Carriage and Gene Content Variability of the pESI-Like Plasmid Associated with Salmonella Infantis Recently Established in United States Poultry Production

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Carriage
2.2. Gene Content Analysis
2.3. Hierarchal Clustering and Dendrogram Visualization
3. Results
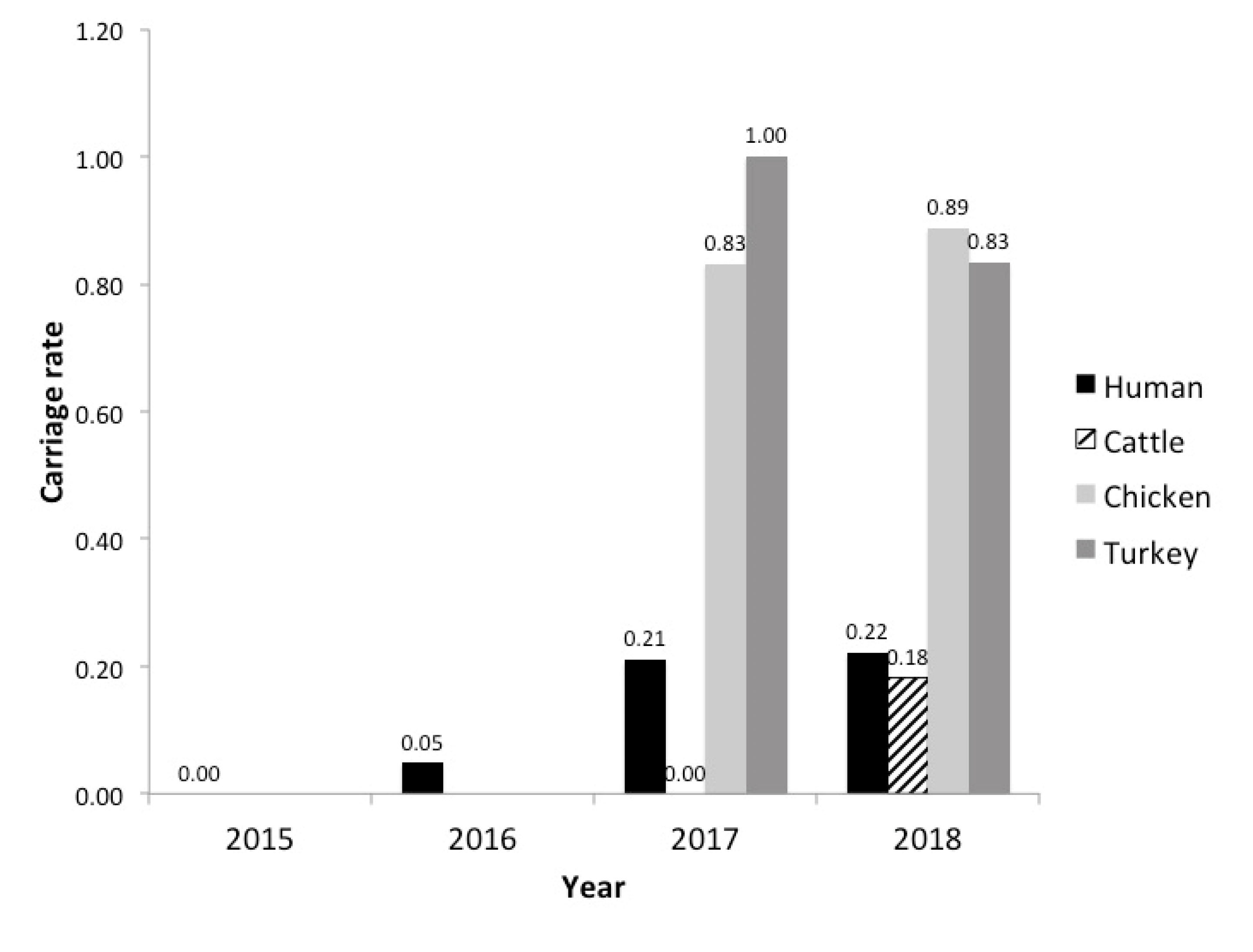
3.1. Plasmid Carriage
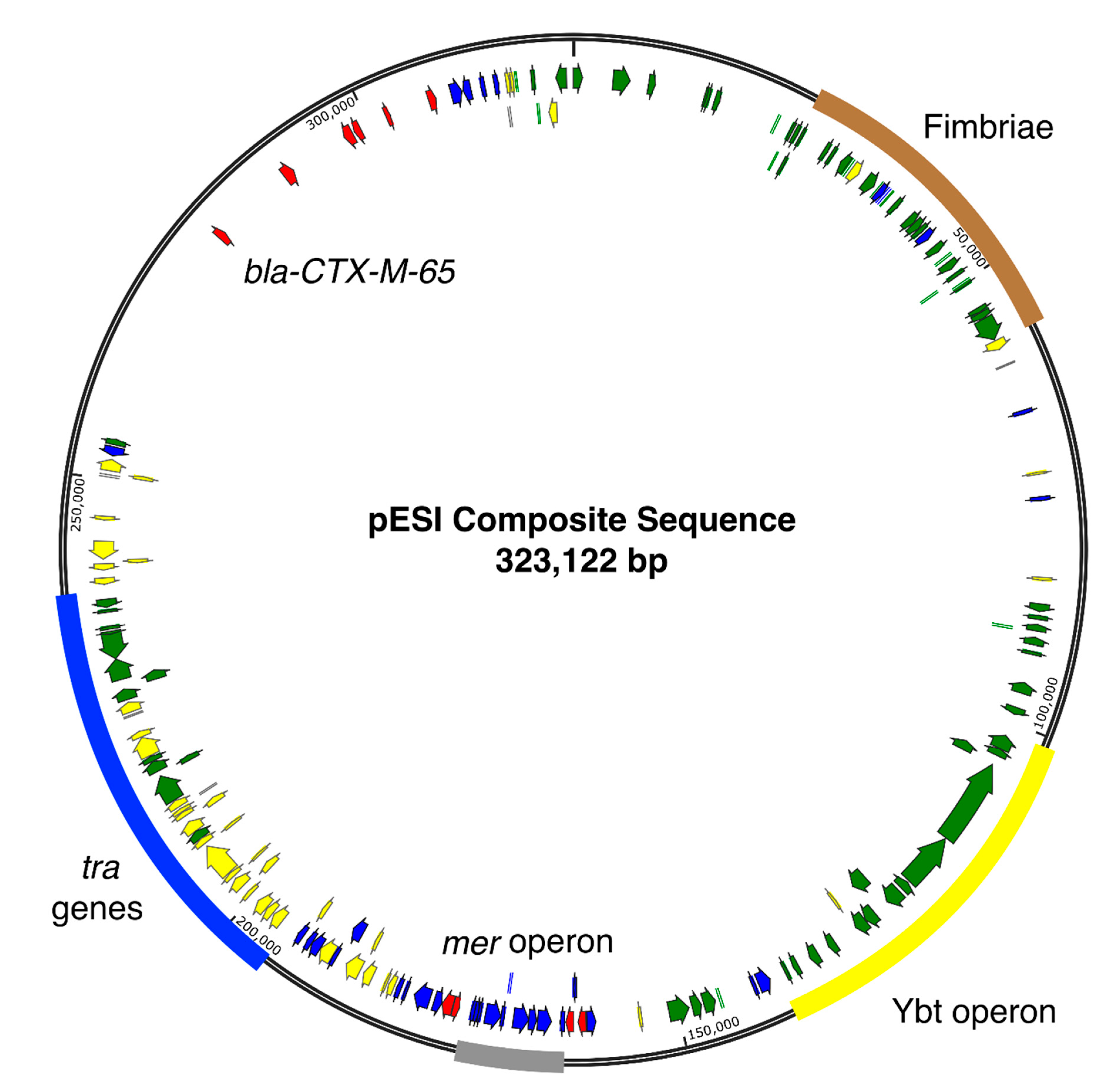
3.2. Gene Content Analysis
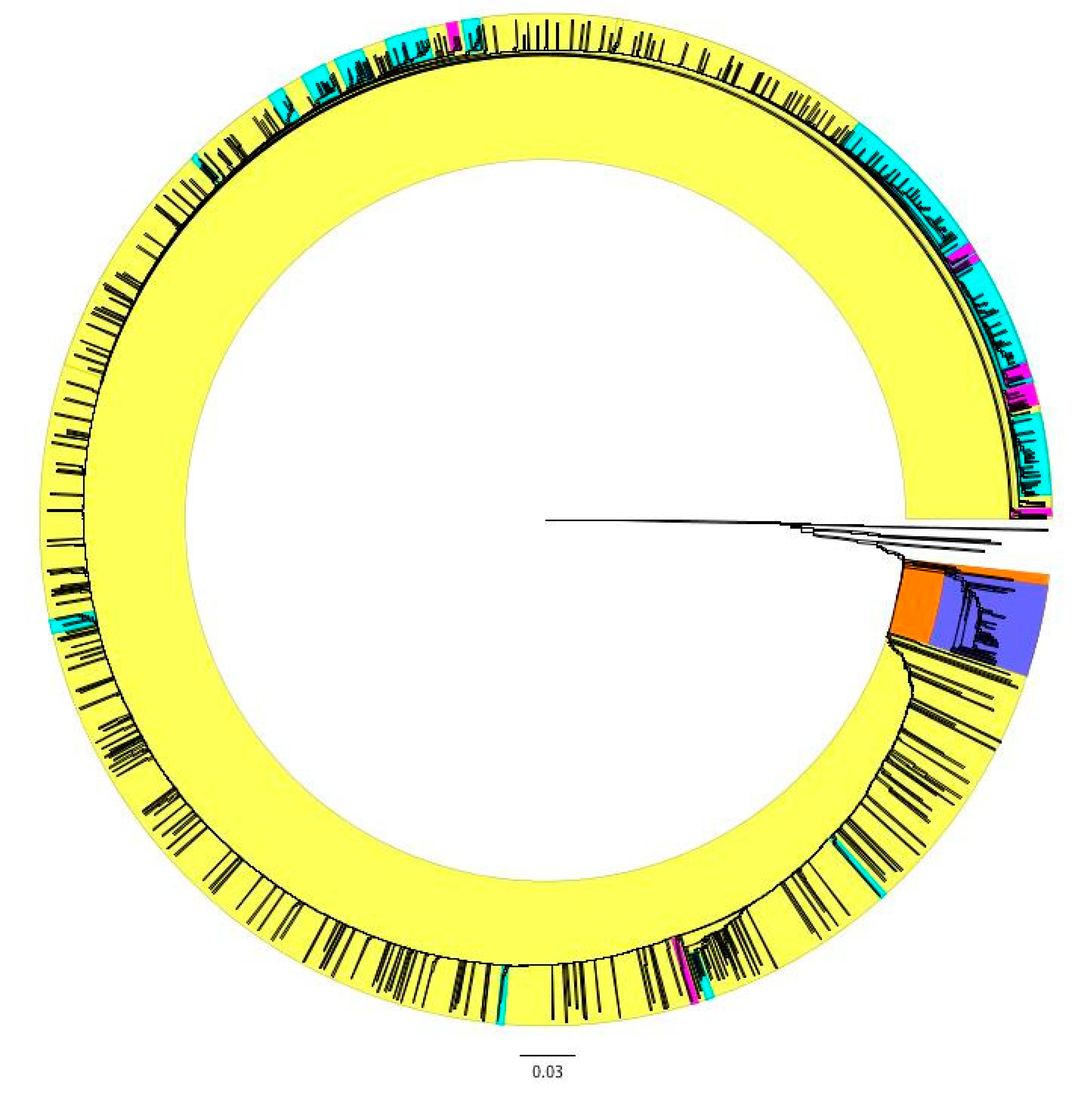
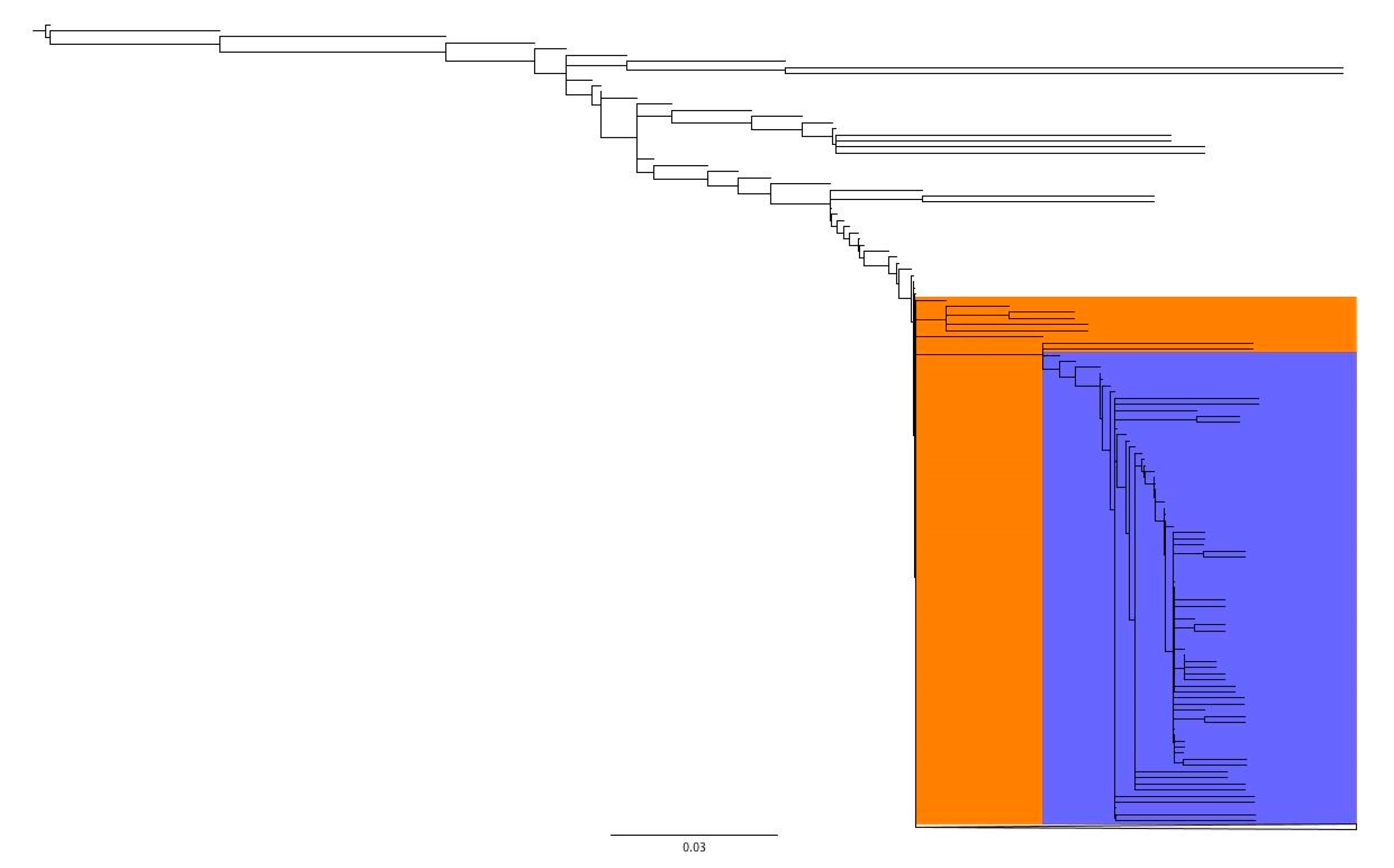
3.3. Hierarchal Clustering
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Note
References
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States--major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- CDC. National Enteric Disease Surveillance: Salmonella Annual Report, 2016; US Department of Health and Human Services, Centers for Disease Control and Prevention: Atlanta, GA, USA, 2018.
- FSIS. Progress Report on Salmonella and Campylobacter Testing of Raw Meat and Poultry Products, 1998–2014; USDA: Washington, DC, USA, 2014.
- FSIS. Serotypes Profile of Salmonella Isolates from Meat and Poultry Products January 1998 through December 2014; USDA: Washington, DC, USA, 2016.
- Tack, D.M.; Ray, L.; Griffin, P.M.; Cieslak, P.R.; Dunn, J.; Rissman, T.; Jervis, R.; Lathrop, S.; Muse, A.; Duwell, M.; et al. Preliminary incidence and trends of infections with pathogens transmitted commonly through food—Foodborne diseases active surveillance network, 10 U.S. sites, 2016–2019. Morb. Mortal. Wkly. Rep. 2020, 69, 509–514. [Google Scholar] [CrossRef] [PubMed]
- CDC. Outbreak of Multidrug-Resistant Salmonella Infections Linked to Raw Chicken Products. Available online: https://www.cdc.gov/salmonella/infantis-10-18/index.html (accessed on 16 September 2019).
- CDC. Antibiotic Resistance Threats in the United States, 2019; CDC: Atlanta, GA, USA, 2019.
- Sjolund-Karlsson, M.; Howie, R.; Krueger, A.; Rickert, R.; Pecic, G.; Lupoli, K.; Folster, J.P.; Whichard, J.M. CTX-M-producing non-Typhi Salmonella spp. isolated from humans, United States. Emerg. Infect. Dis. 2011, 17, 97–99. [Google Scholar] [CrossRef]
- Doi, Y.; Iovleva, A.; Bonomo, R.A. The ecology of extended-spectrum beta-lactamases (ESBLs) in the developed world. J. Travel Med. 2017, 24, S44–S51. [Google Scholar] [CrossRef]
- Crump, J.A.; Sjolund-Karlsson, M.; Gordon, M.A.; Parry, C.M. Epidemiology, clinical presentation, laboratory diagnosis, antimicrobial resistance, and antimicrobial management of invasive Salmonella infections. Clin. Microbiol. Rev. 2015, 28, 901–937. [Google Scholar] [CrossRef] [Green Version]
- Franco, A.; Leekitcharoenphon, P.; Feltrin, F.; Alba, P.; Cordaro, G.; Iurescia, M.; Tolli, R.; D’Incau, M.; Staffolani, M.; Di Giannatale, E.; et al. Emergence of a Clonal Lineage of Multidrug-Resistant ESBL-Producing Salmonella Infantis Transmitted from Broilers and Broiler Meat to Humans in Italy between 2011 and 2014. PLoS ONE 2015, 10, e0144802. [Google Scholar] [CrossRef] [Green Version]
- Cartelle Gestal, M.; Zurita, J.; Paz, Y.M.A.; Ortega-Paredes, D.; Alcocer, I. Characterization of a small outbreak of Salmonella enterica serovar Infantis that harbour CTX-M-65 in Ecuador. Braz. J. Infect. Dis. 2016, 20, 406–407. [Google Scholar] [CrossRef] [Green Version]
- Iriarte, A.; Giner-Lamia, J.; Silva, C.; Betancor, L.; Astocondor, L.; Cestero, J.J.; Ochoa, T.; Garcia, C.; Puente, J.L.; Chabalgoity, J.A.; et al. Draft Genome Sequence of Salmonella enterica subsp. enterica Serovar Infantis Strain SPE101, Isolated from a Chronic Human Infection. Genome Announc. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Fuentes-Castillo, D.; Farfan-Lopez, M.; Esposito, F.; Moura, Q.; Fernandes, M.R.; Lopes, R.; Cardoso, B.; Munoz, M.E.; Cerdeira, L.; Najle, I.; et al. Wild owls colonized by international clones of extended-spectrum beta-lactamase (CTX-M)-producing Escherichia coli and Salmonella Infantis in the Southern Cone of America. Sci. Total Environ. 2019, 674, 554–562. [Google Scholar] [CrossRef]
- Hindermann, D.; Gopinath, G.; Chase, H.; Negrete, F.; Althaus, D.; Zurfluh, K.; Tall, B.D.; Stephan, R.; Nuesch-Inderbinen, M. Salmonella enterica serovar Infantis from Food and Human Infections, Switzerland, 2010-2015: Poultry-Related Multidrug Resistant Clones and an Emerging ESBL Producing Clonal Lineage. Front. Microbiol. 2017, 8, 1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aviv, G.; Rahav, G.; Gal-Mor, O. Horizontal Transfer of the Salmonella enterica Serovar Infantis Resistance and Virulence Plasmid pESI to the Gut Microbiota of Warm-Blooded Hosts. MBio 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aviv, G.; Tsyba, K.; Steck, N.; Salmon-Divon, M.; Cornelius, A.; Rahav, G.; Grassl, G.A.; Gal-Mor, O. A unique megaplasmid contributes to stress tolerance and pathogenicity of an emergent Salmonella enterica serovar Infantis strain. Environ. Microbiol. 2014, 16, 977–994. [Google Scholar] [CrossRef] [PubMed]
- Tate, H.; Folster, J.P.; Hsu, C.-H.; Chen, J.; Hoffmann, M.; Li, C.; Morales, C.; Tyson, G.H.; Murkherjee, S.; Brown, A.C.; et al. Comparative Analysis of Extended-Spectrum-Beta-Lactamase CTX-M-65 Producing Salmonella enterica Serovar Infantis Isolates from Humans, Food Animals, and Retail Chickens in the United States. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.C.; Chen, J.C.; Watkins, L.K.F.; Campbell, D.; Folster, J.P.; Tate, H.; Wasilenko, J.; Van Tubbergen, C.; Friedman, C.R. CTX-M-65 Extended-Spectrum beta-Lactamase-Producing Salmonella enterica Serotype Infantis, United States(1). Emerg. Infect. Dis. 2018, 24, 2284–2291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, B.P.; Lattuada, C.P. (Eds.) Microbiology Laboratory Guidebook; Food Safety and Inspection Service: Washington, DC, USA, 1998.
- CDC. National Antimicrobial Resistance Monitoring System for Enteric Bacteria (NARMS): Human Isolates Surveillance Report for 2015 (Final Report); Department of Health and Human Services: Atlanta, GA, USA, 2018.
- Zhang, S.; Yin, Y.; Jones, M.B.; Zhang, Z.; Deatherage Kaiser, B.L.; Dinsmore, B.A.; Fitzgerald, C.; Fields, P.I.; Deng, X. Salmonella serotype determination utilizing high-throughput genome sequencing data. J. Clin. Microbiol. 2015, 53, 1685–1692. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Alikhan, N.F.; Zhou, Z.; Sergeant, M.J.; Achtman, M. A genomic overview of the population structure of Salmonella. PLoS Genet. 2018, 14, e1007261. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R package version 2.5–6. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 23 November 2019).
- Grimont, P.A.D.; Weill, F.-X. Antigenic Formulae of the Salmonella Serovars; World Health Organization: Geneva, Switzerland, 2007; p. 9. [Google Scholar]
- Bogomazova, A.N.; Gordeeva, V.D.; Krylova, E.V.; Soltynskaya, I.V.; Davydova, E.E.; Ivanova, O.E.; Komarov, A.A. Mega-plasmid found worldwide confers multiple antimicrobial resistance in Salmonella Infantis of broiler origin in Russia. Int. J. Food Microbiol. 2020, 319, 108497. [Google Scholar] [CrossRef]
- Fairchild, B. Environmental Factors to Control When Brooding Chicks. Univ. Ga. Coop. Ext. Bull. 2012, 1287, 26–37. [Google Scholar]
- Thomas, C.M.; Nielsen, K.M. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.E.; Walsh, T.R. Toxin–antitoxin systems and their role in disseminating and maintaining antimicrobial resistance. FEMS Microbiol. Rev. 2017, 41, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Wang, X.; Xu, H.; Xu, Y.; Ling, J.; Zhang, D.; Gao, S.; Liu, X. Roles of iron acquisition systems in virulence of extraintestinal pathogenic Escherichia coli: Salmochelin and aerobactin contribute more to virulence than heme in a chicken infection model. BMC Microbiol. 2012, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepp, D.; Gong, J.; Songer, J.G.; Boerlin, P.; Parreira, V.R.; Prescott, J.F. Identification of Accessory Genome Regions in Poultry Clostridium perfringens Isolates Carrying the netB Plasmid. J. Bacteriol. 2013, 195, 1152–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, M.; Butcher, J.; Stintzi, A. Nutrient acquisition and metabolism by Campylobacter jejuni. Front. In Cell. Infect. Microbiol. 2012, 2. [Google Scholar] [CrossRef] [Green Version]
- Foley, S.L.; Johnson, T.J.; Ricke, S.C.; Nayak, R.; Danzeisen, J. Salmonella pathogenicity and host adaptation in chicken-associated serovars. Microbiol. Mol. Biol. Rev. 2013, 77, 582–607. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Loui, C.; Clavijo, R.I.; Riley, L.W.; Lu, S. Survival characteristics of Salmonella enterica serovar Enteritidis in chicken egg albumen. Epidemiol. Infect. 2006, 134, 967–976. [Google Scholar] [CrossRef]
- FDA. Withdrawal of Approval of New Animal Drug Applications; Carbarsone; Roxarsone. Federal Register, 22 November 2013. [Google Scholar]
- Center for Veterinary Medicine. New Animal Drugs and New Animal Drug Combination Products Administered in or on Medicated Feed or Drinking Water of Food-Producing Animals: Recommendations for Drug Sponsors for Voluntarily Aligning Product Use Conditions with GFI #209; FDA: Silver Spring, MD, USA, 2013.
- Poultry Health Today. More Than Half of US Broilers Raised without Antibiotics in 2018. Available online: https://poultryhealthtoday.com/more-than-half-of-us-broilers-raised-without-antibiotics-in-2018/ (accessed on 12 January 2020).
- FDA. New Animal Drugs; Cephalosporin Drugs; Extralable Animal Drug Use; Order of Prohibition. Federal Register, 6 January 2012; 735–745. [Google Scholar]
- Alba, P.; Leekitcharoenphon, P.; Carfora, V.; Amoruso, R.; Cordaro, G.; Di Matteo, P.; Ianzano, A.; Iurescia, M.; Diaconu, E.L.; Study Group, E.N.; et al. Molecular epidemiology of Salmonella Infantis in Europe: Insights into the success of the bacterial host and its parasitic pESI-like megaplasmid. Microb. Genom. 2020, 6. [Google Scholar] [CrossRef]
- Acar, S.; Bulut, E.; Stasiewicz, M.J.; Soyer, Y. Genome analysis of antimicrobial resistance, virulence, and plasmid presence in Turkish Salmonella serovar Infantis isolates. Int. J. Food Microbiol. 2019, 307, 108275. [Google Scholar] [CrossRef] [PubMed]
- Szmolka, A.; Szabó, M.; Kiss, J.; Pászti, J.; Adrián, E.; Olasz, F.; Nagy, B. Molecular epidemiology of the endemic multiresistance plasmid pSI54/04 of Salmonella Infantis in broiler and human population in Hungary. Food Microbiol. 2018, 71, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, E.; Ando, N.; Ohta, T.; Kanada, A.; Shiwa, Y.; Ishige, T.; Murakami, K.; Kikuchi, T.; Murakami, S. A novel subpopulation of Salmonella enterica serovar Infantis strains isolated from broiler chicken organs other than the gastrointestinal tract. Vet. Microbiol. 2015, 175, 312–318. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Target Function |
|---|---|
| ardA * | Restriction modification enzyme |
| traI | Relaxase (IncI1) |
| sogS * | Primase (IncI1) |
| trbA * | Conjugal transfer protein (IncI1) |
| pESI repA | Plasmid replication |
| blaCTX-M-65 | Extended Spectrum Betalactamase |
| IncP | Iterons associated with IncP |
| hypothetical backbone protein * | Hypothetical protein associated with pESI |
| K88 * | Fimbriae |
| Ybt * | irp2 (Ybt operon); yersiniabactin biosynthesis |
| merA * | Mercuric reductase (mer operon) |
| ipf * | Fimbriae |
| pilL * | Pilus biosynthesis (IncI1) |
| Targets Present in Isolates Containing the pESI Plasmid | 2017 | 2018 | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chicken | Turkey | Chicken | Cattle | Turkey | ||||||||||||||
| I1 | pESI repA | ipf | IncP | blaCTX-M-65 | ardA | pilL | sogS | trbA | K88 | merA | Ytb | Infantis | Rough 1,5- | Infantis | Infantis | Rough 1,5- | Infantis | Infantis |
| + | + | + | + | + | + | + | + | + | + | + | + | 111 | 4 | 4 | 103 | 3 | 1 | 5 |
| + | + | + | + | + | + | + | + | + | + | - | + | 1 | 0 | 0 | 1 | 0 | 0 | 0 |
| + | + | + | + | + | + | + | + | + | - | + | + | 14 | 1 | 1 | 33 | 1 | 1 | 4 |
| + | + | + | + | - | + | + | + | + | + | + | + | 102 | 4 | 4 | 130 | 6 | 0 | 5 |
| + | + | + | + | - | + | + | + | + | - | + | + | 11 | 1 | 0 | 34 | 2 | 0 | 0 |
| + | + | + | + | - | + | + | + | + | - | - | + | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| + | + | + | + | - | - | + | + | + | + | + | + | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| + | + | + | + | - | - | + | + | + | - | + | + | 1 | 0 | 0 | 1 | 0 | 0 | 0 |
| + | + | + | - | + | + | + | + | + | + | - | + | 5 | 0 | 0 | 5 | 0 | 0 | 0 |
| + | + | + | - | + | + | + | + | + | - | + | + | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| + | + | + | - | + | + | + | + | + | - | - | + | 2 | 0 | 0 | 0 | 0 | 0 | 0 |
| + | + | + | - | + | + | - | + | + | + | - | + | 5 | 0 | 0 | 2 | 0 | 0 | 0 |
| + | + | + | - | + | + | - | + | + | - | - | + | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| + | + | + | - | + | + | - | - | + | + | - | + | 0 | 0 | 0 | 2 | 0 | 0 | 0 |
| + | + | + | - | + | + | - | - | + | - | - | + | 0 | 0 | 0 | 2 | 0 | 0 | 0 |
| + | + | + | - | - | + | + | + | + | + | + | + | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| + | + | + | - | - | + | + | + | + | + | - | + | 2 | 0 | 0 | 10 | 0 | 0 | 0 |
| + | + | + | - | - | + | + | + | + | - | - | + | 2 | 0 | 0 | 2 | 0 | 0 | 0 |
| + | + | + | - | - | + | - | + | + | + | - | + | 2 | 0 | 0 | 5 | 0 | 0 | 1 |
| + | + | + | - | - | + | - | + | + | - | - | + | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| + | + | - | + | + | + | + | + | + | + | + | + | 0 | 0 | 0 | 4 | 0 | 0 | 0 |
| + | + | - | + | + | + | + | + | + | - | + | + | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| + | + | - | + | + | + | - | + | + | - | + | + | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| + | + | - | + | - | + | + | + | + | + | + | + | 0 | 0 | 0 | 2 | 0 | 0 | 0 |
| + | + | - | + | - | + | + | + | + | - | + | + | 0 | 0 | 0 | 3 | 0 | 0 | 0 |
| + | + | - | - | + | + | + | + | + | + | - | + | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Total | 261 | 10 | 9 | 345 | 12 | 2 | 15 | |||||||||||
| Targets Present in Isolates Containing the pESI Plasmid | 2015 | 2016 | 2017 | 2018 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I1 | pESI repA | ipf | IncP | blaCTX-M-65 | ardA | pilL | sogS | trbA | K88 | merA | Ytb | hyp | Infantis | Infantis * | Infantis | I 6,7:-:,1,5 | Rough 1,5 | |
| + | + | + | + | + | + | + | + | + | + | + | + | + | 0 | 3 | 0 | 0 | 0 | 0 |
| + | + | + | + | + | + | + | + | + | + | + | + | - | 0 | 0 | 15 | 14 | 2 | 1 |
| + | + | + | + | + | - | + | + | + | + | + | + | - | 0 | 0 | 0 | 1 | 0 | 0 |
| + | + | + | + | - | + | + | + | + | + | + | + | - | 0 | 1 | 5 | 7 | 2 | 0 |
| + | + | + | - | + | + | - | + | + | + | - | + | - | 0 | 0 | 1 | 1 | 0 | 0 |
| + | + | + | - | + | + | + | + | + | + | - | + | - | 0 | 0 | 0 | 1 | 0 | 0 |
| + | + | + | - | - | + | + | + | + | + | - | + | - | 0 | 0 | 0 | 1 | 0 | 0 |
| Total | 0 | 4 | 21 | 25 | 4 | 1 | ||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McMillan, E.A.; Wasilenko, J.L.; Tagg, K.A.; Chen, J.C.; Simmons, M.; Gupta, S.K.; Tillman, G.E.; Folster, J.; Jackson, C.R.; Frye, J.G. Carriage and Gene Content Variability of the pESI-Like Plasmid Associated with Salmonella Infantis Recently Established in United States Poultry Production. Genes 2020, 11, 1516. https://doi.org/10.3390/genes11121516
McMillan EA, Wasilenko JL, Tagg KA, Chen JC, Simmons M, Gupta SK, Tillman GE, Folster J, Jackson CR, Frye JG. Carriage and Gene Content Variability of the pESI-Like Plasmid Associated with Salmonella Infantis Recently Established in United States Poultry Production. Genes. 2020; 11(12):1516. https://doi.org/10.3390/genes11121516
Chicago/Turabian StyleMcMillan, Elizabeth A., Jamie L. Wasilenko, Kaitlin A. Tagg, Jessica C. Chen, Mustafa Simmons, Sushim K. Gupta, Glenn E. Tillman, Jason Folster, Charlene R. Jackson, and Jonathan G. Frye. 2020. "Carriage and Gene Content Variability of the pESI-Like Plasmid Associated with Salmonella Infantis Recently Established in United States Poultry Production" Genes 11, no. 12: 1516. https://doi.org/10.3390/genes11121516
APA StyleMcMillan, E. A., Wasilenko, J. L., Tagg, K. A., Chen, J. C., Simmons, M., Gupta, S. K., Tillman, G. E., Folster, J., Jackson, C. R., & Frye, J. G. (2020). Carriage and Gene Content Variability of the pESI-Like Plasmid Associated with Salmonella Infantis Recently Established in United States Poultry Production. Genes, 11(12), 1516. https://doi.org/10.3390/genes11121516