Schizosaccharomyces pombe Assays to Study Mitotic Recombination Outcomes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
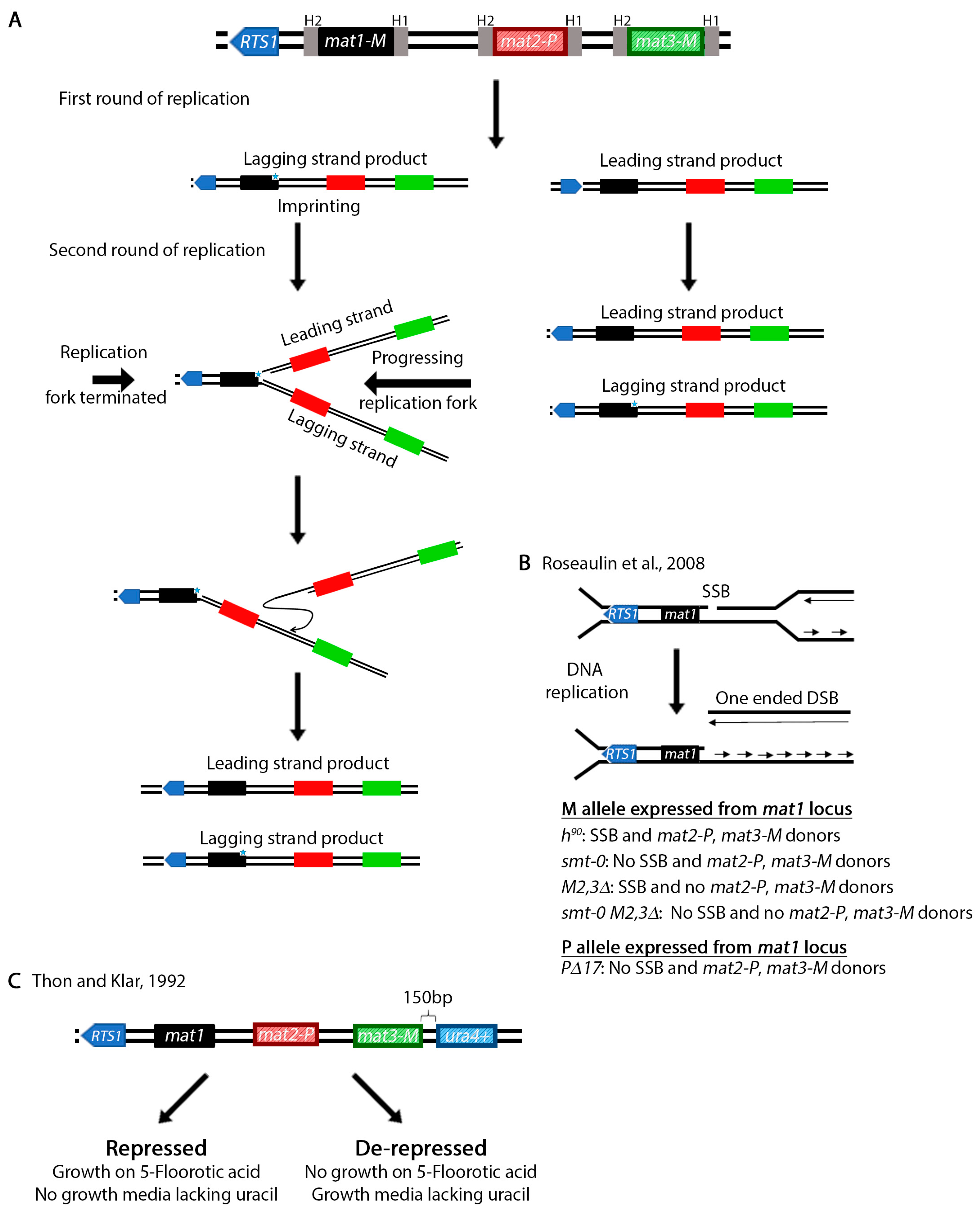{kind=link}
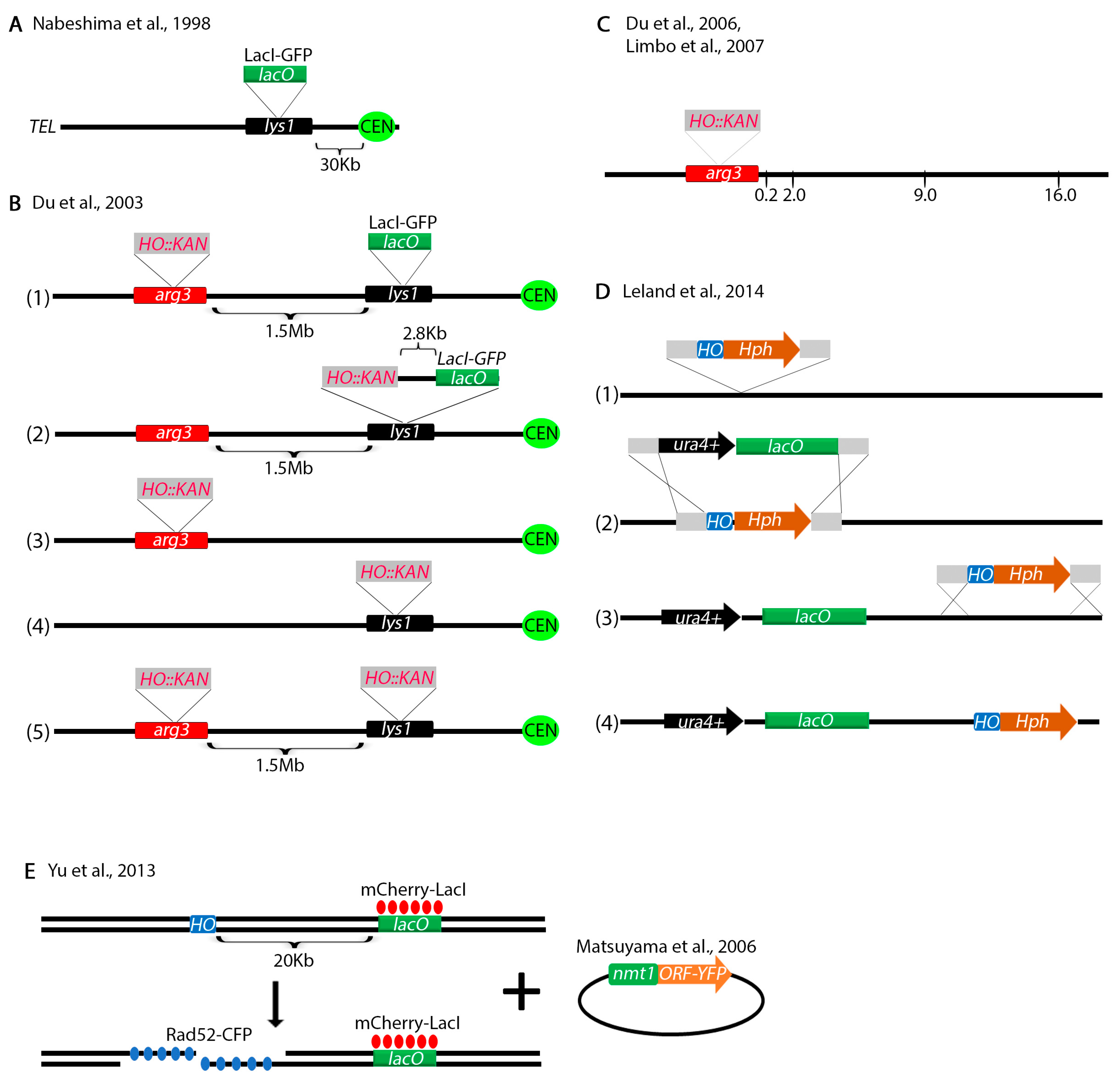{kind=link}
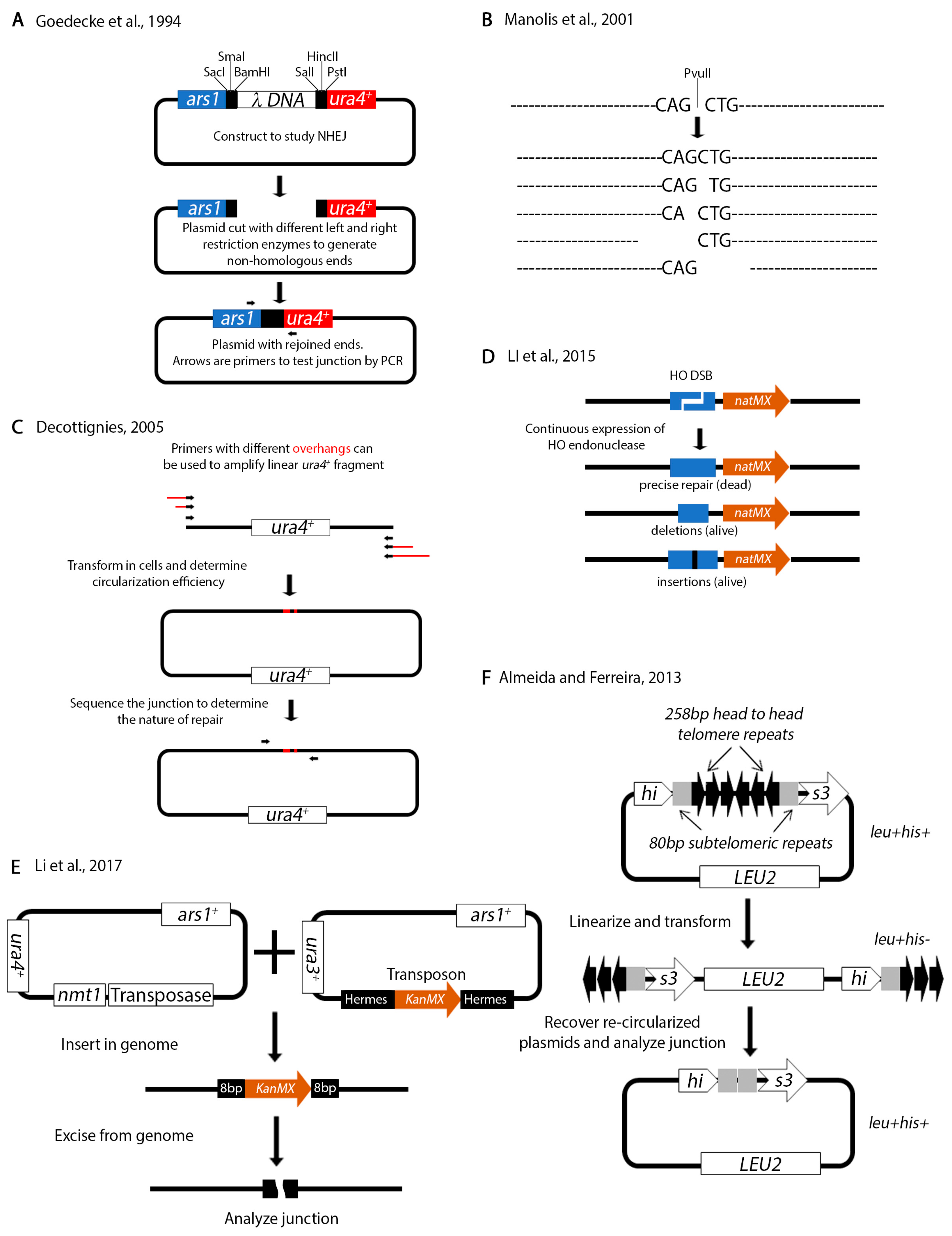{kind=link}
Abstract
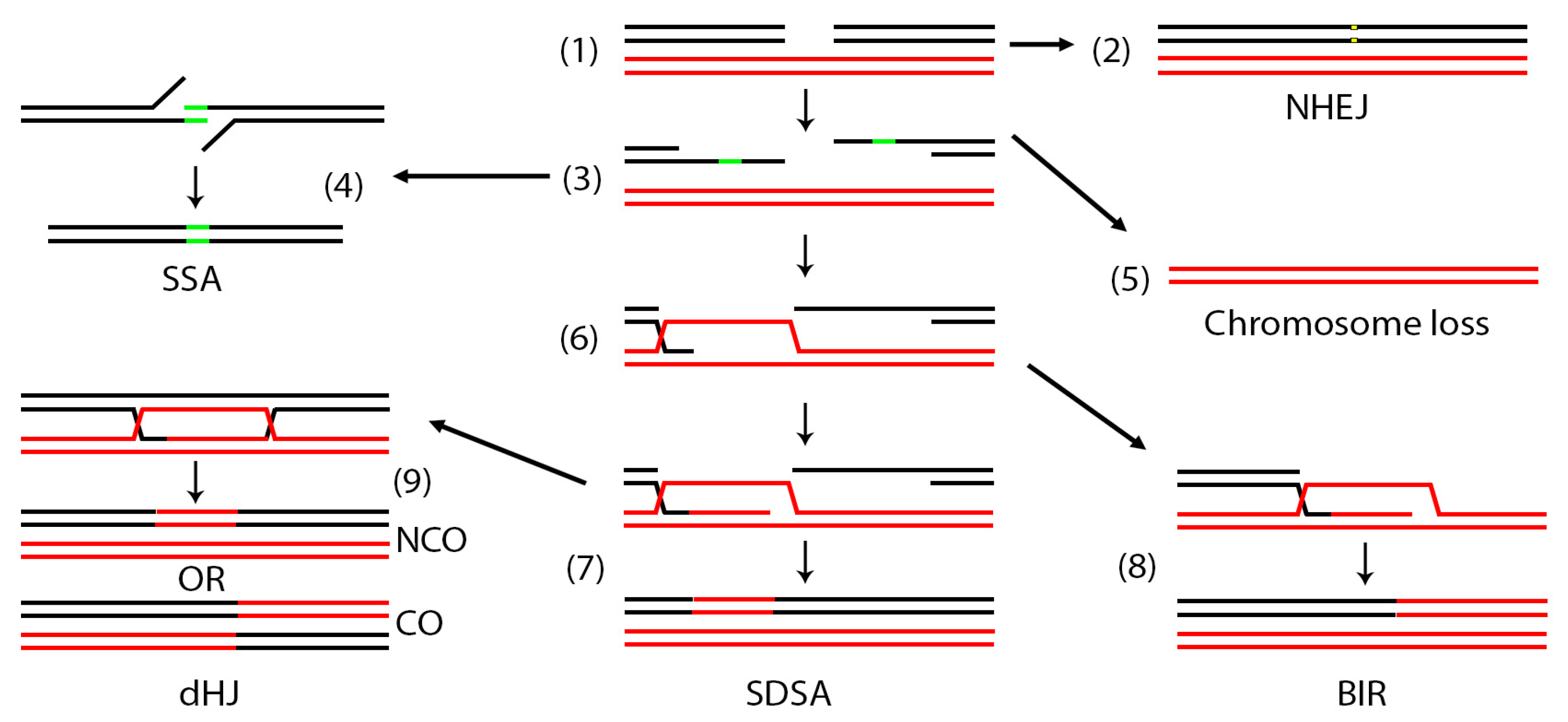
:1. Introduction
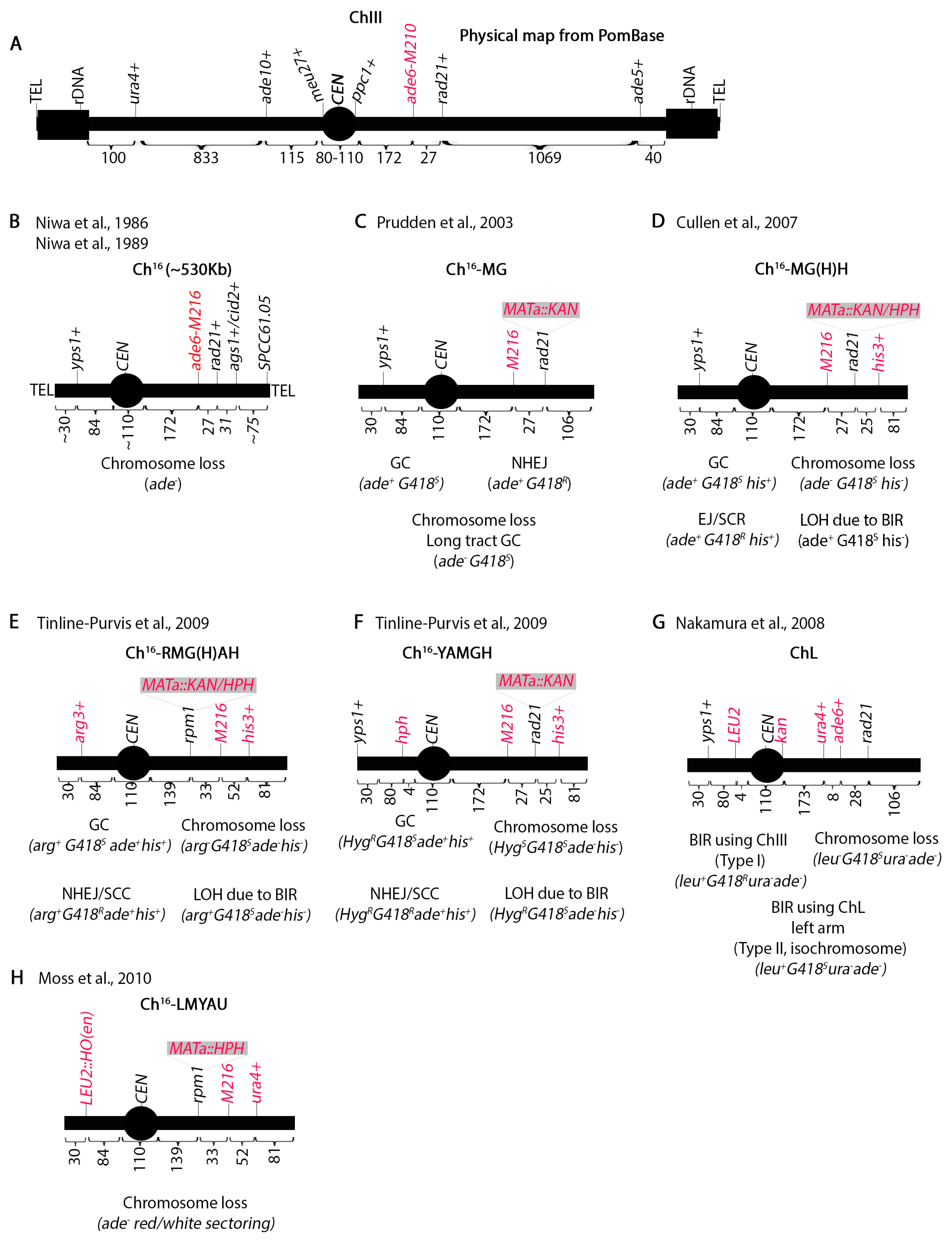
2. Mini-Chromosome Assays
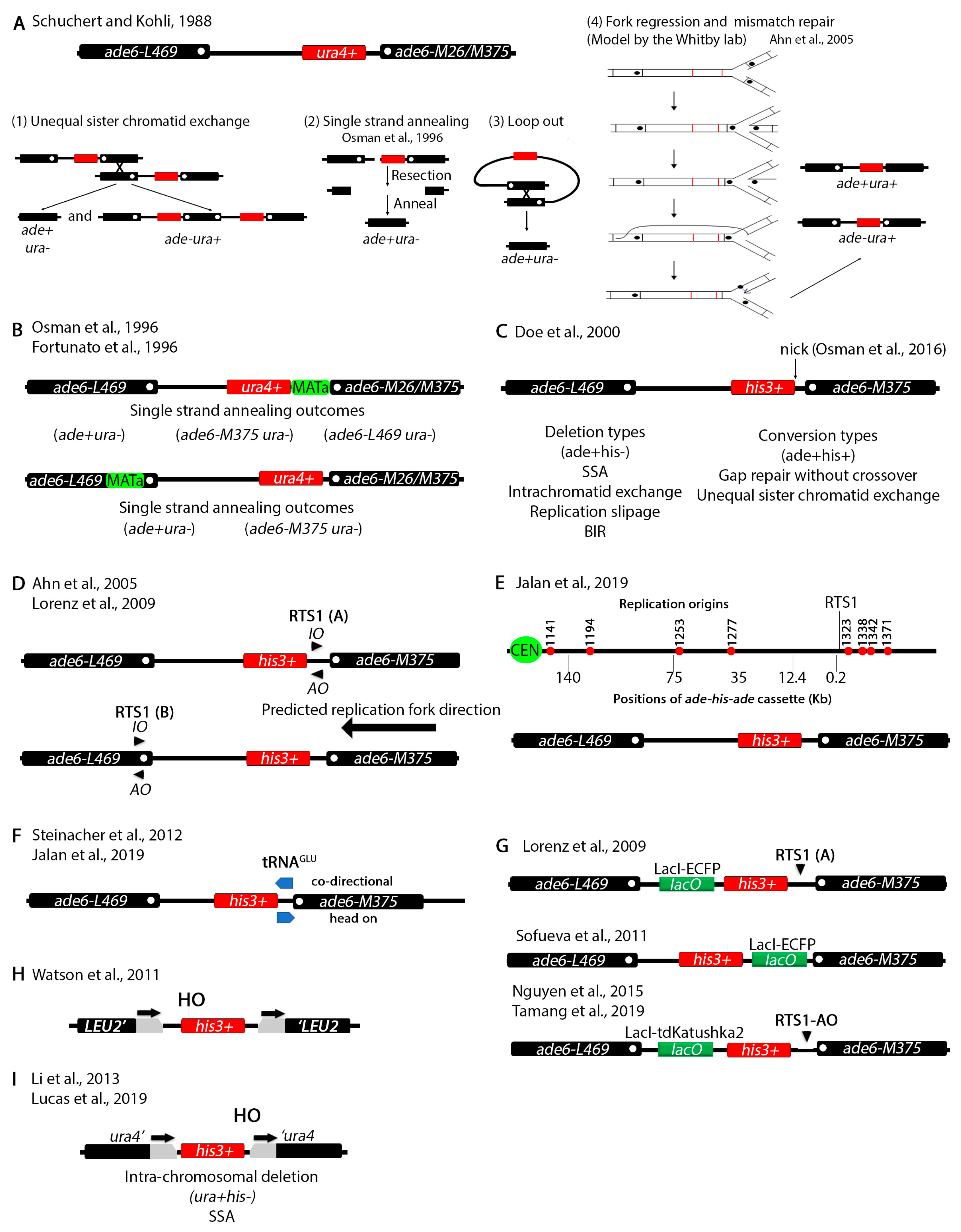
3. Recombination at Repetitive Elements
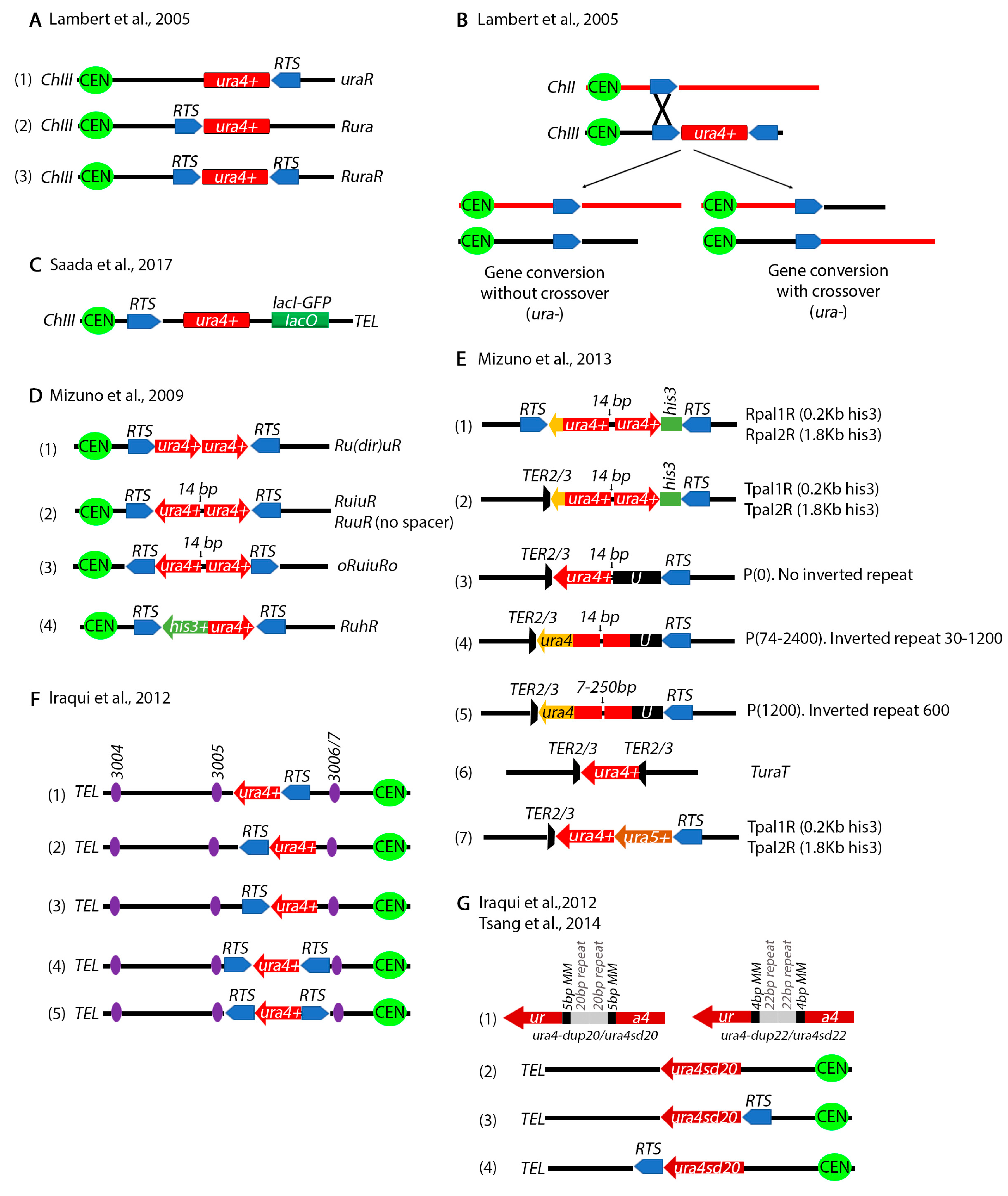
4. Chromosomal Rearrangements Caused by Stalled or Collapsed Replication Forks at Inverted Repeats
5. Mating Type Loci Serve as a Natural Site for Studying Collapsed Replication Forks
6. Other Fluorescence and Biochemical Assays
7. Non-Homologous Repair
8. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kerzendorfer, C.; O’Driscoll, M. Human DNA damage response and repair deficiency syndromes: Linking genomic instability and cell cycle checkpoint proficiency. DNA Repair 2009, 8, 1139–1152. [Google Scholar] [CrossRef]
- White, R.R.; Vijg, J. Do DNA Double-Strand Breaks Drive Aging? Mol. Cell 2016, 63, 729–738. [Google Scholar] [CrossRef] [Green Version]
- Jeggo, P.A.; Lobrich, M. How cancer cells hijack DNA double-strand break repair pathways to gain genomic instability. Biochem. J. 2015, 471, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Janssen, A.; van der Burg, M.; Szuhai, K.; Kops, G.J.; Medema, R.H. Chromosome segregation errors as a cause of DNA damage and structural chromosome aberrations. Science 2011, 333, 1895–1898. [Google Scholar] [CrossRef] [PubMed]
- Lasko, D.; Cavenee, W.; Nordenskjold, M. Loss of constitutional heterozygosity in human cancer. Annu. Rev. Genet. 1991, 25, 281–314. [Google Scholar] [CrossRef] [PubMed]
- Ortega, V.; Chaubey, A.; Mendiola, C.; Ehman, W., Jr.; Vadlamudi, K.; Dupont, B.; Velagaleti, G. Complex Chromosomal Rearrangements in B-Cell Lymphoma: Evidence of Chromoanagenesis? A Case Report. Neoplasia 2016, 18, 223–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, A.; Haber, J.E. Sources of DNA double-strand breaks and models of recombinational DNA repair. Cold Spring Harb. Perspect. Biol. 2014, 6, a016428. [Google Scholar] [CrossRef] [Green Version]
- Nikolov, I.; Taddei, A. Linking replication stress with heterochromatin formation. Chromosoma 2016, 125, 523–533. [Google Scholar] [CrossRef] [Green Version]
- Tabancay, A.P., Jr.; Forsburg, S.L. Eukaryotic DNA replication in a chromatin context. Curr. Top. Dev. Biol. 2006, 76, 129–184. [Google Scholar] [CrossRef]
- Prado, F.; Aguilera, A. Impairment of replication fork progression mediates RNA polII transcription-associated recombination. EMBO J. 2005, 24, 1267–1276. [Google Scholar] [CrossRef] [Green Version]
- Kaushal, S.; Freudenreich, C.H. The role of fork stalling and DNA structures in causing chromosome fragility. Genes Chromosomes Cancer 2019, 58, 270–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irony-Tur Sinai, M.; Kerem, B. Genomic instability in fragile sites-still adding the pieces. Genes Chromosomes Cancer 2019, 58, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Cavalier-Smith, T. Origins of the machinery of recombination and sex. Heredity 2002, 88, 125–141. [Google Scholar] [CrossRef] [PubMed]
- Piazza, A.; Heyer, W.D. Homologous Recombination and the Formation of Complex Genomic Rearrangements. Trends. Cell. Biol. 2019, 29, 135–149. [Google Scholar] [CrossRef]
- West, S.C.; Blanco, M.G.; Chan, Y.W.; Matos, J.; Sarbajna, S.; Wyatt, H.D. Resolution of Recombination Intermediates: Mechanisms and Regulation. Cold Spring Harb. Symp. Quant. Biol. 2015, 80, 103–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prado, F. Homologous recombination maintenance of genome integrity during DNA damage tolerance. Mol. Cell. Oncol. 2014, 1, e957039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- San Filippo, J.; Sung, P.; Klein, H. Mechanism of eukaryotic homologous recombination. Annu. Rev. Biochem. 2008, 77, 229–257. [Google Scholar] [CrossRef] [Green Version]
- Ait Saada, A.; Lambert, S.A.E.; Carr, A.M. Preserving replication fork integrity and competence via the homologous recombination pathway. DNA Repair 2018, 71, 135–147. [Google Scholar] [CrossRef]
- Daley, J.M.; Kwon, Y.; Niu, H.; Sung, P. Investigations of homologous recombination pathways and their regulation. Yale J. Biol. Med. 2013, 86, 453–461. [Google Scholar]
- Pannunzio, N.R.; Li, S.; Watanabe, G.; Lieber, M.R. Non-homologous end joining often uses microhomology: Implications for alternative end joining. DNA Repair 2014, 17, 74–80. [Google Scholar] [CrossRef] [Green Version]
- Malkova, A.; Haber, J.E. Mutations arising during repair of chromosome breaks. Annu. Rev. Genet. 2012, 46, 455–473. [Google Scholar] [CrossRef] [PubMed]
- Hedges, S.B. The origin and evolution of model organisms. Nat. Rev. Genet. 2002, 3, 838–849. [Google Scholar] [CrossRef] [PubMed]
- Heckman, D.S.; Geiser, D.M.; Eidell, B.R.; Stauffer, R.L.; Kardos, N.L.; Hedges, S.B. Molecular evidence for the early colonization of land by fungi and plants. Science 2001, 293, 1129–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, V.; Gwilliam, R.; Rajandream, M.A.; Lyne, M.; Lyne, R.; Stewart, A.; Sgouros, J.; Peat, N.; Hayles, J.; Baker, S.; et al. The genome sequence of Schizosaccharomyces pombe. Nature 2002, 415, 871–880. [Google Scholar] [CrossRef] [Green Version]
- Rubin, G.M.; Yandell, M.D.; Wortman, J.R.; Gabor Miklos, G.L.; Nelson, C.R.; Hariharan, I.K.; Fortini, M.E.; Li, P.W.; Apweiler, R.; Fleischmann, W.; et al. Comparative genomics of the eukaryotes. Science 2000, 287, 2204–2215. [Google Scholar] [CrossRef] [Green Version]
- Forsburg, S.L. The yeasts Saccharomyces cerevisiae and Schizosaccharomyces pombe: Models for cell biology research. Gravit. Space Biol. Bull. 2005, 18, 3–9. [Google Scholar]
- Halazonetis, T.D.; Gorgoulis, V.G.; Bartek, J. An oncogene-induced DNA damage model for cancer development. Science 2008, 319, 1352–1355. [Google Scholar] [CrossRef] [Green Version]
- Branzei, D.; Foiani, M. Maintaining genome stability at the replication fork. Nat. Rev. Mol. Cell Biol. 2010, 11, 208–219. [Google Scholar] [CrossRef]
- Madireddy, A.; Gerhardt, J. Replication Through Repetitive DNA Elements and Their Role in Human Diseases. DNA Replication Old Princ. New Discov. 2017, 1042, 549–581. [Google Scholar] [CrossRef]
- Forsburg, S.L. The CINs of the centromere. Biochem. Soc. Trans. 2013, 41, 1706–1711. [Google Scholar] [CrossRef] [Green Version]
- Li, P.C.; Petreaca, R.C.; Jensen, A.; Yuan, J.P.; Green, M.D.; Forsburg, S.L. Replication fork stability is essential for the maintenance of centromere integrity in the absence of heterochromatin. Cell Rep. 2013, 3, 638–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onaka, A.T.; Toyofuku, N.; Inoue, T.; Okita, A.K.; Sagawa, M.; Su, J.; Shitanda, T.; Matsuyama, R.; Zafar, F.; Takahashi, T.S.; et al. Rad51 and Rad54 promote noncrossover recombination between centromere repeats on the same chromatid to prevent isochromosome formation. Nucleic Acids Res. 2016, 44, 10744–10757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, K.; Okamoto, A.; Katou, Y.; Yadani, C.; Shitanda, T.; Kaweeteerawat, C.; Takahashi, T.S.; Itoh, T.; Shirahige, K.; Masukata, H.; et al. Rad51 suppresses gross chromosomal rearrangement at centromere in Schizosaccharomyces pombe. EMBO J. 2008, 27, 3036–3046. [Google Scholar] [CrossRef] [Green Version]
- Forsburg, S.L.; Shen, K.F. Centromere Stability: The Replication Connection. Genes-Basel 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- He, H.J.; Gonzalez, M.; Zhang, F.; Li, F. DNA replication components as regulators of epigenetic inheritance-lesson from fission yeast centromere. Protein Cell 2014, 5, 411–419. [Google Scholar] [CrossRef] [Green Version]
- Mizuguchi, T.; Barrowman, J.; Grewal, S.I.S. Chromosome domain architecture and dynamic organization of the fission yeast genome. Febs Lett. 2015, 589, 2975–2986. [Google Scholar] [CrossRef] [Green Version]
- Forsburg, S.L. The art and design of genetic screens: Yeast. Nat. Rev. Genet. 2001, 2, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sun, H.; Huang, Y.; Wang, Y.; Liu, Y.; Chen, X. Pathways and assays for DNA double-strand break repair by homologous recombination. Acta Biochim. Biophys. Sin. 2019, 51, 879–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, H.L.; Bacinskaja, G.; Che, J.; Cheblal, A.; Elango, R.; Epshtein, A.; Fitzgerald, D.M.; Gomez-Gonzalez, B.; Khan, S.R.; Kumar, S.; et al. Guidelines for DNA recombination and repair studies: Cellular assays of DNA repair pathways. Microb. Cell 2019, 6, 1–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, H.L.; Ang, K.K.H.; Arkin, M.R.; Beckwitt, E.C.; Chang, Y.H.; Fan, J.; Kwon, Y.; Morten, M.J.; Mukherjee, S.; Pambos, O.J.; et al. Guidelines for DNA recombination and repair studies: Mechanistic assays of DNA repair processes. Microb. Cell 2019, 6, 65–101. [Google Scholar] [CrossRef]
- Kai, M.; Taricani, L.; Wang, T.S. Methods for studying mutagenesis and checkpoints in Schizosaccharomyces pombe. Methods Enzymol. 2006, 409, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Niwa, O.; Matsumoto, T.; Yanagida, M. Construction of a Minichromosome by Deletion and Its Mitotic and Meiotic Behavior in Fission Yeast. Mol. Gen. Genet. 1986, 203, 397–405. [Google Scholar] [CrossRef]
- Niwa, O.; Matsumoto, T.; Chikashige, Y.; Yanagida, M. Characterization of Schizosaccharomyces pombe minichromosome deletion derivatives and a functional allocation of their centromere. EMBO J. 1989, 8, 3045–3052. [Google Scholar] [CrossRef] [PubMed]
- Bloom, K.; Costanzo, V. Centromere Structure and Function. Prog. Mol. Subcell. Biol. 2017, 56, 515–539. [Google Scholar] [CrossRef]
- Kohli, J.; Hottinger, H.; Munz, P.; Strauss, A.; Thuriaux, P. Genetic-Mapping in Schizosaccharomyces-Pombe by Mitotic and Meiotic Analysis and Induced Haploidization. Genetics 1977, 87, 471–489. [Google Scholar]
- Steiner, N.C.; Hahnenberger, K.M.; Clarke, L. Centromeres of the Fission Yeast Schizosaccharomyces-Pombe Are Highly Variable Genetic-Loci. Mol. Cell. Biol. 1993, 13, 4578–4587. [Google Scholar] [CrossRef] [Green Version]
- Gygax, A.; Thuriaux, P. A Revised Chromosome Map of the Fission Yeast Schizosaccharomyces-Pombe. Curr. Genet. 1984, 8, 85–92. [Google Scholar] [CrossRef]
- Murakami, S.; Matsumoto, T.; Niwa, O.; Yanagida, M. Structure of the fission yeast centromere cen3: Direct analysis of the reiterated inverted region. Chromosoma 1991, 101, 214–221. [Google Scholar] [CrossRef]
- Lock, A.; Rutherford, K.; Harris, M.A.; Wood, V. PomBase: The Scientific Resource for Fission Yeast. Eukaryot. Genom. Databases Methods Protoc. 2018, 1757, 49–68. [Google Scholar] [CrossRef] [Green Version]
- PomBase. The Scientific Resource for Fission Yeast. Available online: https://www.pombase.org/ (accessed on 15 December 2019).
- Tinline-Purvis, H.; Savory, A.P.; Cullen, J.K.; Dave, A.; Moss, J.; Bridge, W.L.; Marguerat, S.; Bahler, J.; Ragoussis, J.; Mott, R.; et al. Failed gene conversion leads to extensive end processing and chromosomal rearrangements in fission yeast. EMBO J. 2009, 28, 3400–3412. [Google Scholar] [CrossRef] [Green Version]
- Cromie, G.A.; Rubio, C.A.; Hyppa, R.W.; Smith, G.R. A natural meiotic DNA break site in Schizosaccharomyces pombe is a hotspot of gene conversion, highly associated with crossing over. Genetics 2005, 169, 595–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, J.K.; Hussey, S.P.; Walker, C.; Prudden, J.; Wee, B.Y.; Dave, A.; Findlay, J.S.; Savory, A.P.; Humphrey, T.C. Break-induced loss of heterozygosity in fission yeast: Dual roles for homologous recombination in promoting translocations and preventing de novo telomere addition. Mol. Cell. Biol. 2007, 27, 7745–7757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prudden, J.; Evans, J.S.; Hussey, S.P.; Deans, B.; O’Neill, P.; Thacker, J.; Humphrey, T. Pathway utilization in response to a site-specific DNA double-strand break in fission yeast. EMBO J. 2003, 22, 1419–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.U.; Hayles, J.; Kim, D.; Wood, V.; Park, H.O.; Won, M.; Yoo, H.S.; Duhig, T.; Nam, M.; Palmer, G.; et al. Analysis of a genome-wide set of gene deletions in the fission yeast Schizosaccharomyces pombe. Nat. Biotechnol. 2010, 28, 617–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moss, J.; Tinline-Purvis, H.; Walker, C.A.; Folkes, L.K.; Stratford, M.R.; Hayles, J.; Hoe, K.L.; Kim, D.U.; Park, H.O.; Kearsey, S.E.; et al. Break-induced ATR and Ddb1-Cul4(Cdt)(2) ubiquitin ligase-dependent nucleotide synthesis promotes homologous recombination repair in fission yeast. Genes Dev. 2010, 24, 2705–2716. [Google Scholar] [CrossRef] [Green Version]
- Pai, C.C.; Blaikley, E.; Humphrey, T.C. DNA Double-Strand Break Repair Assay. Cold Spring Harb. Protoc. 2018, 2018. [Google Scholar] [CrossRef]
- Hartsuiker, E.; Vaessen, E.; Carr, A.M.; Kohli, J. Fission yeast Rad50 stimulates sister chromatid recombination and links cohesion with repair. EMBO J. 2001, 20, 6660–6671. [Google Scholar] [CrossRef] [Green Version]
- Jackson, J.A.; Fink, G.R. Gene Conversion between Duplicated Genetic Elements in Yeast. Nature 1981, 292, 306–311. [Google Scholar] [CrossRef]
- Rothstein, R.; Helms, C.; Rosenberg, N. Concerted Deletions and Inversions Are Caused by Mitotic Recombination between Delta-Sequences in Saccharomyces-Cerevisiae. Mol. Cell. Biol. 1987, 7, 1198–1207. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, M.V.; Whitehouse, H.L.K. A Buff Spore Color Mutant in Sordaria-Brevicollis Showing High-Frequency Conversion: 2. Loss of the High-Frequency Conversion. Genet. Res. 1983, 41, 155–163. [Google Scholar] [CrossRef]
- Pukkila, P.J.; Stephens, M.D.; Binninger, D.M.; Errede, B. Frequency and Directionality of Gene Conversion Events Involving the Cyc7-H3 Mutation in Saccharomyces-Cerevisiae. Genetics 1986, 114, 347–361. [Google Scholar] [PubMed]
- Gutz, H. Site Specific Induction of Gene Conversion in schizosaccharomyces pombe. Genetics 1971, 69, 317–337. [Google Scholar] [PubMed]
- Schuchert, P.; Kohli, J. The Ade6-M26 Mutation of Schizosaccharomyces-Pombe Increases the Frequency of Crossing Over. Genetics 1988, 119, 507–515. [Google Scholar] [PubMed]
- Fortunato, E.A.; Osman, F.; Subramani, S. Analysis of spontaneous and double-strand break-induced recombination in rad mutants of S. pombe. Mutat. Res. 1996, 364, 14–60. [Google Scholar] [CrossRef]
- Osman, F.; Fortunato, E.A.; Subramani, S. Double-strand break-induced mitotic intrachromosomal recombination in the fission yeast Schizosaccharomyces pombe. Genetics 1996, 142, 341–357. [Google Scholar] [PubMed]
- Ahamad, N.; Verma, S.K.; Ahmed, S. Activation of Checkpoint Kinase Chk1 by Reactive Oxygen Species Resulting from Disruption of wat1/pop3 in Schizosaccharomyces pombe. Genetics 2016, 204, 1397–1406. [Google Scholar] [CrossRef]
- Bellini, A.; Girard, P.M.; Lambert, S.; Tessier, L.; Sage, E.; Francesconi, S. Stress activated protein kinase pathway modulates homologous recombination in fission yeast. PLoS ONE 2012, 7, e47987. [Google Scholar] [CrossRef] [Green Version]
- Nandi, S.; Whitby, M.C. The ATPase activity of Fml1 is essential for its roles in homologous recombination and DNA repair. Nucleic Acids Res. 2012, 40, 9584–9595. [Google Scholar] [CrossRef]
- Osman, F.; Tsaneva, I.R.; Whitby, M.C.; Doe, C.L. UV irradiation causes the loss of viable mitotic recombinants in Schizosaccharomyces pombe cells lacking the G(2)/M DNA damage checkpoint. Genetics 2002, 160, 891–908. [Google Scholar]
- Bass, K.L.; Murray, J.M.; O’Connell, M.J. Brc1-dependent recovery from replication stress. J. Cell Sci. 2012, 125, 2753–2764. [Google Scholar] [CrossRef] [Green Version]
- Osman, F.; Bjoras, M.; Alseth, I.; Morland, I.; McCready, S.; Seeberg, E.; Tsaneva, I. A new Schizosaccharomyces pombe base excision repair mutant, nth1, reveals overlapping pathways for repair of DNA base damage. Mol. Microbiol. 2003, 48, 465–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommariva, E.; Pellny, T.K.; Karahan, N.; Kumar, S.; Huberman, J.A.; Dalgaard, J.Z. Schizosaccharomyces pombe Swi1, Swi3, and Hsk1 are components of a novel S-phase response pathway to alkylation damage. Mol. Cell. Biol. 2005, 25, 2770–2784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, M.; Inoue, H.; Ishii, C.; Murakami, Y. The novel gene mus7(+) is involved in the repair of replication-associated DNA damage in fission yeast. DNA Repair 2007, 6, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Tomita, K.; Matsuura, A.; Nakagawa, T.; Masukata, H.; Uritani, M.; Ushimaru, T.; Ueno, M. A novel allele of fission yeast rad11 that causes defects in DNA repair and telomere length regulation. Nucleic Acids Res. 2003, 31, 7141–7149. [Google Scholar] [CrossRef] [PubMed]
- Steinacher, R.; Osman, F.; Lorenz, A.; Bryer, C.; Whitby, M.C. Slx8 Removes Pli1-Dependent Protein-SUMO Conjugates Including SUMOylated Topoisomerase I to Promote Genome Stability. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, E.; Miyabe, I.; Iraqui, I.; Zheng, J.; Lambert, S.A.; Carr, A.M. The extent of error-prone replication restart by homologous recombination is controlled by Exo1 and checkpoint proteins. J. Cell Sci. 2014, 127, 2983–2994. [Google Scholar] [CrossRef] [Green Version]
- Octobre, G.; Lorenz, A.; Loidl, J.; Kohli, J. The Rad52 homologs Rad22 and Rti1 of Schizosaccharomyces pombe are not essential for meiotic interhomolog recombination, but are required for meiotic intrachromosomal recombination and mating-type-related DNA repair. Genetics 2008, 178, 2399–2412. [Google Scholar] [CrossRef] [Green Version]
- Doe, C.L.; Dixon, J.; Osman, F.; Whitby, M.C. Partial suppression of the fission yeast rqh1(-) phenotype by expression of a bacterial Holliday junction resolvase. EMBO J. 2000, 19, 2751–2762. [Google Scholar] [CrossRef] [Green Version]
- Doe, C.L.; Osman, F.; Dixon, J.; Whitby, M.C. The Holliday junction resolvase SpCCE1 prevents mitochondrial DNA aggregation in Schizosaccharomyces pombe. Mol. Gen. Genet. 2000, 263, 889–897. [Google Scholar] [CrossRef]
- Doe, C.L.; Whitby, M.C. The involvement of Srs2 in post-replication repair and homologous recombination in fission yeast. Nucleic Acids Res. 2004, 32, 1480–1491. [Google Scholar] [CrossRef] [Green Version]
- Doe, C.L.; Osman, F.; Dixon, J.; Whitby, M.C. DNA repair by a Rad22-Mus81-dependent pathway that is independent of Rhp51. Nucleic Acids Res. 2004, 32, 5570–5581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osman, F.; Ahn, J.S.; Lorenz, A.; Whitby, M.C. The RecQ DNA helicase Rqh1 constrains Exonuclease 1-dependent recombination at stalled replication forks. Sci. Rep. 2016, 6, 22837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzminov, A. Single-strand interruptions in replicating chromosomes cause double-strand breaks. Proc. Natl. Acad. Sci. USA 2001, 98, 8241–8246. [Google Scholar] [CrossRef] [Green Version]
- Asano, S.; Higashitani, A.; Horiuchi, K. Filamentous phage replication initiator protein gpII forms a covalent complex with the 5’ end of the nick it introduced. Nucleic Acids Res. 1999, 27, 1882–1889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalgaard, J.Z.; Klar, A.J. A DNA replication-arrest site RTS1 regulates imprinting by determining the direction of replication at mat1 in S. pombe. Genes Dev. 2001, 15, 2060–2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, J.S.; Osman, F.; Whitby, M.C. Replication fork blockage by RTS1 at an ectopic site promotes recombination in fission yeast. EMBO J. 2005, 24, 2011–2023. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, A.; Osman, F.; Folkyte, V.; Sofueva, S.; Whitby, M.C. Fbh1 limits Rad51-dependent recombination at blocked replication forks. Mol. Cell. Biol. 2009, 29, 4742–4756. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.O.; Jalan, M.; Morrow, C.A.; Osman, F.; Whitby, M.C. Recombination occurs within minutes of replication blockage by RTS1 producing restarted forks that are prone to collapse. Elife 2015, 4, e04539. [Google Scholar] [CrossRef]
- Sun, W.; Nandi, S.; Osman, F.; Ahn, J.S.; Jakovleska, J.; Lorenz, A.; Whitby, M.C. The FANCM Ortholog Fml1 Promotes Recombination at Stalled Replication Forks and Limits Crossing Over during DNA Double-Strand Break Repair. Mol. Cell. 2008, 32, 118–128. [Google Scholar] [CrossRef]
- Steinacher, R.; Osman, F.; Dalgaard, J.Z.; Lorenz, A.; Whitby, M.C. The DNA helicase Pfh1 promotes fork merging at replication termination sites to ensure genome stability. Gene Dev. 2012, 26, 594–602. [Google Scholar] [CrossRef] [Green Version]
- Osman, F.; Whitby, M.C. Monitoring homologous recombination following replication fork perturbation in the fission yeast Schizosaccharomyces pombe. Methods Mol. Biol. 2009, 521, 535–552. [Google Scholar] [CrossRef] [PubMed]
- Jalan, M.; Oehler, J.; Morrow, C.A.; Osman, F.; Whitby, M.C. Factors affecting template switch recombination associated with restarted DNA replication. Elife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- McFarlane, R.J.; Whitehall, S.K. tRNA genes in eukaryotic genome organization and reorganization. Cell Cycle 2009, 8, 3102–3106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sofueva, S.; Osman, F.; Lorenz, A.; Steinacher, R.; Castagnetti, S.; Ledesma, J.; Whitby, M.C. Ultrafine anaphase bridges, broken DNA and illegitimate recombination induced by a replication fork barrier. Nucleic Acids Res. 2011, 39, 6568–6584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamang, S.; Kishkevich, A.; Morrow, C.A.; Osman, F.; Jalan, M.; Whitby, M.C. The PCNA unloader Elg1 promotes recombination at collapsed replication forks in fission yeast. Elife 2019, 8, e47277. [Google Scholar] [CrossRef]
- Watson, A.T.; Werler, P.; Carr, A.M. Regulation of gene expression at the fission yeast Schizosaccharomyces pombe urg1 locus. Gene 2011, 484, 76–86. [Google Scholar] [CrossRef]
- Forsburg, S.L. Comparison of Schizosaccharomyces-Pombe Expression Systems. Nucleic Acids Res. 1993, 21, 2955–2956. [Google Scholar] [CrossRef] [Green Version]
- Maundrell, K. Thiamine-repressible expression vectors pREP and pRIP for fission yeast. Gene 1993, 123, 127–130. [Google Scholar] [CrossRef]
- Muller, S.; Sandal, T.; Kamp-Hansen, P.; Dalboge, H. Comparison of expression systems in the yeasts Saccharomyces cerevisiae, Hansenula polymorpha, Klyveromyces lactis, Schizosaccharomyces pombe and Yarrowia lipolytica. Cloning of two novel promoters from Yarrowia lipolytica. Yeast 1998, 14, 1267–1283. [Google Scholar] [CrossRef]
- Watson, A.T.; Daigaku, Y.; Mohebi, S.; Etheridge, T.J.; Chahwan, C.; Murray, J.M.; Carr, A.M. Optimisation of the Schizosaccharomyces pombe urg1 Expression System. PLoS ONE 2013, 8, e83800. [Google Scholar] [CrossRef] [Green Version]
- Sunder, S.; Greeson-Lott, N.T.; Runge, K.W.; Sanders, S.L. A new method to efficiently induce a site-specific double-strand break in the fission yeast Schizosaccharomyces pombe. Yeast 2012, 29, 275–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zilio, N.; Wehrkamp-Richter, S.; Boddy, M.N. A new versatile system for rapid control of gene expression in the fission yeast Schizosaccharomyces pombe. Yeast 2012, 29, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Lucas, B.E.; McPherson, M.T.; Hawk, T.M.; Wilson, L.N.; Kroh, J.M.; Hickman, K.G.; Fitzgerald, S.R.; Disbennett, W.M.; Rollins, P.D.; Hylton, H.M.; et al. An Assay to Study Intra-Chromosomal Deletions in Yeast. Methods Protoc. 2019, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, S.; Watson, A.; Sheedy, D.M.; Martin, B.; Carr, A.M. Gross chromosomal rearrangements and elevated recombination at an inducible site-specific replication fork barrier. Cell 2005, 121, 689–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saada, A.A.; Teixeira-Silva, A.; Iraqui, I.; Costes, A.; Hardy, J.; Paoletti, G.; Freon, K.; Lambert, S.A.E. Unprotected Replication Forks Are Converted into Mitotic Sister Chromatid Bridges. Mol. Cell 2017, 66, 398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno, K.; Lambert, S.; Baldacci, G.; Murray, J.M.; Carr, A.M. Nearby inverted repeats fuse to generate acentric and dicentric palindromic chromosomes by a replication template exchange mechanism. Gene Dev. 2009, 23, 2876–2886. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, K.; Miyabe, I.; Schalbetter, S.A.; Carr, A.M.; Murray, J.M. Recombination-restarted replication makes inverted chromosome fusions at inverted repeats. Nature 2013, 493, 246–249. [Google Scholar] [CrossRef] [Green Version]
- Mohebi, S.; Mizuno, K.; Watson, A.; Carr, A.M.; Murray, J.M. Checkpoints are blind to replication restart and recombination intermediates that result in gross chromosomal rearrangements. Nat. Commun. 2015, 6, 6357. [Google Scholar] [CrossRef] [Green Version]
- Iraqui, I.; Chekkal, Y.; Jmari, N.; Pietrobon, V.; Freon, K.; Costes, A.; Lambert, S.A.E. Recovery of Arrested Replication Forks by Homologous Recombination Is Error-Prone. Plos Genetics 2012, 8, e1002976. [Google Scholar] [CrossRef] [Green Version]
- Lambert, S.; Mizuno, K.; Blaisonneau, J.; Martineau, S.; Chanet, R.; Freon, K.; Murray, J.M.; Carr, A.M.; Baldacci, G. Homologous Recombination Restarts Blocked Replication Forks at the Expense of Genome Rearrangements by Template Exchange. Mol. Cell 2010, 39, 346–359. [Google Scholar] [CrossRef]
- Teixeira-Silva, A.; Ait Saada, A.; Hardy, J.; Iraqui, I.; Nocente, M.C.; Freon, K.; Lambert, S.A.E. The end-joining factor Ku acts in the end-resection of double strand break-free arrested replication forks. Nat. Commun. 2017, 8, 1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arcangioli, B.; Roseaulin, L.; Holmes, A. Mating-Type Switching in S. Pombe. In Molecular Genetics of Recombination; Springer: Berlin, Germany, 2007; pp. 95–133. [Google Scholar]
- Leupold, U. Studies on recombination in Schizosaccharomyces pombe. Cold Spring Harb. Symp. Quant. Biol. 1958, 23, 161–170. [Google Scholar] [CrossRef]
- Egel, R. Genes involved in mating type expression of fission yeast. Mol. Gen. Genet. 1973, 122, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Egel, R.; Gutz, H. Gene activation by copy transposition in mating-type switching of a homothallic fission yeast. Curr. Genet. 1981, 3, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Egel, R.; Beach, D.H.; Klar, A.J. Genes required for initiation and resolution steps of mating-type switching in fission yeast. Proc. Natl Acad. Sci. USA 1984, 81, 3481–3485. [Google Scholar] [CrossRef] [Green Version]
- Egel, R. Two tightly linked silent cassettes in the mating-type region of Schizosaccharomyces pombe. Curr. Genet. 1984, 8, 199–203. [Google Scholar] [CrossRef]
- Beach, D.H.; Klar, A.J. Rearrangements of the transposable mating-type cassettes of fission yeast. EMBO J. 1984, 3, 603–610. [Google Scholar] [CrossRef]
- Klar, A.J. Lessons learned from studies of fission yeast mating-type switching and silencing. Annu. Rev. Genet. 2007, 41, 213–236. [Google Scholar] [CrossRef]
- Thon, G.; Maki, T.; Haber, J.E.; Iwasaki, H. Mating-type switching by homology-directed recombinational repair: A matter of choice. Curr. Genet. 2019, 65, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Hanson, S.J.; Wolfe, K.H. An Evolutionary Perspective on Yeast Mating-Type Switching. Genetics 2017, 206, 9–32. [Google Scholar] [CrossRef] [Green Version]
- Gutz, H.; Doe, F.J. Two Different h Mating Types in schizosaccharomyces pombe. Genetics 1973, 74, 563–569. [Google Scholar] [PubMed]
- Egel, R. Frequency of mating-type switching in homothallic fission yeast. Nature 1977, 266, 172–174. [Google Scholar] [CrossRef]
- Lee, C.S.; Haber, J.E. Mating-type Gene Switching in Saccharomyces cerevisiae. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef]
- Vengrova, S.; Dalgaard, J.Z. RNase-sensitive DNA modification(s) initiates S. pombe mating-type switching. Gene Dev. 2004, 18, 794–804. [Google Scholar] [CrossRef] [Green Version]
- Vengrova, S.; Dalgaard, J.Z. The wild-type Schizosaccharomyces pombe mat1 imprint consists of two ribonucleotides. EMBO Rep. 2006, 7, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, B.; Bisht, K.K.; Upadhyay, U.; Kushwaha, A.C.; Nanda, J.S.; Srivastava, S.; Saini, J.K.; Klar, A.J.S.; Singh, J. Role of Cdc23/Mcm10 in generating the ribonucleotide imprint at the mat1 locus in fission yeast. Nucleic Acids Res. 2019, 47, 3422–3433. [Google Scholar] [CrossRef] [PubMed]
- Kaykov, A.; Holmes, A.M.; Arcangioli, B. Formation, maintenance and consequences of the imprint at the mating-type locus in fission yeast. EMBO J. 2004, 23, 930–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalgaard, L.Z.; Klar, A.J.S. Orientation of DNA replication establishes mating-type switching pattern in S-pombe. Nature 1999, 400, 181–184. [Google Scholar] [CrossRef]
- Arcangioli, B. A site- and strand-specific DNA break confers asymmetric switching potential in fission yeast. EMBO J. 1998, 17, 4503–4510. [Google Scholar] [CrossRef] [Green Version]
- Arcangioli, B.; de Lahondes, R. Fission yeast switches mating type by a replication-recombination coupled process. Embo J. 2000, 19, 1389–1396. [Google Scholar] [CrossRef] [Green Version]
- Holmes, A.M.; Kaykov, A.; Arcangioli, B. Molecular and cellular dissection of mating-type switching steps in Schizosaccharomyces pombe. Mol. Cell. Biol. 2005, 25, 303–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vengrova, S.; Codlin, S.; Dalgaard, J.Z. RTS1—an eukaryotic terminator of replication. Int. J. Biochem. Cell. B 2002, 34, 1031–1034. [Google Scholar] [CrossRef]
- Klar, A.J.S.; Ishikawa, K.; Moore, S. A Unique DNA Recombination Mechanism of the Mating/Cell-type Switching of Fission Yeasts: A Review. Microbiol. Spectr. 2014, 2, 5. [Google Scholar] [CrossRef]
- Roseaulin, L.; Yamada, Y.; Tsutsui, Y.; Russell, P.; Iwasaki, H.; Arcangioli, B. Mus81 is essential for sister chromatid recombination at broken replication forks. EMBO J. 2008, 27, 1378–1387. [Google Scholar] [CrossRef] [Green Version]
- Styrkarsdottir, U.; Egel, R.; Nielsen, O. The smt-0 mutation which abolishes mating-type switching in fission yeast is a deletion. Curr. Genet. 1993, 23, 184–186. [Google Scholar] [CrossRef]
- Klar, A.J.; Miglio, L.M. Initiation of meiotic recombination by double-strand DNA breaks in S. pombe. Cell 1986, 46, 725–731. [Google Scholar] [CrossRef]
- Zhu, M.; Zhao, H.; Limbo, O.; Russell, P. Mre11 complex links sister chromatids to promote repair of a collapsed replication fork. Proc. Natl. Acad. Sci. USA 2018, 115, 8793–8798. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Zhu, M.; Limbo, O.; Russell, P. RNase H eliminates R-loops that disrupt DNA replication but is nonessential for efficient DSB repair. EMBO Rep. 2018, 19. [Google Scholar] [CrossRef]
- Gutz, H.; Schmidt, H. Switching genes in Schizosaccharomyces pombe. Curr. Genet. 1985, 9, 325–331. [Google Scholar] [CrossRef]
- Inagawa, T.; Yamada-Inagawa, T.; Eydmann, T.; Mian, I.S.; Wang, T.S.; Dalgaard, J.Z. Schizosaccharomyces pombe Rtf2 mediates site-specific replication termination by inhibiting replication restart. Proc. Natl. Acad. Sci. USA 2009, 106, 7927–7932. [Google Scholar] [CrossRef] [Green Version]
- Allshire, R.C.; Ekwall, K. Epigenetic Regulation of Chromatin States in Schizosaccharomyces pombe. Cold Spring Harb. Perspect. Biol. 2015, 7, a018770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volpe, T.A.; Kidner, C.; Hall, I.M.; Teng, G.; Grewal, S.I.; Martienssen, R.A. Regulation of heterochromatic silencing and histone H3 lysine-9 methylation by RNAi. Science 2002, 297, 1833–1837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, S.; Noma, K.; Grewal, S.I. RNAi-independent heterochromatin nucleation by the stress-activated ATF/CREB family proteins. Science 2004, 304, 1971–1976. [Google Scholar] [CrossRef] [PubMed]
- Verdel, A.; Jia, S.; Gerber, S.; Sugiyama, T.; Gygi, S.; Grewal, S.I.; Moazed, D. RNAi-mediated targeting of heterochromatin by the RITS complex. Science 2004, 303, 672–676. [Google Scholar] [CrossRef] [Green Version]
- Thon, G.; Klar, A.J. The clr1 locus regulates the expression of the cryptic mating-type loci of fission yeast. Genetics 1992, 131, 287–296. [Google Scholar]
- Ekwall, K.; Ruusala, T. Mutations in rik1, clr2, clr3 and clr4 genes asymmetrically derepress the silent mating-type loci in fission yeast. Genetics 1994, 136, 53–64. [Google Scholar]
- Thon, G.; Cohen, A.; Klar, A.J. Three additional linkage groups that repress transcription and meiotic recombination in the mating-type region of Schizosaccharomyces pombe. Genetics 1994, 138, 29–38. [Google Scholar]
- Nabeshima, K.; Nakagawa, T.; Straight, A.F.; Murray, A.; Chikashige, Y.; Yamashita, Y.M.; Hiraoka, Y.; Yanagida, M. Dynamics of centromeres during metaphase-anaphase transition in fission yeast: Dis1 is implicated in force balance in metaphase bipolar spindle. Mol. Biol. Cell 1998, 9, 3211–3225. [Google Scholar] [CrossRef] [Green Version]
- Du, L.L.; Nakamura, T.M.; Moser, B.A.; Russell, P. Retention but not recruitment of Crb2 at double-strand breaks requires Rad1 and Rad3 complexes. Mol. Cell. Biol. 2003, 23, 6150–6158. [Google Scholar] [CrossRef] [Green Version]
- Du, L.L.; Nakamura, T.M.; Russell, P. Histone modification-dependent and -independent pathways for recruitment of checkpoint protein Crb2 to double-strand breaks. Genes Dev. 2006, 20, 1583–1596. [Google Scholar] [CrossRef] [Green Version]
- Sofueva, S.; Du, L.L.; Limbo, O.; Williams, J.S.; Russell, P. BRCT domain interactions with phospho-histone H2A target Crb2 to chromatin at double-strand breaks and maintain the DNA damage checkpoint. Mol. Cell. Biol. 2010, 30, 4732–4743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, T.M.; Moser, B.A.; Du, L.L.; Russell, P. Cooperative control of Crb2 by ATM family and Cdc2 kinases is essential for the DNA damage checkpoint in fission yeast. Mol. Cell. Biol. 2005, 25, 10721–10730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limbo, O.; Chahwan, C.; Yamada, Y.; de Bruin, R.A.M.; Wittenberg, C.; Russell, P. Ctp1 is a cell-cycle-regulated protein that functions with Mre11 complex to control double-strand break repair by homologous recombination. Mol. Cell 2007, 28, 134–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, R.S.; Moncalian, G.; Williams, J.S.; Yamada, Y.; Limbo, O.; Shin, D.S.; Groocock, L.M.; Cahill, D.; Hitomi, C.; Guenther, G.; et al. Mre11 dimers coordinate DNA end bridging and nuclease processing in double-strand-break repair. Cell 2008, 135, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limbo, O.; Porter-Goff, M.E.; Rhind, N.; Russell, P. Mre11 Nuclease Activity and Ctp1 Regulate Chk1 Activation by Rad3(ATR) and Tel1(ATM) Checkpoint Kinases at Double-Strand Breaks. Mol. Cell. Biol. 2011, 31, 573–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limbo, O.; Moiani, D.; Kertokalio, A.; Wyman, C.; Tainer, J.A.; Russell, P. Mre11 ATLD17/18 mutation retains Tel1/ATM activity but blocks DNA double-strand break repair. Nucleic Acids Res. 2012, 40, 11435–11449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Wang, H.T.; Zhai, Y.; Russell, P.; Du, L.L. Mdb1, a Fission Yeast Homolog of Human MDC1, Modulates DNA Damage Response and Mitotic Spindle Function. PLoS ONE 2014, 9, e97028. [Google Scholar] [CrossRef]
- Limbo, O.; Yamada, Y.; Russell, P. Mre11-Rad50-dependent activity of ATM/Tel1 at DNA breaks and telomeres in the absence of Nbs1. Mol. Biol. Cell 2018, 29, 1389–1399. [Google Scholar] [CrossRef]
- Langerak, P.; Mejia-Ramirez, E.; Limbo, O.; Russell, P. Release of Ku and MRN from DNA Ends by Mre11 Nuclease Activity and Ctp1 Is Required for Homologous Recombination Repair of Double-Strand Breaks. PLoS Genet. 2011, 7, e1002271. [Google Scholar] [CrossRef] [Green Version]
- Zierhut, C.; Diffley, J.F.X. Break dosage, cell cycle stage and DNA replication influence DNA double strand break response. EMBO J. 2008, 27, 1875–1885. [Google Scholar] [CrossRef] [Green Version]
- Leland, B.A.; King, M.C. Using LacO Arrays to Monitor DNA Double-Strand Break Dynamics in Live Schizosaccharomyces pombe Cells. In Cancer Genomics and Proteomics: Methods and Protocols, 2nd ed.; Humana Press: New York, NY, USA, 2014; Volume 1176, pp. 127–141. [Google Scholar]
- Yu, Y.; Ren, J.Y.; Zhang, J.M.; Suo, F.; Fang, X.F.; Wu, F.; Du, L.L. A proteome-wide visual screen identifies fission yeast proteins localizing to DNA double-strand breaks. DNA Repair 2013, 12, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, A.; Arai, R.; Yashiroda, Y.; Shirai, A.; Kamata, A.; Sekido, S.; Kobayashi, Y.; Hashimoto, A.; Hamamoto, M.; Hiraoka, Y.; et al. ORFeome cloning and global analysis of protein localization in the fission yeast Schizosaccharomyces pombe. Nat. Biotechnol. 2006, 24, 841–847. [Google Scholar] [CrossRef]
- Goedecke, W.; Pfeiffer, P.; Vielmetter, W. Nonhomologous DNA end joining in Schizosaccharomyces pombe efficiently eliminates DNA double-strand-breaks from haploid sequences. Nucleic Acids Res. 1994, 22, 2094–2101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manolis, K.G.; Nimmo, E.R.; Hartsuiker, E.; Carr, A.M.; Jeggo, P.A.; Allshire, R.C. Novel functional requirements for non-homologous DNA end joining in Schizosaccharomyces pombe. EMBO J. 2001, 20, 210–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulton, S.J.; Jackson, S.P. Saccharomyces cerevisiae Ku70 potentiates illegitimate DNA double-strand break repair and serves as a barrier to error-prone DNA repair pathways. EMBO J. 1996, 15, 5093–5103. [Google Scholar] [CrossRef] [PubMed]
- Hentges, P.; Ahnesorg, P.; Pitcher, R.S.; Bruce, C.K.; Kysela, B.; Green, A.J.; Bianchi, J.; Wilson, T.E.; Jackson, S.P.; Doherty, A.J. Evolutionary and functional conservation of the DNA non-homologous end-joining protein, XLF/cernunnos. J. Biol. Chem. 2006, 281, 37517–37526. [Google Scholar] [CrossRef] [Green Version]
- Decottignies, A. Capture of extranuclear DNA at fission yeast double-strand breaks. Genetics 2005, 171, 1535–1548. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Yu, Y.; Suo, F.; Sun, L.L.; Zhao, D.; Du, L.L. Genome-wide Screens for Sensitivity to Ionizing Radiation Identify the Fission Yeast Nonhomologous End Joining Factor Xrc4. G3-Genes Genom Genet. 2014, 4, 1297–1306. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Li, J.; Li, M.; Dou, K.; Zhang, M.J.; Suo, F.; Du, L.L. Multiple end joining mechanisms repair a chromosomal DNA break in fission yeast. DNA Repair 2012, 11, 120–130. [Google Scholar] [CrossRef]
- Li, Y.; Wang, J.; Zhou, G.; Lajeunesse, M.; Le, N.; Stawicki, B.N.; Corcino, Y.L.; Berkner, K.L.; Runge, K.W. Nonhomologous End-Joining with Minimal Sequence Loss Is Promoted by the Mre11-Rad50-Nbs1-Ctp1 Complex in Schizosaccharomyces pombe. Genetics 2017, 206, 481–496. [Google Scholar] [CrossRef] [Green Version]
- Almeida, H.; Godinho Ferreira, M. Spontaneous telomere to telomere fusions occur in unperturbed fission yeast cells. Nucleic Acids Res. 2013, 41, 3056–3067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van, H.T.; Santos, M.A. Histone modifications and the DNA double-strand break response. Cell Cycle 2018, 17, 2399–2410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bader, A.S.; Hawley, B.R.; Wilczynska, A.; Bushell, M. The roles of RNA in DNA double-strand break repair. Br. J. Cancer 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hylton, H.M.; Lucas, B.E.; Petreaca, R.C. Schizosaccharomyces pombe Assays to Study Mitotic Recombination Outcomes. Genes 2020, 11, 79. https://doi.org/10.3390/genes11010079
Hylton HM, Lucas BE, Petreaca RC. Schizosaccharomyces pombe Assays to Study Mitotic Recombination Outcomes. Genes. 2020; 11(1):79. https://doi.org/10.3390/genes11010079
Chicago/Turabian StyleHylton, Hannah M., Bailey E. Lucas, and Ruben C. Petreaca. 2020. "Schizosaccharomyces pombe Assays to Study Mitotic Recombination Outcomes" Genes 11, no. 1: 79. https://doi.org/10.3390/genes11010079
APA StyleHylton, H. M., Lucas, B. E., & Petreaca, R. C. (2020). Schizosaccharomyces pombe Assays to Study Mitotic Recombination Outcomes. Genes, 11(1), 79. https://doi.org/10.3390/genes11010079