Daily Supplementation of L-Glutamine in Atrial Fibrillation Patients: The Effect on Heat Shock Proteins and Metabolites

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
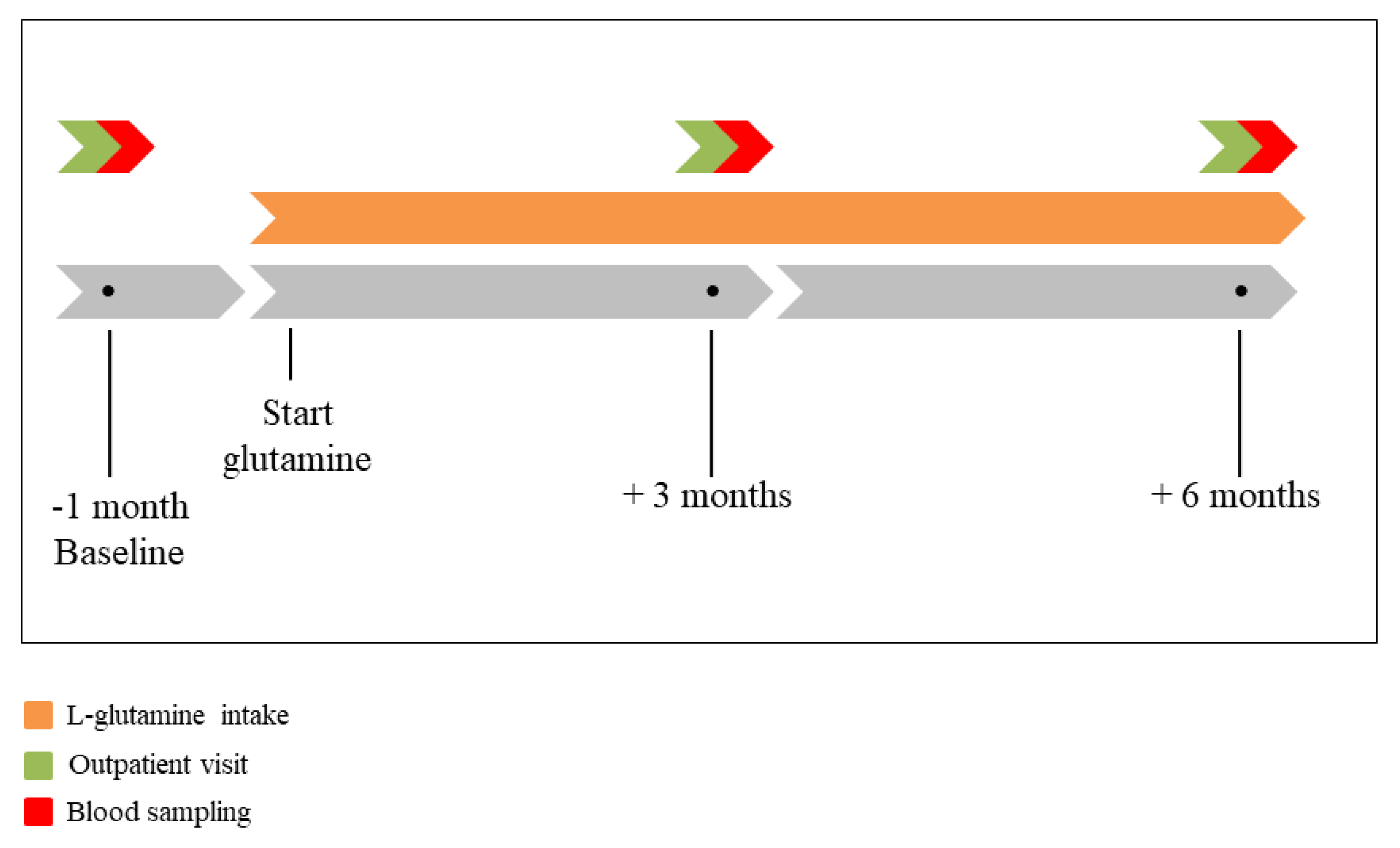
2.2. Study Design
2.3. Data Analysis
2.3.1. HSP Measurements
2.3.2. Metabolomics
2.4. Statistical Analysis
3. Results
3.1. Study Population
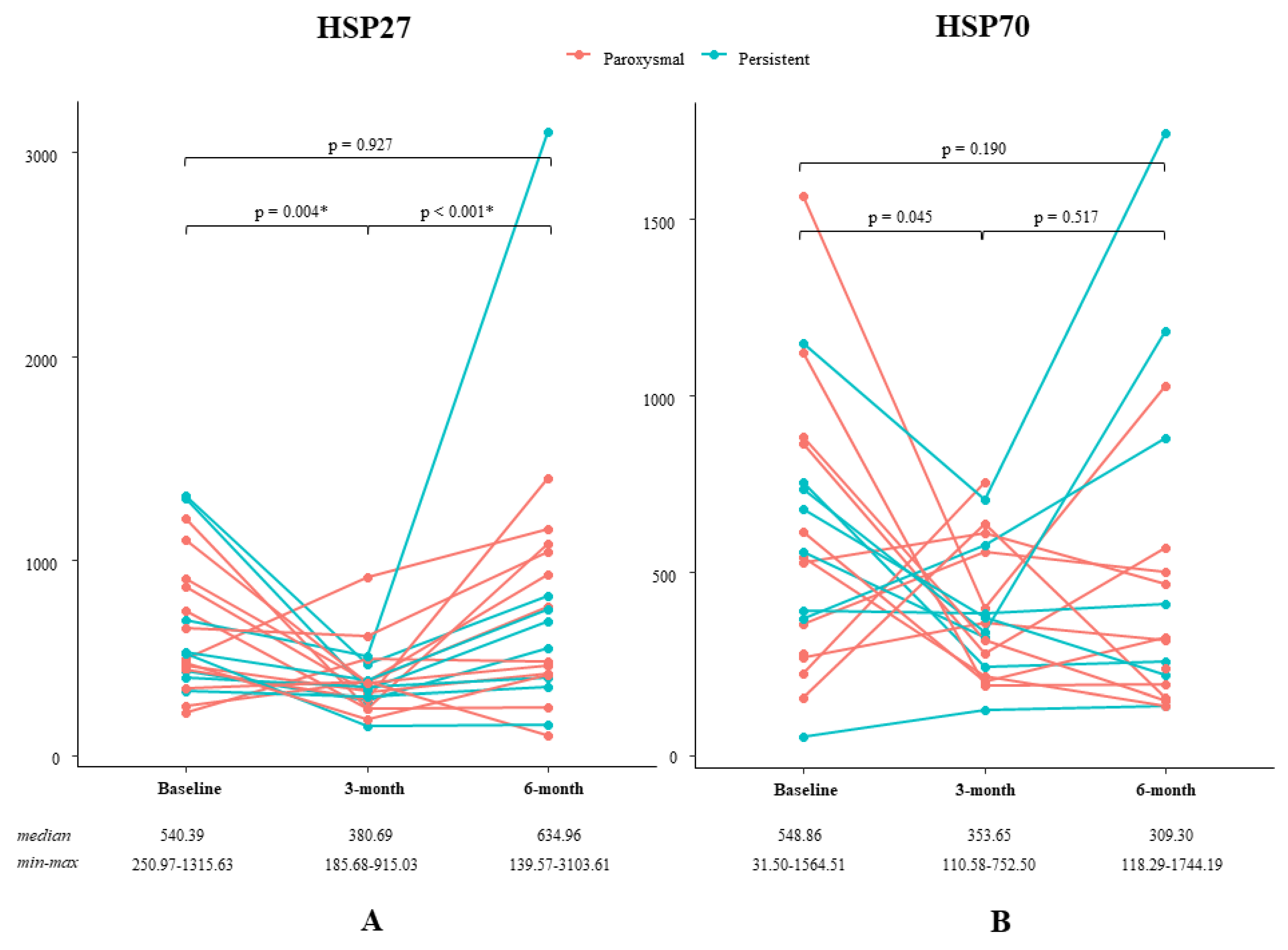
3.2. Effect of L-Glutamine on HSP Levels
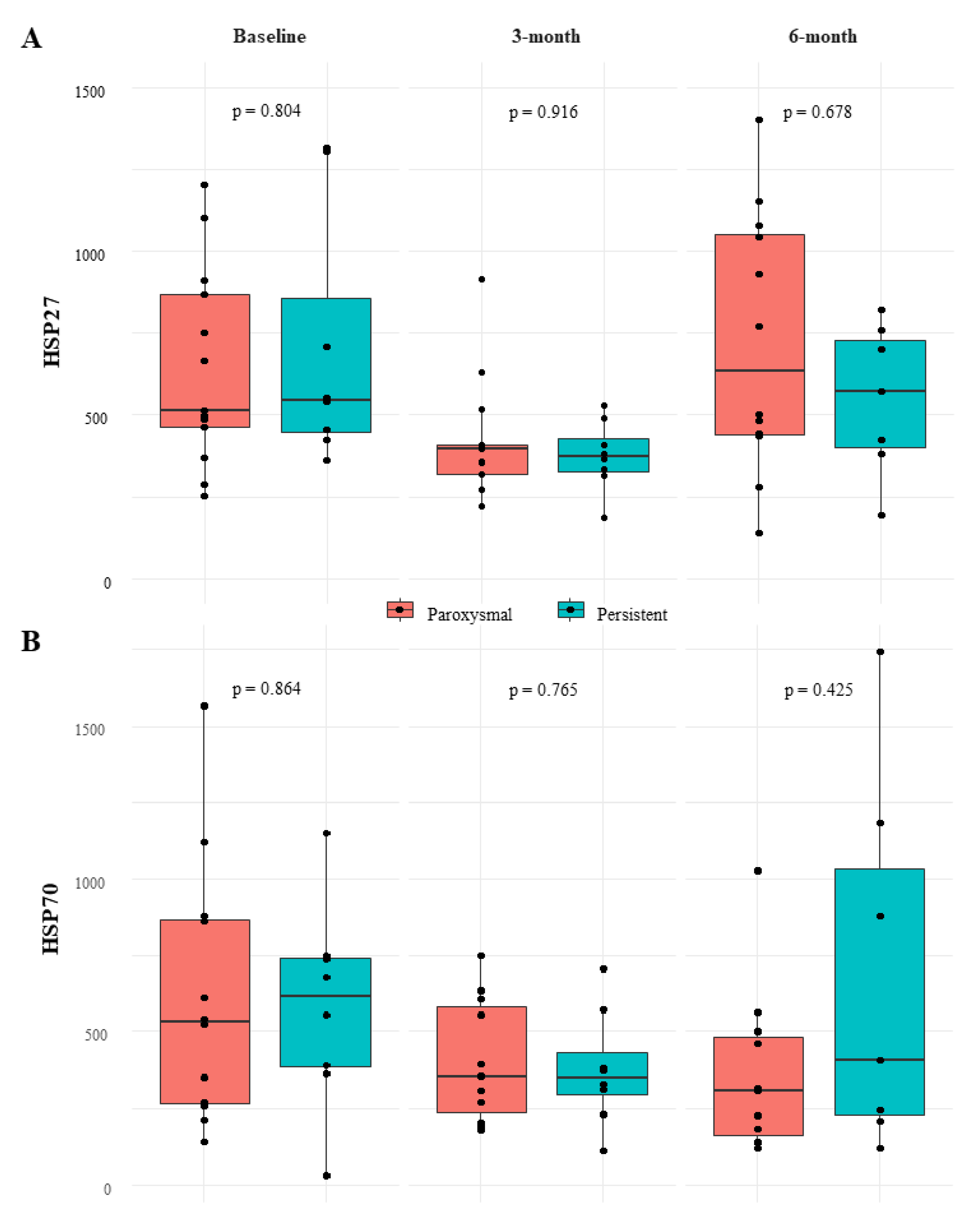
3.3. Relation between HSP Levels and Patient Characteristics
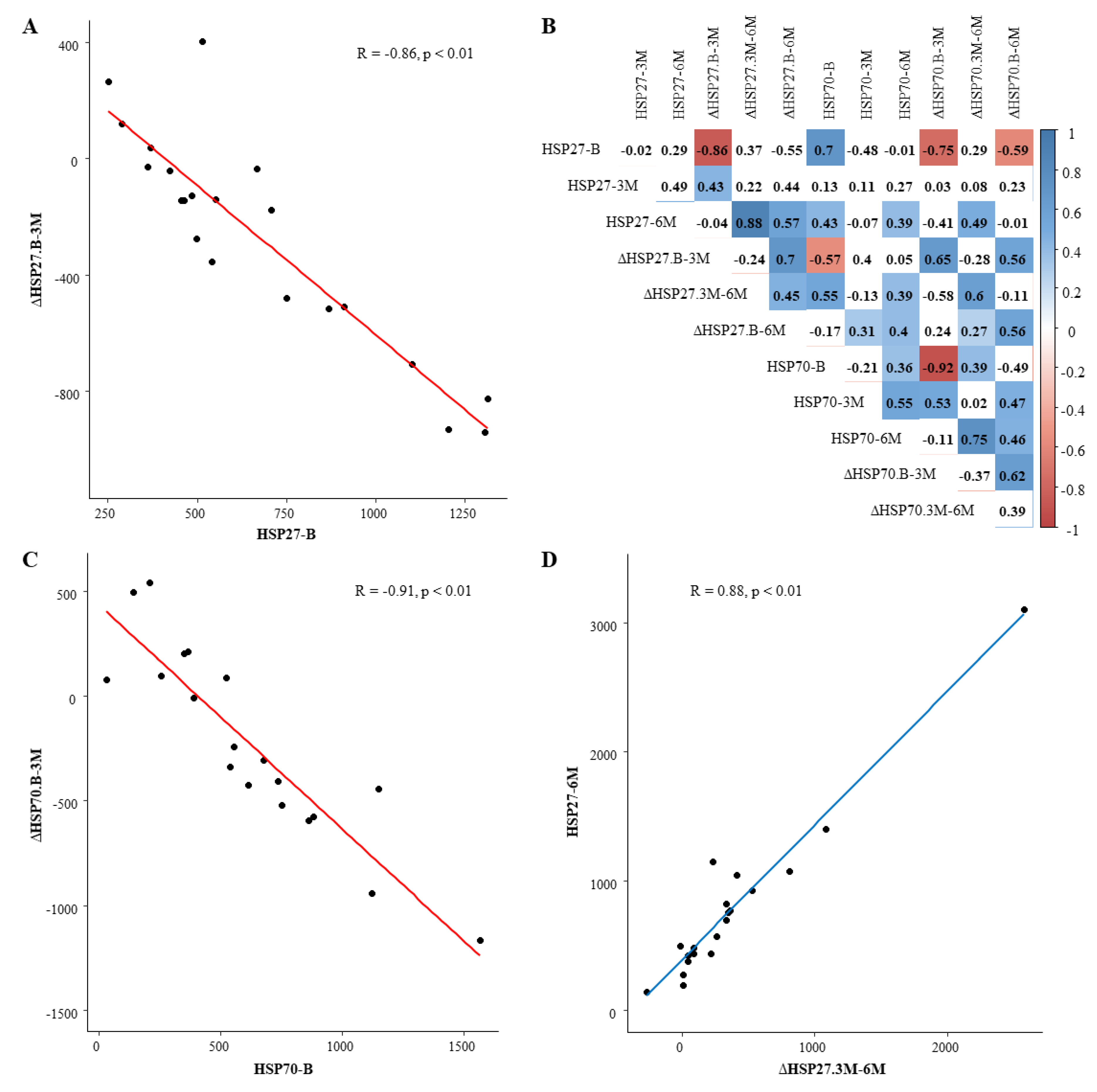
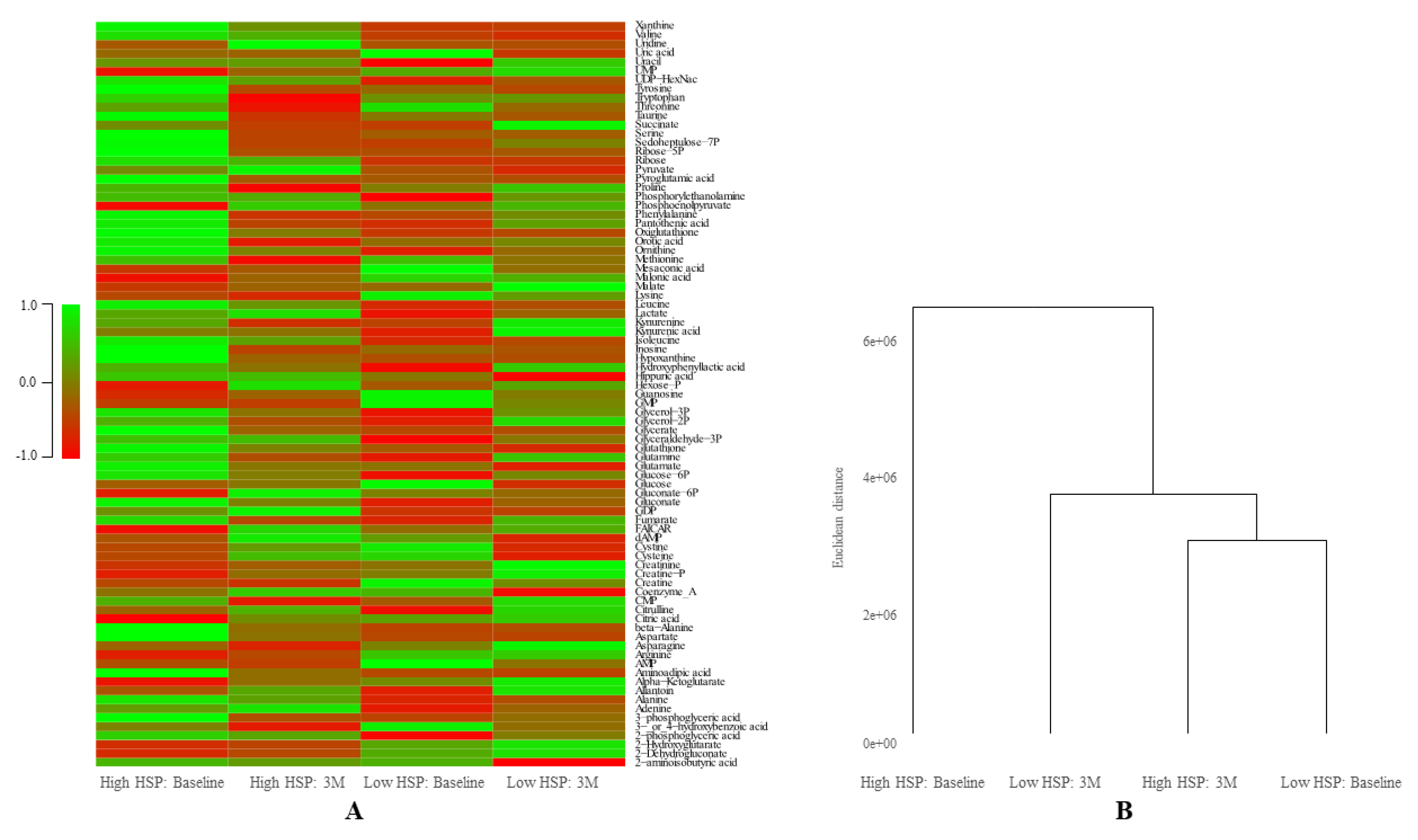
3.4. Relation between HSP27 and Energetic Metabolism
4. Discussion
4.1. Effect of L-glutamine on HSP and Metabolite Levels
4.2. Study Limitations and Future Perspectives
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Internal Standards for Metabolomics
References
- Kirchhof, P.; Benussi, S.; Kotecha, D.; Ahlsson, A.; Atar, D.; Casadei, B.; Castella, M.; Diener, H.C.; Heidbuchel, H.; Hendriks, J.; et al. 2016 esc guidelines for the management of atrial fibrillation developed in collaboration with eacts. Eur. Heart J. 2016, 37, 2893–2962. [Google Scholar] [CrossRef] [Green Version]
- Zoni-Berisso, M.; Lercari, F.; Carazza, T.; Domenicucci, S. Epidemiology of atrial fibrillation: European perspective. Clin. Epidemiol. 2014, 6, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Sultan, A.; Luker, J.; Andresen, D.; Kuck, K.H.; Hoffmann, E.; Brachmann, J.; Hochadel, M.; Willems, S.; Eckardt, L.; Lewalter, T.; et al. Predictors of atrial fibrillation recurrence after catheter ablation: Data from the german ablation registry. Sci. Rep. 2017, 7, 16678. [Google Scholar] [CrossRef] [Green Version]
- van Marion, D.M.; Lanters, E.A.; Wiersma, M.; Allessie, M.A.; Brundel, B.B.; de Groot, N.M. Diagnosis and therapy of atrial fibrillation: The past, the present and the future. J. Atr. Fibrillation 2015, 8, 1216. [Google Scholar] [PubMed]
- Wiersma, M.; van Marion, D.M.S.; Wust, R.C.I.; Houtkooper, R.H.; Zhang, D.; Groot, N.M.S.; Henning, R.H.; Brundel, B. Mitochondrial dysfunction underlies cardiomyocyte remodeling in experimental and clinical atrial fibrillation. Cells 2019, 8, 1202. [Google Scholar] [CrossRef] [Green Version]
- Brundel, B.J.; Henning, R.H.; Ke, L.; van Gelder, I.C.; Crijns, H.J.; Kampinga, H.H. Heat shock protein upregulation protects against pacing-induced myolysis in hl-1 atrial myocytes and in human atrial fibrillation. J. Mol. Cell Cardiol. 2006, 41, 555–562. [Google Scholar] [CrossRef]
- Zhang, D.; Ke, L.; Mackovicova, K.; Van Der Want, J.J.; Sibon, O.C.; Tanguay, R.M.; Morrow, G.; Henning, R.H.; Kampinga, H.H.; Brundel, B.J. Effects of different small hspb members on contractile dysfunction and structural changes in a drosophila melanogaster model for atrial fibrillation. J. Mol. Cell Cardiol. 2011, 51, 381–389. [Google Scholar] [CrossRef]
- Hoogstra-Berends, F.; Meijering, R.A.; Zhang, D.; Heeres, A.; Loen, L.; Seerden, J.P.; Kuipers, I.; Kampinga, H.H.; Henning, R.H.; Brundel, B.J. Heat shock protein-inducing compounds as therapeutics to restore proteostasis in atrial fibrillation. Trends Cardiovasc. Med. 2012, 22, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Li, J.; van Marion, D.M.S.; Zhang, D.; Brundel, B. Heat shock protein inducer gga*-59 reverses contractile and structural remodeling via restoration of the microtubule network in experimental atrial fibrillation. J. Mol. Cell Cardiol. 2019, 134, 86–97. [Google Scholar] [CrossRef] [Green Version]
- Brundel, B.J.; Shiroshita-Takeshita, A.; Qi, X.; Yeh, Y.H.; Chartier, D.; van Gelder, I.C.; Henning, R.H.; Kampinga, H.H.; Nattel, S. Induction of heat shock response protects the heart against atrial fibrillation. Circ. Res. 2006, 99, 1394–1402. [Google Scholar] [CrossRef] [Green Version]
- Sakabe, M.; Shiroshita-Takeshita, A.; Maguy, A.; Brundel, B.J.; Fujiki, A.; Inoue, H.; Nattel, S. Effects of a heat shock protein inducer on the atrial fibrillation substrate caused by acute atrial ischaemia. Cardiovasc. Res. 2008, 78, 63–70. [Google Scholar] [CrossRef]
- Gong, J.; Jing, L. Glutamine induces heat shock protein 70 expression via o-glcnac modification and subsequent increased expression and transcriptional activity of heat shock factor-1. Minerva Anestesiol. 2011, 77, 488–495. [Google Scholar]
- Hamiel, C.R.; Pinto, S.; Hau, A.; Wischmeyer, P.E. Glutamine enhances heat shock protein 70 expression via increased hexosamine biosynthetic pathway activity. Am. J. Physiol. Cell Physiol. 2009, 297, C1509–C1519. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, Y.; Sawa, Y.; Fukuyama, N.; Nakazawa, H.; Matsuda, H. Preoperative glutamine administration induces heat-shock protein 70 expression and attenuates cardiopulmonary bypass-induced inflammatory response by regulating nitric oxide synthase activity. Circulation 2002, 106, 2601–2607. [Google Scholar] [CrossRef]
- Cruzat, V.; Macedo Rogero, M.; Noel Keane, K.; Curi, R.; Newsholme, P. Glutamine: Metabolism and immune function, supplementation and clinical translation. Nutrients 2018, 10, 1564. [Google Scholar] [CrossRef] [Green Version]
- Ropeleski, M.J.; Riehm, J.; Baer, K.A.; Musch, M.W.; Chang, E.B. Anti-apoptotic effects of l-glutamine-mediated transcriptional modulation of the heat shock protein 72 during heat shock. Gastroenterology 2005, 129, 170–184. [Google Scholar] [CrossRef]
- Wu, C.; Kato, T.S.; Ji, R.; Zizola, C.; Brunjes, D.L.; Deng, Y.; Akashi, H.; Armstrong, H.F.; Kennel, P.J.; Thomas, T.; et al. Supplementation of l-alanyl-l-glutamine and fish oil improves body composition and quality of life in patients with chronic heart failure. Circ. Heart Fail. 2015, 8, 1077–1087. [Google Scholar] [CrossRef] [Green Version]
- Shahzad, K.; Chokshi, A.; Schulze, P.C. Supplementation of glutamine and omega-3 polyunsaturated fatty acids as a novel therapeutic intervention targeting metabolic dysfunction and exercise intolerance in patients with heart failure. Curr. Clin. Pharmacol. 2011, 6, 288–294. [Google Scholar] [CrossRef]
- Ziegler, T.R.; Ogden, L.G.; Singleton, K.D.; Luo, M.; Fernandez-Estivariz, C.; Griffith, D.P.; Galloway, J.R.; Wischmeyer, P.E. Parenteral glutamine increases serum heat shock protein 70 in critically ill patients. Intensive Care Med. 2005, 31, 1079–1086. [Google Scholar] [CrossRef]
- Morrison, A.L.; Dinges, M.; Singleton, K.D.; Odoms, K.; Wong, H.R.; Wischmeyer, P.E. Glutamine’s protection against cellular injury is dependent on heat shock factor-1. Am. J. Physiol. Cell Physiol. 2006, 290, C1625–C1632. [Google Scholar] [CrossRef]
- Luo, L.L.; Li, Y.F.; Shan, H.M.; Wang, L.P.; Yuan, F.; Ma, Y.Y.; Li, W.L.; He, T.T.; Wang, Y.Y.; Qu, M.J.; et al. L-glutamine protects mouse brain from ischemic injury via up-regulating heat shock protein 70. CNS Neurosci. Ther. 2019, 25, 1030–1041. [Google Scholar] [CrossRef] [Green Version]
- Boelens, P.G.; van Leeuwen, P.A.; Dejong, C.H.; Deutz, N.E. Intestinal renal metabolism of l-citrulline and l-arginine following enteral or parenteral infusion of l-alanyl-l-[2,15n]glutamine or l-[2,15n]glutamine in mice. Am. J. Physiol. Gastrointest Liver Physiol. 2005, 289, G679–G685. [Google Scholar] [CrossRef]
- Shih, Y.M.; Shih, J.M.; Pai, M.H.; Hou, Y.C.; Yeh, C.L.; Yeh, S.L. Glutamine administration after sublethal lower limb ischemia reduces inflammatory reaction and offers organ protection in ischemia/reperfusion injury. JPEN J. Parenter Enteral Nutr. 2016, 40, 1122–1130. [Google Scholar] [CrossRef]
- Morris, C.R.; Suh, J.H.; Hagar, W.; Larkin, S.; Bland, D.A.; Steinberg, M.H.; Vichinsky, E.P.; Shigenaga, M.; Ames, B.; Kuypers, F.A.; et al. Erythrocyte glutamine depletion, altered redox environment, and pulmonary hypertension in sickle cell disease. Blood 2008, 111, 402–410. [Google Scholar] [CrossRef]
- Forstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837, 837a–837d. [Google Scholar] [CrossRef] [Green Version]
- Molenaars, M.; Janssens, G.E.; Williams, E.G.; Jongejan, A.; Lan, J.; Rabot, S.; Joly, F.; Moerland, P.D.; Schomakers, B.V.; Lezzerini, M.; et al. A conserved mito-cytosolic translational balance links two longevity pathways. Cell Metab. 2020, 31, 549–563.e7. [Google Scholar] [CrossRef]
- Lanters, E.A.; van Marion, D.M.; Steen, H.; de Groot, N.M.; Brundel, B.J. The future of atrial fibrillation therapy: Intervention on heat shock proteins influencing electropathology is the next in line. Neth. Heart J. 2015, 23, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Brundel, B.J.; Ke, L.; Dijkhuis, A.J.; Qi, X.; Shiroshita-Takeshita, A.; Nattel, S.; Henning, R.H.; Kampinga, H.H. Heat shock proteins as molecular targets for intervention in atrial fibrillation. Cardiovasc Res. 2008, 78, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Batulan, Z.; Pulakazhi Venu, V.K.; Li, Y.; Koumbadinga, G.; Alvarez-Olmedo, D.G.; Shi, C.; O’Brien, E.R. Extracellular release and signaling by heat shock protein 27: Role in modifying vascular inflammation. Front. Immunol. 2016, 7, 285. [Google Scholar] [CrossRef] [Green Version]
- Leite, J.S.; Raizel, R.; Hypolito, T.M.; Rosa, T.D.; Cruzat, V.F.; Tirapegui, J. L-glutamine and l-alanine supplementation increase glutamine-glutathione axis and muscle hsp-27 in rats trained using a progressive high-intensity resistance exercise. Appl. Physiol. Nutr. Metab. 2016, 41, 842–849. [Google Scholar] [CrossRef]
- Moura, C.S.; Lollo, P.C.B.; Morato, P.N.; Risso, E.M.; Amaya-Farfan, J. Modulatory effects of arginine, glutamine and branched-chain amino acids on heat shock proteins, immunity and antioxidant response in exercised rats. Food Funct. 2017, 8, 3228–3238. [Google Scholar] [CrossRef]
- Andreasen, A.S.; Pedersen-Skovsgaard, T.; Mortensen, O.H.; van Hall, G.; Moseley, P.L.; Pedersen, B.K. The effect of glutamine infusion on the inflammatory response and hsp70 during human experimental endotoxaemia. Crit. Care 2009, 13, R7. [Google Scholar] [CrossRef] [Green Version]
- Jordan, I.; Balaguer, M.; Esteban, M.E.; Cambra, F.J.; Felipe, A.; Hernandez, L.; Alsina, L.; Molero, M.; Villaronga, M.; Esteban, E. Glutamine effects on heat shock protein 70 and interleukines 6 and 10: Randomized trial of glutamine supplementation versus standard parenteral nutrition in critically ill children. Clin. Nutr. 2016, 35, 34–40. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of patients | 21 |
| Male | 16 (76.2) |
| Age (years) | 58.7 ± 10.5 |
| BMI | 26.7 (22.0–36.3) |
| Type of AF: | |
| Paroxysmal | 13 (61.9) |
| Persistent | 6 (28.6) |
| Longstanding persistent | 2 (9.5) |
| Time since AF diagnosis (years) | 1.8 (0.2–19.5) |
| Hypertension | 6 (28.6) |
| Dyslipidemia | 2 (9.5) |
| Left ventricular function: | |
| Normal | 18 (85.7) |
| Mild impairment | 3 (14.3) |
| Use of anti-arrhythmic drugs (at baseline): | |
| Class I | 8 (38.1) |
| Class II | 6 (28.6) |
| Class III | 7 (33.3) |
| Class IV | 3 (14.3) |
| Class V | 1 (4.8) |
| Follow-up duration: | |
| Completed 3-month follow-up | 21 (100) |
| Completed 6-month follow-up | 20 (95.2) |
| Glutamine compliance: | |
| Baseline to 3-month follow-up (n = 20) | 0.99 (0.77–1.00) |
| 3-month to 6-month follow-up (n = 19) | 0.97 (0.61–1.00) |
| Baseline to 6-month follow-up (n = 20) | 0.97 (0.69–1.00) |
| Reduced kidney function (n = 17) | 1 (5.9) |
| Reduced liver function (n = 17) | 0 (0) |
| Reduced thyroid function (n = 17) | 0 (0) |
| Study ID | AF Type | HSP27-B | HSP27-3M | HSP27-6M | HSP70-B | HSP70-3M | HSP70-6M |
|---|---|---|---|---|---|---|---|
| 1 | L-PER | 1315.63 | 489.79 | 821.92 | 751.86 | 230.31 | 245.39 |
| 2 | L-PER | 455.71 | 312.78 | 571.25 | 31.50 | 110.58 | 118.29 |
| 3 | PAR | 869.73 | 352.14 | 441.98 | 862.09 | 268.50 | 565.83 |
| 4 | PAR | 751.50 | 271.66 | 278.59 | 542.14 | 202.70 | 120.66 |
| 5 | PAR | 288.50 | 407.61 | 769.92 | 524.28 | 610.66 | 463.72 |
| 6 | PAR | 514.07 | 915.03 | 1150.48 | 258.42 | 353.65 | 304.15 |
| 7 | PAR | 250.97 | 516.21 | 501.01 | 352.67 | 555.83 | 500.92 |
| 8 | PAR | 463.89 | 319.85 | 1401.11 | NA | NA | NA |
| 9 | PAR | 1203.41 | 271.80 | 1078.15 | 1564.51 | 396.58 | 1027.39 |
| 10 | PAR | 485.48 | 356.38 | - | 211.71 | 752.50 | - |
| 11 | PER | 360.65 | 332.83 | 379.69 | 390.49 | 382.47 | 409.58 |
| 12 | PER | 706.81 | 528.21 | 3103.61 | 1149.16 | 705.89 | 1744.19 |
| 13 | PER | 423.98 | 380.69 | 421.96 | 365.78 | 574.18 | 878.51 |
| 14 | PER | 1305.84 | 364.30 | 698.66 | 555.57 | 312.52 | NA |
| 15 | PAR | 666.37 | 629.06 | 1044.07 | 1119.96 | 177.28 | 179.68 |
| 16 | PAR | 911.61 | 402.25 | 929.48 | 614.61 | 190.27 | 314.44 |
| 17 | PAR | 369.99 | 407.58 | 139.57 | 143.11 | 635.17 | 142.24 |
| 18 | PER | 550.98 | 409.01 | 758.89 | 735.83 | 328.16 | 1182.55 |
| 19 | PAR | 1102.87 | 394.93 | 483.66 | 880.77 | 304.32 | 135.65 |
| 20 | PER | 540.39 | 185.68 | 192.68 | 678.67 | 371.17 | 208.70 |
| 21 | PAR | 498.64 | 221.64 | 437.17 | 268.38 | NA | 225.46 |
| Median | - | 540.39 | 380.69 | 634.96 | 548.86 | 353.65 | 309.30 |
| Min-Max | - | 250.97–1315.63 | 185.68–915.03 | 139.57–3103.61 | 31.50–1564.51 | 110.58–752.50 | 118.29–1744.19 |
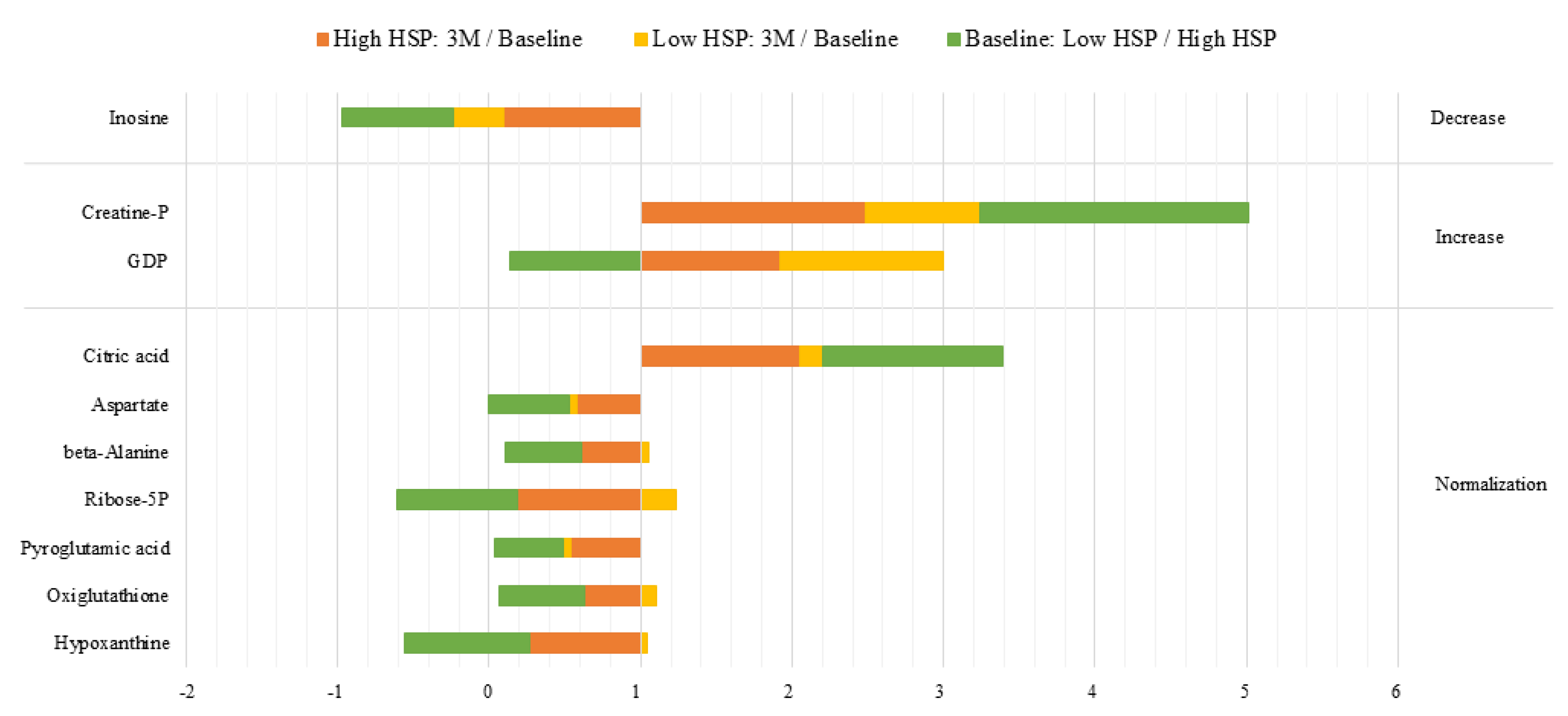
| Behavior | Ratios | Metabolites | Percentage | ||
|---|---|---|---|---|---|
| High HSP: 3M/Baseline | Baseline: Low HSP/High HSP | Low HSP: 3M/Baseline | |||
| Normalization | <0.667 | <0.667 | Stable | Ribose-5P, Aspartate, beta-Alanine, Hypoxanthine, Oxiglutathione, Pyroglutamic acid | 8.43% |
| >1.5 | >1.5 | Stable | Citric Acid | ||
| Increase | >1.5 | >1.5 | >1.5 | Creatine P | 2.4% |
| >1.5 | <0.667 | >1.5 | GDP | ||
| Decrease | <0.667 | <0.667 | <0.667 | Inosine | 1.2% |
| Independent | <0.667 | <0.667 | >1.5 | Sedoheptulose-7P, Fumarate, CMP, 3-phosphoglyceric acid | 31.32% |
| <0.667 | Stable | Stable | 3- and 4-hydroxybenzoic acid | ||
| >1.5 | Stable | Stable | Gluconate-6P | ||
| >1.5 | Stable | <0.667 | dAMP | ||
| Stable | Stable | >1.5 | Allantoin, Malate | ||
| Stable | Stable | <0.667 | Coenzyme A, Hippuric acid | ||
| Stable | <0.667 | >1.5 | Uracil, UDP-HexNac, 2-phosphoglyceric acid, Adenine, Glycerol-3P | ||
| Stable | <0.667 | Stable | Xanthine, Phosphorylethanolanine, Aminoadicpic acid, Glucose 6P, Glutathione, Ribose | ||
| Stable | >1.5 | Stable | Creatine, Guanosine | ||
| Stable | >1.5 | <0.667 | AMP, GMP, Mesaconic acid | ||
| Stable | Stable | Stable | Stable | 2-aminoisobutyric acid, 2-Dehydrogluconate, 2-Hydroxyglutarate, Alanine, Alpha, Ketoglutarate, Arginine, Asparagine, Citrulline, Creatinine, Cysteine, Cystine, FAICAR, Gluconate, Glucose, Glutamate, Glutamine, Glyceraldehyde-3P, Glycerate, Glycerol-2P, Hexose-P, Hydroxyphenyllactic acid, Isoleucine, Kynurenic acid, Kynurenine, Lactate, Leucine, Lysine, Malonic acid, Methionine, Ornithine, Orotic acid, Pantothenic acid, Phenylalanine, Phosphoenolpyruvate, Proline, Pyruvate, Serine, Succinate, Taurine, Threonine, Tryptophan, Tyrosine, UMP, Uric acid, Uridine, Valine | 55,4% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Starreveld, R.; Ramos, K.S.; Muskens, A.J.Q.M.; Brundel, B.J.J.M.; de Groot, N.M.S. Daily Supplementation of L-Glutamine in Atrial Fibrillation Patients: The Effect on Heat Shock Proteins and Metabolites. Cells 2020, 9, 1729. https://doi.org/10.3390/cells9071729
Starreveld R, Ramos KS, Muskens AJQM, Brundel BJJM, de Groot NMS. Daily Supplementation of L-Glutamine in Atrial Fibrillation Patients: The Effect on Heat Shock Proteins and Metabolites. Cells. 2020; 9(7):1729. https://doi.org/10.3390/cells9071729
Chicago/Turabian StyleStarreveld, Roeliene, Kennedy S. Ramos, Agnes J. Q. M. Muskens, Bianca J. J. M. Brundel, and Natasja M. S. de Groot. 2020. "Daily Supplementation of L-Glutamine in Atrial Fibrillation Patients: The Effect on Heat Shock Proteins and Metabolites" Cells 9, no. 7: 1729. https://doi.org/10.3390/cells9071729
APA StyleStarreveld, R., Ramos, K. S., Muskens, A. J. Q. M., Brundel, B. J. J. M., & de Groot, N. M. S. (2020). Daily Supplementation of L-Glutamine in Atrial Fibrillation Patients: The Effect on Heat Shock Proteins and Metabolites. Cells, 9(7), 1729. https://doi.org/10.3390/cells9071729