Role of PB1 Midbody Remnant Creating Tethered Polar Bodies during Meiosis II


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin of the Animals
2.2. Microinjection, Imaging and Reagents
2.3. Synthesis of RNAs
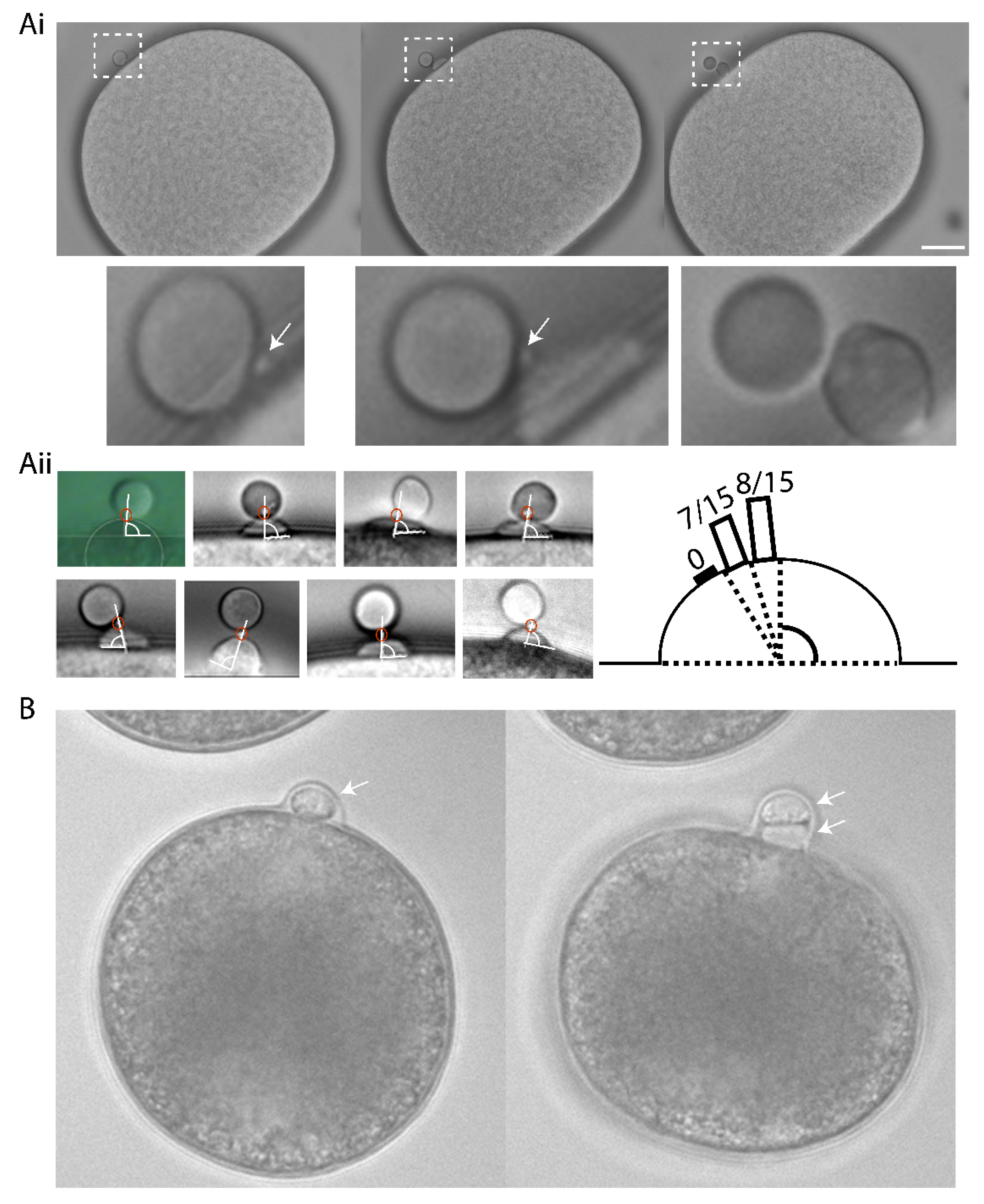
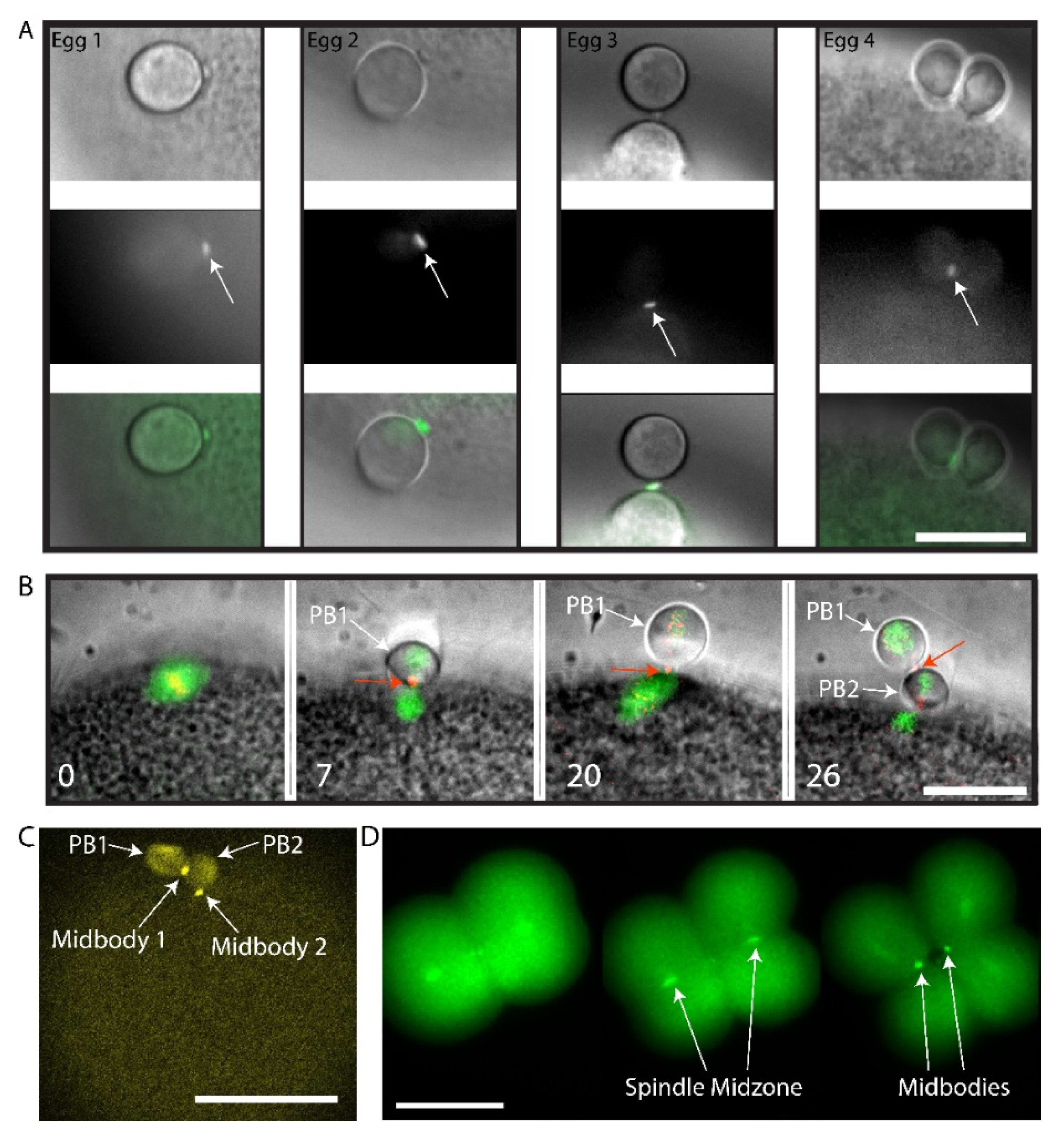
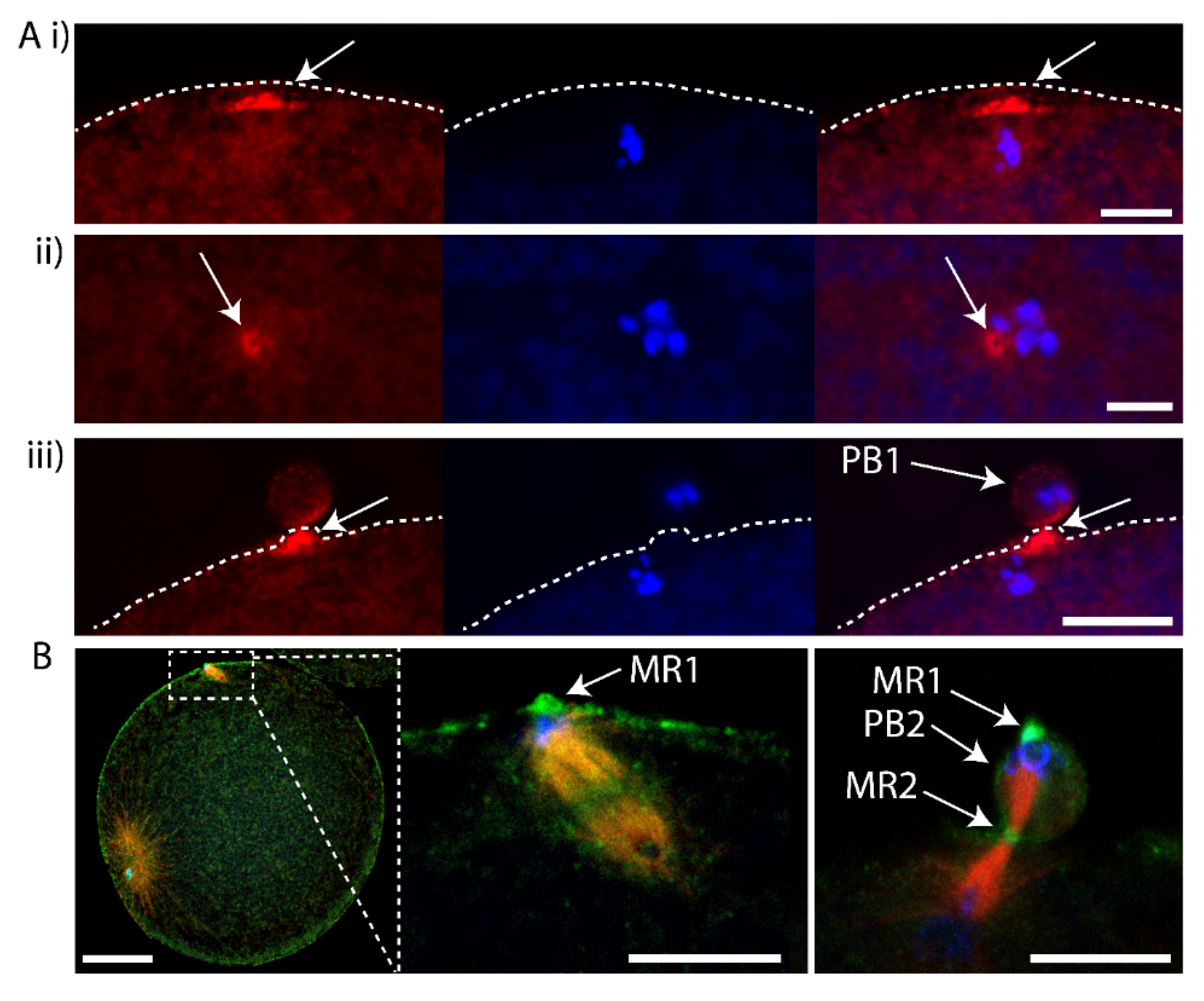
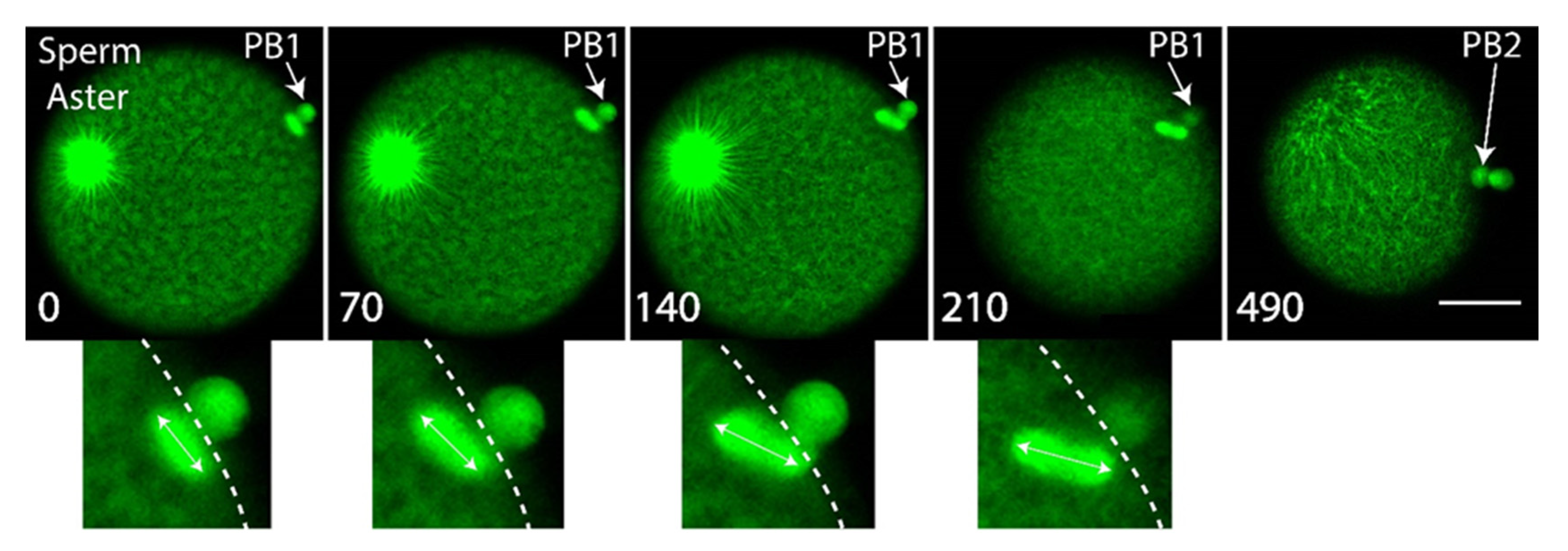
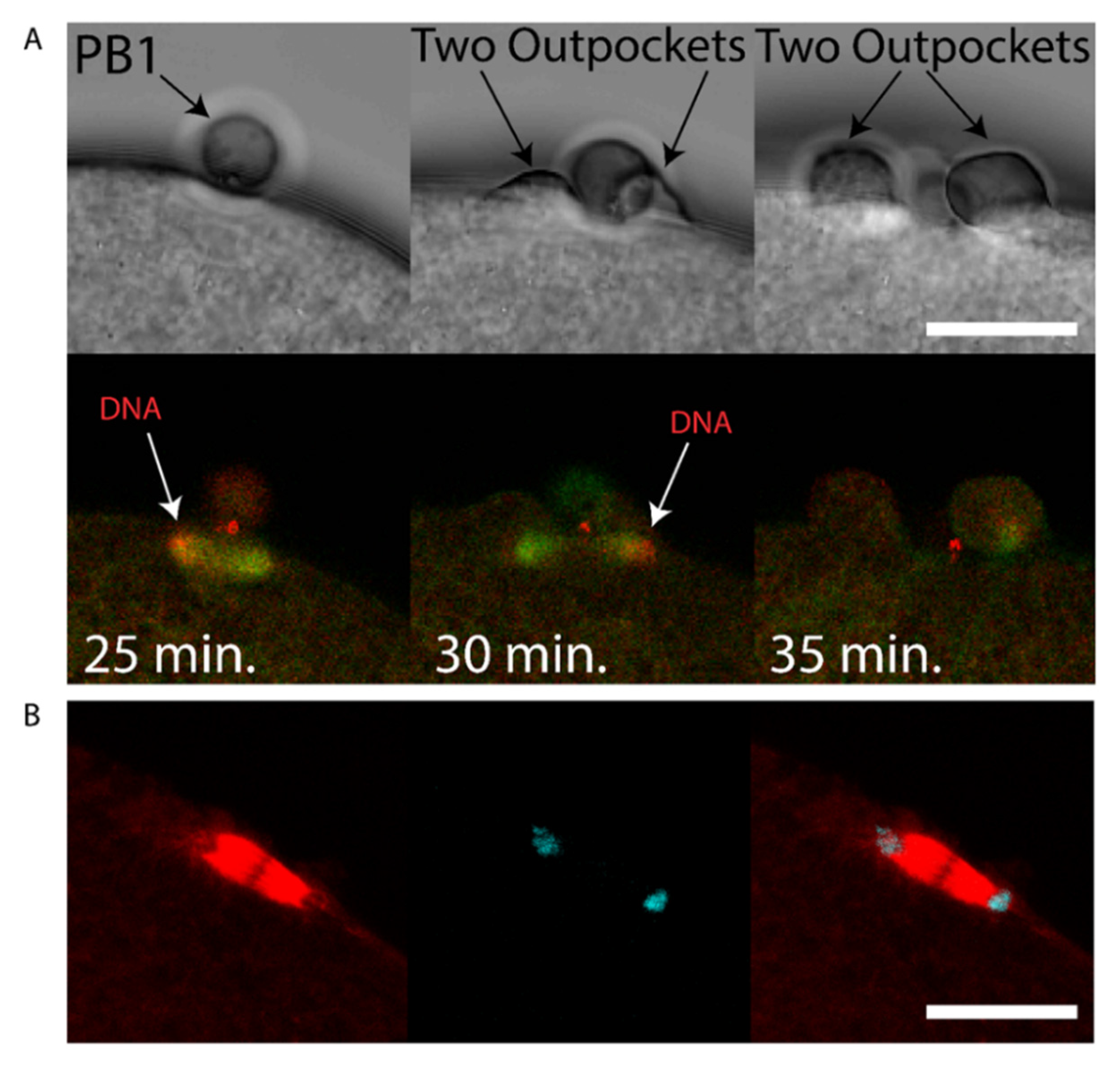
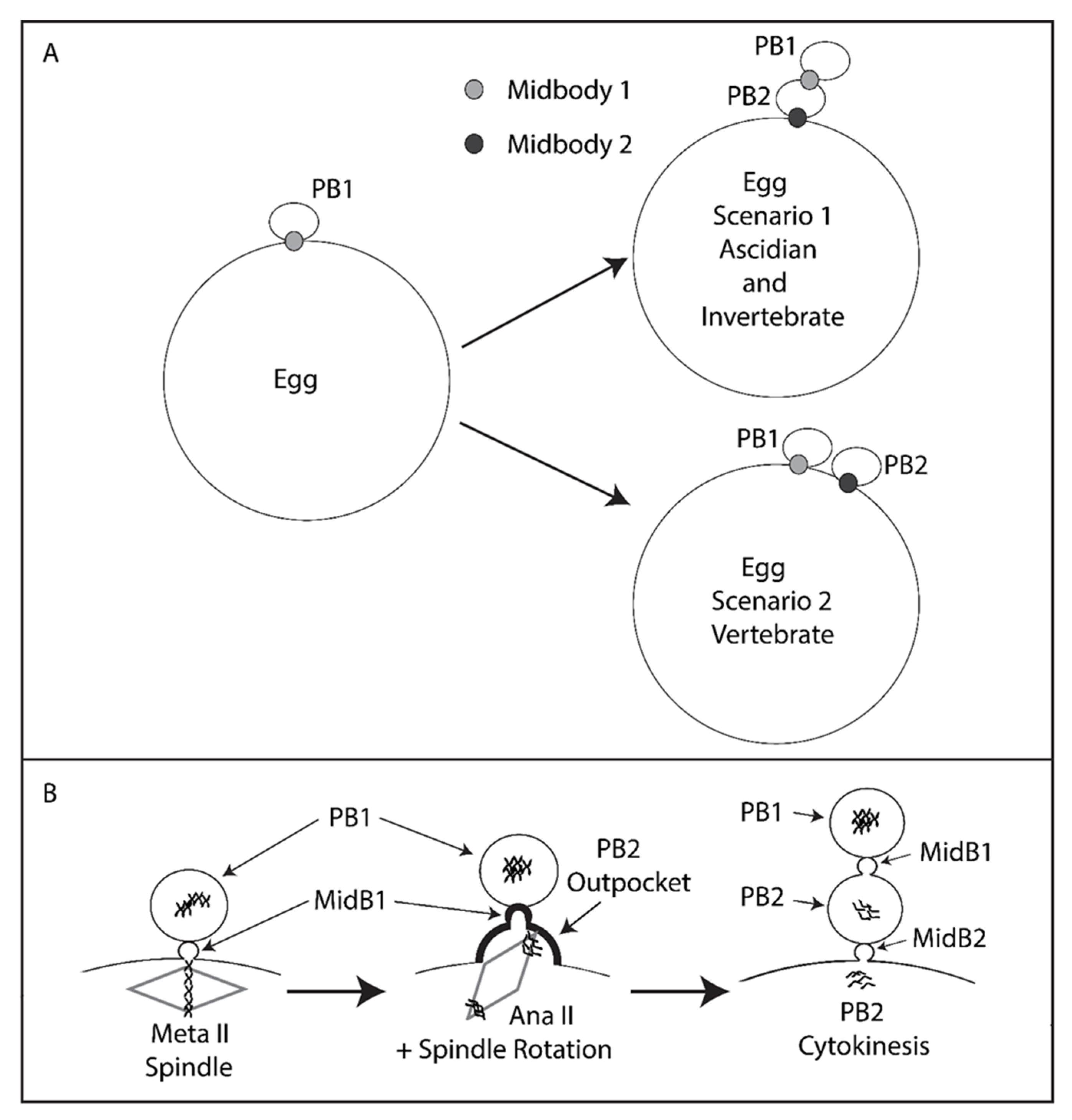
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Azoury, J.; Lee, K.W.; Georget, V.; Rassinier, P.; Leader, B.; Verlhac, M.-H. Spindle Positioning in Mouse Oocytes Relies on a Dynamic Meshwork of Actin Filaments. Curr. Biol. 2008, 18, 1514–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prodon, F.; Chenevert, J.; Sardet, C. Establishment of animal–vegetal polarity during maturation in ascidian oocytes. Dev. Biol. 2006, 290, 297–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuh, M.; Ellenberg, J. A New Model for Asymmetric Spindle Positioning in Mouse Oocytes. Curr. Biol. 2008, 18, 1986–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eager, D.D.; Johnson, M.H.; Thurley, K.W. Ultrastructural studies on the surface membrane of the mouse egg. J. Cell Sci. 1976, 22, 345–353. [Google Scholar]
- Maro, B.; Johnson, M.H.; Pickering, S.J.; Flach, G. Changes in actin distribution during fertilization of the mouse egg. J. Embryol. Exp. Morphol. 1984, 81, 211–237. [Google Scholar]
- Maro, B.; Johnson, M.H.; Webb, M.; Flach, G. Mechanism of polar body formation in the mouse oocyte: An interaction between the chromosomes, the cytoskeleton and the plasma membrane. J. Embryol. Exp. Morphol. 1986, 92, 11–32. [Google Scholar]
- Deng, M.; Suraneni, P.; Schultz, R.M.; Li, R. The Ran GTPase Mediates Chromatin Signaling to Control Cortical Polarity during Polar Body Extrusion in Mouse Oocytes. Dev. Cell 2007, 12, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Dumont, J.; Petri, S.; Pellegrin, F.; Terret, M.-E.; Bohnsack, M.T.; Rassinier, P.; Georget, V.; Kalab, P.; Gruss, O.J.; Verlhac, M.-H. A centriole- and RanGTP-independent spindle assembly pathway in meiosis I of vertebrate oocytes. J. Cell Biol. 2007, 176, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Heald, R.; Tournebize, R.; Blank, T.A.; Sandaltzopoulos, R.; Becker, P.B.; A Hyman, A.; Karsenti, E. Self-organization of microtubules into bipolar spindles around artificial chromosomes in Xenopus egg extracts. Nat. Cell Biol. 1996, 382, 420–425. [Google Scholar] [CrossRef]
- Karsenti, E. The Mitotic Spindle: A Self-Made Machine. Science 2001, 294, 543–547. [Google Scholar] [CrossRef]
- Karsenti, E.; Newport, J.; Kirschner, M. Respective roles of centrosomes and chromatin in the conversion of microtubule arrays from interphase to metaphase. J. Cell Biol. 1984, 99, 47s–54s. [Google Scholar] [CrossRef]
- Dehapiot, B.; Halet, G. Ran GTPase promotes oocyte polarization by regulating ERM (Ezrin/Radixin/Moesin) inactivation. Cell Cycle 2013, 12, 1672–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Benink, H.A.; Cheng, D.; Montplaisir, V.; Wang, L.; Xi, Y.; Zheng, P.-P.; Bement, W.M.; Liu, X.J. Cdc42 activation couples spindle positioning to first polar body formation in oocyte maturation. Curr. Biol. 2006, 16, 214–220. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ma, C.; Miller, A.L.; Katbi, H.A.; Bement, W.M.; Liu, X.J. Polar Body Emission Requires a RhoA Contractile Ring and Cdc42-Mediated Membrane Protrusion. Dev. Cell 2008, 15, 386–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehapiot, B.; Carrière, V.; Carroll, J.; Halet, G. Polarized Cdc42 activation promotes polar body protrusion and asymmetric division in mouse oocytes. Dev. Biol. 2013, 377, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-B.; Jiang, Z.-Z.; Zhang, Q.-H.; Hu, M.-W.; Huang, L.; Ou, X.-H.; Guo, L.; Ouyang, Y.-C.; Hou, Y.; Brakebusch, C.; et al. Specific deletion of Cdc42 does not affect meiotic spindle organization/migration and homologous chromosome segregation but disrupts polarity establishment and cytokinesis in mouse oocytes. Mol. Biol. Cell 2013, 24, 3832–3841. [Google Scholar] [CrossRef] [PubMed]
- Maddox, A.S.; Azoury, J.; Dumont, J. Polar body cytokinesis. Cytoskeleton 2012, 69, 855–868. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, J.; Zhang, X.; McKee, D.; Wang, Z.-B.; Li, R.; Ma, C.; Sun, Q.-Y.; Liu, X.J. The small GTPase Cdc42 promotes membrane protrusion during polar body emission via ARP2-nucleated actin polymerization. Mol. Hum. Reprod. 2011, 17, 305–316. [Google Scholar] [CrossRef] [Green Version]
- Sardet, C.; McDougall, A.; Yasuo, H.; Chenevert, J.; Pruliere, G.; Dumollard, R.; Hudson, C.; Hebras, C.; Le Nguyen, N.; Paix, A. Embryological Methods in Ascidians: The Villefranche-sur-Mer Protocols. Adv. Struct. Saf. Stud. 2011, 770, 365–400. [Google Scholar] [CrossRef]
- McDougall, A.; Chenevert, J.; Prulière, G.; Costache, V.; Hebras, C.; Salez, G.; Dumollard, R. Centrosomes and spindles in ascidian embryos and eggs. Methods Cell Biol. 2015, 129, 317–339. [Google Scholar] [CrossRef]
- Costache, V.; Hebras, C.; Pruliere, G.; Besnardeau, L.; Failla, M.; Copley, R.R.; Burgess, D.; Chenevert, J.; McDougall, A. Kif2 localizes to a subdomain of cortical endoplasmic reticulum that drives asymmetric spindle position. Nat. Commun. 2017, 8, 917. [Google Scholar] [CrossRef] [PubMed]
- McDougall, A.; Sardet, C. Function and characteristics of repetitive calcium waves associated with meiosis. Curr. Biol. 1995, 5, 318–328. [Google Scholar] [CrossRef] [Green Version]
- Amiel, A.; Lechable, M.; Robert, L.; Chevalier, S.; Houliston, E. Conserved Functions for Mos in Eumetazoan Oocyte Maturation Revealed by Studies in a Cnidarian. Curr. Biol. 2009, 19, 305–311. [Google Scholar] [CrossRef] [Green Version]
- Maslakova, S.A. Development to metamorphosis of the nemertean pilidium larva. Front. Zool. 2010, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Goddard, J.H.R. Unusually Large Polar Bodies in an Aeolid Nudibranch: A Novel Mechanism for Producing Extra-Embryonic Yolk Reserves. J. Molluscan Stud. 1991, 57, 143–152. [Google Scholar] [CrossRef]
- Harada, K.; Oita, E.; Chiba, K. Metaphase I arrest of starfish oocytes induced via the MAP kinase pathway is released by an increase of intracellular pH. Development 2003, 130, 4581–4586. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, A.; Kato, K.H.; Nemoto, S.-I. Role of microtubules and centrosomes in the eccentric relocation of the germinal vesicle upon meiosis reinitiation in sea-cucumber oocytes. Dev. Biol. 2005, 280, 237–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorn, J.F.; Zhang, L.; Paradis, V.; Edoh-Bedi, D.; Jusu, S.; Maddox, P.S.; Maddox, A.S. Actomyosin Tube Formation in Polar Body Cytokinesis Requires Anillin in C. elegans. Curr. Biol. 2010, 20, 2046–2051. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, J.M.; Golden, A.; Donovan, P.J. AIR-2: An Aurora/Ipl1-related Protein Kinase Associated with Chromosomes and Midbody Microtubules Is Required for Polar Body Extrusion and Cytokinesis in Caenorhabditis elegans Embryos. J. Cell Biol. 1998, 143, 1635–1646. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Leblanc, J.; He, K.; Liu, X.J. Spindle function in Xenopus oocytes involves possible nanodomain calcium signaling. Mol. Biol. Cell. 2016, 27, 3273–3283. [Google Scholar] [CrossRef]
- Hornick, J.E.; Karanjeet, K.; Collins, E.S.; Hinchcliffe, E.H. Kinesins to the core: The role of microtubule-based motor proteins in building the mitotic spindle midzone. Semin. Cell Dev. Biol. 2010, 21, 290–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.-C.; Liu, H.-L.; Sun, Q.-Y. Survivin regulates Plk1 localization to kinetochore in mouse oocyte meiosis. Biochem. Biophys. Res. Commun. 2012, 421, 797–800. [Google Scholar] [CrossRef]
- Wianny, F.; Tavares, Á.; Evans, M.J.; Glover, D.M.; Zernicka-Goetz, M. Mouse polo-like kinase 1 associates with the acentriolar spindle poles, meiotic chromosomes and spindle midzone during oocyte maturation. Chromosoma 1998, 107, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-T.; Ettinger, A.W.; Huttner, W.B.; Doxsey, S.J. Resurrecting remnants: The lives of post-mitotic midbodies. Trends Cell Biol. 2013, 23, 118–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, D.; Pohl, C. Coupling of Rotational Cortical Flow, Asymmetric Midbody Positioning, and Spindle Rotation Mediates Dorsoventral Axis Formation in C. elegans. Dev. Cell 2014, 28, 253–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowell, E.F.; Gaffuri, A.-L.; Gayraud-Morel, B.; Tajbakhsh, S.; Echard, A. Engulfment of the midbody remnant after cytokinesis in mammalian cells. J. Cell Sci. 2014, 127, 3840–3851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertzler, P.L. Twin meiosis 2 spindles form after suppression of polar body 1 formation in oocytes of the marine shrimp Sicyonia ingentis. Biol. Bull. 2002, 202, 100–103. [Google Scholar] [CrossRef]
- Crowder, M.E.; Strzelecka, M.; Wilbur, J.D.; Good, M.C.; Von Dassow, G.; Heald, R. A Comparative Analysis of Spindle Morphometrics across Metazoans. Curr. Biol. 2015, 25, 1542–1550. [Google Scholar] [CrossRef] [Green Version]
- Herbert, M.; Levasseur, M.; Homer, H.A.; Yallop, K.; Murdoch, A.; McDougall, A. Homologue disjunction in mouse oocytes requires proteolysis of securin and cyclin B1. Nat. Cell Biol. 2003, 5, 1023–1025. [Google Scholar] [CrossRef]
- Kudo, N.R.; Wassmann, K.; Anger, M.; Schuh, M.; Wirth, K.G.; Xu, H.; Helmhart, W.; Kudo, H.; McKay, M.; Maro, B.; et al. Resolution of Chiasmata in Oocytes Requires Separase-Mediated Proteolysis. Cell 2006, 126, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Homer, H.A.; McDougall, A.; Levasseur, M.; Yallop, K.; Murdoch, A.P.; Herbert, M. Mad2 prevents aneuploidy and premature proteolysis of cyclin B and securin during meiosis I in mouse oocytes. Genes Dev. 2005, 19, 202–207. [Google Scholar] [CrossRef] [Green Version]
- Deng, M.; Li, R. Sperm Chromatin-Induced Ectopic Polar Body Extrusion in Mouse Eggs after ICSI and Delayed Egg Activation. PLoS ONE 2009, 4, e7171. [Google Scholar] [CrossRef] [Green Version]
- Zenker, J.; White, M.D.; Templin, R.M.; Parton, R.G.; Thorn-Seshold, O.; Bissiere, S.; Plachta, N. A microtubule-organizing center directing intracellular transport in the early mouse embryo. Science 2017, 357, 925–928. [Google Scholar] [CrossRef] [Green Version]
- Crowder, M.E.; Flynn, J.R.; McNally, K.P.; Cortes, D.B.; Price, K.L.; Kuehnert, P.A.; Panzica, M.T.; Andaya, A.; Leary, J.A.; McNally, F.J. Dynactin-dependent cortical dynein and spherical spindle shape correlate temporally with meiotic spindle rotation in Caenorhabditis elegans. Mol. Biol. Cell 2015, 26, 3030–3046. [Google Scholar] [CrossRef]
- Gard, D.L. Microtubule organization during maturation of Xenopus oocytes: Assembly and rotation of the meiotic spindles. Dev. Biol. 1992, 151, 516–530. [Google Scholar] [CrossRef]
- Chiou, J.; Balasubramanian, M.K.; Lew, D.J. Cell Polarity in Yeast. Annu. Rev. Cell Dev. Biol. 2017, 33, 77–101. [Google Scholar] [CrossRef]
- Komaru, A.; Ookubo, K.; Kiyomoto, M. All meiotic chromosomes and both centrosomes at spindle pole in the zygotes discarded as two polar bodies in clam Corbicula leana: Unusual polar body formation observed by antitubulin immunofluorescence. Dev. Genes Evol. 2000, 210, 263–269. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | PBs Tethered | Publication |
|---|---|---|
| Jellyfish Clytia hemispherica | Yes | [23] |
| Nemertean worms Micura alaskensis | Yes | [24] |
| Sea slug Cuthona lagunae | Yes | [25] |
| Clam * Acila castrensis* | Yes | Von Dassow * Center for Cell Dynamics website * |
| Mussel Mytilus galloprovincialis | Yes | This study |
| Starfish Asterina pectinifera | Yes | [26] |
| Sea cucumber Holothuria moebi | Yes | [27] |
| Ascidian Phallusia mammillata | Yes | This study |
| Nematode Caenorhabditis elegans | Unclear | [28,29] |
| Xenopus | No | [30] |
| Mouse | No | [15] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McDougall, A.; Hebras, C.; Pruliere, G.; Burgess, D.; Costache, V.; Dumollard, R.; Chenevert, J. Role of PB1 Midbody Remnant Creating Tethered Polar Bodies during Meiosis II. Genes 2020, 11, 1394. https://doi.org/10.3390/genes11121394
McDougall A, Hebras C, Pruliere G, Burgess D, Costache V, Dumollard R, Chenevert J. Role of PB1 Midbody Remnant Creating Tethered Polar Bodies during Meiosis II. Genes. 2020; 11(12):1394. https://doi.org/10.3390/genes11121394
Chicago/Turabian StyleMcDougall, Alex, Celine Hebras, Gerard Pruliere, David Burgess, Vlad Costache, Remi Dumollard, and Janet Chenevert. 2020. "Role of PB1 Midbody Remnant Creating Tethered Polar Bodies during Meiosis II" Genes 11, no. 12: 1394. https://doi.org/10.3390/genes11121394
APA StyleMcDougall, A., Hebras, C., Pruliere, G., Burgess, D., Costache, V., Dumollard, R., & Chenevert, J. (2020). Role of PB1 Midbody Remnant Creating Tethered Polar Bodies during Meiosis II. Genes, 11(12), 1394. https://doi.org/10.3390/genes11121394