BTG4 is A Novel p53 Target Gene That Inhibits Cell Growth and Induces Apoptosis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cloning of BTG4 Gene Promoter Region
2.3. Site-Directed Mutagenesis
2.4. Transfection and Luciferase Reporter Assay
2.5. DNA Sequence Alignment and Database Mining
2.6. RNA Extraction and Quantitative Real-Time RT-PCR
2.7. Cell Proliferation Assay
2.8. Wound-Healing Assay
2.9. Apoptosis Assay
3. Results
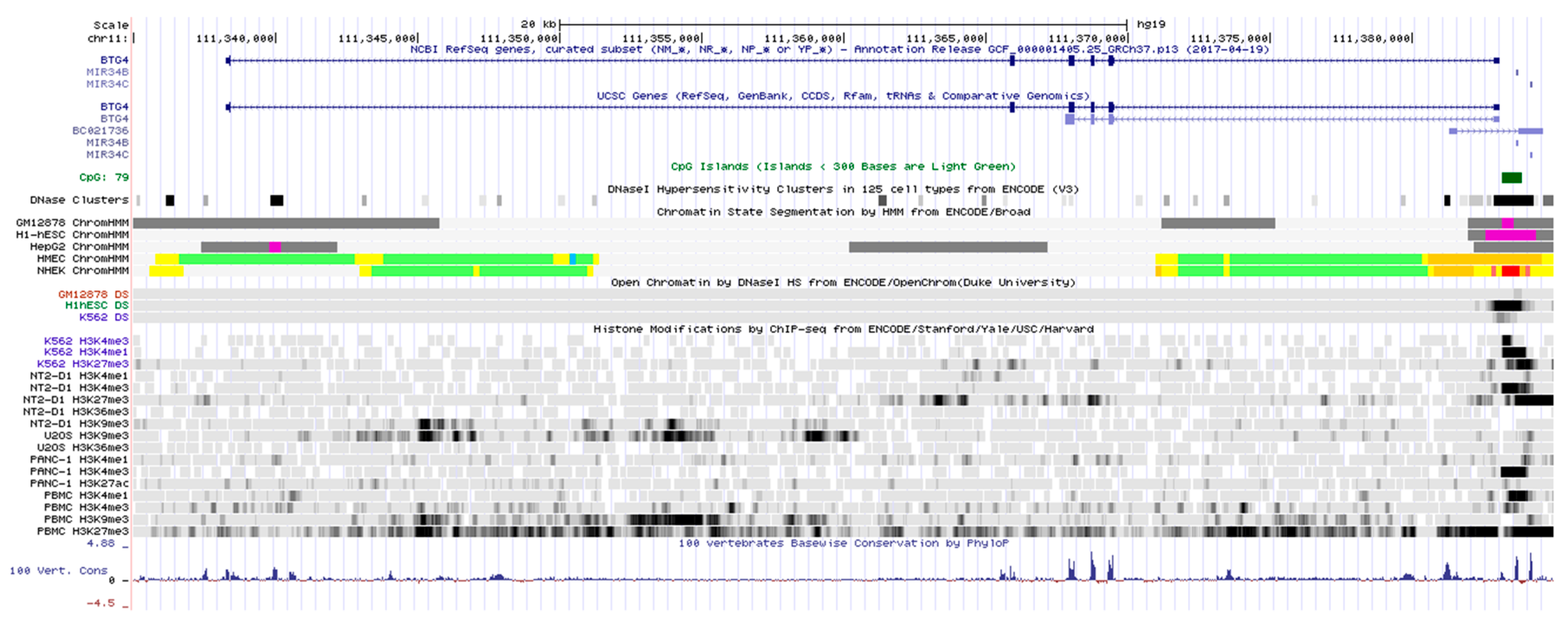
3.1. Gene Organization and Chromatin State of the BTG4 Gene Locus
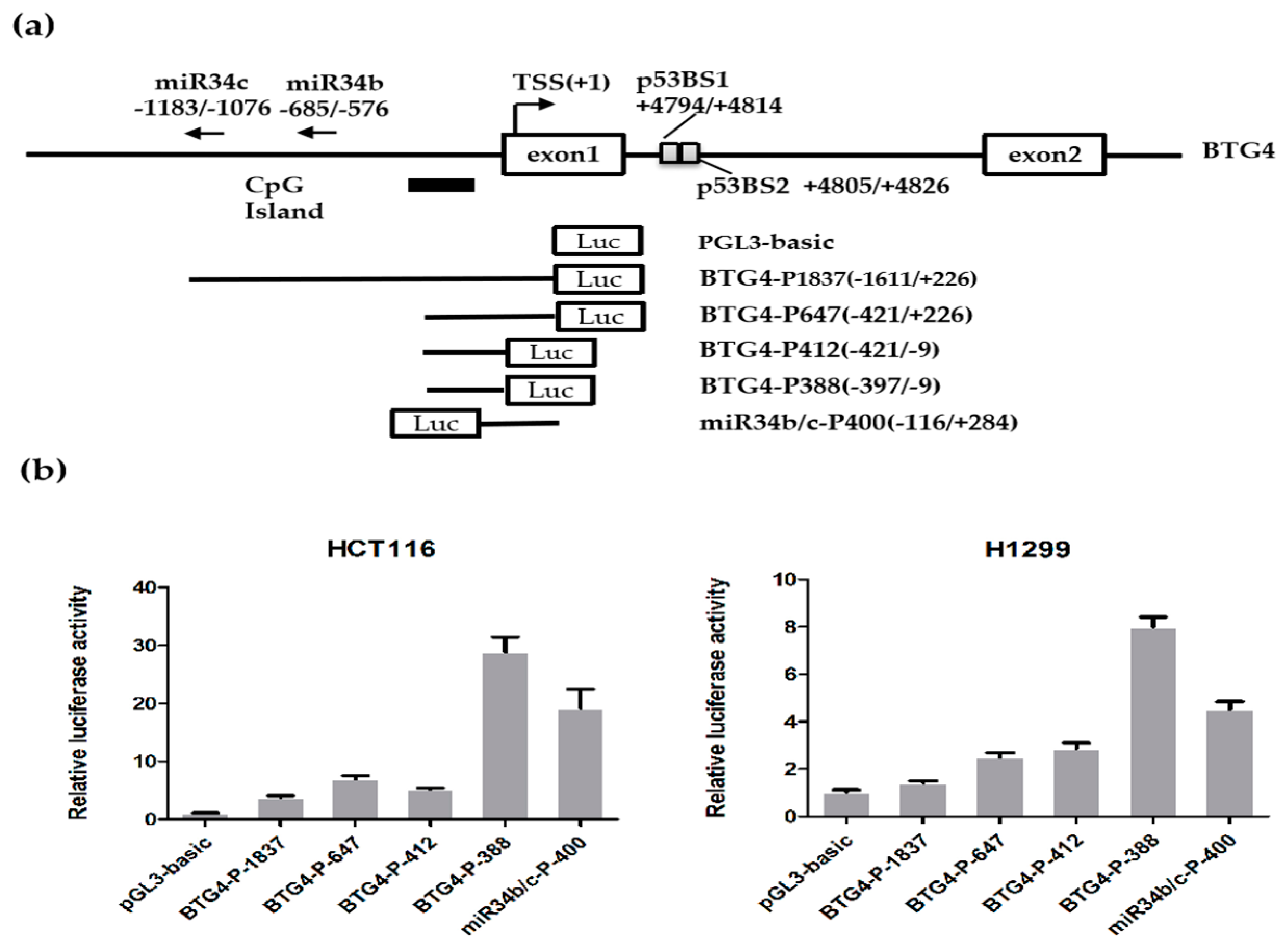
3.2. Identification of the BTG4 Promoter Region
3.3. Sequence Analysis of the BTG4 Promoter Region
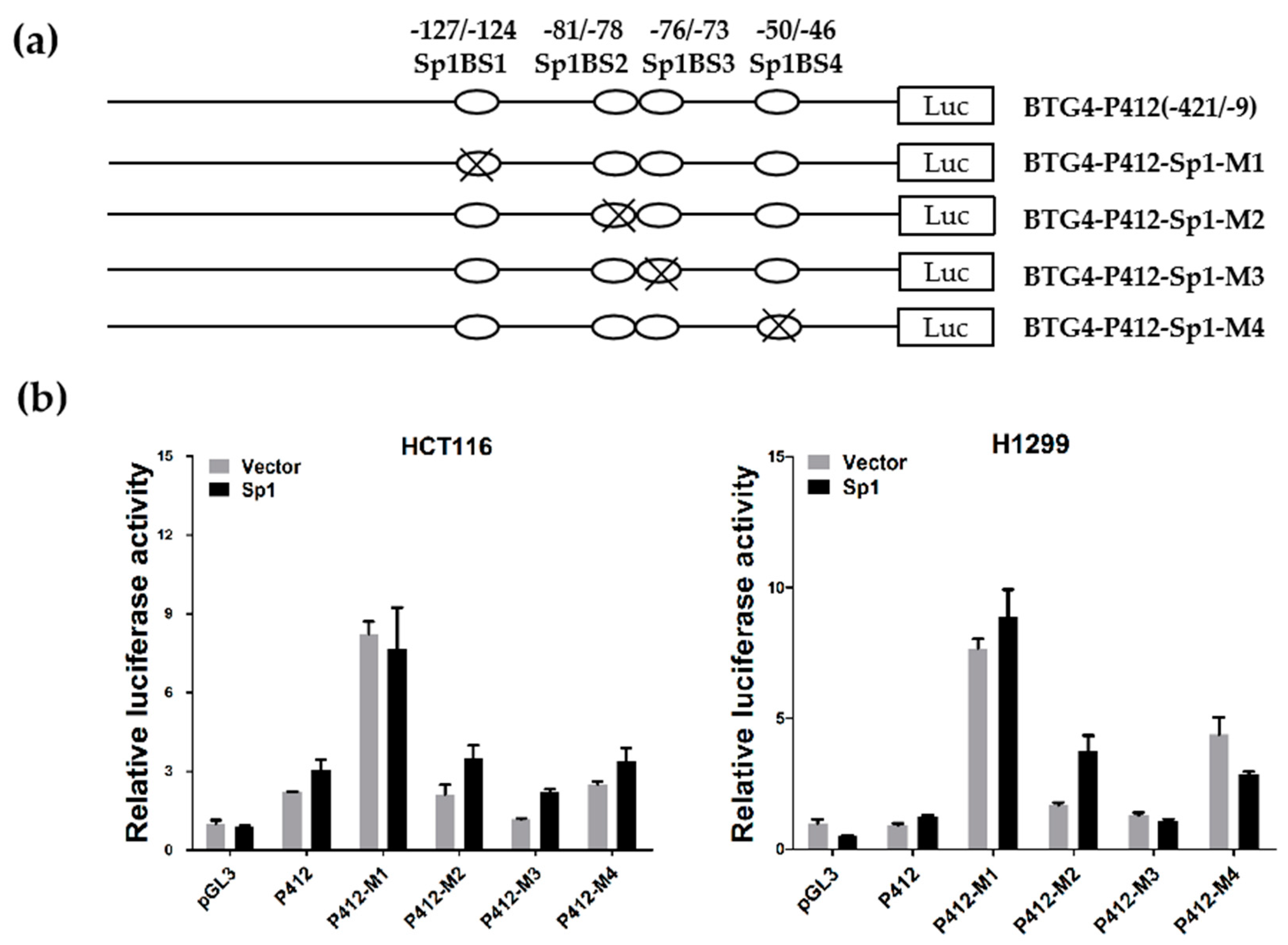
3.4. Functional Analysis of Sp1 Binding Sites in the BTG4 Gene Promoter
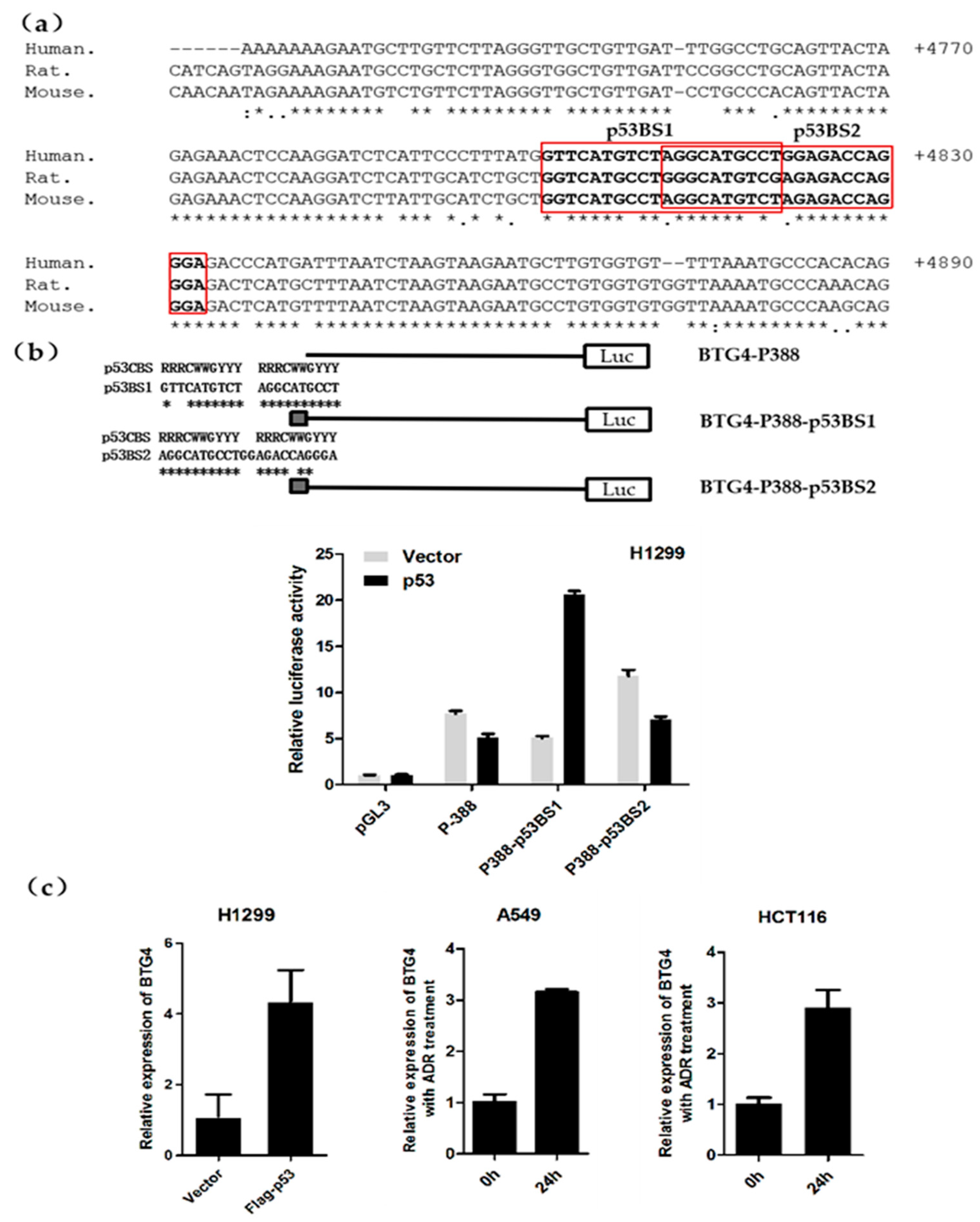
3.5. Identification of a p53-Responsive Element with the First Intron of the BTG4 Gene
3.6. P53 Induces BTG4 Transcription
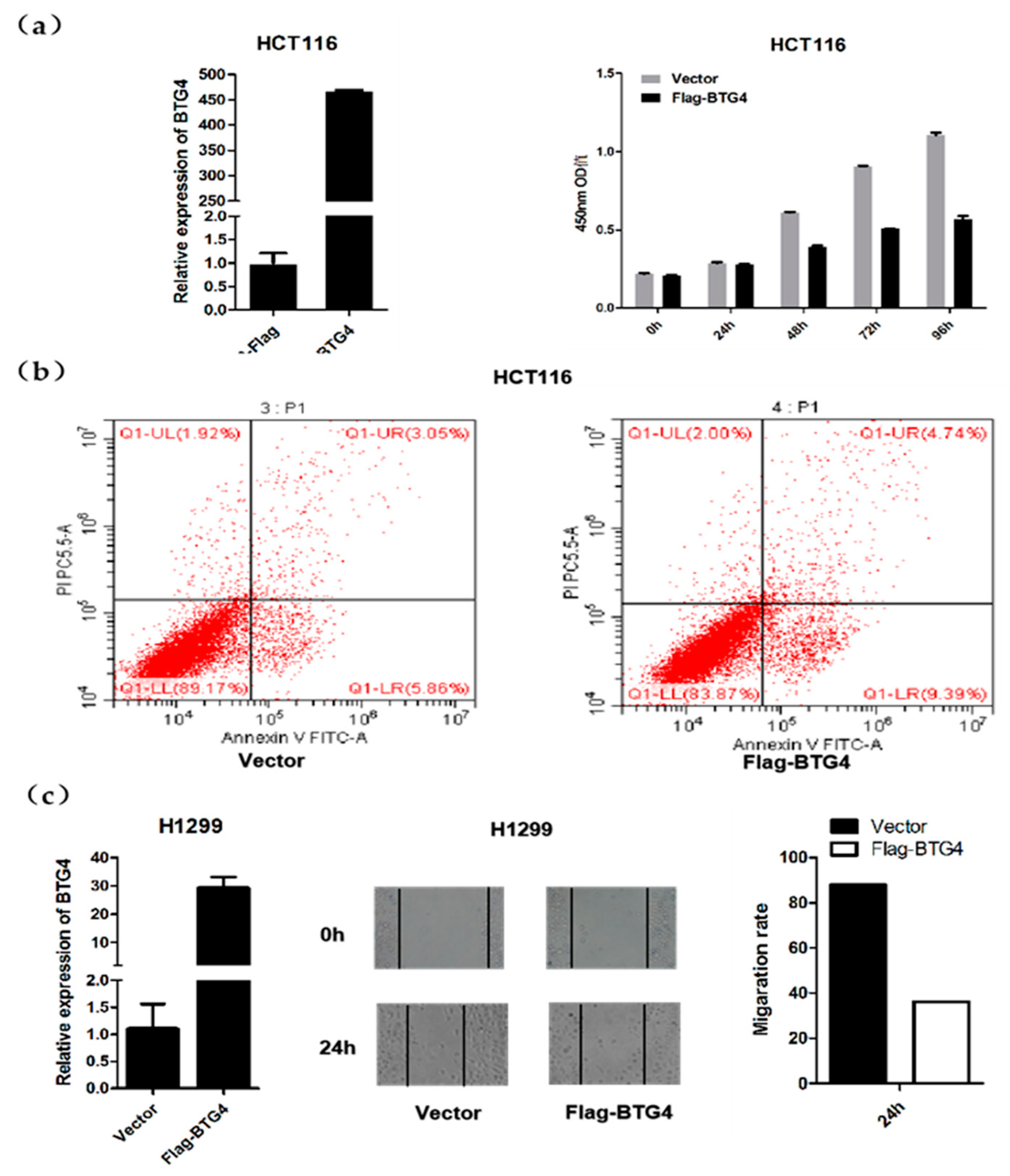
3.7. BTG4 Inhibits Cell Proliferation and Migration, and Induces Apoptosis
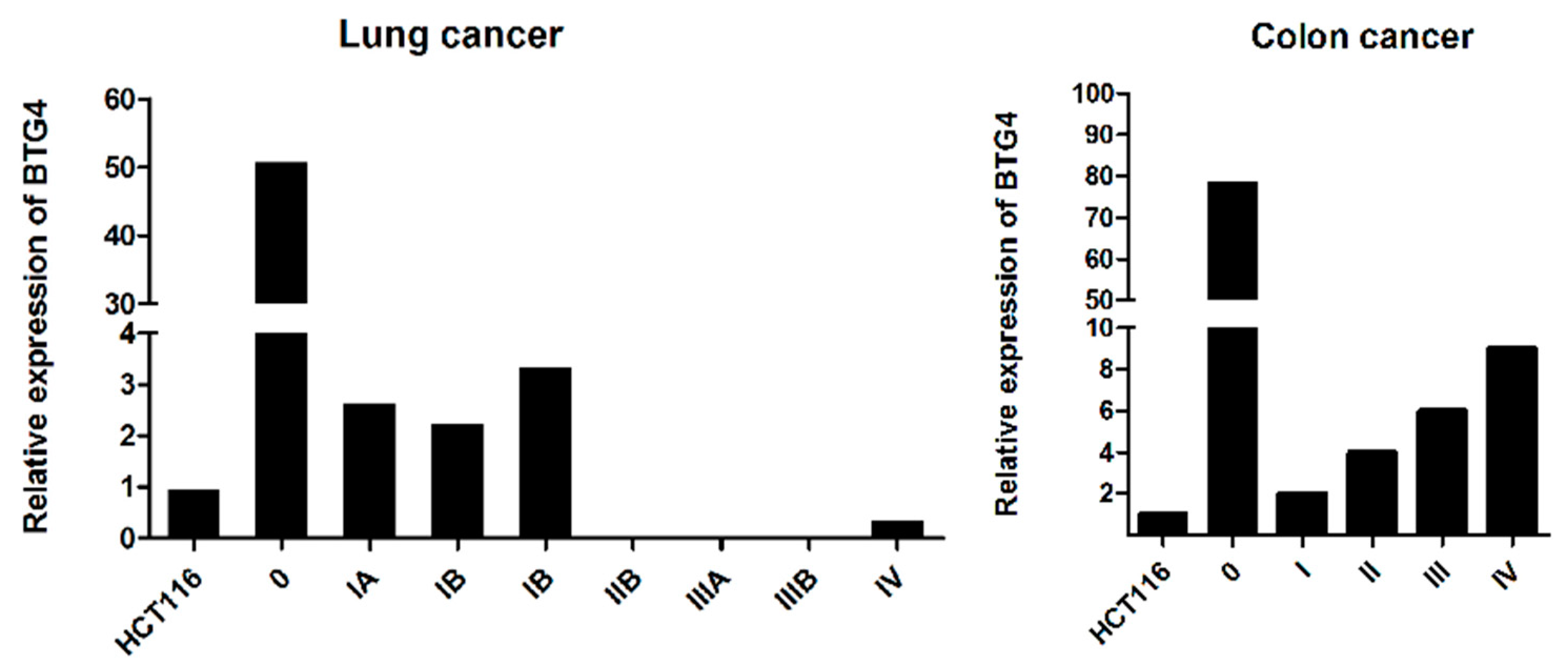
3.8. The Expression of BTG4 in Normal and Cancer Tissue
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Winkler, G. The mammalian anti-proliferative BTG/Tob protein family. J. Cell. Physiol. 2010, 222, 66–72. [Google Scholar] [CrossRef]
- Yuniati, L.; Scheijen, B.; Meer, L.T.; Leeuwen, F. Tumor suppressors BTG1 and BTG2: Beyond growth control. J. Cell. Physiol. 2019, 234, 5379–5389. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, A.; Possenti, R.; Shooter, E.; Tirone, F. Molecular cloning of PC3, a putatively secreted protein whose mRNA is induced by nerve growth factor and depolarization. Proc. Natl. Acad. Sci. USA 1991, 8, 3353–3357. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, B.; Lim, R.W.; Varnum, B.; Kujubu, D.; Koski, R.A.; Herschman, H. Structure and expression of TIS21, a primary response gene induced by growth factors and tumor promoters. J. Biol. Chem. 1991, 266, 14511–14518. [Google Scholar]
- Prevot, D.; Voeltzel, T.; Birot, A.-M.; Morel, A.-P.; Rostan, M.-C.; Magaud, J.-P.; Corbo, L. The Leukemia-associated Protein Btg1 and the p53-regulated Protein Btg2 Interact with the Homeoprotein Hoxb9 and Enhance Its Transcriptional Activation. J. Biol. Chem. 2000, 275, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Ou, Y.; Chung, P.; Hsu, F.; Sun, T.; Chang, W.; Shieh, S. The candidate tumor suppressor BTG3 is a transcriptional target of p53 that inhibits E2F1. EMBO J. 2007, 26, 3968–3980. [Google Scholar] [CrossRef] [Green Version]
- Busson, M.; Carazo, A.; Seyer, P.; Grandemange, S.; Casas, F.; Pessemesse, L.; Rouault, J.; Wrutniak-Cabello, C.; Cabello, G. Coactivation of nuclear receptors and myogenic factors induces the major BTG1 influence on muscle differentiation. Oncogene 2005, 24, 1698–1710. [Google Scholar] [CrossRef] [Green Version]
- Rouault, J.; Prevot, D.; Berthet, C.; Birot, A.; Billaud, M.; Magaud, J.; Corbo, L. Interaction of BTG1 and p53-regulatedBTG2 gene products with Mcaf1, the murine homolog of a component of the yeast CCR4 transcriptional regulatory complex. J. Biol. Chem. 1998, 273, 22563–22569. [Google Scholar] [CrossRef] [Green Version]
- Ikematsu, N.; Yoshida, Y.; Kawamura-Tsuzuku, J.; Ohsugi, M.; Onda, M.; Hirai, M.; Fujimoto, J.; Yamamoto, T. Tob2, a novel anti-proliferative Tob/BTG1 family member, associates with a component of the CCR4 transcriptional regulatory complex capable of binding cyclin-dependent kinases. Oncogene 1999, 18, 7432–7441. [Google Scholar] [CrossRef] [Green Version]
- Prévôt, D.; Morel, A.; Voeltzel, T.; Rostan, M.; Rimokh, R.; Magaud, J.; Corbo, L. Relationships of the antiproliferative proteins BTG1 and BTG2 with CAF1, the human homolog of a component of the yeast CCR4 transcriptional complex. J. Biol. Chem. 2001, 276, 9640–9648. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T. Phosphorylation of three regulatory serines of Tob by Erk1 and Erk2 is requird for Ras-mediated cell proliferation and transformation. Genes 2002, 16, 356–1370. [Google Scholar]
- Yoshida, Y.; Nakamura, T.; Komoda, M.; Satoh, H.; Suzuki, T.; Tsuzuku, J.; Miyasaka, T.; Yoshida, E.; Umemori, H.; Kunisaki, R. Mice lacking a transcriptional corepressor Tob are predisposed to ancer. Genes 2003, 17, 11201–11206. [Google Scholar]
- Boiko, A.; Porteous, S.; Razorenova, O.; Krivokrysenko, V.; Williams, B.; Gudkov, A. A systematic seach for downstream mediators of tumor suppressor function of p53 reveals a major role of BTG2 in suppression of Ras-induced transformation. Genes 2006, 20, 236–252. [Google Scholar] [CrossRef] [Green Version]
- Yoneda, M.; Suzuki, T.; Nakamura, T.; Ajima, R.; Tadashi, Y. Deficiency of antiproliferative family protein Ana correlates with development of lung adenocarcinoma. Cancer Sci. 2009, 100, 225–232. [Google Scholar] [CrossRef]
- Majid, S.; Dar, A.; Ahmad, A.; Hirata, H.; Kawakami, K.; Shahryari, V.; Saini, S.; Tanaka, Y.; Dahiya, A.; Khatri, G. BTG3 tumor suppressor gene promoter demethylation, histone modification and cell cycle arrest by genistein in renal cancer. Carcinogenesis 2009, 30, 662–670. [Google Scholar] [CrossRef] [Green Version]
- Lv, C.; Wang, H.; Tong, Y.; Yin, H.; Wang, D.; Yan, Z.; Liang, Y.; Wu, D.; Su, Q. The function of BTG3 in colorectal cancer cells and its possible signaling pathway. J. Cancer Res. Clin. Oncol. 2018, 144, 295–308. [Google Scholar] [CrossRef] [Green Version]
- Buanne, P.; Corrente, G.; Micheli, L.; Palena, A.; Lavia, P.; Spadafora, C.; Lakshmana, M.; Rinaldi, A.; Banfi, S.; Quarto, M.; et al. Cloning of PC3B, a novel member of the PC3/BTG/TOB family of growth inhibitory genes, highly expressed in the olfactory epithelium. Genomics 2000, 68, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Dean, J. BTG4, a maternal mRNA cleaner. J. Mol. Cell Biol. 2016, 8, 369–370. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Lu, X.; Shi, J.; Yu, X.; Zhang, X.; Zhu, K.; Yi, Z.; Duan, E.; Li, L. BTG4 is a key regulator for maternal mRNA clearance during mouse early embryogenesis. J. Mol. Cell Biol. 2016, 8, 366–368. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Ji, S.; Sha, Q.; Dang, Y.; Zhou, J.; Zhang, Y.; Liu, Y.; Wang, Z.; Hu, B.; Sun, Q.; et al. BTG4 is a meiotic cell cycle-coupled maternal-zygotic-transition licensing factor in oocytes. Nat. Struct. Mol. Biol. 2016, 23, 387–394. [Google Scholar] [CrossRef]
- Dong, W.; Tu, S.; Xie, J.; Sun, P.; Wu, Y.; Wang, L. Frequent promoter hypermethylation and transcriptional downregulation of BTG4 gene in gastric cancer. Biochem. Biophys. Res. Commun. 2009, 387, 132–138. [Google Scholar] [CrossRef]
- Aue, R.; Starczynski, J.; McElwaine, S.; Bertoni, F.; Newland, A.; Fegan, C.; Cotter, F. Identification of a potential role for POU2AF1 and BTG4 in the deletion of 11q23 in chronic lymphocytic leukemia. Genes Chromosomes Cancer 2005, 43, 1–10. [Google Scholar]
- Toyot, M.; Suzuki, H.; Sasaki, Y.; Maruyama, R.; Imai, K.; Shinomura, Y.; Tokino, T. Epigenetic silencing ofmicroRNA-34b/c and B-cell translocation gene 4 is associated with CpG island methylation in colorectal cancer. Cancer Res. 2008, 68, 4123–4132. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, Y.; Zhang, C.; Weng, H.; Li, Y.; Cai, W.; Xie, M.; Long, Y.; Ai, Q.; Liu, Z.; et al. The gene pair PRR11 and SKA2 shares a NF-Y-regulated bidirectional promoter and contributes to lung cancer development. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2015, 1849, 1133–1144. [Google Scholar] [CrossRef]
- Hu, L.; Chen, Q.; Wang, Y.; Zhang, N.; Meng, P.; Liu, T.; Bu, Y. Sp1 Mediates the constitutive expression and repression of the PDSS2 gene in lung cancer cells. Genes 2019, 10, 977. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Xie, M.; Lan, H.; Zhang, Y.; Long, Y.; Weng, H.; Li, D.; Cai, W.; Zhu, H.; Niu, Y.; et al. PRR11 is a novel gene implicated in cell cycle progression and lung cancer. Int. J. Biochem. Cell Biol. 2013, 45, 645–656. [Google Scholar] [CrossRef]
- Bieging, K.; Mello, S.; Attardi, L. Unravelling mechanisms of p53-mediated tumour suppression. Nat. Rev. Cancer 2014, 14, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, C.; Jin, Y.; He, Q.; Liu, Z.; Ai, Q.; Lei, Y.; Li, Y.; Song, F.; Bu, Y. Alkaline ceramidase 2 is a novel direct target of p53 and induces autophagy and apoptosis through ROS generation. Sci. Rep. 2017, 7, 44573. [Google Scholar] [CrossRef]
- Wang, Y.; Weng, H.; Zhang, Y.; Long, Y.; Li, Y.; Niu, Y.; Song, F.; Bu, Y. The PRR11-SKA2 Bidirectional Transcription Unit Is Negatively Regulated by p53 through NF-Y in Lung Cancer Cells. Int. J. Mol. Sci. 2017, 18, 534. [Google Scholar] [CrossRef] [Green Version]
- Rouault, J.; Falette, N.; Guéhenneux, F.; Guillot, C.; Rimokh, R.; Wang, Q.; Berthet, C.; Moyret-Lalle, C.; Savatier, P.; Pain, B.; et al. Identification of BTG2, an antiproliferative p53-dependent component of the DNA damage cellular response pathway. Nat. Genet. 1996, 14, 482–486. [Google Scholar] [CrossRef]
- Hermeking, H. The miR-34 family in cancer and apoptosis. Cell Death Differ. 2010, 17, 193–199. [Google Scholar] [CrossRef]
- Yaman, K.; Tateishi, K.; Klose, R.; Fang, J.; Fabrizio, L.; Erdjument-Bromage, H.; Taylor-Papadimitriou, J.; Tempst, P.; Zhang, Y. PLU-1 is an H3K4 demethylase involved in transcriptional repression and breast cancer cell proliferation. Mol. Cell. 2007, 25, 801–812. [Google Scholar] [CrossRef]
- Scibetta, A.; Santangelo, S.; Coleman, J.; Hall, D.; Chaplin, T.; Copier, J.; Catchpole, S.; Burchell, J.; Taylor-Papadimitriou, J. Functional analysis of the transcription repressor PLU-1/JARID1B. Mol. Cell Biol. 2007, 27, 7220–7235. [Google Scholar] [CrossRef] [Green Version]
- Gou, D.; Wang, J.; Gao, L.; Sun, Y.; Pe, X.; Huang, J.; Li, W. Identification and functional analysis of a novel human KRAB/C2H2 zinc finger gene ZNF300. Biochim. Biophys. Acta 2004, 20, 203–209. [Google Scholar] [CrossRef]
- Sakai, T.; Hino, K.; Wada, S.; Maeda, H. Identification of the DNA binding specificity of the human ZNF219 protein and its function as a transcriptional repressor. DNA Res. 2003, 10, 155–165. [Google Scholar] [CrossRef]
- Liu, W.; Wang, H.; Muguira, M.; Breslin, M.; Lan, M. INSM1 functions as a transcriptional repressor of the neuroD/beta2 gene through the recruitment of cyclin D1 and histone deacetylases. Biochem. J. 2006, 397, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Guo, P.; Lee, C.; Lei, H.; Zheng, Y.; Prieto, P.K.D.; Pan, D. Nerfin-1 represses transcriptional output of Hippo signaling in cell competition. Elife 2019, 8, e38843. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Constructs | Primer Sequences and Templates |
|---|---|
| BTG4-P1837(−1611/+226) | F:5′-GCTAGCCCGGGCTCGAGATCTACTGTGGCAGGAACTGAGATGGA-3′ Bgl II R:5′-CAGTACCGGAATGCCAAGCTTGCTACCCAAGCCCACCTCCATTT-3′ Hind III PCR Template: human genomic DNA |
| BTG4-P647(−421/+226) | F:5′-GCACAACACGCGTAGCTGGGCCA-3′ R:5′-GGTACCTATCGATAGAGAAATGTTCTGGC-3′ Kpn I PCR Template: BTG4-P1837 |
| BTG4-P412 (−421/−9) | F:5′-AAGCTTGGCATTCCGGTACTGTTGGTAA-3′ Hind III R:5′-CAGCGCTTTCTCTCAGCCTCCTCC-3′ PCR Template: BTG4-P647 |
| BTG4-P388(−397/−9) | F:5′-GGGCGGCGCTGCCGGGGAGGA-3′ R:5′-GGTACCTATCGATAGAGAAATGTTCTGGC-3′ Kpn I PCR Template: BTG4-P412 |
| BTG4-P388-p53BS2 | F:5′-AGGCATGCCTGGAGACCAGGGAGCACAACACGCGTAGCTGGGCCA-3′ R:5′-GGTACCTATCGATAGAGAAATGTTCTGGC-3′ Kpn I PCR Template: BTG4-P388 |
| BTG4-P388-p53BS1 | F:5′-GTTCATGTCTAGGCATGCCTGGGCGGCGCTGCCGGGGAGGA-3′ R:5′-GGTACCTATCGATAGAGAAATGTTCTGGC-3′ Kpn I PCR Template: BTG4-P388 |
| BTG4-P412-Sp1-M1 | F:5′-TCTACTCCAACCGTCCTTGGAACCC-3′ R:5′-CCGCCACAGTCACTCGGCCGCTCAG-3′ PCR Template: BTG4-P412 |
| BTG4-P412-Sp1-M2 | F:5′-TTGAGGGGCGGGCGCGAGACGGGGC-3′ R:5′-CTCGGCCCGCGGGGTTCCAAGGACG-3′ PCR Template: BTG4-P412 |
| BTG4-P412-Sp1-M3 | F:5′-CTATGGGCGCGAGACGGGGCGGGTG-3′ R:5′-CGCCCCTCGGCCCGCGGGGTTCCAA-3′ PCR Template: BTG4-P412 |
| BTG4-P412-Sp1-M4 | F:5′-TTCGGGGATGGGGGGGGGGAGGAGG-3′ R:5′-CCCACCCGCCCCGTCTCGCGCCCGC-3′ PCR Template: BTG4-P412 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, N.; Jiang, T.; Wang, Y.; Hu, L.; Bu, Y. BTG4 is A Novel p53 Target Gene That Inhibits Cell Growth and Induces Apoptosis. Genes 2020, 11, 217. https://doi.org/10.3390/genes11020217
Zhang N, Jiang T, Wang Y, Hu L, Bu Y. BTG4 is A Novel p53 Target Gene That Inhibits Cell Growth and Induces Apoptosis. Genes. 2020; 11(2):217. https://doi.org/10.3390/genes11020217
Chicago/Turabian StyleZhang, Na, Tinghui Jiang, Yitao Wang, Lanyue Hu, and Youquan Bu. 2020. "BTG4 is A Novel p53 Target Gene That Inhibits Cell Growth and Induces Apoptosis" Genes 11, no. 2: 217. https://doi.org/10.3390/genes11020217
APA StyleZhang, N., Jiang, T., Wang, Y., Hu, L., & Bu, Y. (2020). BTG4 is A Novel p53 Target Gene That Inhibits Cell Growth and Induces Apoptosis. Genes, 11(2), 217. https://doi.org/10.3390/genes11020217