The Molecular Function and Clinical Role of Thyroid Stimulating Hormone Receptor in Cancer Cells

Abstract
:1. Introduction
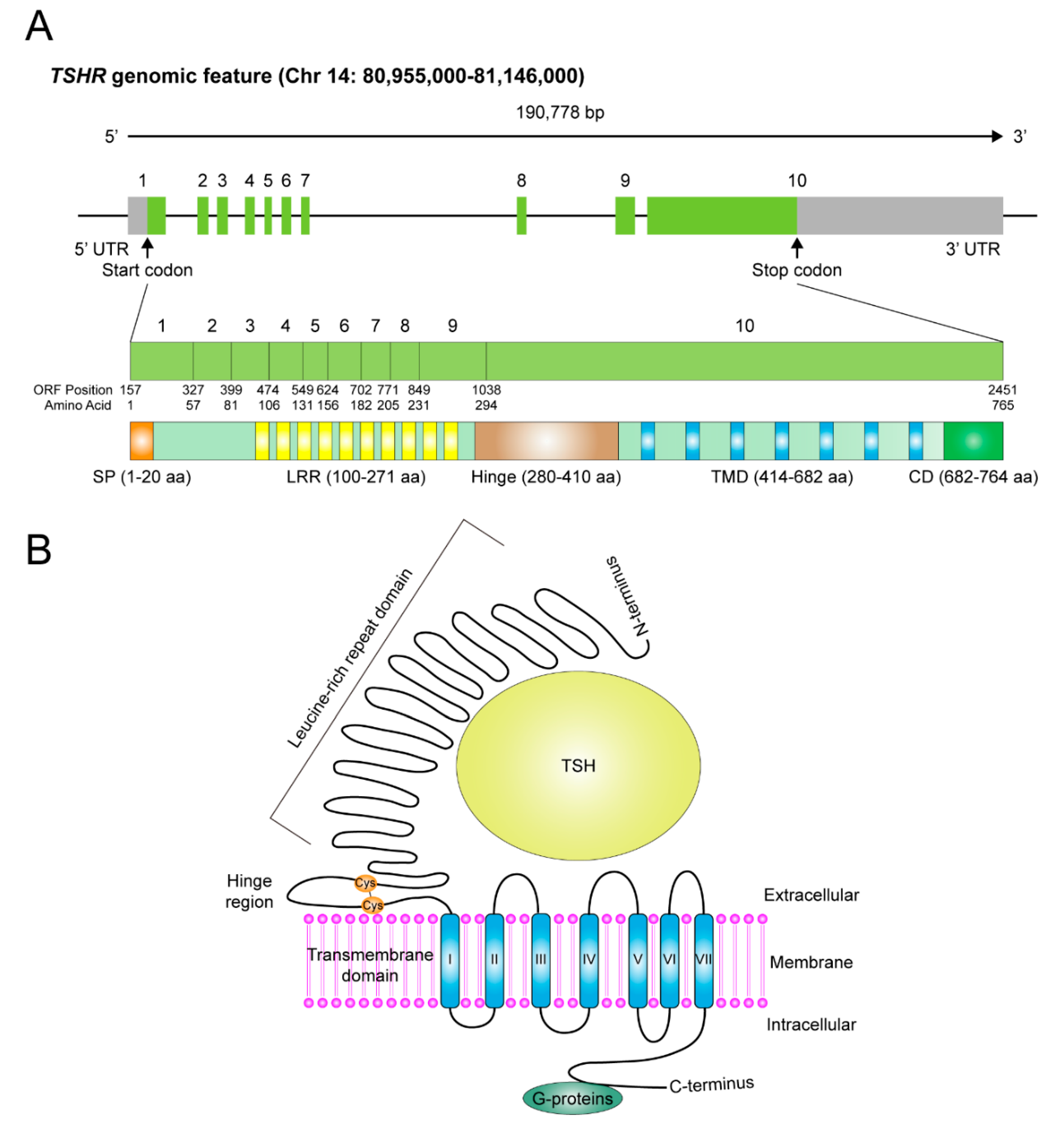
2. Genomic Feature, Protein Structure and Distribution of TSHR
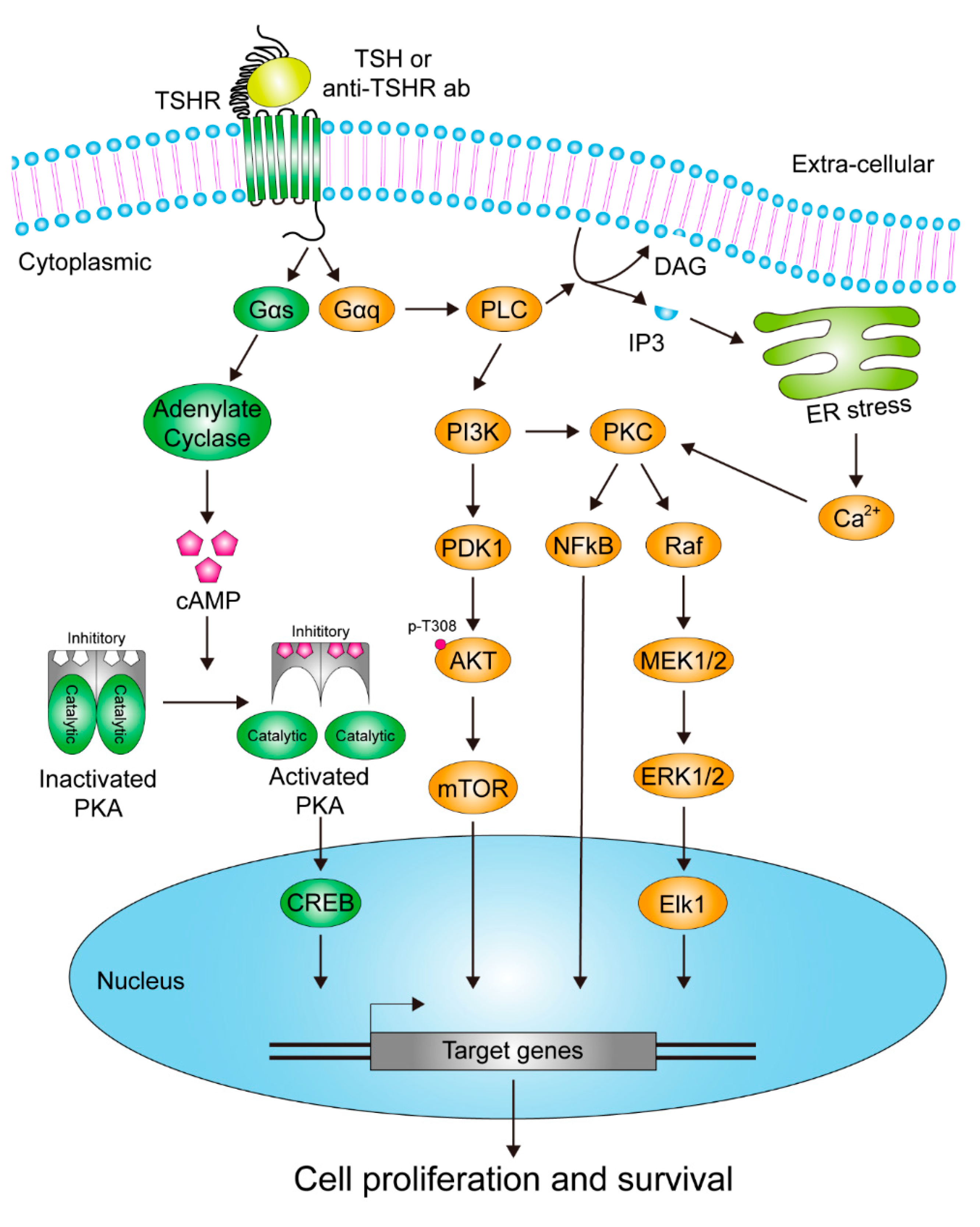
3. The Signaling Pathways Downstream of TSHR in Response to Stimulatory Signals
4. TSHR in Thyroid Cancer Cells
5. TSHR in Extra-Thyroid Cancer Cells
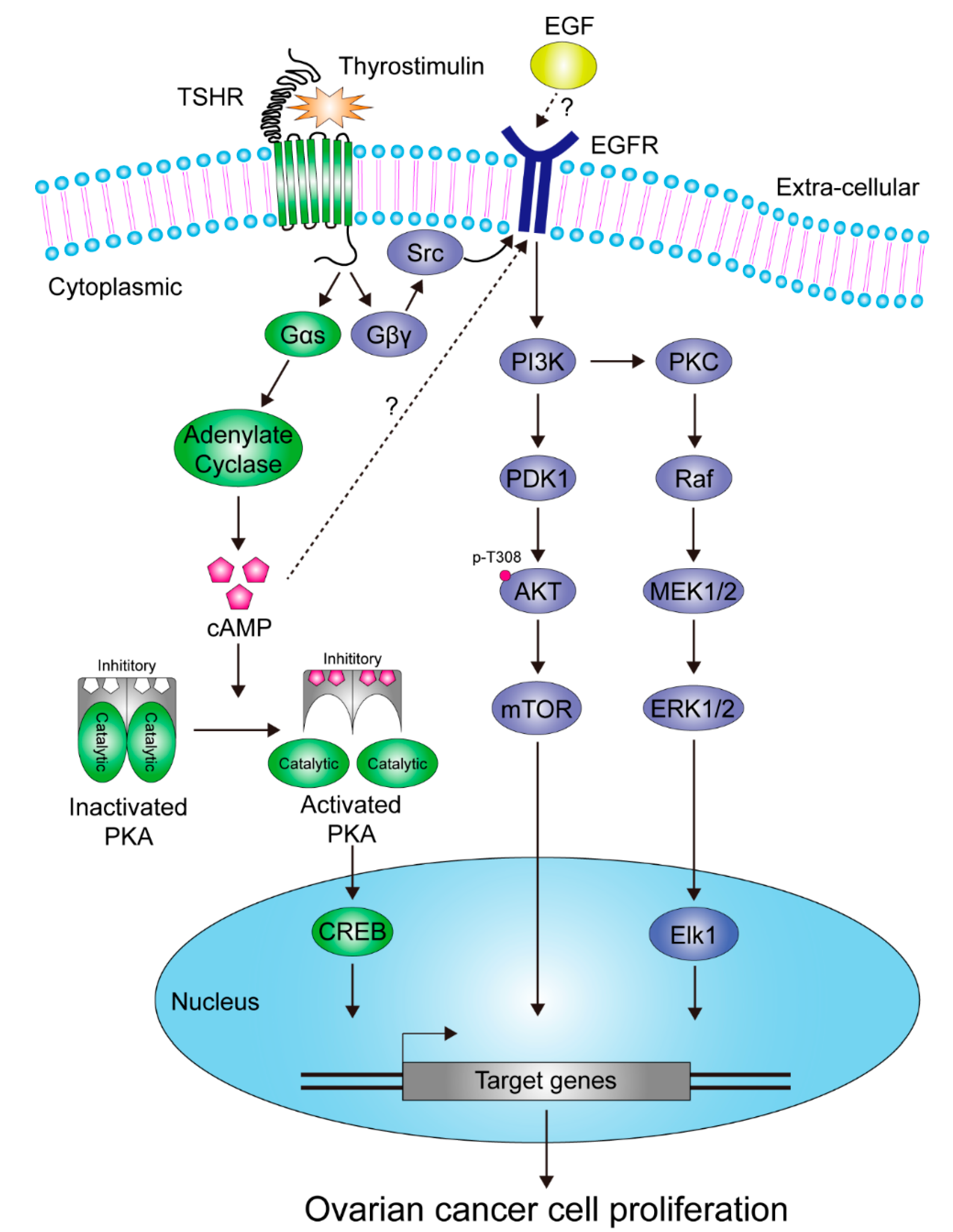
5.1. Ovarian Cancer
5.2. Liver Cancer
6. Clinical Usefulness of TSHR Related Pathways in Anticancer Therapies
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Neumann, S.; Raaka, B.M.; Gershengorn, M.C. Human TSH receptor ligands as pharmacological probes with potential clinical application. Expert Rev. Endocrinol. Metab. 2009, 4, 669–679. [Google Scholar] [CrossRef] [Green Version]
- Troppmann, B.; Kleinau, G.; Krause, G.; Gromoll, J. Structural and functional plasticity of the luteinizing hormone/choriogonadotrophin receptor. Hum. Reprod Update 2013, 19, 583–602. [Google Scholar] [CrossRef] [Green Version]
- Persani, L.; Gelmini, G.; Marelli, F.; Beck-Peccoz, P.; Bonomi, M. Syndromes of resistance to TSH. Ann. Endocrinol. 2011, 72, 60–63. [Google Scholar] [CrossRef]
- Schroeder, A.C.; Privalsky, M.L. Thyroid hormones, t3 and t4, in the brain. Front. Endocrinol. 2014, 5, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maenhaut, C.; Brabant, G.; Vassart, G.; Dumont, J.E. In vitro and in vivo regulation of thyrotropin receptor mRNA levels in dog and human thyroid cells. J. Biol. Chem. 1992, 267, 3000–3007. [Google Scholar]
- Bruno, R.; Ferretti, E.; Tosi, E.; Arturi, F.; Giannasio, P.; Mattei, T.; Scipioni, A.; Presta, I.; Morisi, R.; Gulino, A.; et al. Modulation of thyroid-specific gene expression in normal and nodular human thyroid tissues from adults: An in vivo effect of thyrotropin. J. Clin. Endocrinol. Metab. 2005, 90, 5692–5697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuppert, F.; Deiters, S.; Rambusch, E.; Sierralta, W.; Dralle, H.; Mühlen, A.V.Z. TSH-receptor expression and human thyroid disease: Relation to clinical, endocrine, and molecular thyroid parameters. Thyroid 1996, 6, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Haugen, B.R.; Alexander, E.K.; Bible, K.C.; Doherty, G.M.; Mandel, S.J.; Nikiforov, Y.E.; Pacini, F.; Randolph, G.W.; Sawka, A.M.; Schlumberger, M.; et al. 2015 American Thyroid Association Management Guidelines for Adult Patients with Thyroid Nodules and Differentiated Thyroid Cancer: The American Thyroid Association Guidelines Task Force on Thyroid Nodules and Differentiated Thyroid Cancer. Thyroid 2016, 26, 1–133. [Google Scholar] [CrossRef] [Green Version]
- Williams, G.R. Extrathyroidal expression of TSH receptor. Ann. Endocrinol 2011, 72, 68–73. [Google Scholar] [CrossRef]
- Pastan, I.; Roth, J.; Macchia, V. Binding of hormone to tissue: The first step in polypeptide hormone action. Proc. Natl. Acad. Sci. USA 1966, 56, 1802–1809. [Google Scholar] [CrossRef] [Green Version]
- Amir, S.M.; Carraway, T.F., Jr.; Kohn, L.D.; Winand, R.J. The Binding of Thyrotropin to Isolated Bovine Thyroid Plasma Membranes. J. Biol. Chem. 1973, 248, 4092–4100. [Google Scholar]
- Parmentier, M.; Libert, F.; Maenhaut, C.; Lefort, A.; Gerard, C.; Perret, J.; Sande, J.V.; Dumont, J.; Vassart, G. Molecular cloning of the thyrotropin receptor. Science 1989, 246, 1620–1622. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, Y.; Kaufman, K.D.; Seto, P.; Rapoport, B. Molecular cloning, sequence and functional expression of the cDNA for the human thyrotropin receptor. Biochem. Biophys. Res. Commun. 1989, 165, 1184–1190. [Google Scholar] [CrossRef]
- Misrahi, M.; Loosfelt, H.; Atger, M.; Sar, S.; Guiochon-Mantel, A.; Milgrom, E. Cloning, sequencing and expression of human TSH receptor. Biochem. Biophys. Res. Commun. 1990, 166, 394–403. [Google Scholar] [CrossRef]
- Akamizu, T. Antithyrotropin Receptor Antibody: An Update. Thyroid 2001, 11, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Bahn, R.S. Autoimmunity and Graves’ disease. Clin. Pharm. 2012, 91, 577–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahn, R.S.; Dutton, C.M.; Natt, N.; Joba, W.; Spitzweg, C.; Heufelder, A.E. Thyrotropin receptor expression in Graves’ orbital adipose/connective tissues: Potential autoantigen in Graves’ ophthalmopathy. J. Clin. Endocrinol. Metab. 1998, 83, 998–1002. [Google Scholar] [CrossRef] [PubMed]
- Davies, T.F. Thyrotropin receptor-associated diseases: From adenomata to Graves disease. J. Clin. Investig. 2005, 115, 1972–1983. [Google Scholar] [CrossRef] [Green Version]
- Kohn, L.D.; Harii, N. Thyrotropin receptor autoantibodies (TSHRAbs): Epitopes, origins and clinical significance. Autoimmunity 2003, 36, 331–337. [Google Scholar] [CrossRef]
- Ludgate, M. Animal models of Graves’ disease. Eur. J. Endocrinol. 2000, 142, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalek, K.; Morshed, S.A.; Latif, R.; Davies, T.F. TSH receptor autoantibodies. Autoimmun. Rev. 2009, 9, 113–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapoport, B.; Chazenbalk, G.D.; Jaume, J.C.; Mclachlan, S.M. The thyrotropin (TSH) receptor: Interaction with TSH and autoantibodies. Endocr. Rev. 1998, 19, 673–716. [Google Scholar]
- Rapoport, B.; McLachlan, S.M. The Thyrotropin Receptor in Graves’ Disease. Thyroid 2007, 17, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Diana, T.; Olivo, P.D.; Kahaly, G.J. Thyrotropin Receptor Blocking Antibodies. Horm. Metab. Res. 2018, 50, 853–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, T.F.; Yin, X.; Latif, R. The genetics of the thyroid stimulating hormone receptor: History and relevance. Thyroid 2010, 20, 727–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, D.; Chazenbalk, G.D.; Nagayama, Y.; Wadsworth, H.L.; Seto, P.; Rapoport, B. A new structural model for the thyrotropin (TSH) receptor, as determined by covalent cross-linking of TSH to the recombinant receptor in intact cells: Evidence for a single polypeptide chain. Mol. Endocrinol. 1991, 5, 1607–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, T.; Ikeda, M.; Ohmori, M.; Anzai, E.; Haraguchi, K.; Onaya, T. Single subunit structure of the human thyrotropin receptor. Biochem. Biophys. Res. Commun. 1992, 187, 887–893. [Google Scholar] [CrossRef]
- Chazenbalk, G.D.; Kakinuma, A.; Jaume, J.C.; Mclachlan, S.M.; Rapoport, B. Evidence for negative cooperativity among human thyrotropin receptors overexpressed in mammalian cells. Endocrinology 1996, 137, 4586–4591. [Google Scholar] [CrossRef]
- Chen, C.-R.; Chazenbalk, G.D.; Wawrowsky, K.A.; McLachlan, S.M.; Rapoport, B. Evidence that human thyroid cells express uncleaved, single-chain thyrotropin receptors on their surface. Endocrinology 2006, 147, 3107–3113. [Google Scholar] [CrossRef] [Green Version]
- Buckland, P.R.; Howells, R.D.; Rickards, C.R.; Smith, B.R. Affinity-labelling of the thyrotropin receptor. Characterization of the photoactive ligand. Biochem. J. 1985, 225, 753–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajita, Y.; Rickards, C.R.; Buckland, P.R.; Howells, R.D.; Smith, B.R. Analysis of thyrotropin receptors by photoaffinity labelling. Orientation of receptor subunits in the cell membrane. Biochem. J. 1985, 227, 413–420. [Google Scholar] [CrossRef] [Green Version]
- Buckland, P.R.; Strickland, T.W.; Pierce, J.G.; Smith, B.R. TSH crosslinks to the TSH receptor through the beta subunit. Endocrinology 1985, 116, 2122–2124. [Google Scholar] [CrossRef]
- Loosfelt, H.; Pichon, C.; Jolivet, A.; Misrahi, M.; Caillou, B.; Jamous, M.; Vannier, B.; Milgrom, E. Two-subunit structure of the human thyrotropin receptor. Proc. Natl. Acad. Sci. USA 1992, 89, 3765–3769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapoport, B.; McLachlan, S.M. TSH Receptor Cleavage into Subunits and Shedding of the A-Subunit; A Molecular and Clinical Perspective. Endocr. Rev. 2016, 37, 114–134. [Google Scholar] [CrossRef]
- Kleinau, G.; Worth, C.L.; Kreuchwig, A.; Biebermann, H.; Marcinkowski, P.; Scheerer, P.; Krause, G. Structural-Functional Features of the Thyrotropin Receptor: A Class A G-Protein-Coupled Receptor at Work. Front. Endocrinol. 2017, 8, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kursawe, R.; Paschke, R. Modulation of TSHR signaling by posttranslational modifications. Trends Endocrinol. Metab. 2007, 18, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Zabczynska, M.; Kozlowska, K.; Pochec, E. Glycosylation in the Thyroid Gland: Vital Aspects of Glycoprotein Function in Thyrocyte Physiology and Thyroid Disorders. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [Green Version]
- Vastrad, B.; Vastrad, C.; Godavarthi, A.; Chandrashekar, R. Molecular mechanisms underlying gliomas and glioblastoma pathogenesis revealed by bioinformatics analysis of microarray data. Med. Oncol. 2017, 34, 182. [Google Scholar] [CrossRef]
- Ellerhorst, J.A.; Sendi-Naderi, A.; Johnson, M.K.; Cooke, C.P.; Dang, S.M.; Diwan, A.H. Human melanoma cells express functional receptors for thyroid-stimulating hormone. Endocr. Relat. Cancer 2006, 13, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Lee, S.; Lui, N.; Choi, H.; Mulvihill, M.; Fang, L.T.; Kang, H.C.; Kwon, Y.W.; Jablons, D.; Kim, I.J. A somatic TSHR mutation in a patient with lung adenocarcinoma with bronchioloalveolar carcinoma, coronary artery disease and severe chronic obstructive pulmonary disease. Oncol. Rep. 2012, 28, 1225–1230. [Google Scholar] [CrossRef] [PubMed]
- Govindaraj, V.; Yaduvanshi, N.S.; Krishnamachar, H.; Rao, A.J. Expression of thyroid-stimulating hormone receptor, octamer-binding transcription factor 4, and intracisternal A particle-promoted polypeptide in human breast cancer tissues. Horm. Mol. Biol. Clin. Investig 2012, 9, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Gyftaki, R.; Liacos, C.; Politi, E.; Liontos, M.; Saltiki, K.; Papageorgiou, T.; Thomakos, N.; Haidopoulos, D.; Rodolakis, A.; Alevizaki, M.; et al. Differential transcriptional and protein expression of thyroid-stimulating hormone receptor in ovarian carcinomas. Int. J. Gynecol. Cancer 2014, 24, 851–856. [Google Scholar] [PubMed]
- Shih, Y.L.; Huang, Y.H.; Lin, K.H.; Chu, Y.D.; Yeh, C.T. Identification of Functional Thyroid Stimulating Hormone Receptor and TSHR Gene Mutations in Hepatocellular Carcinoma. Anticancer Res. 2018, 38, 2793–2802. [Google Scholar] [PubMed]
- Janson, A.; Rawet, H.; Perbeck, L.; Marcus, C. Presence of thyrotropin receptor in infant adipocytes. Pediatr. Res. 1998, 43, 555–558. [Google Scholar] [CrossRef] [Green Version]
- Bell, A.; Gagnon, A.; Grunder, L.; Parikh, S.J.; Smith, T.J.; Sorisky, A. Functional TSH receptor in human abdominal preadipocytes and orbital fibroblasts. Am. J. Physiol. Cell Physiol. 2000, 279, C335–C340. [Google Scholar] [CrossRef]
- Murakami, M.; Kamiya, Y.; Morimura, T.; Araki, O.; Imamura, M.; Ogiwara, T.; Mizuma, H.; Mori, M. Thyrotropin receptors in brown adipose tissue: Thyrotropin stimulates type II iodothyronine deiodinase and uncoupling protein-1 in brown adipocytes. Endocrinology 2001, 142, 1195–1201. [Google Scholar] [CrossRef]
- Dutton, C.M.; Joba, W.; Spitzweg, C.; Heufelder, A.E.; Bahn, R.S. Thyrotropin receptor expression in adrenal, kidney, and thymus. Thyroid 1997, 7, 879–884. [Google Scholar] [CrossRef]
- Aghajanova, L.; Stavreus-Evers, A.; Lindeberg, M.; Landgren, B.M.; Sparre, L.S.; Hovatta, O. Thyroid-stimulating hormone receptor and thyroid hormone receptors are involved in human endometrial physiology. Fertil. Steril. 2011, 95, 230–237. [Google Scholar] [CrossRef]
- Balzan, S.; Nicolini, G.; Forini, F.; Boni, G.; Del Carratore, R.; Nicolini, A.; Carpi, A.; Iervasi, G. Presence of a functional TSH receptor on human erythrocytes. Biomed. Pharm. 2007, 61, 463–467. [Google Scholar] [CrossRef]
- Balzan, S.; Carratore, R.D.; Nicolini, G.; Forini, F.; Lubrano, V.; Simili, M.; Benedetti, P.A.; Iervasi, G. TSH induces co-localization of TSH receptor and Na/K-ATPase in human erythrocytes. Cell Biochem. Funct. 2009, 27, 259–263. [Google Scholar] [CrossRef]
- Zhang, W.; Tian, L.M.; Han, Y.; Ma, H.Y.; Wang, L.C.; Guo, J.; Gao, L.; Zhao, J.J. Presence of thyrotropin receptor in hepatocytes: Not a case of illegitimate transcription. J. Cell. Mol. Med. 2009, 13, 4636–4642. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, L.; Zhou, L.; Song, Y.; Ma, S.; Yu, C.; Zhao, J.; Xu, C.; Gao, L. Thyroid stimulating hormone increases hepatic gluconeogenesis via CRTC2. Mol. Cell Endocrinol. 2017, 446, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Coutelier, J.-P.; Kehrl, J.H.; Bellur, S.S.; Kohn, L.D.; Notkins, A.L.; Prabhakar, B.S. Binding and functional effects of thyroid stimulating hormone on human immune cells. J. Clin. Immunol. 1990, 10, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Prummel, M.F.; Brokken, L.J.S.; Meduri, G.; Misrahi, M.; Bakker, O.; Wiersinga, W.M. Expression of the thyroid-stimulating hormone receptor in the folliculo-stellate cells of the human anterior pituitary. J. Clin. Endocrinol. Metab. 2000, 85, 4347–4353. [Google Scholar] [CrossRef] [PubMed]
- Drvota, V.; Janson, A.; Norman, C.; Sylven, C.; Haggblad, J.; Bronnegard, M.; Marcus, C. Evidence for the presence of functional thyrotropin receptor in cardiac muscle. Biochem. Biophys. Res. Commun. 1995, 211, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Bodo, E.; Kromminga, A.; Biro, T.; Borbiro, I.; Gaspar, E.; Zmijewski, M.A.; van Beek, N.; Langbein, L.; Slominski, A.T.; Paus, R. Human female hair follicles are a direct, nonclassical target for thyroid-stimulating hormone. J. Invest. Derm. 2009, 129, 1126–1139. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Ni, J.; Guo, T.; Liu, J.; Dang, Y.; Guo, Q.; Zhang, L. TSH stimulates the proliferation of vascular smooth muscle cells. Endocrine 2014, 46, 651–658. [Google Scholar] [CrossRef]
- Aghajanova, L.; Lindeberg, M.; Carlsson, I.B.; Stavreus-Evers, A.; Zhang, P.; Scott, J.E.; Hovatta, O.; Skjöldebrand-Sparre, L. Receptors for thyroid-stimulating hormone and thyroid hormones in human ovarian tissue. Reprod. Biomed. Online 2009, 18, 337–347. [Google Scholar] [CrossRef]
- Sun, S.C.; Hsu, P.J.; Wu, F.J.; Li, S.H.; Lu, C.H.; Luo, C.W. Thyrostimulin, but not thyroid-stimulating hormone (TSH), acts as a paracrine regulator to activate the TSH receptor in mammalian ovary. J. Biol. Chem. 2010, 285, 3758–3765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seagle, B.L.; Eng, K.H.; Yeh, J.Y.; Dandapani, M.; Schiller, E.; Samuelson, R.; Odunsi, K.; Shahabi, S. Discovery of candidate tumor biomarkers for treatment with intraperitoneal chemotherapy for ovarian cancer. Sci. Rep. 2016, 6, 21591. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.L.; Li, Z.; Lin, T.Y.; Wang, S.W.; Wu, F.J.; Luo, C.W. Thyrostimulin-TSHR signaling promotes the proliferation of NIH:OVCAR-3 ovarian cancer cells via trans-regulation of the EGFR pathway. Sci. Rep. 2016, 6, 27471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassart, G.; Pardo, L.; Costagliola, S. A molecular dissection of the glycoprotein hormone receptors. Trends Biochem. Sci. 2004, 29, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Tuncel, M. Thyroid Stimulating Hormone Receptor. Mol. Imaging. Radionucl. 2017, 26, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Kleinau, G.; Neumann, S.; Grüters, A.; Krude, H.; Biebermann, H. Novel Insights on Thyroid-Stimulating Hormone Receptor Signal Transduction. Endocr. Rev. 2013, 34, 691–724. [Google Scholar] [CrossRef] [Green Version]
- Duprez, L.; Parma, J.; Sande, J.V.; Allgeier, A.; Leclère, J.; Schvartz, C.; Delisle, M.-J.; Decoulx, M.; Orgiazzi, J.; Dumont, J.; et al. Germline mutations in the thyrotropin receptor gene cause non-autoimmune autosomal dominant hyperthyroidism. Nat. Genet. 1994, 7, 396–401. [Google Scholar] [CrossRef]
- Hsu, S.Y.; Nakabayashi, K.; Bhalla, A. Evolution of Glycoprotein Hormone Subunit Genes in Bilateral Metazoa: Identification of Two Novel Human Glycoprotein Hormone Subunit Family Genes, GPA2 and GPB5. Mol. Endocrinol. 2002, 16, 1538–1551. [Google Scholar] [CrossRef]
- Postiglione, M.P.; Parlato, R.; Rodriguez-Mallon, A.; Rosica, A.; Mithbaokar, P.; Maresca, M.; Marians, R.C.; Davies, T.F.; Zannini, M.S.; De Felice, M.; et al. Role of the thyroid-stimulating hormone receptor signaling in development and differentiation of the thyroid gland. Proc. Natl. Acad. Sci. USA 2002, 99, 15462–15467. [Google Scholar] [CrossRef] [Green Version]
- Vassart, G.; Dumont, J.E. The Thyrotropin Receptor and the Regulation of Thyrocyte Function and Growth. Endocr. Rev. 1992, 13, 596–611. [Google Scholar]
- Allgeier, A.; Offermanns, S.; Sande, J.V.; Spicher, K.; Schultz, G.; Dumont, J. The human thyrotropin receptor activates G-proteins Gs and Gq/11. J. Biol. Chem. 1994, 269, 13733–13735. [Google Scholar]
- Laugwitz, K.-L.; Allgeier, A.; Offermanns, S.; Spicher, K.; Sande, J.V.; Dumont, J.E.; Schultz, G. The human thyrotropin receptor: A heptahelical receptor capable of stimulating members of all four G protein families. Proc. Natl. Acad. Sci. USA 1996, 93, 116–120. [Google Scholar] [CrossRef] [Green Version]
- Sande, J.V.; Raspé, E.; Perret, J.; Lejeune, C.; Maenhaut, C.; Vassart, G.; Dumont, J.E. Thyrotropin activates both the cyclic AMP and the PIP2 cascades in CHO cells expressing the human cDNA of TSH receptor. Mol. Cell Endocrinol. 1990, 74, R1–R6. [Google Scholar] [CrossRef]
- Buch, T.R.; Biebermann, H.; Kalwa, H.; Pinkenburg, O.; Hager, D.; Barth, H.; Aktories, K.; Breit, A.; Gudermann, T. G13-dependent activation of MAPK by thyrotropin. J. Biol. Chem. 2008, 283, 20330–20341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krause, K.; Boisnard, A.; Ihling, C.; Ludgate, M.; Eszlinger, M.; Krohn, K.; Sinz, A.; Fuhrer, D. Comparative proteomic analysis to dissect differences in signal transduction in activating TSH receptor mutations in the thyroid. Int. J. Biochem. Cell Biol. 2012, 44, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Kero, J.; Ahmed, K.; Wettschureck, N.; Tunaru, S.; Wintermantel, T.; Greiner, E.; Schutz, G.; Offermanns, S. Thyrocyte-specific Gq/G11 deficiency impairs thyroid function and prevents goiter development. J. Clin. Invest. 2007, 117, 2399–2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledent, C.; Parmentier, M.; Maenhaut, C.; Taton, M.; Pirson, I.; Lamy, F.; Roger, P.; Dumont, J.E. The TSH cyclic AMP cascade in the control of thyroid cell proliferation: The story of a concept. Thyroidology 1991, 3, 97–101. [Google Scholar] [PubMed]
- Verrier, B.; Fayet, G.; Lissitzky, S. Thyrotropin-binding properties of isolated thyroid cells and their purified plasma membranes. Relation of thyrotropin-specific binding to adenylate-cyclase activation. Eur. J. Biochem. 1974, 42, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Winkler, F.; Kleinau, G.; Tarnow, P.; Rediger, A.; Grohmann, L.; Gaetjens, I.; Krause, G.; L’Allemand, D.; Gruters, A.; Krude, H.; et al. A new phenotype of nongoitrous and nonautoimmune hyperthyroidism caused by a heterozygous thyrotropin receptor mutation in transmembrane helix 6. J. Clin. Endocr Metab. 2010, 95, 3605–3610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, G.K.; Weinstein, S.P.; Graves, P.N.; Davies, T.F. The positive regulation of human thyrotropin (TSH) receptor messenger ribonucleic acid by recombinant human TSH is at the intranuclear level. Endocrinology 1992, 130, 2858–2864. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Okajima, F.; Ban, T.; Hidaka, A.; Shenker, A.; Kohn, L.D. Substitutions of different regions of the third cytoplasmic loop of the thyrotropin (TSH) receptor have selective effects on constitutive, TSH-, and TSH receptor autoantibody-stimulated phosphoinositide and 3′,5′-cyclic adenosine monophosphate signal generation. Mol. Endocrinol. 1993, 7, 1009–1020. [Google Scholar]
- Kosugi, S.; Kohn, L.D.; Akamizu, T.; Mori, T. The middle portion in the second cytoplasmic loop of the thyrotropin receptor plays a crucial role in adenylate cyclase activation. Mol. Endocrinol 1994, 8, 498–509. [Google Scholar]
- Kosugi, S.; Shenker, A.; Mori, T. Constitutive activation of cyclic AMP but not phosphatidylinositol signaling caused by four mutations in the 6th transmembrane helix of the human thyrotropin receptor. Febs Lett. 1994, 356, 291–294. [Google Scholar] [CrossRef] [Green Version]
- Morshed, S.A.; Latif, R.; Davies, T.F. Characterization of thyrotropin receptor antibody-induced signaling cascades. Endocrinology 2009, 150, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Gilman, A.G.; Rall, T.W. The role of adenosine 3′,5′-phosphate in mediating effects of thyroid-stimulating hormone on carbohydrate metabolism of bovine thyroid slices. J. Biol. Chem. 1968, 243, 5872–5881. [Google Scholar] [PubMed]
- Rapoport, B. Dog thyroid cells in monolayer tissue culture: Adenosine 3′, 5′-cyclic monophosphate response to thyrotropic hormone. Endocrinology 1976, 98, 1189–1197. [Google Scholar] [CrossRef]
- Allgeier, A.; Laugwitz, K.-L.; Sande, J.V.; Schultz, G.n.; Dumont, J.E. Multiple G-protein coupling of the dog thyrotropin receptor. Mol. Cell. Endocrinol. 1997, 127, 81–90. [Google Scholar] [CrossRef]
- Dremier, S.; Pohl, V.; Poteet-Smith, C.; Roger, P.P.; Corbin, J.; Doskeland, S.O.; Dumont, J.E.; Maenhaut, C. Activation of cyclic AMP-dependent kinase is required but may not be sufficient to mimic cyclic AMP-dependent DNA synthesis and thyroglobulin expression in dog thyroid. Mol. Cell Biol. 1997, 17, 6717–6726. [Google Scholar] [CrossRef] [Green Version]
- Kosugi, S.; Mori, T. The first cytoplasmic loop of the thyrotropin receptor is important for phosphoinositide signaling but not for agonist-induced adenylate cyclase activation. Febs. Lett. 1994, 341, 162–166. [Google Scholar] [CrossRef] [Green Version]
- Kosugi, S.; Mori, T. The intracellular region adjacent to plasma membrane (residues 684-692) of the thyrotropin receptor is important for phosphoinositide signaling but not for agonist-induced adenylate cyclase activation. Biochem. Biophys. Res. Commun. 1994, 199, 1497–1503. [Google Scholar] [CrossRef]
- Laurent, E.; Mockel, J.; Sande, J.V.; Graff, I.; Dumont, J.E. Dual activation by thyrotropin of the phospholipase C and cyclic AMP cascades in human thyroid. Mol. Cell Endocrinol. 1987, 52, 273–278. [Google Scholar] [CrossRef]
- Karponis, D.; Ananth, S. The role of thyrostimulin and its potential clinical significance. Endocr. Regul. 2017, 51, 117–128. [Google Scholar] [CrossRef]
- Nakabayashi, K.; Matsumi, H.; Bhalla, A.; Bae, J.; Mosselman, S.; Hsu, S.Y.; Hsueh, A.J.W. Thyrostimulin, a heterodimer of two new human glycoprotein hormone subunits, activates the thyroid-stimulating hormone receptor. J. Clin. Investig. 2002, 109, 1445–1452. [Google Scholar] [CrossRef] [PubMed]
- Okajima, Y.; Nagasaki, H.; Suzuki, C.; Suga, H.; Ozaki, N.; Arima, H.; Hamada, Y.; Civelli, O.; Oiso, Y. Biochemical roles of the oligosaccharide chains in thyrostimulin, a heterodimeric hormone of glycoprotein hormone subunits alpha2 (GPA2) and beta5 (GPB5). Regul. Pept. 2008, 148, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Okada, S.L.; Ellsworth, J.L.; Durnam, D.M.; Haugen, H.S.; Holloway, J.L.; Kelley, M.L.; Lewis, K.E.; Ren, H.; Sheppard, P.O.; Storey, H.M.; et al. A Glycoprotein Hormone Expressed in Corticotrophs Exhibits Unique Binding Properties on Thyroid-Stimulating Hormone Receptor. Mol. Endocrinol. 2006, 20, 414–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegedüs, L.; Hansen, J.M.; Karstrup, S. High incidence of normal thyroid gland volume in patients with Graves’ disease. Clin. Endocrinol. 1983, 19, 603–607. [Google Scholar] [CrossRef]
- Beck-Peccoz, P.; Persani, L.; Mannavola, D.; Campi, I. Pituitary tumours: TSH-secreting adenomas. Best Pr. Res. Clin. Endocrinol Metab 2009, 23, 597–606. [Google Scholar] [CrossRef]
- Nieto, H.; Boelaert, K. WOMEN IN CANCER THEMATIC REVIEW: Thyroid-stimulating hormone in thyroid cancer: Does it matter? Endocr. Relat. Cancer 2016, 23, T109–T121. [Google Scholar] [CrossRef] [Green Version]
- Franco, A.T.; Malaguarnera, R.; Refetoff, S.; Liao, X.H.; Lundsmith, E.; Kimura, S.; Pritchard, C.; Marais, R.; Davies, T.F.; Weinstein, L.S.; et al. Thyrotrophin receptor signaling dependence of Braf-induced thyroid tumor initiation in mice. Proc. Natl. Acad. Sci. USA 2011, 108, 1615–1620. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Choi, Y.W.; Han, J.H.; Lee, J.; Soh, E.Y.; Park, S.H.; Kim, J.H.; Park, T.J. TSH signaling overcomes B-RafV600E-induced senescence in papillary thyroid carcinogenesis through regulation of DUSP6. Neoplasia 2014, 16, 1107–1120. [Google Scholar] [CrossRef] [Green Version]
- Moulana, F.I.; Priyani, A.A.H.; de Silva, M.V.C.; Dassanayake, R.S. BRAF-Oncogene-Induced Senescence and the Role of Thyroid-Stimulating Hormone Signaling in the Progression of Papillary Thyroid Carcinoma. Horm. Cancer 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Orim, F.; Bychkov, A.; Shimamura, M.; Nakashima, M.; Ito, M.; Matsuse, M.; Kurashige, T.; Suzuki, K.; Saenko, V.; Nagayama, Y.; et al. Thyrotropin Signaling Confers More Aggressive Features with Higher Genomic Instability on BRAFV600E-Induced Thyroid Tumors in a Mouse Model. Thyroid 2014, 24, 502–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.; Zhao, L.; Ying, H.; Willingham, M.C.; Cheng, S.-y. Growth Activation Alone Is Not Sufficient to Cause Metastatic Thyroid Cancer in a Mouse Model of Follicular Thyroid Carcinoma. Endocrinology 2010, 151, 1929–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournes, B.; Monier, R.; Michiels, F.; Milgrom, E.; Misrahi, M.; Feunteun, J. Oncogenic potential of a mutant human thyrotropin receptor expressed in FRTL-5 cells. Oncogene 1998, 16, 985–990. [Google Scholar] [CrossRef] [Green Version]
- Rowe, C.W.; Paul, J.W.; Gedye, C.; Tolosa, J.M.; Bendinelli, C.; McGrath, S.; Smith, R. Targeting the TSH receptor in thyroid cancer. Endocr Relat Cancer 2017, 24, R191–R202. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Xu, J.; Bo, T.; Zhou, X.; Jiang, X.; Gao, L.; Zhao, J. Decreased fasting blood glucose is associated with impaired hepatic glucose production in thyroid-stimulating hormone receptor knockout mice. Endocr. J. 2013, 60, 941–950. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Wang, Q.; Lu, M.; Chen, W.; Song, Y.; Jing, F.; Guan, Y.; Wang, L.; Lin, Y.; Bo, T.; et al. Thyrotropin increases hepatic triglyceride content through upregulation of SREBP-1c activity. J. Hepatol. 2014, 61, 1358–1364. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Song, Y.; Feng, M.; Zhou, X.; Lu, Y.; Gao, L.; Yu, C.; Jiang, X.; Zhao, J. Thyroid-stimulating hormone decreases HMG-CoA reductase phosphorylation via AMP-activated protein kinase in the liver. J. Lipid. Res. 2015, 56, 963–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Xu, C.; Shao, S.; Liu, J.; Xing, W.; Xu, J.; Qin, C.; Li, C.; Hu, B.; Yi, S.; et al. Thyroid-stimulating hormone regulates hepatic bile acid homeostasis via SREBP-2/HNF-4alpha/CYP7A1 axis. J. Hepatol. 2015, 62, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zheng, D.; Zhao, M.; Qin, Y.; Wang, T.; Xing, W.; Gao, L.; Zhao, J. Thyroid-Stimulating Hormone Increases HNF-4alpha Phosphorylation via cAMP/PKA Pathway in the Liver. Sci. Rep. 2015, 5, 13409. [Google Scholar] [CrossRef] [Green Version]
- Niu, S.; Li, H.; Chen, W.; Zhao, J.; Gao, L.; Bo, T. Beta-Arrestin 1 Mediates Liver Thyrotropin Regulation of Cholesterol Conversion Metabolism via the Akt-Dependent Pathway. Int. J. Endocrinol. 2018, 2018, 4371396. [Google Scholar]
- Zhou, L.; Wu, K.; Zhang, L.; Gao, L.; Chen, S. Liver-specific deletion of TSHR inhibits hepatic lipid accumulation in mice. Biochem. Biophys. Res. Commun. 2018, 497, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Mato, J.M.; Alonso, C.; Noureddin, M.; Lu, S.C. Biomarkers and subtypes of deranged lipid metabolism in non- alcoholic fatty liver disease. World. J. Gastroenterol. 2019, 25, 3009–3020. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Lin, K.H.; Yeh, C.T. Thyroid Hormone in Hepatocellular Carcinoma: Cancer Risk, Growth Regulation, and Anticancer Drug Resistance. Front. Med. 2020, 7, 174. [Google Scholar] [CrossRef]
- Pinter, M.; Haupt, L.; Hucke, F.; Bota, S.; Bucsics, T.; Trauner, M.; Peck-Radosavljevic, M.; Sieghart, W. The impact of thyroid hormones on patients with hepatocellular carcinoma. PLoS ONE 2017, 12, e0181878. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.D.; Lin, K.H.; Huang, Y.H.; Lin, C.C.; Hung, C.F.; Yeh, T.S.; Lee, W.C.; Yeh, C.T. A novel thyroid function index associated with opposite therapeutic outcomes in advanced hepatocellular carcinoma patients receiving chemotherapy or sorafenib. Asia Pac. J. Clin. Oncol. 2018, 14, e341–e351. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.C.; Tsai, C.Y.; Tsai, M.M.; Yeh, C.T.; Lin, K.H. Molecular functions and clinical impact of thyroid hormone-triggered autophagy in liver-related diseases. J. Biomed. Sci. 2019, 26, 24. [Google Scholar] [CrossRef] [Green Version]
- Krashin, E.; Piekielko-Witkowska, A.; Ellis, M.; Ashur-Fabian, O. Thyroid Hormones and Cancer: A Comprehensive Review of Preclinical and Clinical Studies. Front. Endocrinol. 2019, 10, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schottenfeld, D.; Gershman, S.T. Epidemiology of thyroid cancer. Ca. Cancer J. Clin. 1978, 28, 66–86. [Google Scholar] [CrossRef] [PubMed]
- Reverter, J.L.; Lucas, A.; Salinas, I.; Audi, L.; Foz, M.; Sanmarti, A. Suppressive therapy with levothyroxine for solitary thyroid nodules. Clin. Endocrinol. 1992, 36, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.R. Historical note: TSH suppression for thyroid cancer. Thyroid 2011, 21, 1175–1176. [Google Scholar] [CrossRef]
- McGriff, N.J.; Csako, G.; Gourgiotis, L.; Guthrie, L.C.; Pucino, F.; Sarlis, N.J. Effects of thyroid hormone suppression therapy on adverse clinical outcomes in thyroid cancer. Ann. Med. 2002, 34, 554–564. [Google Scholar] [CrossRef]
- Mazzaferri, E.L.; Jhiang, S.M. Long-term impact of initial surgical and medical therapy on papillary and follicular thyroid cancer. Am. J. Med. 1994, 97, 418–428. [Google Scholar] [CrossRef]
- Yoon, B.H.; Lee, Y.; Oh, H.J.; Kim, S.H.; Lee, Y.K. Influence of Thyroid-stimulating Hormone Suppression Therapy on Bone Mineral Density in Patients with Differentiated Thyroid Cancer: A Meta-analysis. J. Bone Metab. 2019, 26, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Suh, B.; Shin, D.W.; Park, Y.; Lim, H.; Yun, J.M.; Song, S.O.; Park, J.H.; Cho, B.; Guallar, E. Increased cardiovascular risk in thyroid cancer patients taking levothyroxine: A nationwide cohort study in Korea. Eur. J. Endocrinol. 2019, 180, 11–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edmonds, C.J.; Hayes, S.; Kermode, J.C.; Thompson, B.D. Measurement of serum TSH and thyroid hormones in the management of treatment of thyroid carcinoma with radioiodine. Br. J. Radiol. 1977, 50, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Fallahi, B.; Beiki, D.; Takavar, A.; Fard-Esfahani, A.; Gilani, K.A.; Saghari, M.; Eftekhari, M. Low versus high radioiodine dose in postoperative ablation of residual thyroid tissue in patients with differentiated thyroid carcinoma: A large randomized clinical trial. Nucl. Med. Commun. 2012, 33, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Paolino, D.; Cosco, D.; Gaspari, M.; Celano, M.; Wolfram, J.; Voce, P.; Puxeddu, E.; Filetti, S.; Celia, C.; Ferrari, M.; et al. Targeting the thyroid gland with thyroid-stimulating hormone (TSH)-nanoliposomes. Biomaterials 2014, 35, 7101–7109. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.-j.; Li, A.-q.; Zhang, X.; Liu, P.; Wang, J.-R.; Cai, X. Thyroid-stimulating hormone (TSH)-armed polymer–lipid nanoparticles for the targeted delivery of cisplatin in thyroid cancers: Therapeutic efficacy evaluation. Rsc. Adv. 2015, 5, 106413–106420. [Google Scholar] [CrossRef]
- Neumann, S.; Gershengorn, M.C. Small molecule TSHR agonists and antagonists. Ann. Endocrinol. 2011, 72, 74–76. [Google Scholar] [CrossRef] [Green Version]
- Neumann, S.; Pope, A.; Geras-Raaka, E.; Raaka, B.M.; Bahn, R.S.; Gershengorn, M.C. A drug-like antagonist inhibits thyrotropin receptor-mediated stimulation of cAMP production in Graves’ orbital fibroblasts. Thyroid 2012, 22, 839–843. [Google Scholar] [CrossRef] [Green Version]
- Neumann, S.; Eliseeva, E.; McCoy, J.G.; Napolitano, G.; Giuliani, C.; Monaco, F.; Huang, W.; Gershengorn, M.C. A new small-molecule antagonist inhibits Graves’ disease antibody activation of the TSH receptor. J. Clin. Endocr. Metab. 2011, 96, 548–554. [Google Scholar] [CrossRef]
- Marcinkowski, P.; Hoyer, I.; Specker, E.; Furkert, J.; Rutz, C.; Neuenschwander, M.; Sobottka, S.; Sun, H.; Nazare, M.; Berchner-Pfannschmidt, U.; et al. A New Highly Thyrotropin Receptor-Selective Small-Molecule Antagonist with Potential for the Treatment of Graves’ Orbitopathy. Thyroid 2019, 29, 111–123. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Benign Tissues | Expression a | Functionality b | Reference |
|---|---|---|---|
| Adipose tissue | Both mRNA and protein | Yes | [44,45,46] |
| Adrenal | Both mRNA and protein | No | [47] |
| Endometrium | Both mRNA and protein | No | [48] |
| Erythrocytes | Protein | Yes | [49,50] |
| Kidney | Both mRNA and protein | No | [47] |
| Liver | Both mRNA and protein | Yes | [51,52] |
| Lymphocytes | Both mRNA and protein | No | [53] |
| Pituitary | Both mRNA and protein | No | [54] |
| Cardiac muscle | mRNA | Yes | [55] |
| Hair follicles | Both mRNA and protein | Yes | [56] |
| Vascular smooth muscle | Both mRNA and protein | Yes | [57] |
| Ovary | Both mRNA and protein | Yes | [58,59] |
| Malignant tissues | Expression a | Functionality b | Reference |
| Melanoma | Both mRNA and protein | Yes | [39] |
| Glioma | mRNA | No | [38] |
| Lung cancer | Protein | No | [40] |
| Breast cancer | Both mRNA and protein | No | [41] |
| Ovarian cancer | Both mRNA and protein | Yes | [42,59,60,61] |
| Hepatocellular carcinoma | Protein | Yes | [43] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, Y.-D.; Yeh, C.-T. The Molecular Function and Clinical Role of Thyroid Stimulating Hormone Receptor in Cancer Cells. Cells 2020, 9, 1730. https://doi.org/10.3390/cells9071730
Chu Y-D, Yeh C-T. The Molecular Function and Clinical Role of Thyroid Stimulating Hormone Receptor in Cancer Cells. Cells. 2020; 9(7):1730. https://doi.org/10.3390/cells9071730
Chicago/Turabian StyleChu, Yu-De, and Chau-Ting Yeh. 2020. "The Molecular Function and Clinical Role of Thyroid Stimulating Hormone Receptor in Cancer Cells" Cells 9, no. 7: 1730. https://doi.org/10.3390/cells9071730
APA StyleChu, Y. -D., & Yeh, C. -T. (2020). The Molecular Function and Clinical Role of Thyroid Stimulating Hormone Receptor in Cancer Cells. Cells, 9(7), 1730. https://doi.org/10.3390/cells9071730