A CNTNAP1 Missense Variant Is Associated with Canine Laryngeal Paralysis and Polyneuropathy

 , , ,
, , ,  , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animal Selection
2.3. Sample Preparation
2.4. Axonal Size Frequency Distributions and G-Ratios
2.5. Single Nucleotide Polymorphism Array Genotyping and Imputation
2.6. Genome-Wide Association Study and Fine-Mapping
2.7. Whole-Genome Sequencing
2.8. Targeted Genotyping
2.9. Protein Predictions
2.10. Availability of Data and Material
3. Results
3.1. Phenotype
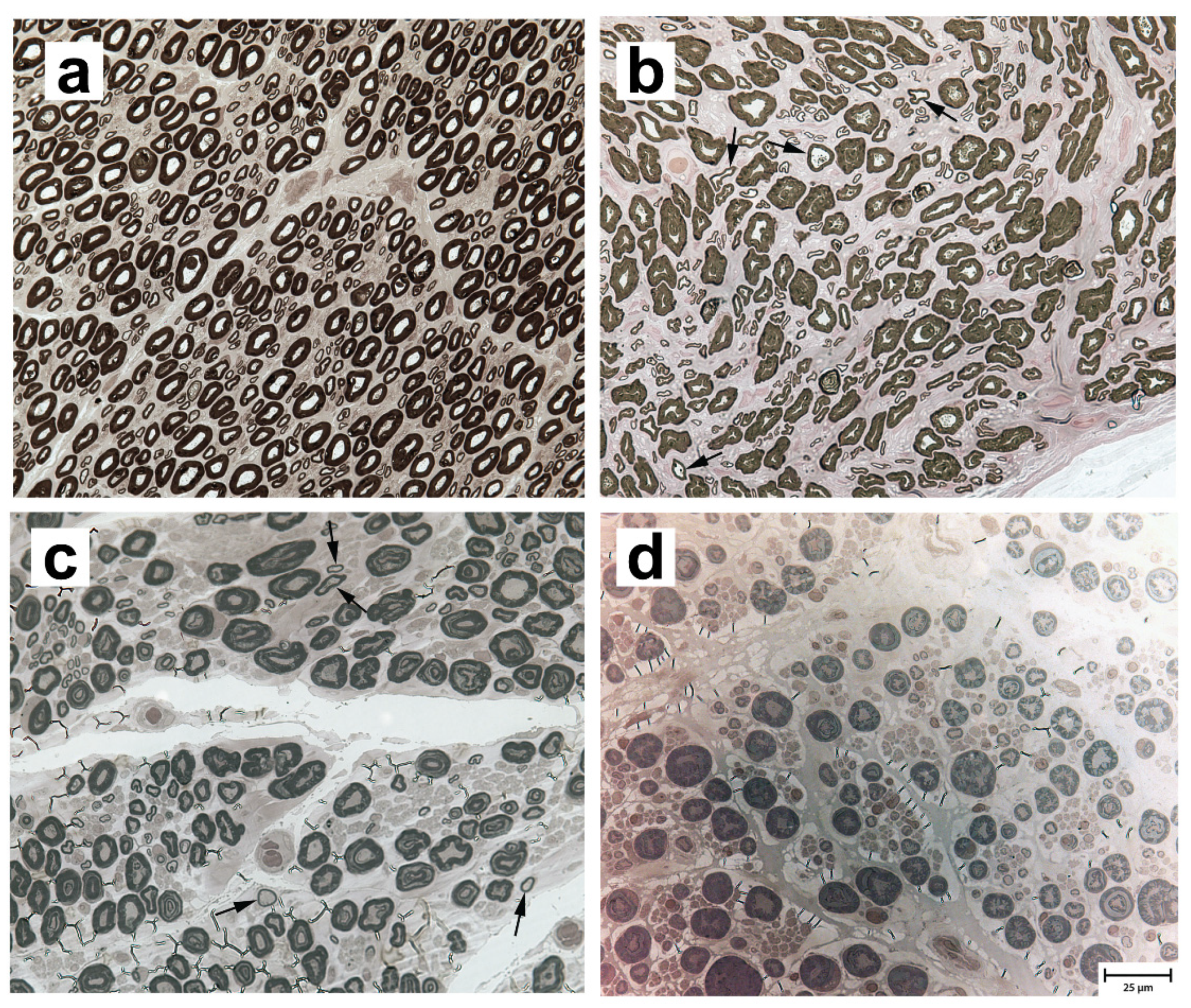
3.2. Neuropathological and Morphometric Findings
3.3. Genome-Wide Association Study and Fine-Mapping
3.4. Identification of the Candidate Causative Variant
3.5. The CNTNAP1 Variant Occurs in Several Breeds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kitshoff, A.M.; Van Goethem, B.; Stegen, L.; Vandekerckhov, P.; De Rooster, H. Laryngeal paralysis in dogs: An update on recent knowledge. J. S. Afr. Vet. Assoc. 2013, 84, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Monnet, E. Surgical Treatment of Laryngeal Paralysis. Vet. Clin. N. Am. Small Anim. Pract. 2016, 46, 709–717. [Google Scholar] [CrossRef]
- Mackin, G.A. Diagnosis of patients with peripheral nerve disease. Clin. Podiatr. Med. Surg. 1994, 11, 545–569. [Google Scholar] [PubMed]
- Gabriel, A.; Poncelet, L.; Van Ham, L.; Clercx, C.; Braund, K.G.; Bhatti, S.; Detilleux, J.; Peeters, D. Laryngeal paralysis-polyneuropathy complex in young related Pyrenean mountain dogs. J. Small Anim. Pract. 2006, 47, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Granger, N. Canine inherited motor and sensory neuropathies: An updated classification in 22 breeds and comparison to Charcot-Marie-Tooth disease. Vet. J. 2011, 188, 274–285. [Google Scholar] [CrossRef] [PubMed]
- GOLPP|College of Veterinary Medicine at MSU. Available online: https://cvm.msu.edu/scs/research-initiatives/golpp (accessed on 7 September 2020).
- Letko, A.; Minor, K.M.; Jagannathan, V.; Seefried, F.R.; Mickelson, J.R.; Oliehoek, P.; Drögemüller, C. Genomic diversity and population structure of the Leonberger dog breed. Genet. Sel. Evol. 2020, 52, 61. [Google Scholar] [CrossRef]
- Ekenstedt, K.J.; Becker, D.; Minor, K.M.; Shelton, G.D.; Patterson, E.E.; Bley, T.; Oevermann, A.; Bilzer, T.; Leeb, T.; Drögemüller, C.; et al. An ARHGEF10 Deletion Is Highly Associated with a Juvenile-Onset Inherited Polyneuropathy in Leonberger and Saint Bernard Dogs. PLoS Genet. 2014, 10, e1004635. [Google Scholar] [CrossRef] [Green Version]
- Becker, D.; Minor, K.M.; Letko, A.; Ekenstedt, K.J.; Jagannathan, V.; Leeb, T.; Shelton, G.D.; Mickelson, J.R.; Drögemüller, C. A GJA9 frameshift variant is associated with polyneuropathy in Leonberger dogs. BMC Genom. 2017, 18, 662. [Google Scholar] [CrossRef] [Green Version]
- Wiedmer, M.; Oevermann, A.; Borer-Germann, S.E.; Gorgas, D.; Shelton, G.D.; Drögemüller, M.; Jagannathan, V.; Henke, D.; Leeb, T. A RAB3GAP1 SINE Insertion in Alaskan Huskies with Polyneuropathy, Ocular Abnormalities, and Neuronal Vacuolation (POANV) Resembling Human Warburg Micro Syndrome 1 (WARBM1). G3 Genes Genomes Genet. 2016, 6, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Mhlanga-Mutangadura, T.; Johnson, G.S.; Schnabel, R.D.; Taylor, J.F.; Johnson, G.C.; Katz, M.L.; Shelton, G.D.; Lever, T.E.; Giuliano, E.; Granger, N.; et al. A mutation in the Warburg syndrome gene, RAB3GAP1, causes a similar syndrome with polyneuropathy and neuronal vacuolation in Black Russian Terrier dogs. Neurobiol. Dis. 2016, 86, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Mhlanga-Mutangadura, T.; Johnson, G.S.; Ashwini, A.; Shelton, G.D.; Wennogle, S.A.; Johnson, G.C.; Kuroki, K.; O’Brien, D.P. A Homozygous RAB3GAP1:c.743delC Mutation in Rottweilers with Neuronal Vacuolation and Spinocerebellar Degeneration. J. Vet. Intern. Med. 2016, 30, 813–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasouliha, S.H.; Barrientos, L.; Anderegg, L.; Klesty, C.; Lorenz, J.; Chevallier, L.; Jagannathan, V.; Rösch, S.; Leeb, T. A RAPGEF6 variant constitutes a major risk factor for laryngeal paralysis in dogs. PLoS Genet. 2019, 15, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Minor, K.M.; Letko, A.; Becker, D.; Drögemüller, M.; Mandigers, P.J.J.; Bellekom, S.R.; Leegwater, P.A.J.; Stassen, Q.E.M.; Putschbach, K.; Fischer, A.; et al. Canine NAPEPLD-associated models of human myelin disorders. Sci. Rep. 2018, 8, 5818. [Google Scholar] [CrossRef] [PubMed]
- Parker, H.G.; Dreger, D.L.; Rimbault, M.; Davis, B.W.; Mullen, A.B.; Carpintero-Ramirez, G.; Ostrander, E.A. Genomic Analyses Reveal the Influence of Geographic Origin, Migration, and Hybridization on Modern Dog Breed Development. Cell Rep. 2017, 19, 697–708. [Google Scholar] [CrossRef]
- Huang, M.; Hayward, J.J.; Corey, E.; Garrison, S.J.; Wagner, G.R.; Krotscheck, U.; Hayashi, K.; Schweitzer, P.A.; Lust, G.; Boyko, A.R.; et al. A novel iterative mixed model to remap three complex orthopedic traits in dogs. PLoS ONE 2017, 12, e0176932. [Google Scholar] [CrossRef]
- Browning, S.R.; Browning, B.L. Rapid and Accurate Haplotype Phasing and Missing-Data Inference for Whole-Genome Association Studies by Use of Localized Haplotype Clustering. Am. J. Hum. Genet. 2007, 81, 1084–1097. [Google Scholar] [CrossRef] [Green Version]
- Browning, B.L.; Browning, S.R. Genotype Imputation with Millions of Reference Samples. Am. J. Hum. Genet. 2016, 98, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Browning, B. Conform-gt Program. Available online: https://faculty.washington.edu/browning/conform-gt.html (accessed on 22 September 2020).
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.A.M.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. Gigascience 2015, 4, 7. [Google Scholar] [CrossRef]
- Zhou, X.; Stephens, M. Genome-wide efficient mixed-model analysis for association studies. Nat. Genet. 2012, 44, 821–824. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Turner, S.D.; Turner, D.S. qqman: An R package for visualizing GWAS results using Q-Q and manhattan plots. bioRxiv 2014, 81, 559–575. [Google Scholar] [CrossRef]
- Jagannathan, V.; Drögemüller, C.; Leeb, T. Dog Biomedical Variant Database Consortium, (DBVDC) A comprehensive biomedical variant catalogue based on whole genome sequences of 582 dogs and eight wolves. Anim. Genet. 2019, 50, 695–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pejaver, V.; Urresti, J.; Lugo-Martinez, J.; Pagel, K.A.; Lin, G.N.; Nam, H.-J.; Mort, M.; Cooper, D.N.; Sebat, J.; Iakoucheva, L.M.; et al. MutPred2: Inferring the molecular and phenotypic impact of amino acid variants. bioRxiv 2017, 134981. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.; Chan, A.P. PROVEAN web server: A tool to predict the functional effect of amino acid substitutions and indels. Bioinformatics 2015, 31, 2745–2747. [Google Scholar] [CrossRef] [Green Version]
- Niroula, A.; Urolagin, S.; Vihinen, M. PON-P2: Prediction method for fast and reliable identification of harmful variants. PLoS ONE 2015, 10, e117380. [Google Scholar] [CrossRef] [Green Version]
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alföldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef]
- Low, K.; Stals, K.; Caswell, R.; Clayton-Smith, J.; Donaldson, A.; Foulds, N.; Splitt, M.; Norman, A.; Urankar, K.; Vijayakumar, K.; et al. CNTNAP1: Extending the phenotype of congenital hypomyelinating neuropathy in 6 further patients. Neuromuscul. Disord. 2017, 27, S148. [Google Scholar] [CrossRef]
- Freed, A.S.; Weiss, M.D.; Malouf, E.A.; Hisama, F.M. CNTNAP1 mutations in an adult with Charcot Marie Tooth disease. Muscle Nerve 2019, 60, E28–E30. [Google Scholar] [CrossRef]
- Sabbagh, S.; Antoun, S.; Mégarbané, A. CNTNAP1 Mutations and Their Clinical Presentations: New Case Report and Systematic Review. Case Rep. Med. 2020, 2020. [Google Scholar] [CrossRef] [Green Version]
- Low, K.; Stals, K.; Caswell, R.; Wakeling, M.; Clayton-Smith, J.; Donaldson, A.; Foulds, N.; Norman, A.; Splitt, M.; Urankar, K.; et al. Phenotype of CNTNAP1: A study of patients demonstrating a specific severe congenital hypomyelinating neuropathy with survival beyond infancy. Eur. J. Hum. Genet. 2018, 26, 796–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallat, J.-M.; Nizon, M.; Magee, A.; Isidor, B.; Magy, L.; Péréon, Y.; Richard, L.; Ouvrier, R.; Cogné, B.; Devaux, J.; et al. Contactin-Associated Protein 1 (CNTNAP1) Mutations Induce Characteristic Lesions of the Paranodal Region. J. Neuropathol. Exp. Neurol. 2016, 75, 1155–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meola, S.D. Brachycephalic Airway Syndrome. Top. Companion Anim. Med. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyko, A.R.; Brooks, S.A.; Behan-Braman, A.; Castelhano, M.; Corey, E.; Oliveira, K.C.; Swinburne, J.E.; Todhunter, R.J.; Zhang, Z.; Ainsworth, D.M.; et al. Genomic analysis establishes correlation between growth and laryngeal neuropathy in Thoroughbreds. BMC Genom. 2014, 15, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Cohort 1 | Dog Breed/Species | Total | Phenotype | ||
|---|---|---|---|---|---|
| LPPN-Affected 2 | LPPN Non-Affected 3 | Unknown | |||
| Discovery (n = 517) | Leonberger | 426 | 126 | 300 | 0 |
| Saint Bernard | 91 | 18 | 14 | 59 | |
| Validation (n = 1070) | Leonberger | 859 | 500 | 359 | 0 |
| Saint Bernard | 11 | 11 | 0 | 0 | |
| Labrador retriever 4 | 200 | 150 | 50 | 0 | |
| Population (n = 14,174) | 243 dog breeds | 13,798 | 0 | 0 | 13,798 |
| Unknown/mixed heritage | 314 | 0 | 0 | 314 | |
| Wolf | 58 | 0 | 0 | 58 | |
| Golden jackal | 2 | 0 | 0 | 2 | |
| Andean fox | 1 | 0 | 0 | 1 | |
| Dhole | 1 | 0 | 0 | 1 | |
| CNTNAP1 Genotypes | ||||
|---|---|---|---|---|
| Breed | LPPN Status | G/G | G/A | A/A 1 |
| Leonberger (n = 2738) | Affected (n = 434) 2 | 358 | 58 | 18 |
| LPN1/LPN2 (n = 192) 3 | 180 | 11 | 1 | |
| Non-affected (n = 659) 4 | 605 | 54 | 0 | |
| Population controls (n = 1453) | 1258 | 192 | 3 | |
| Saint Bernard (n = 305) | Affected (n = 24) 2 | 9 | 5 | 10 |
| LPN1 (n = 5) 3 | 2 | 3 | 0 | |
| Non-affected (n = 14) | 9 | 5 | 0 | |
| Population controls (n = 262) | 213 | 46 | 3 | |
| Labrador retriever (n = 1524) | Affected (n = 148) | 132 | 9 | 7 |
| Non-affected (n = 45) | 42 | 3 | 0 | |
| Population controls (n = 1331) | 1200 | 128 | 3 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Letko, A.; Minor, K.M.; Friedenberg, S.G.; Shelton, G.D.; Salvador, J.P.; Mandigers, P.J.J.; Leegwater, P.A.J.; Winkler, P.A.; Petersen-Jones, S.M.; Stanley, B.J.; et al. A CNTNAP1 Missense Variant Is Associated with Canine Laryngeal Paralysis and Polyneuropathy. Genes 2020, 11, 1426. https://doi.org/10.3390/genes11121426
Letko A, Minor KM, Friedenberg SG, Shelton GD, Salvador JP, Mandigers PJJ, Leegwater PAJ, Winkler PA, Petersen-Jones SM, Stanley BJ, et al. A CNTNAP1 Missense Variant Is Associated with Canine Laryngeal Paralysis and Polyneuropathy. Genes. 2020; 11(12):1426. https://doi.org/10.3390/genes11121426
Chicago/Turabian StyleLetko, Anna, Katie M. Minor, Steven G. Friedenberg, G. Diane Shelton, Jill Pesayco Salvador, Paul J. J. Mandigers, Peter A. J. Leegwater, Paige A. Winkler, Simon M. Petersen-Jones, Bryden J. Stanley, and et al. 2020. "A CNTNAP1 Missense Variant Is Associated with Canine Laryngeal Paralysis and Polyneuropathy" Genes 11, no. 12: 1426. https://doi.org/10.3390/genes11121426
APA StyleLetko, A., Minor, K. M., Friedenberg, S. G., Shelton, G. D., Salvador, J. P., Mandigers, P. J. J., Leegwater, P. A. J., Winkler, P. A., Petersen-Jones, S. M., Stanley, B. J., Ekenstedt, K. J., Johnson, G. S., Hansen, L., Jagannathan, V., Mickelson, J. R., & Drögemüller, C. (2020). A CNTNAP1 Missense Variant Is Associated with Canine Laryngeal Paralysis and Polyneuropathy. Genes, 11(12), 1426. https://doi.org/10.3390/genes11121426