D4 receptors are heterogeneously distributed in the striosomes/matrix compartments of the striatum. J. Neurochem. 2002, 80, 219–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferre, S.; Quiroz, C.; Woods, A.S.; Cunha, R.; Popoli, P.; Ciruela, F.; Lluis, C.; Franco, R.; Azdad, K.; Schiffmann, S.N. An update on adenosine A2A-dopamine D2 receptor interactions: implications for the function of G protein-coupled receptors. Curr. Pharm. Des. 2008, 14, 1468–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferre, S.; Quiroz, C.; Orru, M.; Guitart, X.; Navarro, G.; Cortes, A.; Casado, V.; Canela, E.I.; Lluis, C.; Franco, R. Adenosine A(2A) Receptors and A(2A) Receptor Heteromers as Key Players in Striatal Function. Front. Neuroanat. 2011, 5, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuxe, K.; Marcellino, D.; Borroto-Escuela, D.O.; Guescini, M.; Fernandez-Duenas, V.; Tanganelli, S.; Rivera, A.; Ciruela, F.; Agnati, L.F. Adenosine-dopamine interactions in the pathophysiology and treatment of CNS disorders. CNS Neurosci. Ther. 2010, 16, e18–e42. [Google Scholar] [CrossRef] [PubMed]
- Tozzi, A.; de Iure, A.; Di Filippo, M.; Tantucci, M.; Costa, C.; Borsini, F.; Ghiglieri, V.; Giampà, C.; Fusco, F.R.; Picconi, B.; et al. The Distinct Role of Medium Spiny Neurons and Cholinergic Interneurons in the D2/A2AReceptor Interaction in the Striatum: Implications for Parkinson’s Disease. J. Neurosci. 2011, 31, 1850. [Google Scholar] [CrossRef] [Green Version]
- Cervetto, C.; Venturini, A.; Passalacqua, M.; Guidolin, D.; Genedani, S.; Fuxe, K.; Borroto-Esquela, D.O.; Cortelli, P.; Woods, A.; Maura, G.; et al. A2A-D2 receptor-receptor interaction modulates gliotransmitter release from striatal astrocyte processes. J. Neurochem. 2017, 140, 268–279. [Google Scholar] [CrossRef]
- Cervetto, C.; Venturini, A.; Guidolin, D.; Maura, G.; Passalacqua, M.; Tacchetti, C.; Cortelli, P.; Genedani, S.; Candiani, S.; Ramoino, P.; et al. Homocysteine and A2A-D2 Receptor-Receptor Interaction at Striatal Astrocyte Processes. J. Mol. Neurosci. 2018, 65, 456–466. [Google Scholar] [CrossRef]
- Trifilieff, P.; Rives, M.L.; Urizar, E.; Piskorowski, R.A.; Vishwasrao, H.D.; Castrillon, J.; Schmauss, C.; Slättman, M.; Gullberg, M.; Javitch, J.A. Detection of antigen interactions ex vivo by proximity ligation assay: endogenous dopamine D2-adenosine A2A receptor complexes in the striatum. Biotechniques 2011, 51, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Garriga, P.; Ciruela, F.; Narvaez, M.; Tarakanov, A.O.; Palkovits, M.; Agnati, L.F.; Fuxe, K. G protein-coupled receptor heterodimerization in the brain. Methods Enzymol. 2013, 521, 281–294. [Google Scholar] [CrossRef] [Green Version]
- Tarakanov, A.O.; Fuxe, K.G. Triplet puzzle: Homologies of receptor heteromers. J. Mol. Neurosci. 2010, 41, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Pinton, L.; O Borroto-Escuela, D.; Narváez, M.; Oflijan, J.; F Agnati, L.; Fuxe, K. Evidence for the existence of dopamine D2R and Sigma 1 allosteric receptor-receptor interaction in the rat brain: Role in brain plasticity and cocaine action. SpringerPlus 2015, 4, P37. [Google Scholar] [CrossRef] [Green Version]
- Turgeon, S.M.; Pollack, A.E.; Schusheim, L.; Fink, J.S. Effects of selective adenosine A1 and A2a agonists on amphetamine-induced locomotion and c-Fos in striatum and nucleus accumbens. Brain Res. 1996, 707, 75–80. [Google Scholar] [CrossRef]
- Rimondini, R.; Ferre, S.; Ogren, S.O.; Fuxe, K. Adenosine A2A agonists: A potential new type of atypical antipsychotic. Neuropsychopharmacology 1997, 17, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Kavanagh, K.A.; Schreiner, D.C.; Levis, S.C.; O’Neill, C.E.; Bachtell, R.K. Role of adenosine receptor subtypes in methamphetamine reward and reinforcement. Neuropharmacology 2015, 89, 265–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poleszak, E.; Malec, D. Cocaine-induced hyperactivity is more influenced by adenosine receptor agonists than amphetamine-induced hyperactivity. Pol. J. Pharmacol. 2002, 54, 359–366. [Google Scholar] [PubMed]
- Filip, M.; Frankowska, M.; Zaniewska, M.; Przegalinski, E.; Muller, C.E.; Agnati, L.; Franco, R.; Roberts, D.C.; Fuxe, K. Involvement of adenosine A2A and dopamine receptors in the locomotor and sensitizing effects of cocaine. Brain Res. 2006, 1077, 67–80. [Google Scholar] [CrossRef]
- Czyzyk, J.; Zaniewska, M.; Bader, M.; Fuxe, K.; Filip, M. Attenuated behavioral responses to acute and chronic cocaine in adenosine A2A overexpressed transgenic rats. In European Neuropsychopharmacology. Proceedings of the The 24th ECNP Congress, Paris, France, 3–7 September 2011; ECNP: Utrecht, The Netherlands; pp. S597–S598.
- Poleszak, E.; Malec, D. Adenosine receptor ligands and cocaine in conditioned place preference (CPP) test in rats. Pol. J. Pharmacol. 2002, 54, 119–126. [Google Scholar]
- Haynes, N.S.; O’Neill, C.E.; Hobson, B.D.; Bachtell, R.K. Effects of adenosine A2A receptor antagonists on cocaine-induced locomotion and cocaine seeking. Psychopharmacology (Berl). 2019, 236, 699–708. [Google Scholar] [CrossRef]
- Chen, J.F.; Beilstein, M.; Xu, Y.H.; Turner, T.J.; Moratalla, R.; Standaert, D.G.; Aloyo, V.J.; Fink, J.S.; Schwarzschild, M.A. Selective attenuation of psychostimulant-induced behavioral responses in mice lacking A(2A) adenosine receptors. Neuroscience 2000, 97, 195–204. [Google Scholar] [CrossRef]
- Wright, S.R.; Zanos, P.; Georgiou, P.; Yoo, J.H.; Ledent, C.; Hourani, S.M.; Kitchen, I.; Winsky-Sommerer, R.; Bailey, A. A critical role of striatal A2A R-mGlu5 R interactions in modulating the psychomotor and drug-seeking effects of methamphetamine. Addict. Biol. 2016, 21, 811–825. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.Y.; Coelho, J.E.; Ohtsuka, N.; Canas, P.M.; Day, Y.J.; Huang, Q.Y.; Rebola, N.; Yu, L.; Boison, D.; Cunha, R.A.; et al. A critical role of the adenosine A2A receptor in extrastriatal neurons in modulating psychomotor activity as revealed by opposite phenotypes of striatum and forebrain A2A receptor knock-outs. J. Neurosci. 2008, 28, 2970–2975. [Google Scholar] [CrossRef]
- Shimazoe, T.; Yoshimatsu, A.; Kawashimo, A.; Watanabe, S. Roles of adenosine A(1) and A(2A) receptors in the expression and development of methamphetamine-induced sensitization. Eur. J. Pharmacol. 2000, 388, 249–254. [Google Scholar] [CrossRef]
- Bastia, E.; Xu, Y.H.; Scibelli, A.C.; Day, Y.J.; Linden, J.; Chen, J.F.; Schwarzschild, M.A. A crucial role for forebrain adenosine A(2A) receptors in amphetamine sensitization. Neuropsychopharmacology 2005, 30, 891–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.F.; Moratalla, R.; Yu, L.; Martin, A.B.; Xu, K.; Bastia, E.; Hackett, E.; Alberti, I.; Schwarzschild, M.A. Inactivation of adenosine A2A receptors selectively attenuates amphetamine-induced behavioral sensitization. Neuropsychopharmacology 2003, 28, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Soria, G.; Castane, A.; Ledent, C.; Parmentier, M.; Maldonado, R.; Valverde, O. The lack of A2A adenosine receptors diminishes the reinforcing efficacy of cocaine. Neuropsychopharmacology 2006, 31, 978–987. [Google Scholar] [CrossRef]
- Listos, J.; Talarek, S.; Fidecka, S. Involvement of adenosine receptor agonists on the development of hypersensitivity to acute dose of morphine during morphine withdrawal period. Pharmacol. Rep. 2008, 60, 679–685. [Google Scholar]
- Listos, J.; Talarek, S.; Poleszak, E.; Wrobel, A.; Fidecka, S. Attenuating effect of adenosine receptor agonists on the development of behavioral sensitization induced by sporadic treatment with morphine. Pharmacol. Biochem. Behav. 2011, 98, 356–361. [Google Scholar] [CrossRef]
- Castane, A.; Wells, L.; Soria, G.; Hourani, S.; Ledent, C.; Kitchen, I.; Opacka-Juffry, J.; Maldonado, R.; Valverde, O. Behavioural and biochemical responses to morphine associated with its motivational properties are altered in adenosine A(2A) receptor knockout mice. Br. J. Pharmacol. 2008, 155, 757–766. [Google Scholar] [CrossRef] [Green Version]
- Soria, G.; Castane, A.; Berrendero, F.; Ledent, C.; Parmentier, M.; Maldonado, R.; Valverde, O. Adenosine A2A receptors are involved in physical dependence and place conditioning induced by THC. Eur. J. Neurosci. 2004, 20, 2203–2213. [Google Scholar] [CrossRef]
- Fritz, B.M.; Boehm, S.L. 2nd. Adenosinergic regulation of binge-like ethanol drinking and associated locomotor effects in male C57BL/6J mice. Pharmacol. Biochem. Behav. 2015, 135, 83–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houchi, H.; Warnault, V.; Barbier, E.; Dubois, C.; Pierrefiche, O.; Ledent, C.; Daoust, M.; Naassila, M. Involvement of A2A receptors in anxiolytic, locomotor and motivational properties of ethanol in mice. Genes Brain Behav. 2008, 7, 887–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jastrzębska, J.; Nowak, E.; Smaga, I.; Bystrowska, B.; Frankowska, M.; Bader, M.; Filip, M.; Fuxe, K. Adenosine (A)(2A)receptor modulation of nicotine-induced locomotor sensitization. A pharmacological and transgenic approach. Neuropharmacology 2014, 81, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Castane, A.; Soria, G.; Ledent, C.; Maldonado, R.; Valverde, O. Attenuation of nicotine-induced rewarding effects in A2A knockout mice. Neuropharmacology 2006, 51, 631–640. [Google Scholar] [CrossRef]
- Justinova, Z.; Ferre, S.; Segal, P.N.; Antoniou, K.; Solinas, M.; Pappas, L.A.; Highkin, J.L.; Hockemeyer, J.; Munzar, P.; Goldberg, S.R. Involvement of adenosine A1 and A2A receptors in the adenosinergic modulation of the discriminative-stimulus effects of cocaine and methamphetamine in rats. J. Pharmacol. Exp. Ther. 2003, 307, 977–986. [Google Scholar] [CrossRef] [Green Version]
- Justinova, Z.; Ferre, S.; Barnes, C.; Wertheim, C.E.; Pappas, L.A.; Goldberg, S.R.; Le Foll, B. Effects of chronic caffeine exposure on adenosinergic modulation of the discriminative-stimulus effects of nicotine, methamphetamine, and cocaine in rats. Psychopharmacology (Berl). 2009, 203, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Munzar, P.; Justinova, Z.; Kutkat, S.W.; Ferre, S.; Goldberg, S.R. Adenosinergic modulation of the discriminative-stimulus effects of methamphetamine in rats. Psychopharmacol. Berl. 2002, 161, 348–355. [Google Scholar] [CrossRef]
- Knapp, C.M.; Foye, M.M.; Cottam, N.; Ciraulo, D.A.; Kornetsky, C. Adenosine agonists CGS 21680 and NECA inhibit the initiation of cocaine self-administration. Pharmacol. Biochem. Behav. 2001, 68, 797–803. [Google Scholar] [CrossRef]
- Wydra, K.; Golembiowska, K.; Suder, A.; Kaminska, K.; Fuxe, K.; Filip, M. On the role of adenosine (A)(2)A receptors in cocaine-induced reward: A pharmacological and neurochemical analysis in rats. Psychopharmacology (Berl). 2015, 232, 421–435. [Google Scholar] [CrossRef]
- Wydra, K.; Suder, A.; Frankowska, M.; Borroto Escuela, D.O.; Fuxe, K.; Filip, M. Effects of intra-accumbal or intra-prefrontal cortex microinjections of adenosine 2A receptor ligands on responses to cocaine reward and seeking in rats. Psychopharmacology (Berl). 2018, 235, 3509–3523. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Wydra, K.; Romero-Fernandez, W.; Zhou, Z.; Frankowska, M.; Filip, M.; Fuxe, K. A2AR Transmembrane 2 Peptide Administration Disrupts the A2AR-A2AR Homoreceptor but Not the A2AR-D2R Heteroreceptor Complex: Lack of Actions on Rodent Cocaine Self-Administration. Int. J. Mol. Sci. 2019, 20, 6100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borroto-Escuela, D.O.; Wydra, K.; Li, X.; Rodriguez, D.; Carlsson, J.; Jastrzebska, J.; Filip, M.; Fuxe, K. Disruption of A2AR-D2R Heteroreceptor Complexes After A2AR Transmembrane 5 Peptide Administration Enhances Cocaine Self-Administration in Rats. Mol. Neurobiol. 2018, 55, 7038–7048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poleszak, E.; Malec, D. Effects of adenosine receptor agonists and antagonists in amphetamine-induced conditioned place preference test in rats. Pol. J. Pharmacol. 2003, 55, 319–326. [Google Scholar] [PubMed]
- Justinova, Z.; Ferre, S.; Redhi, G.H.; Mascia, P.; Stroik, J.; Quarta, D.; Yasar, S.; Muller, C.E.; Franco, R.; Goldberg, S.R. Reinforcing and neurochemical effects of cannabinoid CB1 receptor agonists, but not cocaine, are altered by an adenosine A2A receptor antagonist. Addict. Biol. 2011, 16, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Medina, J.; Ledent, C.; Carreton, O.; Valverde, O. The A2a adenosine receptor modulates the reinforcement efficacy and neurotoxicity of MDMA. J. Psychopharmacol. 2011, 25, 550–564. [Google Scholar] [CrossRef]
- Gill, W.D.; Shelton, H.W.; Burgess, K.C.; Brown, R.W. Effects of an adenosine A2A agonist on the rewarding associative properties of nicotine and neural plasticity in a rodent model of schizophrenia. J. Psychopharmacol. 2020, 34, 137–144. [Google Scholar] [CrossRef]
- Sahraei, H.; Motamedi, F.; Khoshbaten, A.; Zarrindast, M.R. Adenosine A(2) receptors inhibit morphine self-administration in rats. Eur. J. Pharmacol. 1999, 383, 107–113. [Google Scholar] [CrossRef]
- Listos, J.; Talarek, S.; Listos, P.; Orzelska, J.; Lupina, M.; Fidecka, S. Effects of the adenosinergic system on the expression and acquisition of sensitization to conditioned place preference in morphine-conditioned rats. Naunyn Schmiedebergs Arch. Pharmacol. 2016, 389, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Brown, R.M.; Short, J.L.; Cowen, M.S.; Ledent, C.; Lawrence, A.J. A differential role for the adenosine A2A receptor in opiate reinforcement vs opiate-seeking behavior. Neuropsychopharmacology 2009, 34, 844–856. [Google Scholar] [CrossRef] [Green Version]
- Houchi, H.; Persyn, W.; Legastelois, R.; Naassila, M. The adenosine A2A receptor agonist CGS 21680 decreases ethanol self-administration in both non-dependent and dependent animals. Addict. Biol. 2013, 18, 812–825. [Google Scholar] [CrossRef]
- Zaniewska, M.; Gawlinski, D.; Wyczesana, M.; Nowak, E.; Kula, K.; Maciow-Glab, M.; Jastrzebska, J.; Sadakierska-Chudy, A.; Bader, M.; Fuxe, K. Increased Ethanol Consumption and Locomotion Develop upon Ethanol Deprivation in Rats Overexpressing the Adenosine (A)2A Receptor. Neuroscience 2019, 418, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Arolfo, M.P.; Yao, L.; Gordon, A.S.; Diamond, I.; Janak, P.H. Ethanol operant self-administration in rats is regulated by adenosine A2 receptors. Alcohol. Clin. Exp. Res. 2004, 28, 1308–1316. [Google Scholar] [CrossRef] [PubMed]
- Thorsell, A.; Johnson, J.; Heilig, M. Effect of the adenosine A2a receptor antagonist 3,7-dimethyl-propargylxanthine on anxiety-like and depression-like behavior and alcohol consumption in Wistar Rats. Alcohol. Clin. Exp. Res. 2007, 31, 1302–1307. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.L.; Cowen, M.S.; Short, J.L.; Lawrence, A.J. Combined antagonism of glutamate mGlu5 and adenosine A2A receptors interact to regulate alcohol-seeking in rats. Int. J. Neuropsychopharmacol. 2008, 11, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Nam, H.W.; Hinton, D.J.; Kang, N.Y.; Kim, T.; Lee, M.R.; Oliveros, A.; Adams, C.; Ruby, C.L.; Choi, D.S. Adenosine transporter ENT1 regulates the acquisition of goal-directed behavior and ethanol drinking through A2A receptor in the dorsomedial striatum. J. Neurosci. 2013, 33, 4329–4338. [Google Scholar] [CrossRef] [Green Version]
- Naassila, M.; Ledent, C.; Daoust, M. Low ethanol sensitivity and increased ethanol consumption in mice lacking adenosine A2A receptors. J. Neurosci. 2002, 22, 10487–10493. [Google Scholar] [CrossRef] [Green Version]
- Mukhara, D.; Banks, M.L.; Neigh, G.N. Stress as a Risk Factor for Substance Use Disorders: A Mini-Review of Molecular Mediators. Front. Behav. Neurosci. 2018, 12, 309. [Google Scholar] [CrossRef] [Green Version]
- Bachtell, R.K.; Self, D.W. Effects of adenosine A2A receptor stimulation on cocaine-seeking behavior in rats. Psychopharmacol. Berl. 2009, 206, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Wydra, K.; Suder, A.; Borroto-Escuela, D.O.; Filip, M.; Fuxe, K. On the role of A(2)A and D(2) receptors in control of cocaine and food-seeking behaviors in rats. Psychopharmacol. Berl. 2015, 232, 1767–1778. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, C.E.; LeTendre, M.L.; Bachtell, R.K. Adenosine A2A receptors in the nucleus accumbens bi-directionally alter cocaine seeking in rats. Neuropsychopharmacology 2012, 37, 1245–1256. [Google Scholar] [CrossRef] [Green Version]
- Larson, T.A.; Winkler, M.C.; Stafford, J.; Levis, S.C.; O’Neill, C.E.; Bachtell, R.K. Role of dopamine D2-like receptors and their modulation by adenosine receptor stimulation in the reinstatement of methamphetamine seeking. Psychopharmacol. Berl. 2019, 236, 1207–1218. [Google Scholar] [CrossRef] [PubMed]
- Weerts, E.M.; Griffiths, R.R. The adenosine receptor antagonist CGS15943 reinstates cocaine-seeking behavior and maintains self-administration in baboons. Psychopharmacol. Berl. 2003, 168, 155–163. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, C.E.; Hobson, B.D.; Levis, S.C.; Bachtell, R.K. Persistent reduction of cocaine seeking by pharmacological manipulation of adenosine A1 and A 2A receptors during extinction training in rats. Psychopharmacol. Berl. 2014, 231, 3179–3188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, L.; McFarland, K.; Fan, P.; Jiang, Z.; Ueda, T.; Diamond, I. Adenosine A2a blockade prevents synergy between mu-opiate and cannabinoid CB1 receptors and eliminates heroin-seeking behavior in addicted rats. Proc. Natl. Acad. Sci. USA 2006, 103, 7877–7882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldo, B.A.; Koob, G.F.; Markou, A. Role of adenosine A2 receptors in brain stimulation reward under baseline conditions and during cocaine withdrawal in rats. J. Neurosci. 1999, 19, 11017–11026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dionyssopoulos, T.; Hope, W.; Coupar, I.M. Effect of adenosine analogues on the expression of opiate withdrawal in rats. Pharmacol. Biochem. Behav. 1992, 42, 201–206. [Google Scholar] [CrossRef]
- Kaplan, G.B.; Sears, M.T. Adenosine receptor agonists attenuate and adenosine receptor antagonists exacerbate opiate withdrawal signs. Psychopharmacol. Berl. 1996, 123, 64–70. [Google Scholar] [CrossRef]
- Salem, A.; Hope, W. Effect of adenosine receptor agonists and antagonists on the expression of opiate withdrawal in rats. Pharmacol. Biochem. Behav. 1997, 57, 671–679. [Google Scholar] [CrossRef]
- Bailey, A.; Davis, L.; Lesscher, H.M.; Kelly, M.D.; Ledent, C.; Hourani, S.M.; Kitchen, I. Enhanced morphine withdrawal and micro -opioid receptor G-protein coupling in A2A adenosine receptor knockout mice. J. Neurochem. 2004, 88, 827–834. [Google Scholar] [CrossRef]
- Bilbao, A.; Cippitelli, A.; Martin, A.B.; Granado, N.; Ortiz, O.; Bezard, E.; Chen, J.F.; Navarro, M.; Rodriguez de Fonseca, F.; Moratalla, R. Absence of quasi-morphine withdrawal syndrome in adenosine A2A receptor knockout mice. Psychopharmacol. Berl. 2006, 185, 160–168. [Google Scholar] [CrossRef]
- Berrendero, F.; Castane, A.; Ledent, C.; Parmentier, M.; Maldonado, R.; Valverde, O. Increase of morphine withdrawal in mice lacking A2a receptors and no changes in CB1/A2a double knockout mice. Eur. J. Neurosci. 2003, 17, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G.B.; Bharmal, N.H.; Leite-Morris, K.A.; Adams, W.R. Role of adenosine A1 and A2A receptors in the alcohol withdrawal syndrome. Alcohol 1999, 19, 157–162. [Google Scholar] [CrossRef]
- Prediger, R.D.; da Silva, G.E.; Batista, L.C.; Bittencourt, A.L.; Takahashi, R.N. Activation of adenosine A1 receptors reduces anxiety-like behavior during acute ethanol withdrawal (hangover) in mice. Neuropsychopharmacology 2006, 31, 2210–2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Yacoubi, M.; Ledent, C.; Parmentier, M.; Daoust, M.; Costentin, J.; Vaugeois, J. Absence of the adenosine A(2A) receptor or its chronic blockade decrease ethanol withdrawal-induced seizures in mice. Neuropharmacology 2001, 40, 424–432. [Google Scholar] [CrossRef]
- Bailey, A.; Gianotti, R.; Ho, A.; Kreek, M.J. Persistent upregulation of mu-opioid, but not adenosine, receptors in brains of long-term withdrawn escalating dose "binge" cocaine-treated rats. Synapse 2005, 57, 160–166. [Google Scholar] [CrossRef]
- Marcellino, D.; Roberts, D.C.; Navarro, G.; Filip, M.; Agnati, L.; Lluis, C.; Franco, R.; Fuxe, K. Increase in A2A receptors in the nucleus accumbens after extended cocaine self-administration and its disappearance after cocaine withdrawal. Brain Res. 2007, 1143, 208–220. [Google Scholar] [CrossRef]
- Frankowska, M.; Marcellino, D.; Adamczyk, P.; Filip, M.; Fuxe, K. Effects of cocaine self-administration and extinction on D2 -like and A2A receptor recognition and D2 -like/Gi protein coupling in rat striatum. Addict. Biol. 2013, 18, 455–466. [Google Scholar] [CrossRef]
- Yang, S.L.; Han, J.Y.; Kim, Y.B.; Nam, S.Y.; Song, S.; Hong, J.T.; Oh, K.W. Increased non-rapid eye movement sleep by cocaine withdrawal: possible involvement of A2A receptors. Arch. Pharm. Res. 2011, 34, 281–287. [Google Scholar] [CrossRef]
- Aso, E.; Fernández-Dueñas, V.; López-Cano, M.; Taura, J.; Watanabe, M.; Ferrer, I.; Luján, R.; Ciruela, F. Adenosine A2A-Cannabinoid CB1 Receptor Heteromers in the Hippocampus: Cannabidiol Blunts Δ9-Tetrahydrocannabinol-Induced Cognitive Impairment. Mol. Neurobiol. 2019, 56, 5382–5391. [Google Scholar] [CrossRef] [Green Version]
- Franco, R.; Reyes-Resina, I.; Aguinaga, D.; Lillo, A.; Jiménez, J.; Raïch, I.; Borroto-Escuela, D.O.; Ferreiro-Vera, C.; Canela, E.I.; Sánchez de Medina, V.; et al. Potentiation of cannabinoid signaling in microglia by adenosine A(2A) receptor antagonists. Glia 2019, 67, 2410–2423. [Google Scholar] [CrossRef]
- Kaplan, G.B.; Leite-Morris, K.A.; Sears, M.T. Alterations of adenosine A1 receptors in morphine dependence. Brain Res. 1994, 657, 347–350. [Google Scholar] [CrossRef]
- De Montis, M.; Devoto, P.; Meloni, D.; Saba, P.; Tagliamonte, A. Deacresed adenosine A2A receptor function in morphine dependent rats. Pharmacol. Res. 1992, 25, 232–233. [Google Scholar] [CrossRef]
- Dar, M.S.; Mustafa, S.J.; Wooles, W.R. Possible role of adenosine in the CNS effects of ethanol. Life Sci. 1983, 33, 1363–1374. [Google Scholar] [CrossRef]
- Clark, M.; Dar, M.S. In vitro autoradiographic evidence for adenosine modulation of ethanol-induced motor disturbances in rats. Alcohol Alcohol. Suppl. 1991, 1, 203–206. [Google Scholar]
- Daly, J.W.; Shi, D.; Wong, V.; Nikodijevic, O. Chronic effects of ethanol on central adenosine function of mice. Brain Res. 1994, 650, 153–156. [Google Scholar] [CrossRef]
- Jarvis, M.F.; Becker, H.C. Single and repeated episodes of ethanol withdrawal increase adenosine A1, but not A2A, receptor density in mouse brain. Brain Res. 1998, 786, 80–88. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Wallman-Johansson, A. Effects of ethanol and acetate on adenosine production in rat hippocampal slices. Pharmacol. Toxicol. 1996, 79, 120–123. [Google Scholar] [CrossRef]
- Bolewska, P.; Martin, B.I.; Orlando, K.A.; Rhoads, D.E. Sequential Changes in Brain Glutamate and Adenosine A1 Receptors May Explain Severity of Adolescent Alcohol Withdrawal after Consumption of High Levels of Alcohol. Neurosci. J. 2019, 2019, 5950818. [Google Scholar] [CrossRef] [Green Version]
- Feltmann, K.; Borroto-Escuela, D.O.; Rüegg, J.; Pinton, L.; de Oliveira Sergio, T.; Narváez, M.; Jimenez-Beristain, A.; Ekström, T.J.; Fuxe, K.; Steensland, P. Effects of Long-Term Alcohol Drinking on the Dopamine D2 Receptor: Gene Expression and Heteroreceptor Complexes in the Striatum in Rats. Alcohol. Clin. Exp. Res. 2018, 42, 338–351. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, C.E.; Levis, S.C.; Schreiner, D.C.; Amat, J.; Maier, S.F.; Bachtell, R.K. Effects of adolescent caffeine consumption on cocaine sensitivity. Neuropsychopharmacology 2015, 40, 813–821. [Google Scholar] [CrossRef] [Green Version]
- Fuxe, K.; Ferre, S.; Zoli, M.; Agnati, L.F. Integrated events in central dopamine transmission as analyzed at multiple levels. Evidence for intramembrane adenosine A2A/dopamine D2 and adenosine A1/dopamine D1 receptor interactions in the basal ganglia. Brain Res. Brain Res. Rev. 1998, 26, 258–273. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Marcellino, D.; Narvaez, M.; Flajolet, M.; Heintz, N.; Agnati, L.; Ciruela, F.; Fuxe, K. A serine point mutation in the adenosine A2AR C-terminal tail reduces receptor heteromerization and allosteric modulation of the dopamine D2R. Biochem. Biophys. Res. Commun. 2010, 394, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Narvaez, M.; Romero-Fernandez, W.; Pinton, L.; Wydra, K.; Filip, M.; Beggiato, S.; Tanganelli, S.; Ferraro, L.; Fuxe, K. Acute Cocaine Enhances Dopamine D2R Recognition and Signaling and Counteracts D2R Internalization in Sigma1R-D2R Heteroreceptor Complexes. Mol. Neurobiol. 2019, 56, 7045–7055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferraro, L.; Beggiato, S.; Marcellino, D.; Frankowska, M.; Filip, M.; Agnati, L.F.; Antonelli, T.; Tomasini, M.C.; Tanganelli, S.; Fuxe, K. Nanomolar concentrations of cocaine enhance D2-like agonist-induced inhibition of the K+-evoked [3H]-dopamine efflux from rat striatal synaptosomes: a novel action of cocaine. J. Neural Transm. (Vienna) 2010, 117, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, L.; Frankowska, M.; Marcellino, D.; Zaniewska, M.; Beggiato, S.; Filip, M.; Tomasini, M.C.; Antonelli, T.; Tanganelli, S.; Fuxe, K. A novel mechanism of cocaine to enhance dopamine d2-like receptor mediated neurochemical and behavioral effects. An in vivo and in vitro study. Neuropsychopharmacology 2012, 37, 1856–1866. [Google Scholar] [CrossRef] [Green Version]
- Romero-Fernandez, W.; Zhou, Z.; Beggiato, S.; Wydra, K.; Filip, M.; Tanganelli, S.; Borroto-Escuela, D.O.; Ferraro, L.; Fuxe, K. Acute cocaine treatment enhances the antagonistic allosteric adenosine A2A-dopamine D2 receptor-receptor interactions in rat dorsal striatum without increasing significantly extracellular dopamine levels. Pharmacol. Rep. 2020, 72, 332–339. [Google Scholar] [CrossRef] [Green Version]
- Wydra, K.; Golembiowska, K.; Zaniewska, M.; Kaminska, K.; Ferraro, L.; Fuxe, K.; Filip, M. Accumbal and pallidal dopamine, glutamate and GABA overflow during cocaine self-administration and its extinction in rats. Addict. Biol. 2013, 18, 307–324. [Google Scholar] [CrossRef]
- Pintsuk, J.; Borroto-Escuela, D.O.; Pomierny, B.; Wydra, K.; Zaniewska, M.; Filip, M.; Fuxe, K. Cocaine self-administration differentially affects allosteric A2A-D2 receptor-receptor interactions in the striatum. Relevance for cocaine use disorder. Pharmacol. Biochem. Behav. 2016, 144, 85–91. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Wydra, K.; Pintsuk, J.; Narvaez, M.; Corrales, F.; Zaniewska, M.; Agnati, L.F.; Franco, R.; Tanganelli, S.; Ferraro, L.; et al. Understanding the Functional Plasticity in Neural Networks of the Basal Ganglia in Cocaine Use Disorder: A Role for Allosteric Receptor-Receptor Interactions in A2A-D2 Heteroreceptor Complexes. Neural Plast. 2016, 2016, 4827268. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Narvaez, M.; Wydra, K.; Pintsuk, J.; Pinton, L.; Jimenez-Beristain, A.; Di Palma, M.; Jastrzebska, J.; Filip, M.; Fuxe, K. Cocaine self-administration specifically increases A2AR-D2R and D2R-sigma1R heteroreceptor complexes in the rat nucleus accumbens shell. Relevance for cocaine use disorder. Pharmacol. Biochem. Behav. 2017, 155, 24–31. [Google Scholar] [CrossRef]
- Romieu, P.; Meunier, J.; Garcia, D.; Zozime, N.; Martin-Fardon, R.; Bowen, W.D.; Maurice, T. The sigma1 (sigma1) receptor activation is a key step for the reactivation of cocaine conditioned place preference by drug priming. Psychopharmacol. Berl. 2004, 175, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Kourrich, S.; Su, T.P.; Fujimoto, M.; Bonci, A. The sigma-1 receptor: roles in neuronal plasticity and disease. Trends Neurosci. 2012, 35, 762–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, G.; Moreno, E.; Bonaventura, J.; Brugarolas, M.; Farre, D.; Aguinaga, D.; Mallol, J.; Cortes, A.; Casado, V.; Lluis, C.; et al. Cocaine inhibits dopamine D2 receptor signaling via sigma-1-D2 receptor heteromers. PLoS ONE 2013, 8, e61245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahlholm, K.; Arhem, P.; Fuxe, K.; Marcellino, D. The dopamine stabilizers ACR16 and (-)-OSU6162 display nanomolar affinities at the sigma-1 receptor. Mol. Psychiatry 2013, 18, 12–14. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Wydra, K.; Zhou, Z.; Suder, A.; Filip, M.; Fuxe, K. OSU-6162, a Sigma1R Ligand in Low Doses, Can Further Increase the Effects of Cocaine Self-Administration on Accumbal D2R Heteroreceptor Complexes. Neurotox. Res. 2020, 37, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Wydra, K.; Filip, M.; Fuxe, K. A2AR-D2R Heteroreceptor Complexes in Cocaine Reward and Addiction. Trends Pharmacol. Sci. 2018, 39, 1008–1020. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Rodriguez, D.; Romero-Fernandez, W.; Kapla, J.; Jaiteh, M.; Ranganathan, A.; Lazarova, T.; Fuxe, K.; Carlsson, J. Mapping the Interface of a GPCR Dimer: A Structural Model of the A2A Adenosine and D2 Dopamine Receptor Heteromer. Front. Pharmacol. 2018, 9, 829. [Google Scholar] [CrossRef] [Green Version]
- Gimenez-Llort, L.; Schiffmann, S.N.; Shmidt, T.; Canela, L.; Camon, L.; Wassholm, M.; Canals, M.; Terasmaa, A.; Fernandez-Teruel, A.; Tobena, A.; et al. Working memory deficits in transgenic rats overexpressing human adenosine A2A receptors in the brain. Neurobiol. Learn Mem. 2007, 87, 42–56. [Google Scholar] [CrossRef] [Green Version]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wydra, K.; Gawliński, D.; Gawlińska, K.; Frankowska, M.; Borroto-Escuela, D.O.; Fuxe, K.; Filip, M. Adenosine A2AReceptors in Substance Use Disorders: A Focus on Cocaine. Cells 2020, 9, 1372. https://doi.org/10.3390/cells9061372
Wydra K, Gawliński D, Gawlińska K, Frankowska M, Borroto-Escuela DO, Fuxe K, Filip M. Adenosine A2AReceptors in Substance Use Disorders: A Focus on Cocaine. Cells. 2020; 9(6):1372. https://doi.org/10.3390/cells9061372
Chicago/Turabian StyleWydra, Karolina, Dawid Gawliński, Kinga Gawlińska, Małgorzata Frankowska, Dasiel O. Borroto-Escuela, Kjell Fuxe, and Małgorzata Filip. 2020. "Adenosine A2AReceptors in Substance Use Disorders: A Focus on Cocaine" Cells 9, no. 6: 1372. https://doi.org/10.3390/cells9061372
APA StyleWydra, K., Gawliński, D., Gawlińska, K., Frankowska, M., Borroto-Escuela, D. O., Fuxe, K., & Filip, M. (2020). Adenosine A2AReceptors in Substance Use Disorders: A Focus on Cocaine. Cells, 9(6), 1372. https://doi.org/10.3390/cells9061372