Hepatogenic Potential and Liver Regeneration Effect of Human Liver-derived Mesenchymal-Like Stem Cells

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of hLD-SCs from Human Liver Tissues
2.2. Preparation of Human Umbilical Cord Matrix-Derived Stem Cells (hUC-MSC) and Human Bone Marrow Stem Cells (hBM-MSC)
2.3. Flow Cytometric Analysis of Human Stem Cells
2.4. Total mRNA Extraction and qRT-PCR Analysis
2.5. Mixed Lymphocyte Reaction (MLR) Assay
2.6. Immunofluorescence Assay
2.7. In Vitro Hepatic Differentiation of Stem Cells
2.8. Mitochondrial Function Measurement (Seahorse Assay)
2.9. Liver Injury Models with In Vivo Stem Cell Treatments
2.10. Cell Labeling with DiR and In Vivo Fluorescence Imaging
2.11. Serum Measurement Assays
2.12. Detection of Human DNA in Mouse Livers
2.13. Histological Analysis and Immunohistochemistry
2.14. Statistical Analysis
3. Results
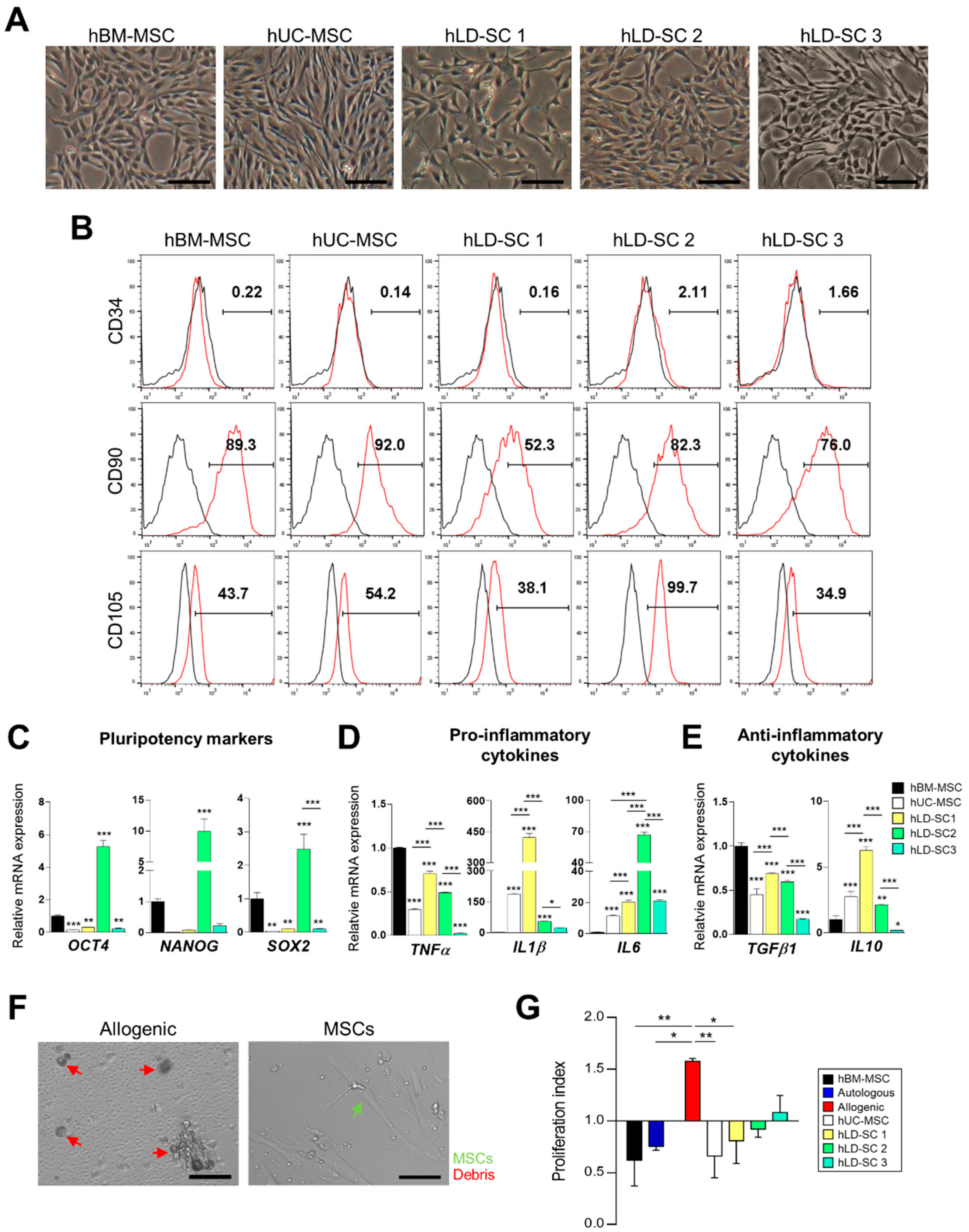
3.1. Characterization of hLD-SCs
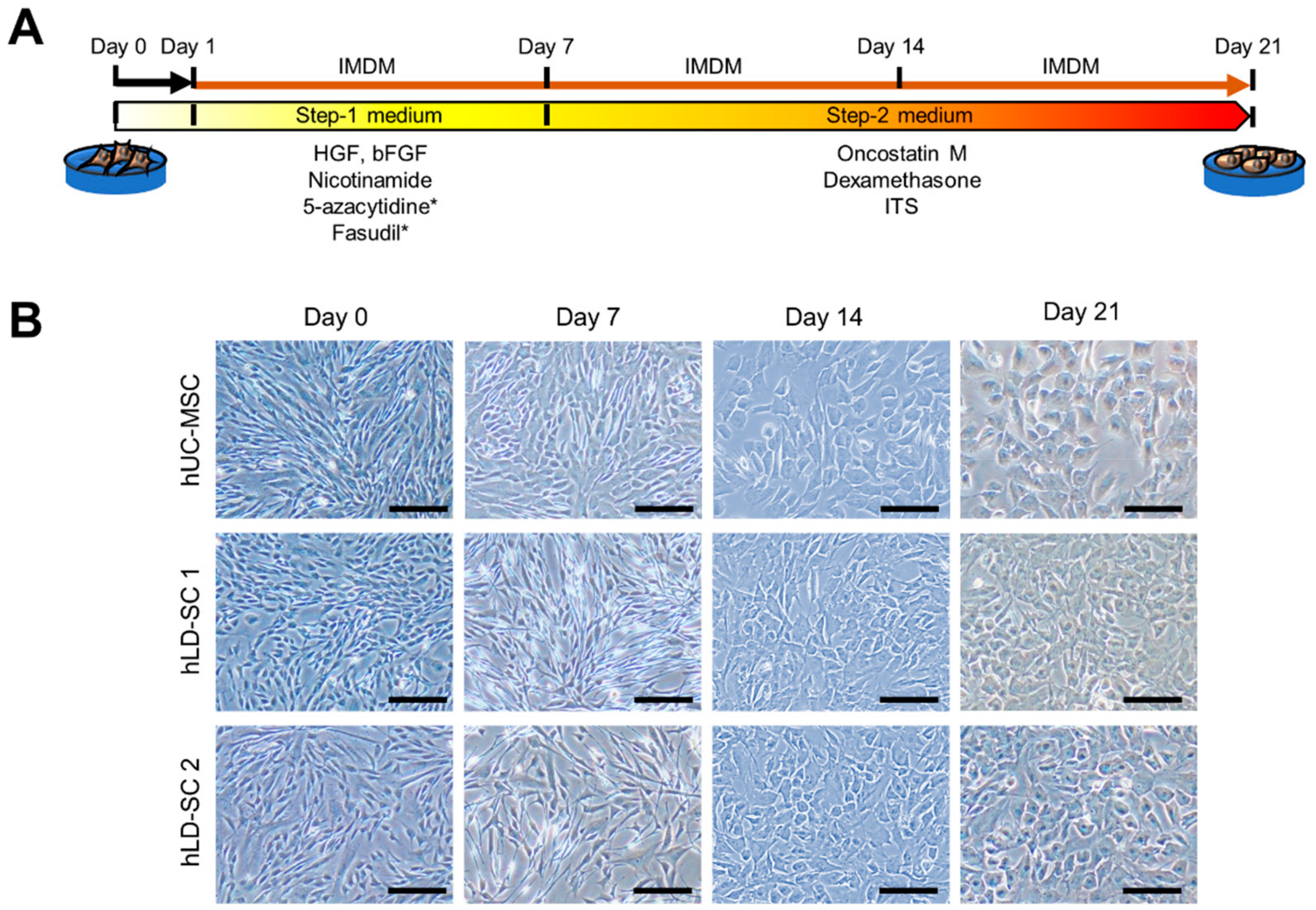
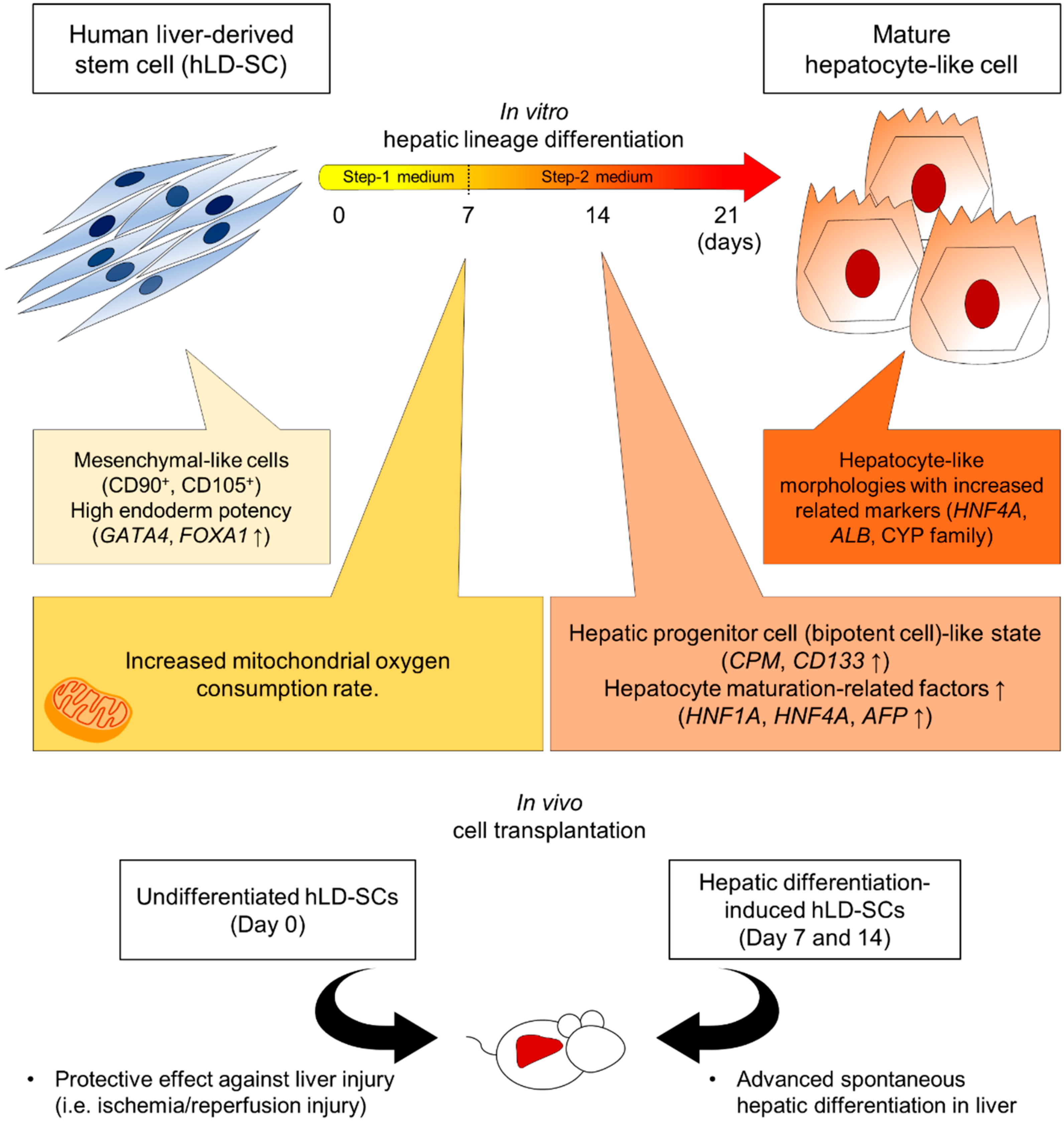
3.2. The Modified Two-Step Hepatic Differentiation Protocol Produced Hepatocyte-Like Cells from hLD-SCs
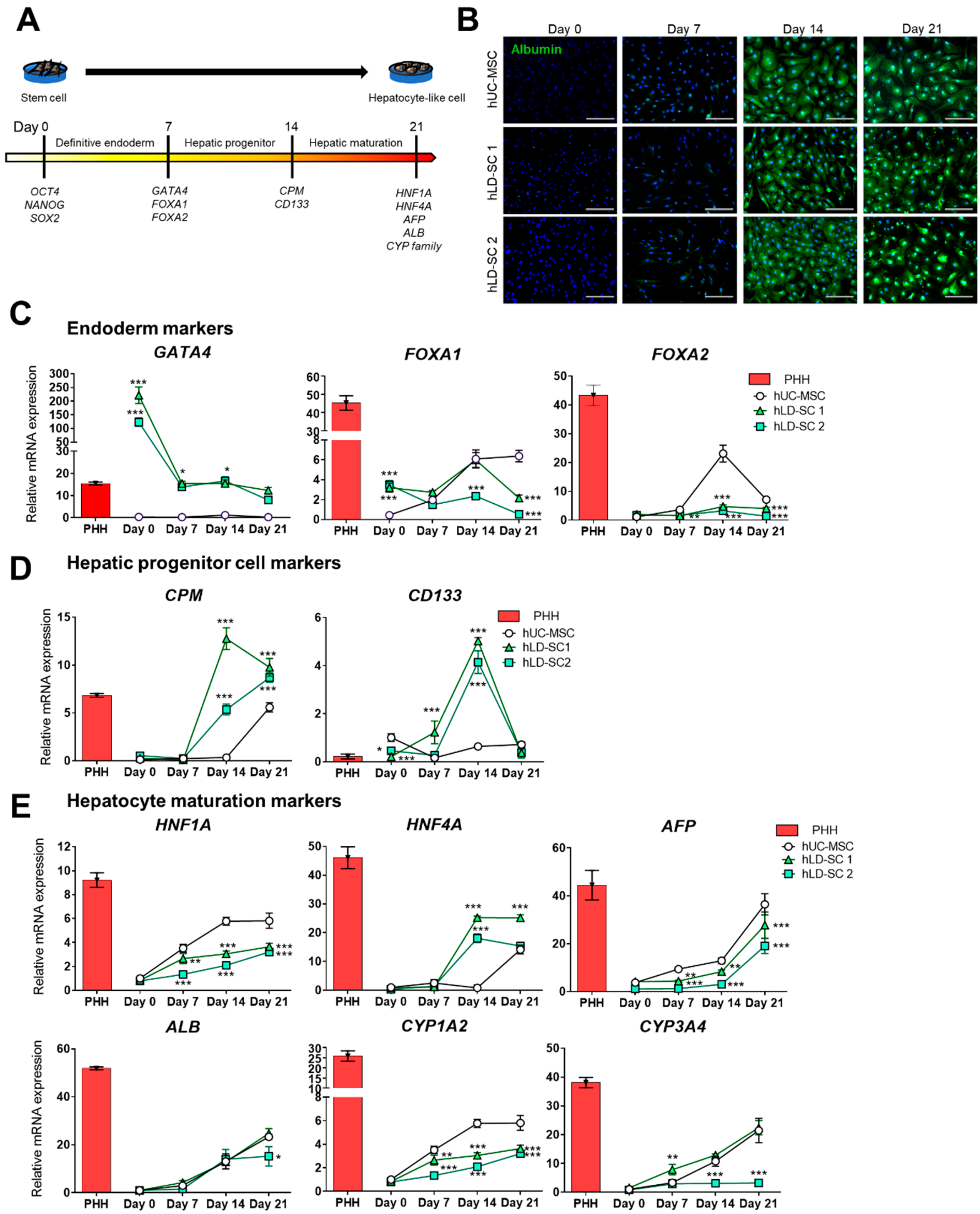
3.3. Comparison of Gene Expression Patterns during Hepatic Lineage Development
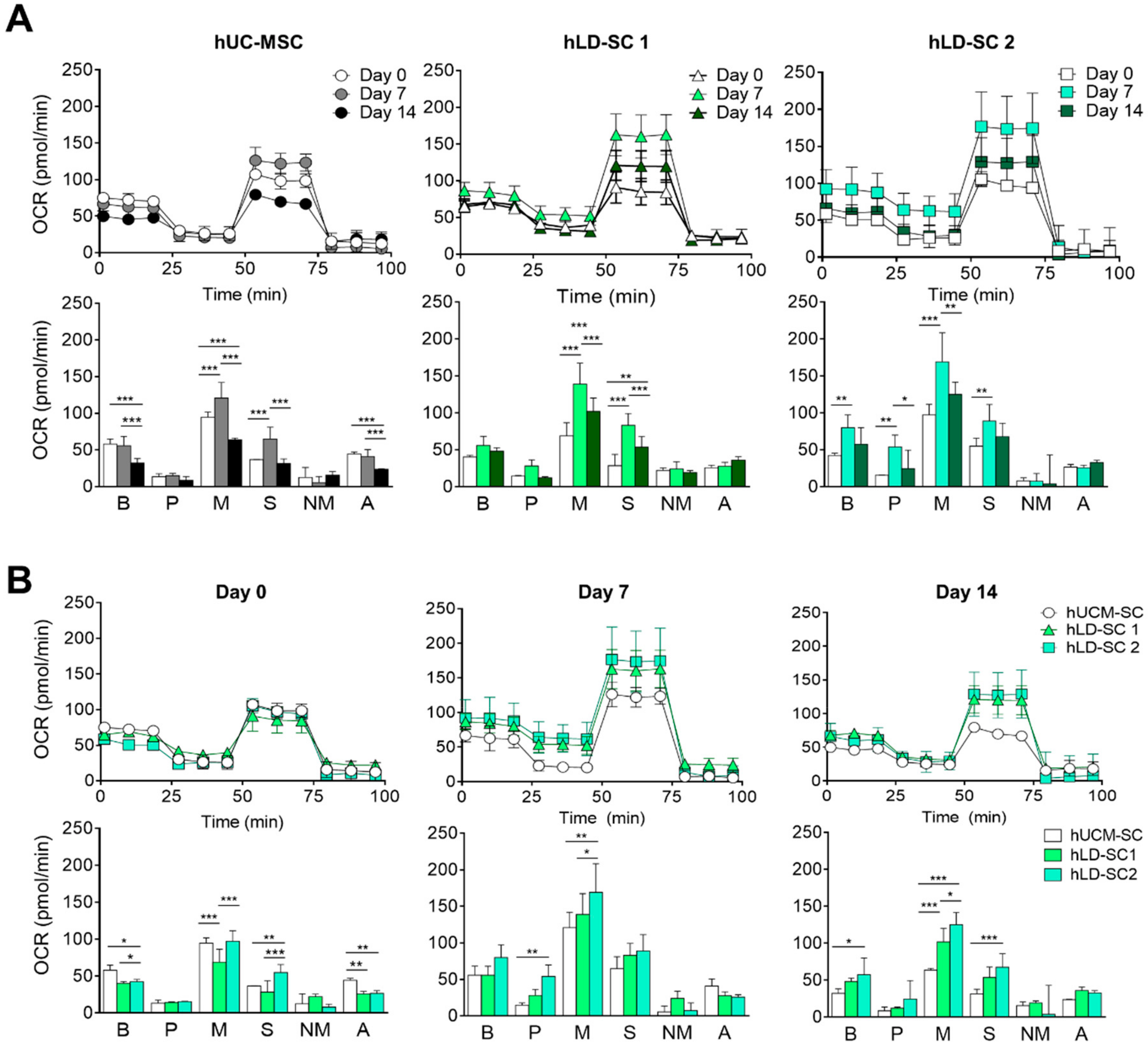
3.4. Mitochondrial Oxygen Consumption Rate (OCR) and Metabolic Profile Comparison
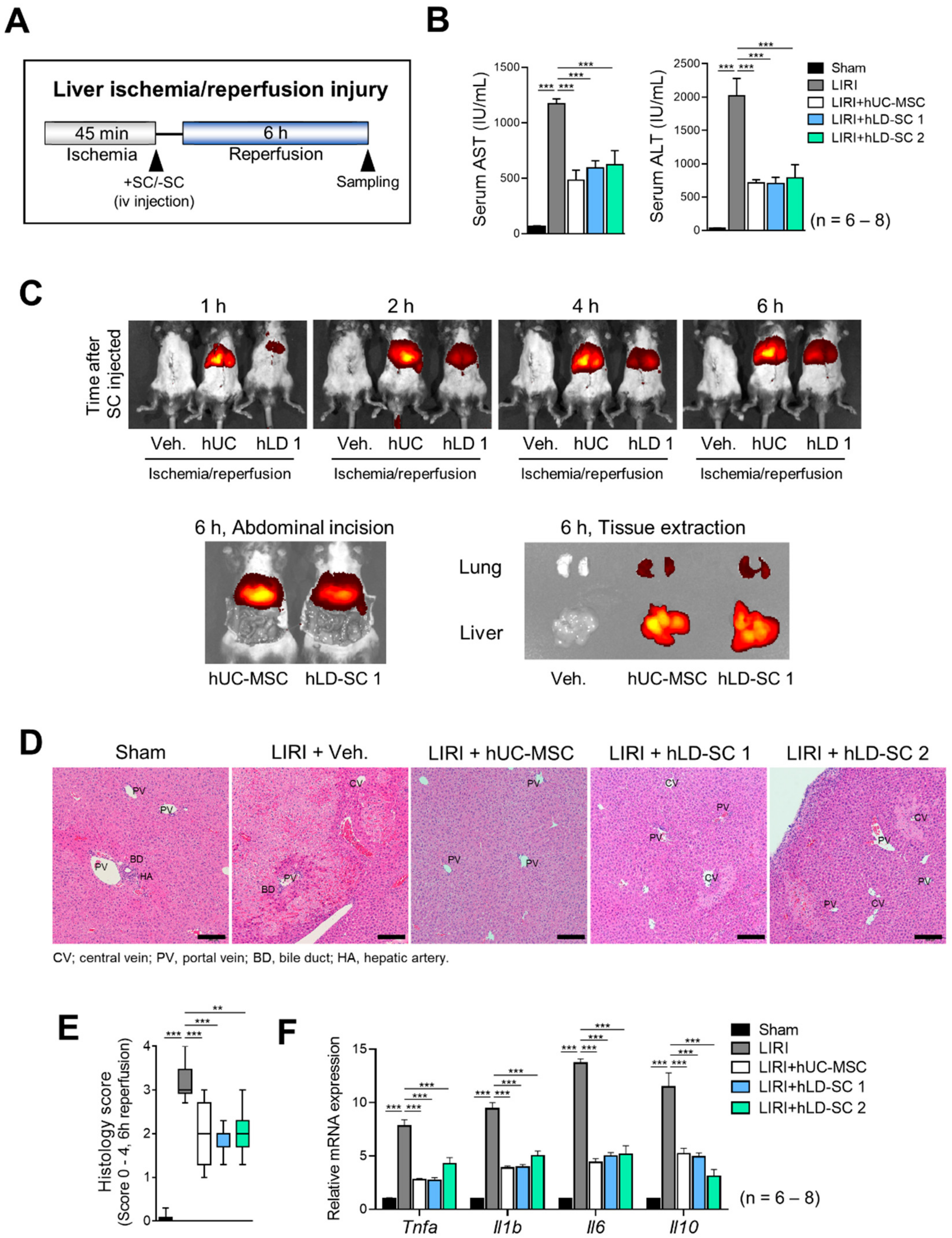
3.5. Trafficking of hLD-SCs during Liver Ischemia-Reperfusion Injury (LIRI)
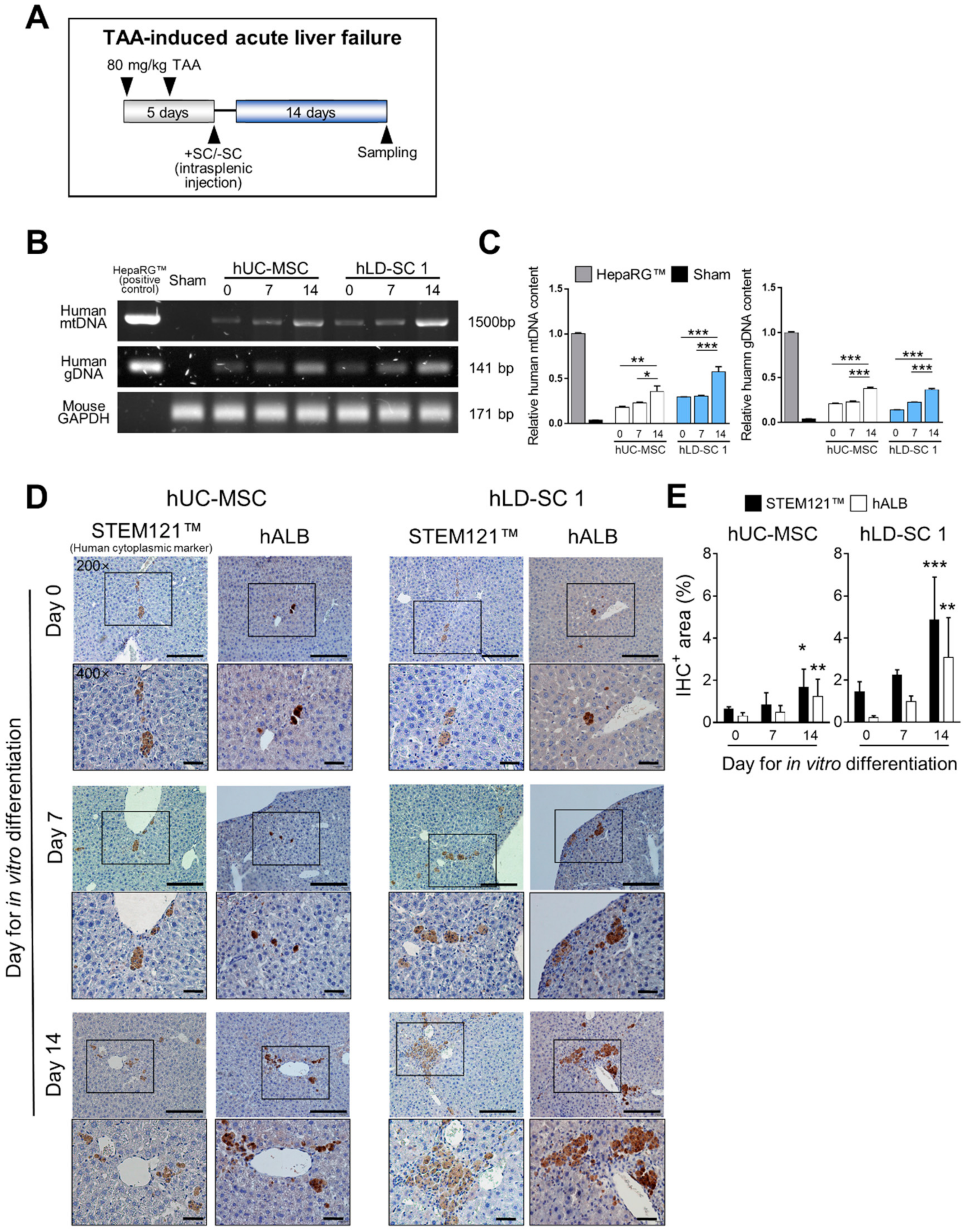
3.6. Spontaneous Differentiation and Regeneration Potential of Differentiation-Induced hLD-SCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Keeffe, E.B. Liver transplantation: Current status and novel approaches to liver replacement. Gastroenterology 2001, 120, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.B.; Lee, S.G. Liver transplantation. Gut Liver 2009, 3, 145–165. [Google Scholar] [CrossRef] [PubMed]
- Lantieri, L.; Grimbert, P.; Ortonne, N.; Suberbielle, C.; Bories, D.; Gil-Vernet, S.; Lemogne, C.; Bellivier, F.; Lefaucheur, J.P.; Schaffer, N.; et al. Face transplant: Long-term follow-up and results of a prospective open study. Lancet 2016, 388, 1398–1407. [Google Scholar] [CrossRef]
- Burra, P.; Tomat, S.; Villa, E.; Gasbarrini, A.; Costa, A.N.; Conconi, M.T.; Forbes, S.J.; Farinati, F.; Cozzi, E.; Alison, M.R.; et al. Experimental hepatology applied to stem cells. Dig. Liver Dis. 2008, 40, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Soto-Gutierrez, A.; Navarro-Alvarez, N.; Rivas-Carrillo, J.D.; Jun, H.S.; Kobayashi, N. Functional hepatocyte culture and its application to cell therapies. Cell Transplant. 2006, 15, 855–864. [Google Scholar] [CrossRef]
- Denker, H.W. Potentiality of embryonic stem cells: An ethical problem even with alternative stem cell sources. J. Med. Eth. 2006, 32, 665–671. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, X.; Nyberg, S.L. Potential and Challenges of Induced Pluripotent Stem Cells in Liver Diseases Treatment. J. Clin. Med. 2014, 3, 997–1017. [Google Scholar] [CrossRef] [Green Version]
- Meirelles, L.D.S.; Chagastelles, P.C.; Nardi, N.B. Mesenchymal stem cells reside in virtually all post-natal organs and tissues. J. Cell Sci. 2006, 119, 2204–2213. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.C.; Hung, S.C. Functional roles of pluripotency transcription factors in mesenchymal stem cells. Cell Cycle 2012, 11, 3711–3712. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, S.; Pittenger, M.F. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood 2005, 105, 1815–1822. [Google Scholar] [CrossRef] [Green Version]
- Kuo, T.K.; Ho, J.H.; Lee, O.K. Mesenchymal stem cell therapy for nonmusculoskeletal diseases: Emerging applications. Cell Transplant. 2009, 18, 1013–1028. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Liu, G.; Halim, A.; Ju, Y.; Luo, Q.; Song, A.G. Mesenchymal Stem Cell Migration and Tissue Repair. Cells 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Burra, P.; Tomat, S.; Bizzaro, D.; Pellicanò, D.; Masier, A.; Conconi, M.T.; Russo, F.; Bo, P.; Parnigotto, P.P. Stem cells in hepatology. Dig. Liver Dis. 2008, 40, 54–61. [Google Scholar] [CrossRef] [PubMed]
- van Poll, D.; Parekkadan, B.; Cho, C.H.; Berthiaume, F.; Nahmias, Y.; Tilles, A.W.; Yarmush, M.L. Mesenchymal stem cell-derived molecules directly modulate hepatocellular death and regeneration in vitro and in vivo. Hepatology 2008, 47, 1634–1643. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Wu, D.B.; Chen, B.; Chen, E.Q.; Tang, H. Progress in mesenchymal stem cell-based therapy for acute liver failure. Stem Cell Res. Ther. 2018, 9, 227. [Google Scholar] [CrossRef] [PubMed]
- Francois, S.; Mouiseddine, M.; Allenet-Lepage, B.; Voswinkel, J.; Douay, L.; Benderitter, M.; Chapel, A. Human mesenchymal stem cells provide protection against radiation-induced liver injury by antioxidative process, vasculature protection, hepatocyte differentiation, and trophic effects. BioMed Res. Int. 2013, 2013, 151679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campard, D.; Lysy, P.A.; Najimi, M.; Sokal, E.M. Native umbilical cord matrix stem cells express hepatic markers and differentiate into hepatocyte-like cells. Gastroenterology 2008, 134, 833–848. [Google Scholar] [CrossRef]
- Lee, K.D.; Kuo, T.K.; Whang-Peng, J.; Chung, Y.F.; Lin, C.T.; Chou, S.H.; Chen, J.R.; Chen, Y.P.; Lee, O.K. In vitro hepatic differentiation of human mesenchymal stem cells. Hepatology 2004, 40, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Y.; Zhang, L.; Li, J.; Zhu, C. Mesenchymal stem cells: Potential application for the treatment of hepatic cirrhosis. Stem Cell Res. Ther. 2018, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Tan, E.K.; Shuh, M.; Francois-Vaughan, H.; Sanders, J.A.; Cohen, A.J. Negligible Oval Cell Proliferation Following Ischemia-Reperfusion Injury With and Without Partial Hepatectomy. Ochsner J. 2017, 17, 31–37. [Google Scholar] [PubMed]
- Fausto, N. Liver regeneration and repair: Hepatocytes, progenitor cells, and stem cells. Hepatology 2004, 39, 1477–1487. [Google Scholar] [CrossRef] [PubMed]
- Shupe, T.D.; Piscaglia, A.C.; Oh, S.H.; Gasbarrini, A.; Petersen, B.E. Isolation and characterization of hepatic stem cells, or “oval cells,” from rat livers. Methods Mol. Biol. 2009, 482, 387–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Xin, J.; Zhang, L.; Wu, J.; Jiang, L.; Zhou, Q.; Li, J.; Guo, J.; Cao, H.; Li, L. Human hepatic progenitor cells express hematopoietic cell markers CD45 and CD109. Int. J. Med. Sci. 2014, 11, 65–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Kang, K.; Lee, S.B.; Seo, D.; Yoon, S.; Kim, S.J.; Jang, K.; Jung, Y.K.; Lee, K.G.; Factor, V.M.; et al. Small molecule-mediated reprogramming of human hepatocytes into bipotent progenitor cells. J. Hepatol. 2019, 70, 97–107. [Google Scholar] [CrossRef]
- Xie, B.Q.; Sun, D.; Du, Y.Y.; Jia, J.; Sun, S.C.; Xu, J.; Liu, Y.F.; Xiang, C.G.; Chen, S.T.; Xie, H.F.; et al. A two-step lineage reprogramming strategy to generate functionally competent human hepatocytes from fibroblasts. Cell Res. 2019, 29, 696–710. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Zhang, L.; Liu, W.; Ma, X.; Cen, J.; Sun, Z.; Wang, C.; Feng, S.; Zhang, Z.; Yue, L.; et al. In Vitro Expansion of Primary Human Hepatocytes with Efficient Liver Repopulation Capacity. Cell Stem Cell 2018, 23, 806–819. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liu, D.F.; Ma, Y.T.; Du, X.M.; Jing, J.Z.; Wang, L.P.; Xie, B.Q.; Sun, D.; Sun, S.Q.; Jin, X.Q.; et al. Direct Reprogramming of Fibroblasts via a Chemically Induced XEN-like State. Cell Stem Cell 2017, 21, 264–273. [Google Scholar] [CrossRef]
- Herrera, M.B.; Bruno, S.; Buttiglieri, S.; Tetta, C.; Gatti, S.; Deregibus, M.C.; Bussolati, B.; Camussi, G. Isolation and characterization of a stem cell population from adult human liver. Stem Cells 2006, 24, 2840–2850. [Google Scholar] [CrossRef]
- Najimi, M.; Khuu, D.N.; Lysy, P.A.; Jazouli, N.; Abarca, J.; Sempoux, C.; Sokal, E.M. Adult-derived human liver mesenchymal-like cells as a potential progenitor reservoir of hepatocytes? Cell Transplant. 2007, 16, 717–728. [Google Scholar] [CrossRef]
- Pan, Q.W.; Fouraschen, S.M.G.; Kaya, F.S.F.A.; Verstegen, M.M.; Pescatori, M.; Stubbs, A.P.; van IJcken, W.; van der Sloot, A.; Smits, R.; Kwekkeboom, J.; et al. Mobilization of Hepatic Mesenchymal Stem Cells From Human Liver Grafts. Liver Transplant. 2011, 17, 596–609. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Park, H.J.; Kim, Y.A.; Lee, D.H.; Noh, J.K.; Kwon, C.H.D.; Jung, S.M.; Lee, S.K. The Phenotypic Characteristic of Liver-Derived Stem Cells From Adult Human Deceased Donor Liver. Transplant. Proc. 2012, 44, 1110–1112. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.B.; Gupta, K.; Ananthanarayanan, A.; Wang, Z.N.; Xia, L.; Li, A.M.; Sakban, R.B.; Liu, S.D.; Yu, H. Directed Differentiation of Adult Liver Derived Mesenchymal Like Stem Cells into Functional Hepatocytes. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Yu, X.; Chen, E.; Li, L. Liver-derived human mesenchymal stem cells: A novel therapeutic source for liver diseases. Stem Cell Res. Ther. 2016, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- Kholodenko, I.V.; Kurbatov, L.K.; Kholodenko, R.V.; Manukyan, G.V.; Yarygin, K.N. Mesenchymal Stem Cells in the Adult Human Liver: Hype or Hope? Cells 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Li, K.; Wei, W.; Ding, S. Chemical approaches to stem cell biology and therapeutics. Cell Stem Cell 2013, 13, 270–283. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Chang, W.; Song, B.W.; Hwang, K.C. Specific differentiation of mesenchymal stem cells by small molecules. Am. J. Stem Cells 2012, 1, 22–30. [Google Scholar]
- Korostylev, A.; Mahaddalkar, P.U.; Keminer, O.; Hadian, K.; Schorpp, K.; Gribbon, P.; Lickert, H. A high-content small molecule screen identifies novel inducers of definitive endoderm. Mol. Metab. 2017, 6, 640–650. [Google Scholar] [CrossRef]
- So, S.; Lee, Y.; Choi, J.; Kang, S.; Lee, J.Y.; Hwang, J.; Shin, J.; Dutton, J.R.; Seo, E.J.; Lee, B.H.; et al. The Rho-associated kinase inhibitor fasudil can replace Y-27632 for use in human pluripotent stem cell research. PLoS ONE 2020, 15, e0233057. [Google Scholar] [CrossRef]
- He, Y.; Cui, J.; He, T.; Bi, Y. 5-azacytidine promotes terminal differentiation of hepatic progenitor cells. Mol. Med. Rep. 2015, 12, 2872–2878. [Google Scholar] [CrossRef]
- Kim, J.; Piao, Y.; Pak, Y.K.; Chung, D.; Han, Y.M.; Hong, J.S.; Jun, E.J.; Shim, J.-Y.; Choi, J.; Kim, C.J. Umbilical cord mesenchymal stromal cells affected by gestational diabetes mellitus display premature aging and mitochondrial dysfunction. Stem Cells Dev. 2014, 24, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Yang, G.; Park, J.; Choi, J.; Kang, E.; Lee, B.K. Therapeutic effect of mesenchymal stem cells derived from the human umbilical cord in a rabbit temporomandibular joint model of osteoarthritis. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, B.-W.; Kang, E.-J.; Byun, J.-H.; Son, M.-G.; Kim, H.-J.; Hah, Y.-S.; Kim, T.-H.; Mohana Kumar, B.; Ock, S.-A.; Rho, G.-J. In vitro and in vivo osteogenesis of human mesenchymal stem cells derived from skin, bone marrow and dental follicle tissues. Differentiation 2012, 83, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.J.; Wang, X.J.; Tippner-Hedges, R.; Ma, H.; Folmes, C.D.L.; Gutierrez, N.M.; Lee, Y.M.; Van Dyken, C.; Ahmed, R.; Li, Y.; et al. Age-Related Accumulation of Somatic Mitochondrial DNA Mutations in Adult-Derived Human iPSCs. Cell Stem Cell 2016, 18, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Toledo-Pereyra, L.H.; Rodriguez, F.J.; Cejalvo, D. Neutrophil infiltration as an important factor in liver ischemia and reperfusion injury. Transplantation 1993, 55, 1265–1271. [Google Scholar]
- Kyurkchiev, D.; Bochev, I.; Ivanova-Todorova, E.; Mourdjeva, M.; Oreshkova, T.; Belemezova, K.; Kyurkchiev, S. Secretion of immunoregulatory cytokines by mesenchymal stem cells. World J. Stem Cells 2014, 6, 552–570. [Google Scholar] [CrossRef]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef]
- Bloom, D.D.; Centanni, J.M.; Bhatia, N.; Emler, C.A.; Drier, D.; Leverson, G.E.; McKenna, D.H., Jr.; Gee, A.P.; Lindblad, R.; Hei, D.J.; et al. A reproducible immunopotency assay to measure mesenchymal stromal cell-mediated T-cell suppression. Cytotherapy 2015, 17, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Meseguer-Ripolles, J.; Khetani, S.R.; Blanco, J.G.; Iredale, M.; Hay, D.C. Pluripotent Stem Cell-Derived Human Tissue: Platforms to Evaluate Drug Metabolism and Safety. AAPS J. 2018, 20, 20. [Google Scholar] [CrossRef] [Green Version]
- Chaudhari, P.; Tian, L.; Deshmukh, A.; Jang, Y.Y. Expression kinetics of hepatic progenitor markers in cellular models of human liver development recapitulating hepatocyte and biliary cell fate commitment. Exp. Biol. Med. 2016, 241, 1653–1662. [Google Scholar] [CrossRef] [Green Version]
- Lau, H.H.; Ng, N.H.J.; Loo, L.S.W.; Jasmen, J.B.; Teo, A.K.K. The molecular functions of hepatocyte nuclear factors - In and beyond the liver. J. Hepatol. 2018, 68, 1033–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchiya, A.; Lu, W.Y. Liver stem cells: Plasticity of the liver epithelium. World J. Gastroenterol. 2019, 25, 1037–1049. [Google Scholar] [CrossRef] [PubMed]
- Kido, T.; Koui, Y.; Suzuki, K.; Kobayashi, A.; Miura, Y.; Chern, E.Y.; Tanaka, M.; Miyajima, A. CPM Is a Useful Cell Surface Marker to Isolate Expandable Bi-Potential Liver Progenitor Cells Derived from Human iPS Cells. Stem Cell Rep. 2015, 5, 508–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizrak, D.; Brittan, M.; Alison, M.R. CD133: Molecule of the moment. J. Pathol. 2008, 214, 3–9. [Google Scholar] [CrossRef]
- Zhao, D.X.; Chen, S.; Cai, J.; Guo, Y.S.; Song, Z.H.; Che, J.; Liu, C.; Wu, C.; Ding, M.X.; Deng, H.K. Derivation and Characterization of Hepatic Progenitor Cells from Human Embryonic Stem Cells. PLoS ONE 2009, 4, e6468. [Google Scholar] [CrossRef] [Green Version]
- Okabe, M.; Tsukahara, Y.; Tanaka, M.; Suzuki, K.; Saito, S.; Kamiya, Y.; Tsujimura, T.; Nakamura, K.; Miyajima, A. Potential hepatic stem cells reside in EpCAM+ cells of normal and injured mouse liver. Development 2009, 136, 1951–1960. [Google Scholar] [CrossRef] [Green Version]
- Gires, O. EpCAM in hepatocytes and their progenitors. J. Hepatol. 2012, 56, 490–492. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Chen, Y.; Fu, G.B.; Yuan, T.J.; Huang, W.J.; Wang, Z.Y.; Li, W.J.; Jiao, Y.F.; Yu, W.F.; Yan, H.X. EpCAM inhibits differentiation of human liver progenitor cells into hepatocytes in vitro by activating Notch1 signaling. Biochem. Biophys. Res. Commun. 2020. [Google Scholar] [CrossRef]
- Furuyama, K.; Kawaguchi, Y.; Akiyama, H.; Horiguchi, M.; Kodama, S.; Kuhara, T.; Hosokawa, S.; Elbahrawy, A.; Soeda, T.; Koizumi, M.; et al. Continuous cell supply from a Sox9-expressing progenitor zone in adult liver, exocrine pancreas and intestine. Nat. Genet. 2011, 43, 34–41. [Google Scholar] [CrossRef]
- Shen, Y.L.; Wu, L.; Qin, D.D.; Xia, Y.K.; Zhou, Z.; Zhang, X.M.; Wu, X. Carbon black suppresses the osteogenesis of mesenchymal stem cells: The role of mitochondria. Part. Fibre Toxicol. 2018, 15, 16. [Google Scholar] [CrossRef]
- Forni, M.F.; Peloggia, J.; Trudeau, K.; Shirihai, O.; Kowaltowski, A.J. Murine Mesenchymal Stem Cell Commitment to Differentiation Is Regulated by Mitochondrial Dynamics. Stem Cells 2016, 34, 743–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, M.; Liu, D.D.; Thakor, A.S. Mesenchymal Stromal Cell Homing: Mechanisms and Strategies for Improvement. iScience 2019, 15, 421–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, M.F. Applications for ROCK kinase inhibition. Curr. Opin. Cell Biol. 2008, 20, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.; Li, Q.Y.; Yu, J.Z.; Wang, X.; Sun, C.H.; Lu, C.Z.; Xiao, B.G. Fasudil, a Rho kinase inhibitor, drives mobilization of adult neural stem cells after hypoxia/reoxygenation injury in mice. Mol. Cell. Neurosci. 2010, 43, 201–208. [Google Scholar] [CrossRef]
- Tsuno, A.; Nasu, K.; Kawano, Y.; Yuge, A.; Li, H.L.; Abe, W.; Narahara, H. Fasudil Inhibits the Proliferation and Contractility and Induces Cell Cycle Arrest and Apoptosis of Human Endometriotic Stromal Cells: A Promising Agent for the Treatment of Endometriosis. J. Clin. Endocrinol. Metab. 2011, 96, E1944–E1952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.-W.; Huang, W.-C.; Huang, H.-D.; Huang, Y.-H.; Ho, J.H.; Yang, M.-H.; Yang, V.W.; Lee, O.K. DNA methyltransferases modulate hepatogenic lineage plasticity of mesenchymal stromal cells. Stem Cell Rep. 2017, 9, 247–263. [Google Scholar] [CrossRef] [Green Version]
- Fisher, J.B.; Pulakanti, K.; Rao, S.; Duncan, S.A. GATA6 is essential for endoderm formation from human pluripotent stem cells. Biol. Open 2017, 6, 1084–1095. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; McKnight, K.D.; Wong, D.J.; Rodriguez, R.T.; Sugiyama, T.; Gu, X.Y.; Ghodasara, A.; Qu, K.; Chang, H.Y.; Kim, S.K. A Molecular Signature for Purified Definitive Endoderm Guides Differentiation and Isolation of Endoderm from Mouse and Human Embryonic Stem Cells. Stem Cells Dev. 2012, 21, 2273–2287. [Google Scholar] [CrossRef] [Green Version]
- Kordes, C.; Sawitza, I.; Muller-Marbach, A.; Ale-Agha, N.; Keitel, V.; Klonowski-Stumpe, H.; Haussinger, D. CD133(+) hepatic stellate cells are progenitor cells. Biochem. Biophys. Res. Commun. 2007, 352, 410–417. [Google Scholar] [CrossRef] [Green Version]
- Van Eyken, P.; Sciot, R.; Callea, F.; Van der Steen, K.; Moerman, P.; Desmet, V.J. The development of the intrahepatic bile ducts in man: A keratin-immunohistochemical study. Hepatology 1988, 8, 1586–1595. [Google Scholar] [CrossRef]
- Wruck, W.; Adjaye, J. Human pluripotent stem cell derived HLC transcriptome data enables molecular dissection of hepatogenesis. Sci. Data 2018, 5, 180035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanet, A.; Arnould, T.; Najimi, M.; Renard, P. Connecting Mitochondria, Metabolism, and Stem Cell Fate. Stem Cells Dev. 2015, 24, 1957–1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birket, M.J.; Orr, A.L.; Gerencser, A.A.; Madden, D.T.; Vitelli, C.; Swistowski, A.; Brand, M.D.; Zeng, X.M. A reduction in ATP demand and mitochondrial activity with neural differentiation of human embryonic stem cells. J. Cell Sci. 2011, 124, 348–358. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.Q.; Gao, Z.W.; Chen, Y.; Guan, M.X. The role of mitochondria in osteogenic, adipogenic and chondrogenic differentiation of mesenchymal stem cells. Protein Cell 2017, 8, 439–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.X.; Fan, L.X.; Cen, P.P.; Chen, E.M.; Jiang, Z.Y.; Li, L.J. Energy Metabolism Plays a Critical Role in Stem Cell Maintenance and Differentiation. Int. J. Mol. Sci. 2016, 17, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkinson, B.M.; Desler, C.; Kalisz, M.; Vestentoft, P.S.; Rasmussen, L.J.; Bisgaard, H.C. Bioenergetic Changes during Differentiation of Human Embryonic Stem Cells along the Hepatic Lineage. Oxid. Med. Cell. Longev. 2017, 2017, 5080128. [Google Scholar] [CrossRef] [PubMed]
- Abu-Amara, M.; Yang, S.Y.; Tapuria, N.; Fuller, B.; Davidson, B.; Seifalian, A. Liver ischemia/reperfusion injury: Processes in inflammatory networks—A review. Liver Transplant. 2010, 16, 1016–1032. [Google Scholar] [CrossRef]
- Teoh, N.; Leclercq, I.; Pena, A.D.; Farrell, G. Low-dose TNF-α protects against hepatic ischemia-reperfusion injury in mice: Implications for preconditioning. Hepatology 2003, 37, 118–128. [Google Scholar] [CrossRef]
- Najimi, M.; Berardis, S.; El-Kehdy, H.; Rosseels, V.; Evraerts, J.; Lombard, C.; El Taghdouini, A.; Henriet, P.; van Grunsven, L.; Sokal, E.M. Human liver mesenchymal stem/progenitor cells inhibit hepatic stellate cell activation: In vitro and in vivo evaluation. Stem Cell Res. Ther. 2017, 8, 131. [Google Scholar] [CrossRef]
- Khuu, D.N.; Nyabi, O.; Maerckx, C.; Sokal, E.; Najimi, M. Adult Human Liver Mesenchymal Stem/Progenitor Cells Participate in Mouse Liver Regeneration After Hepatectomy. Cell Transplant. 2013, 22, 1369–1380. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Liver Donor | 1 | 2 | 3 | Reference Range |
|---|---|---|---|---|
| Ethnicity | Asian | Asian | Asian | |
| Sex | Male | Female | Female | |
| Age (years) | 21 | 38 | 27 | |
| Body mass index | 21.18 | 25.02 | 20.56 | |
| Diagnosis | Living liverdonor | Living liverdonor | Living liverdonor | |
| Operation | Rightlobectomy | Rightlobectomy | Rightlobectomy | |
| Albumin (g/dL) | 4.2 | 3.9 | 4.6 | 3.5–5.2 |
| Total bilirubin (mg/dL) | 0.2 | 0.4 | 0.7 | 0.2–1.2 |
| AST (U/L) | 20 | 20 | 21 | < 40 |
| ALT (U/L) | 14 | 19 | 16 | < 40 |
| ABO group | O+ | B+ | B+ | |
| HAV (COI) | Negative | Negative | Negative | Negative |
| HBV (U/mL) | Negative | Negative | Negative | Negative |
| HCV (COI) | Negative | Negative | Negative | Negative |
| Glucose (mg/dL) | 94 | 89 | 94 | 70–99 |
| Creatinine (mg/dL) | 0.81 | 0.79 | 0.77 | 0.70–1.40 |
| BUN (mg/dL) | 12 | 8 * | 10 | 10–26 |
| Uric acid (mg/dL) | 6.7 | 4.6 | 5.4 | 3–7 |
| Total cholesterol (mg/dL) | 146 | 140 | 235 * | < 199 |
| Total protein (g/dL) | 7.1 | 7.0 | 7.7 | 6–8 |
| Alkaline phosphatase (U/L) | 73 | 63 | 58 | 40–120 |
| Gamma-GT (U/L) | 13 | 19 | 17 | 5–36 |
| Triglyceride (mg/dL) | 51 | 157 | 56 | < 199 |
| HDL-cholesterol (mg/dL) | 66 | 52 | 90 * | 40 < |
| Gene Name | Primer Sequences (5′ to 3′) |
|---|---|
| Human | |
| OCT4 | Forward: GAAGGATGTGGTCCGAGTGT Reverse: GTGAAGTGAGGGCTCCCATA |
| NANOG | Forward: CAAAGGCAAACAACCCACTT Reverse: TCTGCTGGAGGCTGAGGTAT |
| SOX2 | Forward: AACCCCAAGATGCACAACTC Reverse: CGGGGCCGGTATTTATAATC |
| TNFα | Forward: CAGAGGGCCTGTACCTCATC Reverse: GGAAGACCCCTCCCAGATAG |
| IL1β | Forward: GGGCCTCAAGGAAAAGAATC Reverse: TTCTGCTTGAGAGGTGCTGA |
| IL6 | Forward: TACCCCCAGGAGAAGATTCC Reverse: TTTTCTGCCAGTGCCTCTTT |
| TGFβ | Forward: GCCATTTAATGGCAATGGTAGTCT Reverse: CACAGGGAGCTTGCAGAGAT |
| IL10 | Forward: TGCCTTCAGCAGAGTGAAGA Reverse: GGTCTTGGTTCTCAGCTTGG |
| GATA4 | Forward: GGCCTGTCATCTCACTACGG Reverse: ATGGCCAGACATCGCACT |
| FOXA1 | Forward: GACTCCAGCCTCCTCAACTG Reverse: CTCCGAGGAGGACATGAGGT |
| FOXA2 | Forward: CTTCAAGCACCTGCAGATTC Reverse: AGACCTGGATTTCACCGTGT |
| CPM | Forward: GGATGGAAGCGTTTTTGAAG Reverse: CCACAACAAGAACCCACAGG |
| CD133 | Forward: AAGCATTGGCATCTTCTATGG Reverse: AAGCACAGAGGGTCATTGAGA |
| HNF1A | Forward: TGGGTCCTACGTTCACCAAC Reverse: TCTGCACAGGTGGCATGAGC |
| HNF4A | Forward: CAGGCTCAAGAAATGCTTCC Reverse: GGCTGCTGTCCTCATAGCTT |
| AFP | Forward: AGACTGCTGCAGCCAAAGTGA Reverse: GTGGGATCGATGCTGGAGTG |
| ALB | Forward: TGCTGATGAGTCAGCTGAAAA Reverse: TCAGCCATTTCACCATAGGTT |
| CYP1A2 | Forward: CTTCGTAAACCAGTGGCAGG Reverse: AGGGCTTGTTAATGGCAGTG |
| CYP3A4 | Forward: TTTTGTCCTACCATAAGGGCTTT Reverse: CACAGGCTGTTGACCATCAT |
| GAPDH | Forward: GCCTCAAGATCATCAGCAATGC Reverse: TGGTCATGAGTCCTTCCACGAT |
| Human mitochondrial DNA | Forward: CCCAAGACAACCAACCAAAA Reverse: ACTAGCTTATATGCTTGGGG |
| Human genomic DNA | Forward: ATGCTGATGTCTGGGTAGGGTG Reverse: TGAGTCAGGAGCCAGCGTATG |
| Mouse | |
| Tnfa | Forward: AGCCCCCAGTCTGTATCCTT Reverse: CTCCCTTTGCAGAACTCAGG |
| Il1b | Forward: GCCCATCCTCTGTGACTCAT Reverse: AGGCCACAGGTATTTTGTCG |
| Il6 | Forward: AGTTGCCTTCTTGGGACTGA Reverse: TCCACGATTTCCCAGAGAAC |
| Il10 | Forward: CCAAGCCTTATCGGAAATGA Reverse: TTTTCACAGGGGAGAAATCG |
| GAPDH | Forward: AGGTCGGTGTGAACGGATTTG Reverse: TGTAGACCATGTAGTTGAGGTCA |
| GAPDH genomic DNA | Forward: ACCCAGAAGACTGTGGATGG Reverse: CACATTGGGGGTAGGAACAC |
| Score | Congestion | Cytoplasmic Vacuolization | Parenchymal Necrosis |
|---|---|---|---|
| 0 | No | No | No |
| 1 | Minimal | Minimal | Single-cell necrosis |
| 2 | Mild | Mild | <30% |
| 3 | Moderate | Moderate | <60% |
| 4 | Severe | Severe | >60% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.; Choi, J.; Kang, S.; Kim, J.; Lee, R.; So, S.; Yoon, Y.-I.; Kirchner, V.A.; Song, G.-W.; Hwang, S.; et al. Hepatogenic Potential and Liver Regeneration Effect of Human Liver-derived Mesenchymal-Like Stem Cells. Cells 2020, 9, 1521. https://doi.org/10.3390/cells9061521
Lee J, Choi J, Kang S, Kim J, Lee R, So S, Yoon Y-I, Kirchner VA, Song G-W, Hwang S, et al. Hepatogenic Potential and Liver Regeneration Effect of Human Liver-derived Mesenchymal-Like Stem Cells. Cells. 2020; 9(6):1521. https://doi.org/10.3390/cells9061521
Chicago/Turabian StyleLee, Jooyoung, Jiwan Choi, Seoon Kang, Jiye Kim, Ryunjin Lee, Seongjun So, Young-In Yoon, Varvara A. Kirchner, Gi-Won Song, Shin Hwang, and et al. 2020. "Hepatogenic Potential and Liver Regeneration Effect of Human Liver-derived Mesenchymal-Like Stem Cells" Cells 9, no. 6: 1521. https://doi.org/10.3390/cells9061521
APA StyleLee, J., Choi, J., Kang, S., Kim, J., Lee, R., So, S., Yoon, Y. -I., Kirchner, V. A., Song, G. -W., Hwang, S., Lee, S. -G., Kang, E., & Tak, E. (2020). Hepatogenic Potential and Liver Regeneration Effect of Human Liver-derived Mesenchymal-Like Stem Cells. Cells, 9(6), 1521. https://doi.org/10.3390/cells9061521