Development and Cell Cycle Activity of the Root Apical Meristem in the Fern Ceratopteris richardii

 ,
, 
 and
and 
Abstract
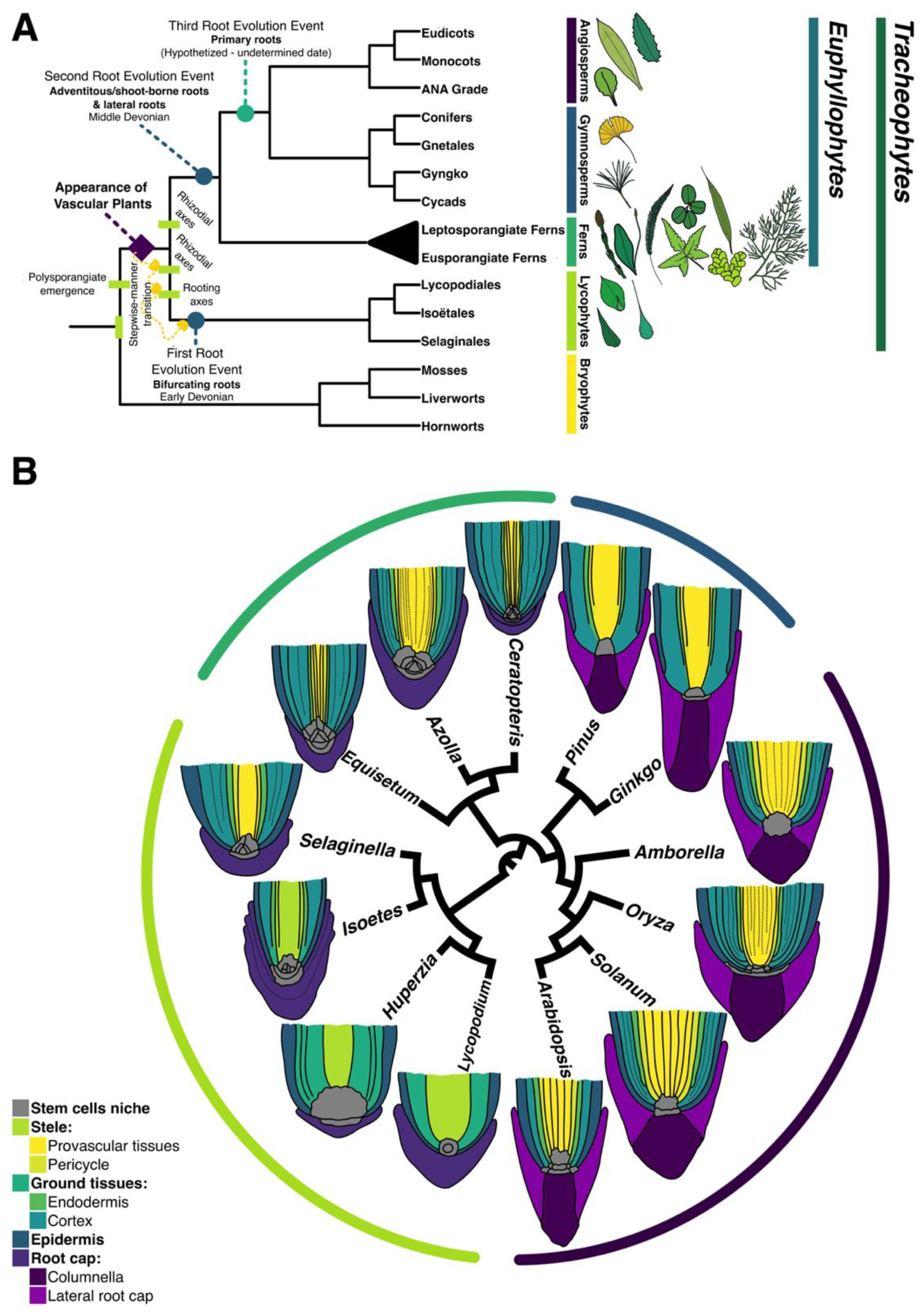
:1. Introduction
2. Materials and Methods
2.1. Plant Growth and Culture Conditions
2.2. Fixation and Clearing
2.3. Tissue Sections and Staining
2.4. Meristem Cell Size and Root Growth Analysis
2.5. EdU Incorporation Treatment
2.6. Microscopy and Image Analysis
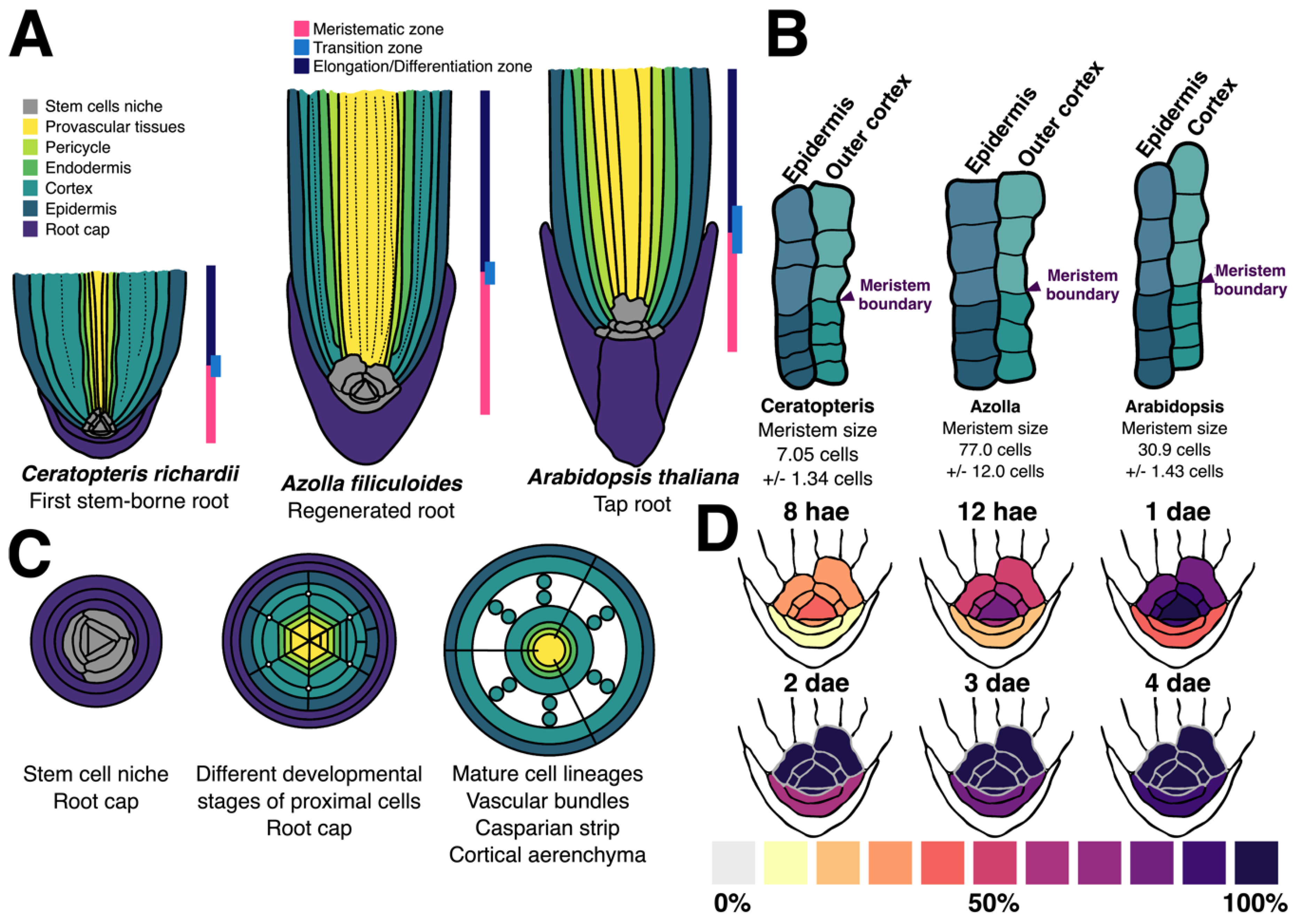
3. Results
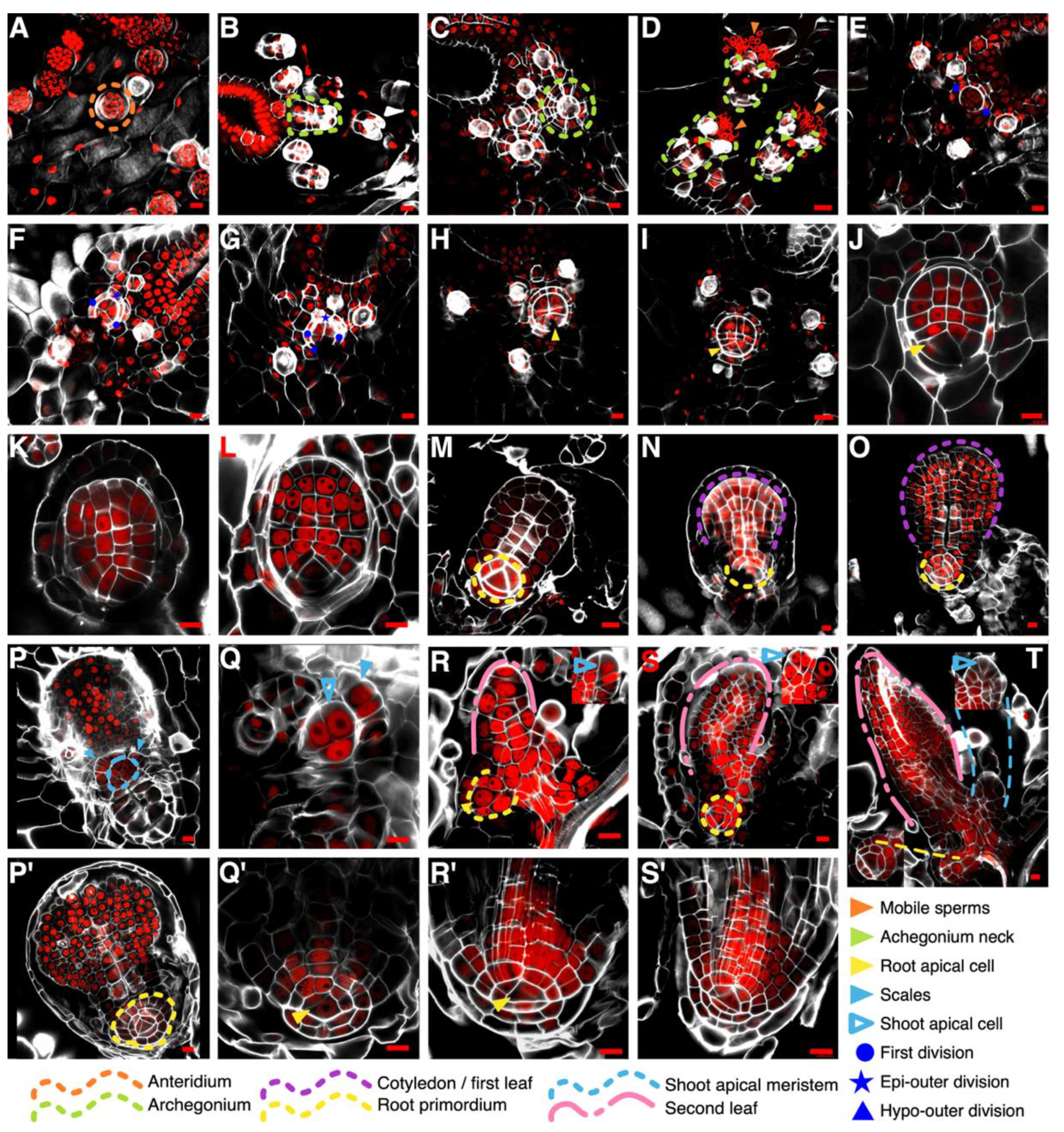
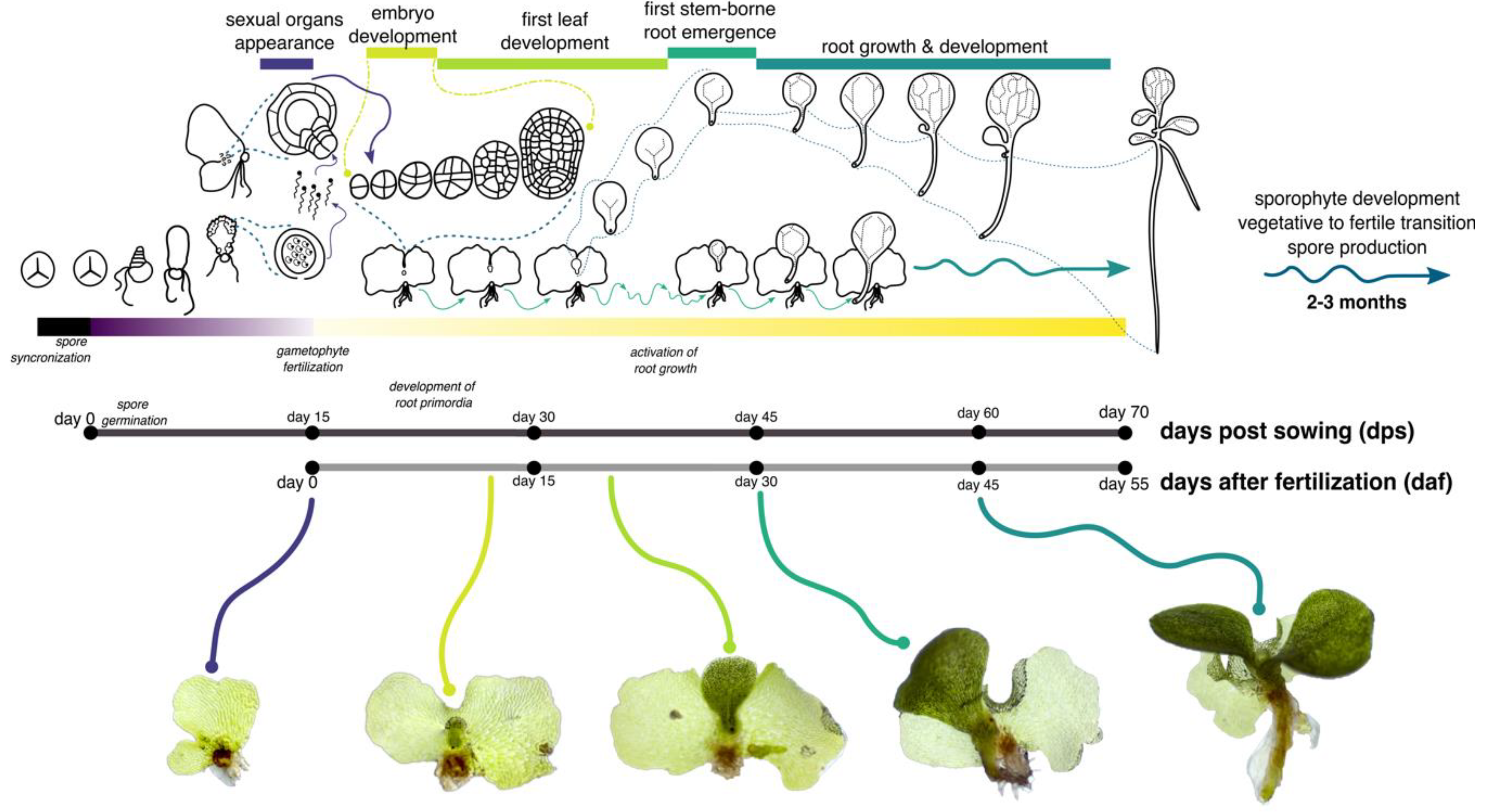
3.1. Establishment of the First Root during Embryonic Development in Ceratopteris Sporophyte
3.2. Zonation of the First Shoot-Borne Root
3.3. Histochemical Identification of the Different Root Cell Layers
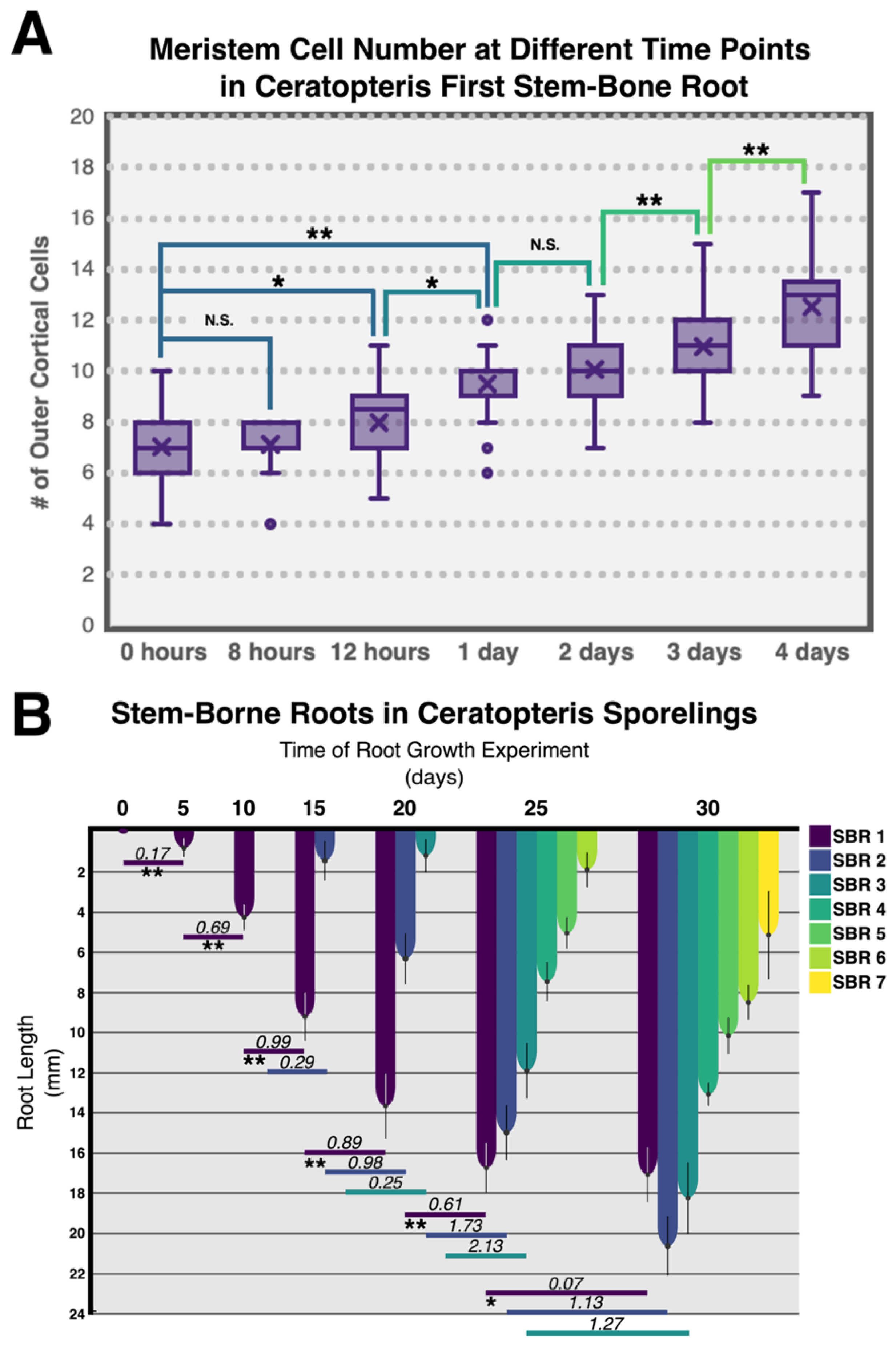
3.4. Growth Analysis of Ceratopteris Stem-Borne Roots in Sporelings
3.5. Cell Cycle Activity in the Root Apical Meristem
4. Discussion
4.1. How Early is the First Root Specified during Ceratopteris Embryogenesis?
4.2. The Ceratopteris Root Develops from a Narrow Multicellular Meristem
4.3. Ceratopteris Root Cell Layers Exhibit the Conservation of Specific Traits
4.4. Ceratopteris Stem-Borne Roots Display Growth Cessation
4.5. The Ceratopteris RAC Exhibits High Mitotic Activity
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| RAM | Root Apical Meristem |
| RAC | Root Apical Cell |
| RCI | Root Cap Initial |
| RSCN | Root Stem Cell Niche |
| SBR | Stem-Borne Root |
| QC | Quiescent Center |
| dps | days post sowing |
| daf | days after fertilization |
| hae | hours after exposure |
| dae | days after exposure |
| RGR | Root Growth Rate |
| EdU | 5-ethynyl-2′-deoxyuridine |
| PI | Propidium Iodide |
| CW | Calcofluor White |
| MZ | Meristematic Zone |
| EDZ | Elongation-Differentiation Zone |
References
- Delaux, P.-M.; Hetherington, A.J.; Coudert, Y.; Delwiche, C.; Dunand, C.; Gould, S.; Kenrick, P.; Li, F.-W.; Philippe, H.; Rensing, S.A.; et al. Reconstructing trait evolution in plant evo–devo studies. Curr. Biol. 2019, 29, R1110–R1118. [Google Scholar] [CrossRef]
- Fujinami, R.; Yamada, T.; Imaichi, R. Root apical meristem diversity and the origin of roots: Insights from extant lycophytes. J. Plant Res. 2020, 133, 291–296. [Google Scholar] [CrossRef]
- Hetherington, A.J.; Dolan, L. Stepwise and independent origins of roots among land plants. Nature 2018, 561, 235–238. [Google Scholar] [CrossRef]
- Kenrick, P.; Strullu-Derrien, C. The origin and early evolution of roots. Plant Physiol. 2014, 166, 570–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raven, J.A.; Edwards, D. Roots: Evolutionary origins and biogeochemical significance. J. Exp. Bot. 2001, 52 (Suppl. 1), 381–401. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xu, L. Recruitment of IC-WOX Genes in Root Evolution. Trends Plant Sci. 2018, 23, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Vasco, A.; Smalls, T.L.; Graham, S.W.; Cooper, E.D.; Wong, G.K.-S.; Stevenson, D.W.; Moran, R.C.; Ambrose, B.A. Challenging the paradigms of leaf evolution: Class III HD-Zips in ferns and lycophytes. New Phytol. 2016, 212, 745–758. [Google Scholar] [CrossRef] [PubMed]
- Bui, L.T.; Cordle, A.R.; Irish, E.E.; Cheng, C.L. Transient and stable transformation of Ceratopteris richardii gametophytes. BMC Res. Notes 2015, 8, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickok, L.G.; Warne, T.R.; Fribourg, R.S. The biology of the fern Ceratopteris and its use as a model system. Int. J. Plant Sci. 1995, 156, 332–345. [Google Scholar] [CrossRef]
- Marchant, D.B. Ferns with Benefits: Incorporating Ceratopteris into the Genomics Era. Am. Fern J. 2019, 109, 183. [Google Scholar] [CrossRef]
- Marchant, D.B.; Sessa, E.B.; Wolf, P.G.; Heo, K.; Barbazuk, W.B.; Soltis, P.S.; Soltis, D.E. The C-Fern (Ceratopteris richardii) genome: Insights into plant genome evolution with the first partial homosporous fern genome assembly. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Plackett, A.R.G.; Huang, L.; Sanders, H.L.; Langdale, J.A. High-efficiency stable transformation of the model fern species Ceratopteris richardii via microparticle bombardment. Plant Physiol. 2014, 165, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plackett, A.R.G.; Conway, S.J.; Hazelton, K.D.H.; Rabbinowitsch, E.H.; Langdale, J.A.; di Stilio, V.S. LEAFY maintains apical stem cell activity during shoot development in the fern ceratopteris richardii. Elife 2018, 7, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Conway, S.J.; di Stilio, V.S. An ontogenetic framework for functional studies in the model fern Ceratopteris richardii. Dev. Biol. 2020, 457, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Hetherington, A.J.; Dolan, L. The evolution of lycopsid rooting structures: Conservatism and disparity. New Phytol. 2017, 215, 538–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hetherington, A.J.; Dolan, L. Bilaterally symmetric axes with rhizoids composed the rooting structure of the common ancestor of vascular plants. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170042. [Google Scholar] [CrossRef] [Green Version]
- Hetherington, A.J.; Dolan, L. Rhynie chert fossils demonstrate the independent origin and gradual evolution of lycophyte roots. Curr. Opin. Plant Biol. 2019, 47, 119–126. [Google Scholar] [CrossRef]
- Kenrick, P. The origin of roots. In Plant Roots: The Hidden Half, 4th ed.; Fourth, E., Eshel, A., Beeckman, T., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 3–16. [Google Scholar]
- Dolan, L.; Janmaat, K.; Willemsen, V.; Linstead, P.; Poethig, S.; Roberts, K.; Scheres, B. Cellular organisation of the Arabidopsis thaliana root. Development 1993, 119, 71–84. [Google Scholar]
- Ron, M.; Dorrity, M.W.; De Lucas, M.; Toal, T.; Hernandez, R.I.; Little, S.A.; Maloof, J.; Kliebenstein, D.J.; Brady, S.M. Identification of novel loci regulating interspecific variation in root morphology and cellular development in tomato. Plant Physiol. 2013, 162, 755–768. [Google Scholar] [CrossRef] [Green Version]
- Ni, J.; Shen, Y.; Zhang, Y.; Wu, P. Definition and stabilisation of the quiescent centre in rice roots. Plant Biol. 2014, 16, 1014–1019. [Google Scholar] [CrossRef]
- Seago, J.L.; Fernando, D.D. Anatomical aspects of angiosperm root evolution. Ann. Bot. 2013, 112, 223–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, A. Root apical organization in gymnosperms—some cycads and Ginkgo biloba. Proc. Indian Acad. Sci. 1962, 57, 211–222. [Google Scholar]
- Peterson, C.A.; Enstone, D.E.; Taylor, J.H. Pine root structure and its potential significance for root function. Plant Soil 1999, 217, 205–213. [Google Scholar] [CrossRef]
- Hou, G.-C.; Hill, J.P. Developmental anatomy of the fifth shoot-borne root in young sporophytes of Ceratopteris richardii. Planta 2004, 219, 212–220. [Google Scholar] [CrossRef]
- Gunning, B.E.S.; Hughes, J.E.; Hardham, A.R. Formative and proliferative cell divisions, cell differentiation, and developmental changes in the meristem of Azolla roots. Planta 1978, 143, 121–144. [Google Scholar] [CrossRef]
- Gifford, E.M. The Root Apical Meristem of Equisetum diffusum: Structure and Development. Am. J. Bot. 1993, 80, 468. [Google Scholar] [CrossRef]
- Otreba, P.; Gola, E.M. Specific intercalary growth of rhizophores and roots in Selaginella kraussiana (Selaginellaceae) is related to unique dichotomous branching. Flora Morphol. Distrib. Funct. Ecol. Plants 2011, 206, 227–232. [Google Scholar] [CrossRef]
- Fujinami, R.; Yamada, T.; Nakajima, A.; Takagi, S.; Idogawa, A.; Kawakami, E.; Tsutsumi, M.; Imaichi, R. Root apical meristem diversity in extant lycophytes and implications for root origins. New Phytol. 2017, 215, 1210–1220. [Google Scholar] [CrossRef] [Green Version]
- Evkaikina, A.I.; Berke, L.; Romanova, M.A.; Proux-Wéra, E.; Ivanova, A.N.; Rydin, C.; Pawlowski, K.; Voitsekhovskaja, O.V. The Huperzia selago shoot tip transcriptome sheds new light on the evolution of leaves. Genome Biol. Evol. 2017, 9, 2444–2460. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Rodríguez, J.F.; Shishkova, S.; Napsucialy-Mendivil, S.; Dubrovsky, J.G. Apical meristem organization and lack of establishment of the quiescent center in Cactaceae roots with determinate growth. Planta 2003, 217, 849–857. [Google Scholar] [CrossRef]
- Kirschner, G.K.; Stahl, Y.; von Korff, M.; Simon, R. Unique and Conserved Features of the Barley Root Meristem. Front. Plant Sci. 2017, 8, 1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshpande, B.D.; Bahtnagar, P. Apical Meristems of Ephedra foliata. Int. J. Plant Sci. 1961, 122, 279–284. [Google Scholar] [CrossRef]
- Allen, G.S. Embryogeny and the Development of the Apical Meristems of Pseudotsuga. III. Development of the Apical Meristems. Am. J. Bot. 1947, 34, 204–211. [Google Scholar] [CrossRef]
- Johnson-Flanagan, A.M.; Owens, J.N. Development of white spruce (Picea glauca) seedling roots. Can. J. Bot. 1985, 63, 456–462. [Google Scholar] [CrossRef]
- Hetherington, A.J.; Dubrovsky, J.G.; Dolan, L. Unique Cellular Organization in the Oldest Root Meristem. Curr. Biol. 2016, 26, 1629–1633. [Google Scholar] [CrossRef] [Green Version]
- Bhambie, S.; Prakasa, C.G. Rao Studies in Pteridophytes IX. The Root Apex Organization in Some Pteridophytes. Proc. Plant Sci. 1972, 39, 150–156. [Google Scholar]
- Chapple, C.C.S.; Peterson, R.L. Root Structure in the Fern Platycerium bifurcatum (Cav.) C. Chr. (Polypodiaceae). Int. J. Plant Sci. 1987, 148, 180–187. [Google Scholar] [CrossRef]
- Pillai, A. Root apical organization in gymnosperms. Planta 1966, 70, 26–33. [Google Scholar] [CrossRef]
- Hou, G.; Hill, J.P. Heteroblastic Root Development in Ceratopteris richardii (Parkeriaceae). Int. J. Plant Sci. 2002, 163, 341–351. [Google Scholar] [CrossRef]
- Heimsch, C.; Seago, J.L. Organization of the root apical meristem in angiosperms. Am. J. Bot. 2008, 95, 1–21. [Google Scholar] [CrossRef]
- Schneider, H. Evolutionary Morphology of Ferns (Monilophytes). Evol. Plant Form 2012, 45, 115–140. [Google Scholar]
- Scheres, B.; Wolkenfelt, H.; Willemsen, V.; Terlouw, M.; Lawson, E.; Dean, C.; Weisbeek, P. Embryonic origin of the Arabidopsis primary root and root meristem initials. Development 1994, 120, 2475–2487. [Google Scholar]
- Warne, T.R.; Lloyd, R.M. The Role of Spore Germination and Gametophyte Development in Habitat Selection: Temperature Responses in Certain Temperature and Tropical Ferns. Torrey Bot. Soc. 1980, 107, 57–64. [Google Scholar] [CrossRef]
- Kurihara, D.; Mizuta, Y.; Sato, Y.; Higashiyama, T. ClearSee: A rapid optical clearing reagent for whole-plant fluorescence imaging. Development 2015, 142, 4168–4179. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Schiefelbein, J. Conserved gene expression programs in developing roots from diverse plants. Plant Cell 2015, 27, 2119–2132. [Google Scholar] [CrossRef] [Green Version]
- Lux, A.; Morita, S.; Abe, J.; Ito, K. An improved method for clearing and staining free-hand sections and whole-mount samples. Ann. Bot. 2005, 96, 989–996. [Google Scholar] [CrossRef]
- Cruz-Ramírez, A.; Díaz-Triviño, S.; Wachsman, G.; Du, Y.; Arteága-Vázquez, M.; Zhang, H.; Benjamins, R.; Blilou, I.; Neef, A.B.; Chandler, V.; et al. A SCARECROW-RETINOBLASTOMA Protein Network Controls Protective Quiescence in the Arabidopsis Root Stem Cell Organizer. PLoS Biol. 2013, 11. [Google Scholar] [CrossRef] [Green Version]
- Neef, A.B.; Luedtke, N.W. Dynamic metabolic labeling of DNA in vivo with arabinosyl nucleosides. Proc. Natl. Acad. Sci. USA 2011, 108, 20404–20409. [Google Scholar] [CrossRef] [Green Version]
- Johnson, G.P.; Renzaglia, K.S. Embryology of Ceratopteris richardii (Pteridaceae, tribe Ceratopterideae), with emphasis on placental development. J. Plant Res. 2008, 121, 581–592. [Google Scholar] [CrossRef]
- Wardlaw, C.W. Embryogenesis in Plants; Methuen: London, UK, 1955. [Google Scholar]
- DeMaggio, A.E. Cytological Aspects of Reproduction in Ferns. Bot. Rev. 1977, 43, 427–448. [Google Scholar] [CrossRef]
- Chiang, Y.-L.; Chiang, S.-H.T. The Sporeling of Ceratopteris. Taiwania 1960, 8, 35–50. [Google Scholar]
- Johnson, G.P.; Renzaglia, K.S. Evaluating the diversity of pteridophyte embryology in the light of recent phylogenetic analyses leads to new inferences on character evolution. Plant Syst. Evol. 2009, 283, 149–164. [Google Scholar] [CrossRef]
- Jenik, P.D.; Gillmor, C.S.; Lukowitz, W. Embryonic patterning in Arabidopsis thaliana. Annu. Rev. Cell Dev. Biol. 2007, 23, 207–236. [Google Scholar] [CrossRef] [PubMed]
- Bennett, T.; Scheres, B. Root development-two meristems for the price of one? Curr. Top. Dev. Biol. 2010, 91, 67–102. [Google Scholar] [PubMed]
- Nardmann, J.; Werr, W. The invention of WUS-like stem cell-promoting functions in plants predates leptosporangiate ferns. Plant Mol. Biol. 2012, 78, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Youngstrom, C.E.; Geadelmann, L.F.; Irish, E.E.; Cheng, C.L. A fern WUSCHEL-RELATED HOMEOBOX gene functions in both gametophyte and sporophyte generations. BMC Plant Biol. 2019, 19, 416. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Zhang, Y.; Liu, W.; Wang, H.; Wen, S.; Zhang, Y.; Xu, L. Molecular evolution of auxin-mediated root initiation in plants. Mol. Biol. Evol. 2019, 37, 1–19. [Google Scholar] [CrossRef] [Green Version]
- De Vries, J.; Fischer, A.M.; Roettger, M.; Rommel, S.; Schluepmann, H.; Bräutigam, A.; Carlsbecker, A.; Gould, S.B. Cytokinin-induced promotion of root meristem size in the fern Azolla supports a shoot-like origin of euphyllophyte roots. New Phytol. 2016, 209, 705–720. [Google Scholar] [CrossRef]
- Perilli, S.; Sabatini, S. Analysis of Root Meristem Size Development. Plant Dev. Biol. 2010, 655, 177–187. [Google Scholar]
- Moubayidin, L.; Salvi, E.; Giustini, L.; Terpstra, I.; Heidstra, R.; Costantino, P.; Sabatini, S. A SCARECROW-based regulatory circuit controls Arabidopsis thaliana meristem size from the root endodermis. Planta 2016, 243, 1159–1168. [Google Scholar] [CrossRef] [Green Version]
- Sebastian, J.; Ryu, K.H.; Zhou, J.; Tarkowská, D.; Tarkowski, P.; Cho, Y.; Yoo, S.; Kim, E.; Lee, J. PHABULOSA Controls the Quiescent Center-Independent Root Meristem Activities in Arabidopsis thaliana. PLoS Genet. 2015, 11, e1004973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motte, H.; Parizot, B.; Fang, T.; Beeckman, T. The evolutionary trajectory of root stem cells. Curr. Opin. Plant Biol. 2020, 53, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Ambrose, B.A.; Vasco, A. Bringing the multicellular fern meristem into focus. New Phytol. 2016, 210, 790–793. [Google Scholar] [CrossRef]
- Cruz, R.; Melo-De-pinna, G.F.A.; Vasco, A.; Prado, J.; Ambrose, B.A. Class I KNOX is related to determinacy during the leaf development of the fern Mickelia scandens (Dryopteridaceae). Int. J. Mol. Sci. 2020, 21, 4295. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, C.; Shivhare, D.; Hansen, B.O.; Pasha, A.; Esteban, E.; Provart, N.J.; Kragler, F.; Fernie, A.R.; Tohge, T.; Mutwil, M. Expression atlas of selaginella moellendorffii provides insights into the evolution of vasculature, secondary metabolism, and roots. Plant Cell 2020, 32, 853–870. [Google Scholar] [CrossRef] [PubMed]
- Imaichi, R.; Moritoki, N.; Solvang, H.K. Evolution of root apical meristem structures in vascular plants: Plasmodesmatal networks. Am. J. Bot. 2018, 105, 1453–1468. [Google Scholar] [CrossRef] [PubMed]
- Freeberg, J.A.; Gifford, E.M. The Root Apical Meristem of Osmunda regalis. Am. J. Bot. 1984, 71, 558–563. [Google Scholar] [CrossRef]
- White, R.A.; Turner, M.D. Anatomy and development of the fern sporophyte. Bot. Rev. 1995, 61, 281–305. [Google Scholar] [CrossRef]
- Gifford, E.M. Concept of Apical Cells in Bryophytes and Pteridophytes. Annu. Rev. Plant Physiol. 1983, 34, 419–440. [Google Scholar] [CrossRef]
- Barlow, P.W. The Root Cap: Cell Dynamics, Cell Differentiation and Cap Function. J. Plant Growth Regul. 2003, 21, 261–286. [Google Scholar] [CrossRef]
- Geldner, N. The Endodermis. Annu. Rev. Plant Biol. 2013, 64, 531–558. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Lee, S.C.; Choi, H.K. Anatomical patterns of aerenchyma in aquatic and wetland plants. J. Plant Biol. 2008, 51, 428–439. [Google Scholar] [CrossRef]
- Pires, N.D.; Dolan, L. Morphological evolution in land plants: New designs with old genes. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 508–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, H.; Yamauchi, T.; Colmer, T.D.; Nakazono, M. Aerenchyma Formation in Plants. Plant Cell Monogr. 2014, 21, 247–265. [Google Scholar] [CrossRef]
- Lucas, W.J.; Groover, A.; Lichtenberger, R.; Furuta, K.; Yadav, S.R.; Helariutta, Y.; Patrick, J.W. The Plant Vascular System: Evolution, Development and Functions. J. Integr. Plant Biol. 2013, 55, 294–388. [Google Scholar] [CrossRef]
- Chiang, S.-H.T.; Chou, T. Tissue Differentiation in the Roots of Some Ferns. Taiwania 1974, 19, 7–18. [Google Scholar]
- Pittermann, J.; Watkins, J.E.; Cary, K.L.; Schuettpelz, E.; Brodersen, C.R.; Smith, A.R.; Baer, A. The Structure and Function of Xylem in Seed-Free Vascular Plants: An Evolutionary Perspective. In Functional and Ecological Xylem Anatomy; Springer International Publishing: Cham, Switzerland, 2015; pp. 1–37. [Google Scholar]
- De Rybel, B.; Mähönen, A.P.; Helariutta, Y.; Weijers, D. Plant vascular development: From early specification to differentiation. Nat. Rev. Mol. Cell Biol. 2016, 17, 30–40. [Google Scholar] [CrossRef]
- Eeckhout, S.; Leroux, O.; Willats, W.G.T.; Popper, Z.A.; Viane, R.L.L. Comparative glycan profiling of Ceratopteris richardii ‘C-Fern’ gametophytes and sporophytes links cell-wall composition to functional specialization. Ann. Bot. 2014, 114, 1295–1307. [Google Scholar] [CrossRef] [Green Version]
- PriestleyI, J.H.; Radcliffe, F.M. A Study in the Endodermis in the Filicineae. New Phytol. 1924, 23, 161–193. [Google Scholar] [CrossRef]
- Damus, M.; Peterson, R.L.; Enstone, D.E.; Peterson, C.A. Modifications of cortical cell walls in roots of seedless vascular plants. Bot. Acta 1997, 110, 190–195. [Google Scholar] [CrossRef]
- Kumpf, R.P.; Nowack, M.K. The root cap: A short story of life and death. J. Exp. Bot. 2015, 66, 5651–5662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eastman, A.; Peterson, R.L. Root Apex Structure in Regnellidium diphyllum (Marsileaceae). Int. J. Plant Sci. 1985, 146, 44–55. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, G.; Wang, X.; Zhang, X.; Friml, J. Evolution of fast root gravitropism in seed plants. Nat. Commun. 2019, 10, 4–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bui, L.T.; Pandzic, D.; Youngstrom, C.E.; Wallace, S.; Irish, E.E.; Szövényi, P.; Cheng, C. A fern AINTEGUMENTA gene mirrors BABY BOOM in promoting apogamy in Ceratopteris richardii. Plant J. 2017, 90, 122–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasebe, M.; Wen, C.K.; Kato, M.; Banks, J.A. Characterization of MADS homeotic genes in the fern Ceratopteris richardii. Proc. Natl. Acad. Sci. USA 1998, 95, 6222–6227. [Google Scholar] [CrossRef] [Green Version]
- Sano, R.; Juarez, C.M.; Hass, B.; Sakakibara, K.; Ito, M.; Banks, J.A.; Hasebe, M. KNOX homeobox genes potentially have similar function in both diploid unicellular and multicellular meristems, but not in haploid meristems. Evol. Dev. 2005, 7, 9–78. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiao, Y.; Jiao, H.; Zhao, H.; Zhu, Y.X. Two-step functional innovation of the stem-cell factors WUS/WOX5 during plant evolution. Mol. Biol. Evol. 2017, 34, 640–653. [Google Scholar] [CrossRef] [Green Version]
- Zumajo-Cardona, C.; Vasco, A.; Ambrose, B.A. The Evolution of the KANADI Gene Family and Leaf Development in Lycophytes and Ferns. Plants 2019, 8, 313. [Google Scholar] [CrossRef] [Green Version]
- Bennett, T.; Brockington, S.F.; Rothfels, C.; Graham, S.W.; Stevenson, D.; Kutchan, T.; Rolf, M.; Thomas, P.; Wong, G.K.-S.; Leyser, O.; et al. Paralogous radiations of PIN proteins with multiple origins of noncanonical PIN structure. Mol. Biol. Evol. 2014, 31, 2042–2060. [Google Scholar] [CrossRef] [Green Version]
- Kurth, E. Mitotic Activity in the Root Apex of the Water Fern Marsilea vestita Hook. and Grev. Am. J. Bot. 1981, 68, 881–896. [Google Scholar] [CrossRef]
- Kurth, E.; Gifford, E.M. Ontogenetic Changes in DNA Content in Roots of the Water Fern Azolla filiculoides. Am. J. Bot. 1985, 72, 1676–1683. [Google Scholar] [CrossRef]
- Nitayangkura, S.; Gifford, E.M.; Rost, T.L. Mitotic Activity in the Root Apical Meristem of Azolla filiculoides Lam., with Special Reference to the Apical Cell. Bot. Soc. Am. 1980, 67, 1484–1492. [Google Scholar] [CrossRef]
- Benesova, S. Development Related Termination of the Root Apical Meristem Activity. Master’s Thesis, Experimental Plant Biology. Charles University in Prague Faculty of Science, Nové Město, Czech Republic, 2016. [Google Scholar]
- Gutiérrez-Alanís, D.; Yong-Villalobos, L.; Jiménez-Sandoval, P.; Alatorre-Cobos, F.; Oropeza-Aburto, A.; Mora-Macías, J.; Sánchez-Rodríguez, F.; Cruz-Ramírez, A.; Herrera-Estrella, L. Phosphate Starvation-Dependent Iron Mobilization Induces CLE14 Expression to Trigger Root Meristem Differentiation through CLV2/PEPR2 Signaling. Dev. Cell 2017, 41, 555–570.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Alonso, G.; Matvienko, M.; López-Valle, M.L.; Lázaro-Mixteco, P.E.; Napsucialy-Mendivil, S.; Dubrovsky, J.G.; Shishkova, S. Transcriptomics insights into the genetic regulation of root apical meristem exhaustion and determinate primary root growth in Pachycereus pringlei (Cactaceae). Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Gunning, B.E.S. Age-related and origin-related control of the numbers of plasmodesmata in cell walls of developing Azolla roots. Planta 1978, 143, 181–190. [Google Scholar]
- Gifford, E.M.; Kurth, E. Quantitative Studies of the Root Apical Meristem of Equisetum scirpoides. Am. J. Bot. 1982, 69, 464. [Google Scholar] [CrossRef]
- Chiang, S.-H.T. The Time of Mitosis in the Root Apical Meristem of Ceratopteris pteridoides. Taiwania 1972, 17, 14–26. [Google Scholar]
- Barlow, P.W. Meristems, metamers and modules and the development of shoot and root systems. Bot. J. Linn. Soc. 1989, 100, 255–279. [Google Scholar] [CrossRef]
- Pi, L.; Aichinger, E.; Van Der Graaff, E.; Llavata-Peris, C.I.; Weijers, D.; Hennig, L.; Groot, E.; Laux, T. Organizer-Derived WOX5 Signal Maintains Root Columella Stem Cells through Chromatin-Mediated Repression of CDF4 Expression. Dev. Cell 2015, 33, 576–588. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | Cell Size | B | Cell Number | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Length (µm) | Width (µm) | n | |||||||
| Mean | SD (+/−) | Mean | SD (+/−) | Mean | SD (+/−) | n | |||
| Meristematic Zone | 16.32 | 2.42 | 18.36 | 2.28 | 20 | 7.00 | 1.34 | 50 | |
| Elongation/Differentiation Zone | 36.79 | 12.09 | 25.22 | 3.62 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aragón-Raygoza, A.; Vasco, A.; Blilou, I.; Herrera-Estrella, L.; Cruz-Ramírez, A. Development and Cell Cycle Activity of the Root Apical Meristem in the Fern Ceratopteris richardii. Genes 2020, 11, 1455. https://doi.org/10.3390/genes11121455
Aragón-Raygoza A, Vasco A, Blilou I, Herrera-Estrella L, Cruz-Ramírez A. Development and Cell Cycle Activity of the Root Apical Meristem in the Fern Ceratopteris richardii. Genes. 2020; 11(12):1455. https://doi.org/10.3390/genes11121455
Chicago/Turabian StyleAragón-Raygoza, Alejandro, Alejandra Vasco, Ikram Blilou, Luis Herrera-Estrella, and Alfredo Cruz-Ramírez. 2020. "Development and Cell Cycle Activity of the Root Apical Meristem in the Fern Ceratopteris richardii" Genes 11, no. 12: 1455. https://doi.org/10.3390/genes11121455
APA StyleAragón-Raygoza, A., Vasco, A., Blilou, I., Herrera-Estrella, L., & Cruz-Ramírez, A. (2020). Development and Cell Cycle Activity of the Root Apical Meristem in the Fern Ceratopteris richardii. Genes, 11(12), 1455. https://doi.org/10.3390/genes11121455