Prolonged Glucocorticoid Exposure Does Not Accelerate Telomere Shortening in Cultured Human Fibroblasts




{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatments
2.2. DNA and RNA Extraction
2.3. qPCR for Telomere Length and Glucocorticoid-Responsive Gene Expression
2.4. Statistical Analysis
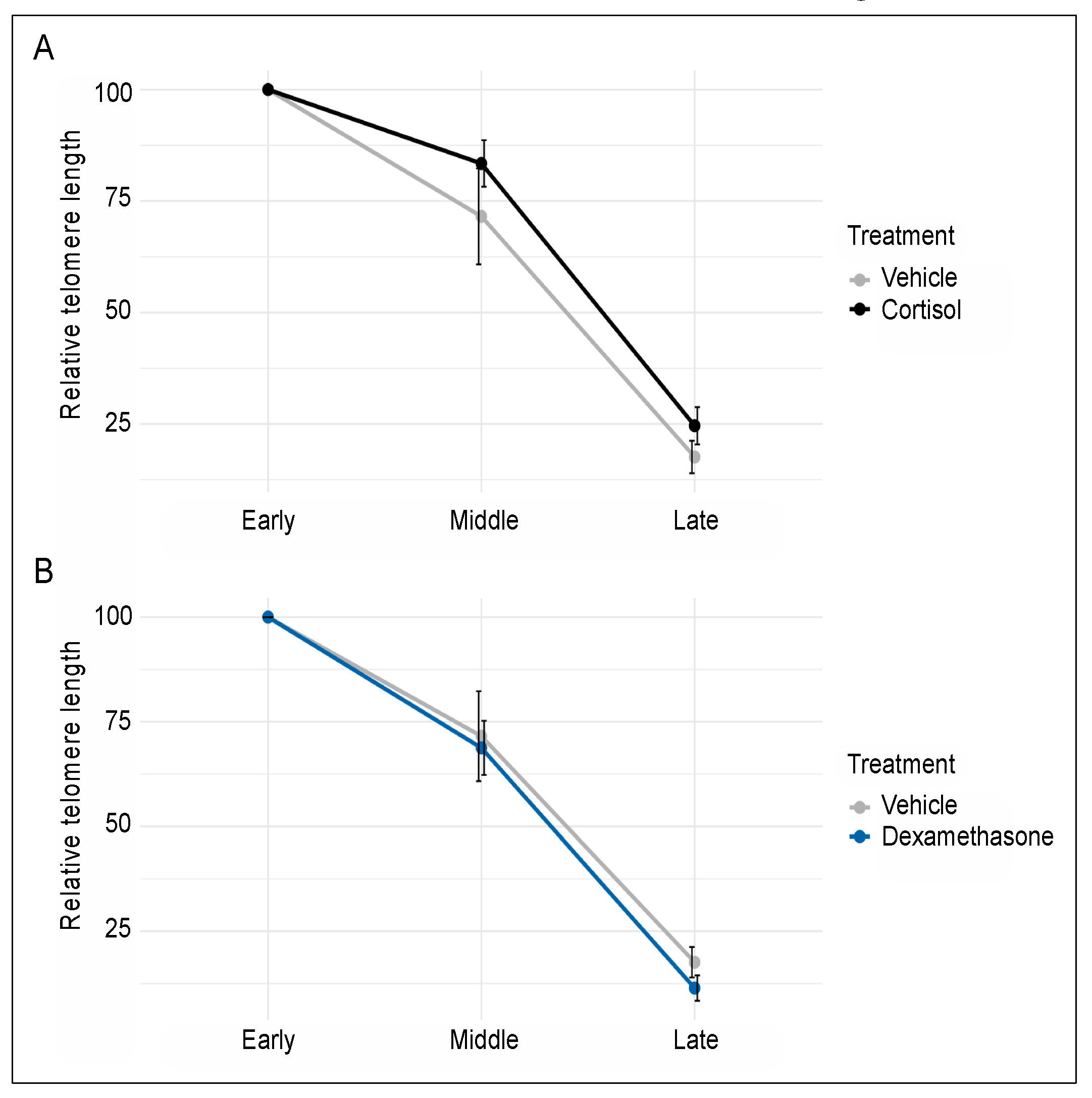
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zannas, A.S. Epigenetics as a Key Link between Psychosocial Stress and Aging: Concepts, Evidence, Mechanisms. Dialogues Clin. Neurosci. 2019, 21, 389–396. [Google Scholar] [PubMed]
- Epel, E.S.; Blackburn, E.H.; Lin, J.; Dhabhar, F.S.; Adler, N.E.; Morrow, J.D.; Cawthon, R.M. Accelerated Telomere Shortening in Response to Life Stress. Proc. Natl. Acad. Sci. USA 2004, 101, 17312–17315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Entringer, S.; Epel, E.S.; Kumsta, R.; Lin, J.; Hellhammer, D.H.; Blackburn, E.H.; Wust, S.; Wadhwa, P.D. Stress Exposure in Intrauterine Life is Associated with Shorter Telomere Length in Young Adulthood. Proc. Natl. Acad. Sci. USA 2011, 108, E513–E518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahola, K.; Siren, I.; Kivimaki, M.; Ripatti, S.; Aromaa, A.; Lonnqvist, J.; Hovatta, I. Work-Related Exhaustion and Telomere Length: A Population-Based Study. PLoS ONE 2012, 7, e40186. [Google Scholar] [CrossRef]
- Savolainen, K.; Eriksson, J.G.; Kananen, L.; Kajantie, E.; Pesonen, A.-K.; Heinonen, K.; Raikkonen, K. Associations between Early Life Stress, Self-Reported Traumatic Experiences Across the Lifespan and Leukocyte Telomere Length in Elderly Adults. Biol. Psychol. 2014, 97, 35–42. [Google Scholar] [CrossRef]
- Chrousos, G.P.; Gold, P.W. The Concepts of Stress and Stress System Disorders. Overview of Physical and Behavioral Homeostasis. JAMA 1992, 267, 1244–1252. [Google Scholar] [CrossRef]
- Zannas, A.S.; Chrousos, G.P. Epigenetic Programming by Stress and Glucocorticoids along the Human Lifespan. Mol. Psychiatry 2017, 22, 640–646. [Google Scholar] [CrossRef]
- Athanasoulia-Kaspar, A.P.; Auer, M.K.; Stalla, G.K.; Jakovcevski, M. Shorter Telomeres Associated with High Doses of Glucocorticoids: The Link to Increased Mortality? Endocr. Connect. 2018, 7, 1217–1226. [Google Scholar] [CrossRef] [Green Version]
- Casagrande, S.; Stier, A.; Monaghan, P.; Loveland, J.L.; Boner, W.; Lupi, S.; Trevisi, R.; Hau, M. Increased Glucocorticoid Concentrations in Early Life Cause Mitochondrial Inefficiency and Short Telomeres. J. Exp. Biol. 2020, 223 Pt 15, jeb222513. [Google Scholar] [CrossRef]
- Dantzer, B.; van Kesteteren, F.; Westrick, S.E.; Boutin, S.; McAdam, A.G.; Lane, J.E.; Gillespie, R.; Majer, A.; Haussmann, M.F.; Monaghan, P. Maternal Glucocorticoids Promote Offspring Growth without Inducing Oxidative Stress or Shortening Telomeres in Wild Red Squirrels. J. Exp. Biol. 2020, 223 Pt 1, jeb212373. [Google Scholar] [CrossRef]
- Lin, J.; Cheon, J.; Brown, R.; Coccia, M.; Puterman, K.; Sinclair, E.; Epel, E.; Blackburn, E.H. Systematic and Cell Type-Specific Telomere Length Changes in Subsets of Lymphocytes. J. Immunol. Res. 2016, 2016, 5371050. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.R.; Blackburn, E.H. Telomeres and Telomerase. Philos. Trans. R Soc. Lond. B Biol. Sci. 2004, 359, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-I.; Moon, S.-H.; Lee, W.-C.; Lee, H.-J.; Shivakumar, S.B.; Lee, S.-H.; Park, B.-W.; Rho, G.-J.; Jeon, B.-G. Inhibition of Cell Growth by Cellular Differentiation into Adipocyte-Like Cells in Dexamethasone Sensitive Cancer Cell Lines. Anim. Cells Syst. 2018, 22, 178–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, L.F.; Richardson, L.S.; Guimaraes da Silva, M.; Sheller-Miller, S.; Menon, R. Dexamethasone Induces Primary Amnion Epithelial Cell Senescence Through Telomere-P21 Associated Pathway. Biol. Reprod. 2019, 100, 1605–1616. [Google Scholar] [CrossRef]
- Britt-Compton, B.; Wyllie, F.; Rowson, J.; Capper, R.; Jones, R.E.; Baird, D.M. Telomere Dynamics during Replicative Senescence Are Not Directly Modulated by Conditions of Oxidative Stress in IMR90 Fibroblast Cells. Biogerontology 2009, 10, 683–693. [Google Scholar] [CrossRef]
- Laberge, R.-M.; Zhou, L.; Sarantos, M.R.; Rodier, F.; Freund, A.; De Keizer, P.L.J.; Liu, S.; DeMaria, M.; Cong, Y.-S.; Kapahi, P.; et al. Glucocorticoids Suppress Selected Components of the Senescence-Associated Secretory Phenotype. Aging Cell 2012, 11, 569–578. [Google Scholar] [CrossRef] [Green Version]
- Zannas, A.S.; Jia, M.; Hafner, K.; Baumert, J.; Weichmann, T.; Pape, J.C.; Arloth, J.; Kodel, M.; Martinelli, S.; Roitman, M.; et al. Epigenetic Upregulation of FKBP5 by Aging and Stress Contributes to NF-kappaB-Driven Inflammation and Cardiovascular Risk. Proc. Natl. Acad. Sci. USA 2019, 116, 11370–11379. [Google Scholar] [CrossRef] [Green Version]
- Zannas, A.S.; Wiechmann, T.; Gassen, N.C.; Binder, E.B. Gene-Stress-Epigenetic Regulation of FKBP5: Clinical and Translational Implications. Neuropsychopharmacology 2016, 41, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Cawthon, R.M. Telomere Measurement by Quantitative PCR. Nucleic Acids Res. 2002, 30, e47. [Google Scholar] [CrossRef]
- O’Callaghan, N.J.; Fenech, M. A Quantitative PCR Method for Measuring Absolute Telomere Length. Biol. Proced. Online 2011, 13, 3. [Google Scholar] [CrossRef] [Green Version]
- Wust, S.; Federenko, I.S.; Van Rossum, E.F.C.; Koper, J.W.; Hellhammer, D.H. Habituation of Cortisol Responses to Repeated Psychosocial Stress-Further Characterization and Impact of Genetic Factors. Psychoneuroendocrinology 2005, 30, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Bhake, R.C.; Leendertz, J.A.; Linthorst, A.C.E.; Lightman, S.L. Automated 24-Hours Sampling of Subcutaneous Tissue Free Cortisol in Humans. J. Med. Eng. Technol. 2013, 37, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Hamrahian, A.H.; Oseni, T.S.; Arafah, B.M. Measurements of Serum Free Cortisol in Critically Ill Patients. N. Engl. J. Med. 2004, 350, 1629–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Yi, W.; Suino-Powell, K.; Zhou, X.E.; Tolbert, W.D.; Tang, X.; Yang, J.; Yang, H.; Shi, J.; Hou, L.; et al. Structures and Mechanism for the Design of Highly Potent Glucocorticoids. Cell Res. 2014, 24, 713–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimanoe, C.; Hara, M.; Nishida, Y.; Nanri, H.; Horita, M.; Yamada, Y.; Li, Y.-S.; Kasai, H.; Kawai, K.; Higaki, Y.; et al. Perceived Stress, Depressive Symptoms, and Oxidative DNA Damage. Psychosom. Med. 2018, 80, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Fouquerel, E.; Barnes, R.P.; Uttam, S.; Watkins, S.C.; Bruchez, M.P.; Opresko, P.L. Targeted and Persistent 8-Oxoguanine Base Damage at Telomeres Promotes Telomere Loss and Crisis. Mol. Cell 2019, 75, 117–130. [Google Scholar] [CrossRef]
- Von Zglinicki, T. Oxidative Stress Shortens Telomeres. Trends Biochem. Sci. 2002, 27, 339–344. [Google Scholar] [CrossRef]
- Makpol, S.; Abidin, A.Z.; Sairin, K.; Mazlan, M.; Top, G.M.; Ngah, W.Z.W. Gamma-Tocotrienol Prevents Oxidative Stress-Induced Telomere Shortening in Human Fibroblasts Derived from Different Aged Individuals. Oxid. Med. Cell Longev. 2010, 3, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Casagrande, S.; Hau, M. Telomere Attrition: Metabolic Regulation and Signalling Function? Biol. Lett. 2019, 15, 20180885. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zannas, A.S.; Kosyk, O.; Leung, C.S. Prolonged Glucocorticoid Exposure Does Not Accelerate Telomere Shortening in Cultured Human Fibroblasts. Genes 2020, 11, 1425. https://doi.org/10.3390/genes11121425
Zannas AS, Kosyk O, Leung CS. Prolonged Glucocorticoid Exposure Does Not Accelerate Telomere Shortening in Cultured Human Fibroblasts. Genes. 2020; 11(12):1425. https://doi.org/10.3390/genes11121425
Chicago/Turabian StyleZannas, Anthony S., Oksana Kosyk, and Calvin S. Leung. 2020. "Prolonged Glucocorticoid Exposure Does Not Accelerate Telomere Shortening in Cultured Human Fibroblasts" Genes 11, no. 12: 1425. https://doi.org/10.3390/genes11121425
APA StyleZannas, A. S., Kosyk, O., & Leung, C. S. (2020). Prolonged Glucocorticoid Exposure Does Not Accelerate Telomere Shortening in Cultured Human Fibroblasts. Genes, 11(12), 1425. https://doi.org/10.3390/genes11121425