Population Genetic Structure Is Unrelated to Shell Shape, Thickness and Organic Content in European Populations of the Soft-Shell Clam Mya Arenaria

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
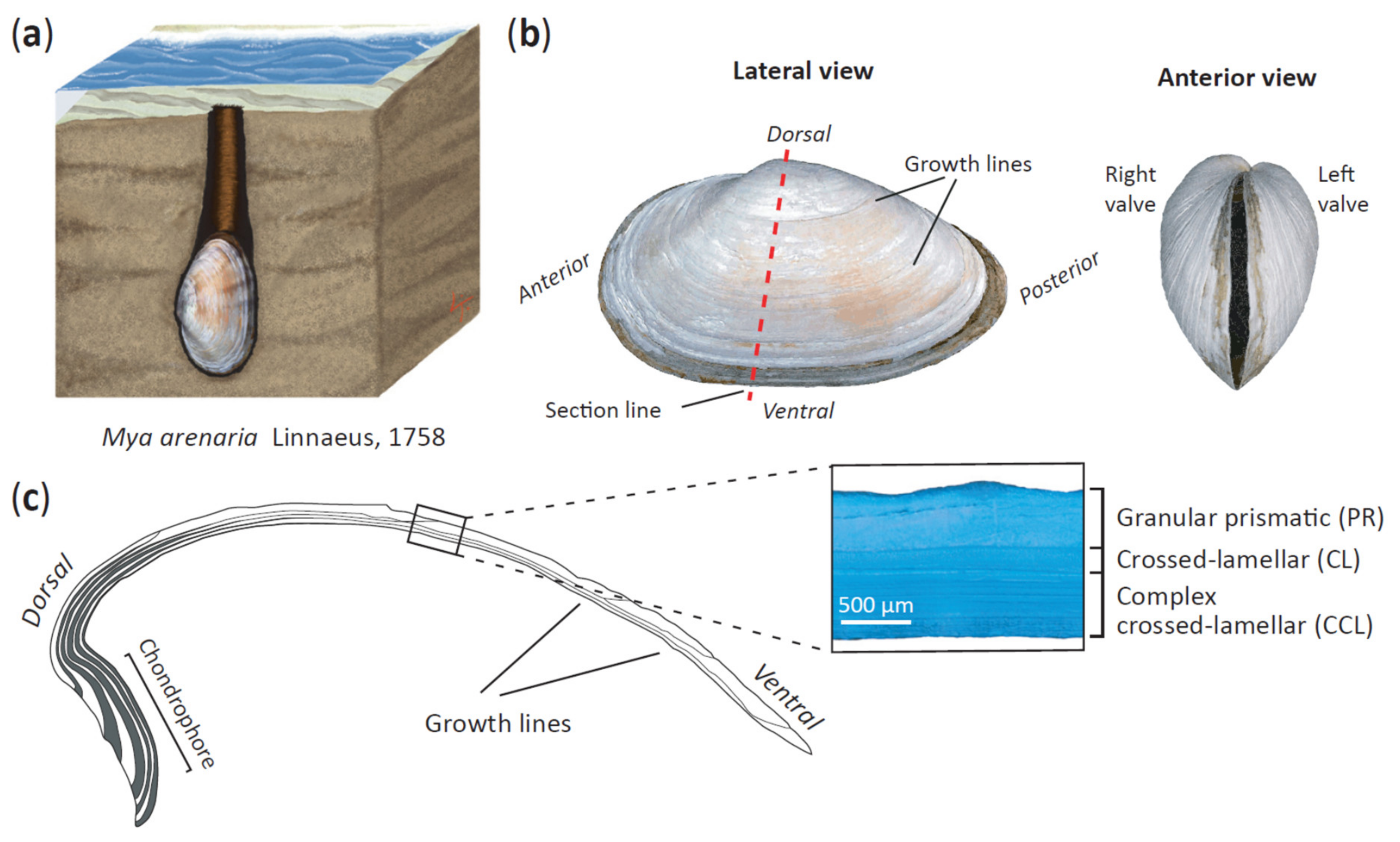
2.1. Sample Collection and DNA Extraction
2.2. Microsatellite Genotyping
2.3. Genetic Summary Statistics
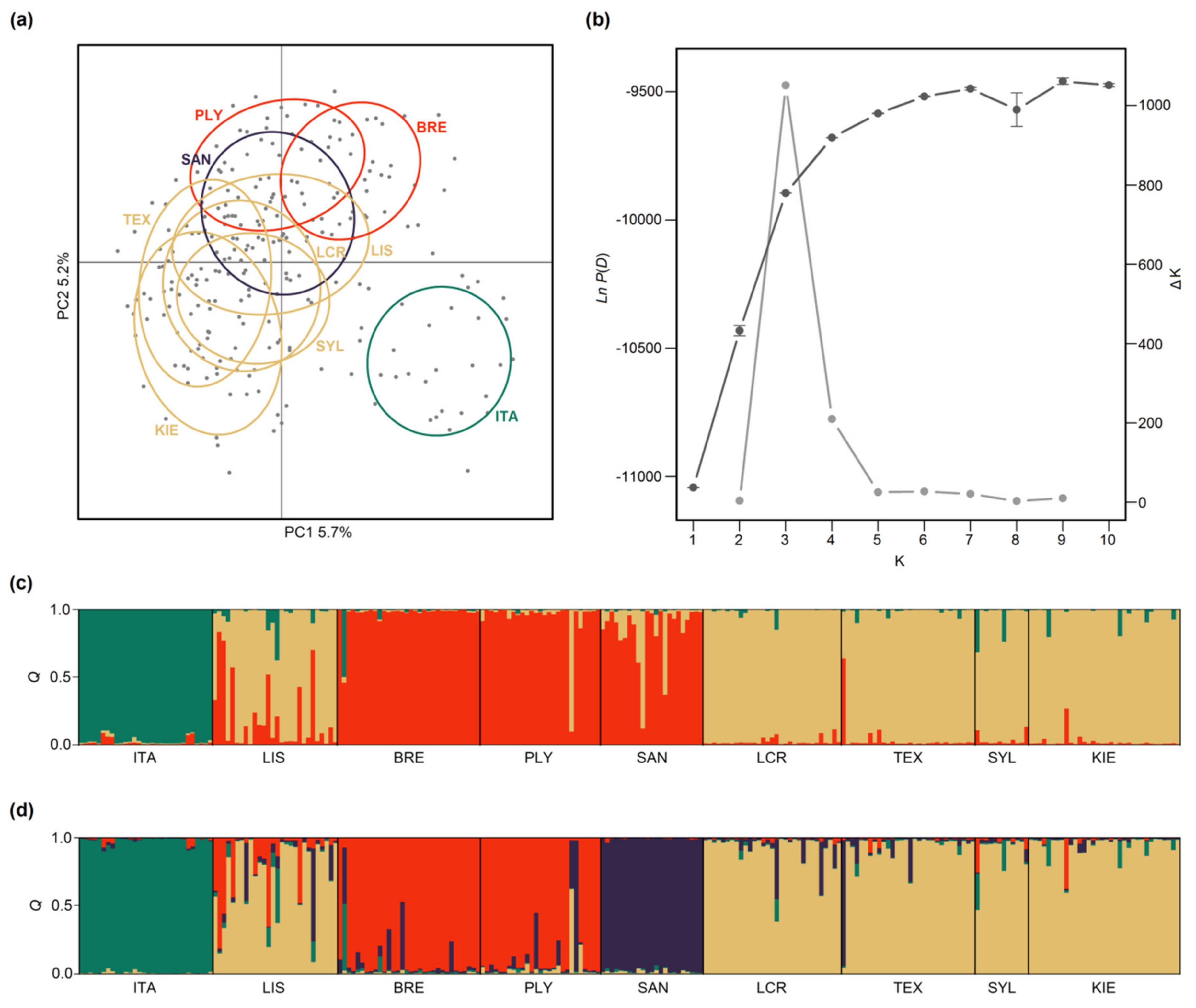
2.4. Analysis of Population Structure
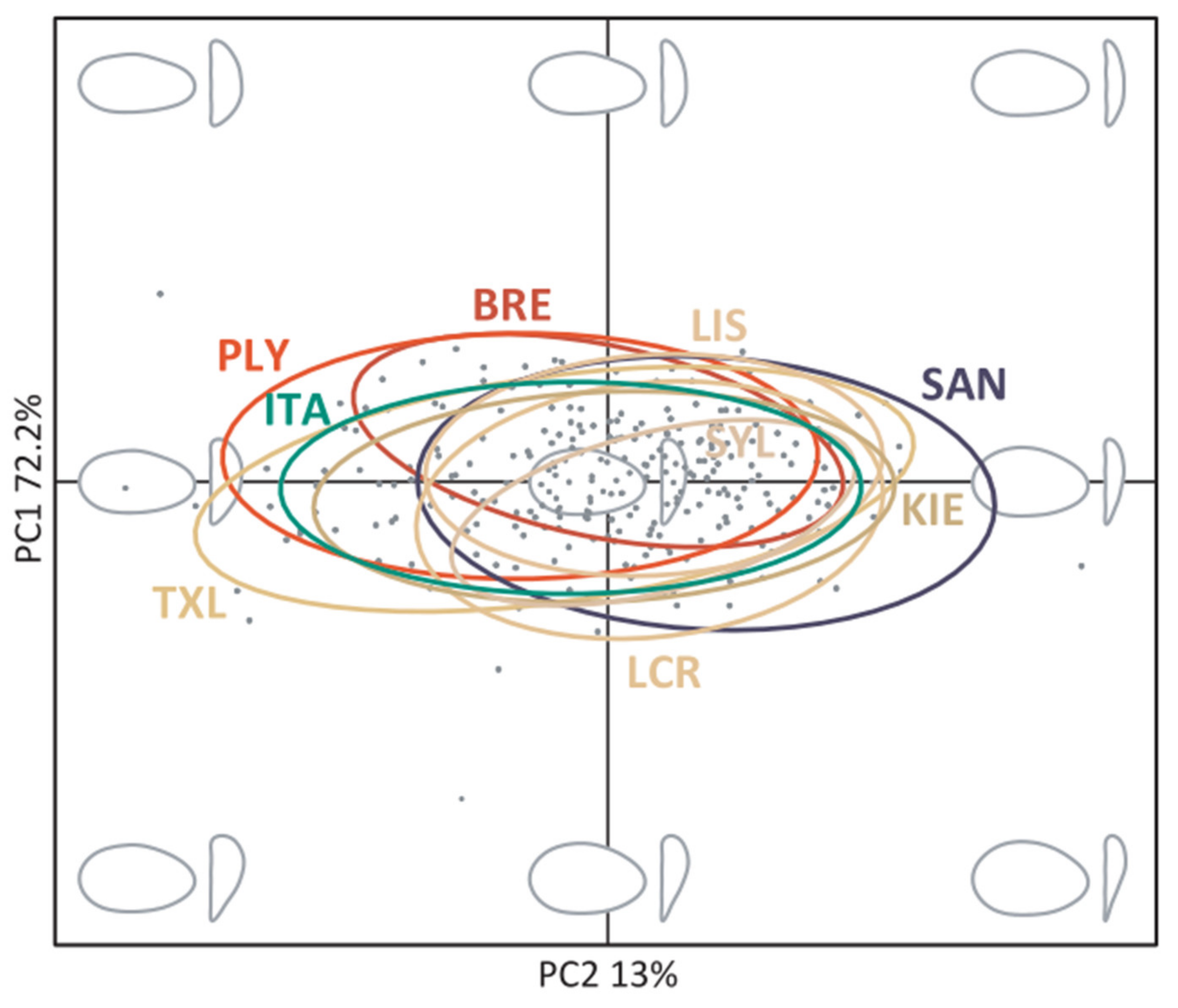
2.5. Elliptic Fourier Analysis of Shell Outlines
2.6. Analysis of Shell Thickness
2.7. Analysis of Organic Shell Content
2.8. Data accessibility
3. Results
3.1. Population Structure
3.2. Shell Shape Variation
3.3. Variation in Shell Thickness
3.4. Variation in Organic Content
4. Discussion
4.1. Population Genetic Structure
4.2. Shell Shape Variation
4.3. Variation in Shell Thickness, Microstructure and Organic Content
4.4. Genetic Versus Plastic Contributions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Strasser, M. Mya arenaria—An ancient invader of the North Sea coast. Helgoländer Meeresunters. 1999, 52, 309. [Google Scholar] [CrossRef] [Green Version]
- Hanna, G.D. Introduced mollusks of western North America. Occ. Pap. Calif. Acad. Sci. 1966, 48, 1–108. [Google Scholar]
- Carlton, J.T. Man’s role in changing the face of the ocean: Biological invasions and implications for conservation of near-shore environments. Conserv. Biol. 1989, 3, 265–273. [Google Scholar] [CrossRef]
- Leppäkoski, E. The Baltic and the Black Sea-Seriously contaminated by nonindigenous species? In Proceedings of the Conference and Workshop Nonindigenous Estuarine and Marine Organisms (NEMO), Seattle, WA, USA, April 1993; pp. 37–44. [Google Scholar]
- Lasota, R.; Pierscieniak, K.; Garcia, P.; Simon-Bouhet, B.; Wolowicz, M. Large-scale mitochondrial COI gene sequence variability reflects the complex colonization history of the invasive soft-shell clam, Mya arenaria (L.)(Bivalvia). Estuar. Coast. Shelf Sci. 2016, 181, 256–265. [Google Scholar] [CrossRef]
- Strauch, F. Phylogenese, Adaptation und Migration einiger nordischer-mariner Molluskengenera (Neptunea, Panomya,-Cyrtodaria und Mya). Abh. Senckenberg. Naturforsch. Ges. 1972, 531, 1–211. [Google Scholar]
- Hessland, I. On the quaternary Mya period in Europe. Ark. Zool. 1946, 37A, 1–51. [Google Scholar]
- Petersen, K.S.; Rasmussen, K.L.; Heinemeier, J.; Rud, N. Clams before Columbus? Nature 1992, 359, 679. [Google Scholar] [CrossRef]
- Essink, K.; Oost, A.P. How did Mya arenaria (Mollusca; Bivalvia) repopulate European waters in mediaeval times? Mar. Biodivers. 2019, 49, 1–10. [Google Scholar] [CrossRef]
- Reise, K.; Gollasch, S.; Wolff, W.J. Introduced marine species of the North Sea coasts. Helgoländer Meeresunters. 1998, 52, 219. [Google Scholar] [CrossRef] [Green Version]
- Crocetta, F.; Turolla, E. Mya arenaria Linné, 1758 (Mollusca: Bivalvia) in the Mediterranean Sea: Its distribution revisited. J. Biol. Res. 2011, 16, 188–193. [Google Scholar]
- Zenetos, A.; Koutsoubas, D.; Vardala-Theodorou, E. Origin and vectors of introduction of exotic molluscs in Greek waters. Belg. J. Zool. 2004, 134, 161–168. [Google Scholar]
- Conde, A.; Novais, J.M.; Domínguez, J. The presence of Mya arenaria in the Ria de Aveiro is the third confirmed record of this invasive clam on the Portuguese coast. Mar. Biodivers. Rec. 2012, 5. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, D.; Le Loc’h, F.; Désaunay, Y.; Hamon, D.; Blanchet, A.; Le Pape, O. Relationships between benthic macrofauna and habitat suitability for juvenile common sole (Solea solea, L.) in the Vilaine estuary (Bay of Biscay, France) nursery ground. Estuar. Coast. Shelf Sci. 2007, 73, 639–650. [Google Scholar] [CrossRef] [Green Version]
- Hughes, R.G.; Lloyd, D.; Ball, L.; Emson, D. The effects of the polychaete Nereis diversicolor on the distribution and transplanting success of Zostera noltii. Helgol. Mar. Res. 2000, 54, 129. [Google Scholar] [CrossRef] [Green Version]
- Seaward, D.R. Distribution of the Marine Molluscs of North West Europe (No. 165); Joint Nature Conservation Committee: Peterborough, UK, 1990. [Google Scholar]
- Ysebaert, T.; Meire, P.; Coosen, J.; Essink, K. Zonation of intertidal macrobenthos in the estuaries of Schelde and Ems. Aquat. Ecol. 1998, 32, 53–71. [Google Scholar] [CrossRef]
- Maximovich, N.V.; Guerassimova, A.V. Life history characteristics of the clam Mya arenaria in the White Sea. Helgol. Mar. Res. 2003, 57, 91. [Google Scholar] [CrossRef]
- Vendrami, D.L.; Houston, R.D.; Gharbi, K.; Telesca, L.; Gutierrez, A.P.; Gurney-Smith, H.; Hoffman, J.I. Detailed insights into pan-European population structure and inbreeding in wild and hatchery Pacific oysters (Crassostrea gigas) revealed by genome-wide SNP data. Evol. Appl. 2019, 12, 519–534. [Google Scholar] [CrossRef] [Green Version]
- Vendrami, D.L.; De Noia, M.; Telesca, L.; Handal, W.; Charrier, G.; Boudry, P.; Hoffman, J.I. RAD sequencing sheds new light on the genetic structure and local adaptation of European scallops and resolves their demographic histories. Sci. Rep. 2019, 9, 7455. [Google Scholar] [CrossRef]
- Zbawicka, M.; Drywa, A.; Śmietanka, B.; Wenne, R. Identification and validation of novel SNP markers in European populations of marine Mytilus mussels. Mar. Biol. 2012, 159, 1347–1362. [Google Scholar] [CrossRef]
- Launey, S.; Ledu, C.; Boudry, P.; Bonhomme, F.; Naciri-Graven, Y. Geographic structure in the European flat oyster (Ostrea edulis L.) as revealed by microsatellite polymorphism. J. Hered. 2002, 93, 331–351. [Google Scholar] [CrossRef]
- Daguin, C.; Bonhomme, F.; Borsa, P. The zone of sympatry and hybridization of Mytilus edulis and M. galloprovincialis, as described by intron length polymorphism at locus mac-1. Heredity 2001, 86, 342. [Google Scholar] [CrossRef] [PubMed]
- Strasser, C.A.; Barber, P.H. Limited genetic variation and structure in softshell clams (Mya arenaria) across their native and introduced range. Conserv. Genet. 2009, 10, 803. [Google Scholar] [CrossRef] [Green Version]
- St-Onge, P.; Sévigny, J.M.; Strasser, C.; Tremblay, R. Strong population differentiation of softshell clams (Mya arenaria) sampled across seven biogeographic marine ecoregions: Possible selection and isolation by distance. Mari. Biol. 2013, 160, 1065–1081. [Google Scholar] [CrossRef]
- Lasota, R.; Hummel, H.; Wolowicz, M. Genetic diversity of European populations of the invasive soft-shell clam Mya arenaria (Bivalvia). J. Mar. Biol. Assoc. U.K 2004, 84, 1051–1056. [Google Scholar] [CrossRef] [Green Version]
- Cross, M.E.; Bradley, C.R.; Cross, T.F.; Culloty, S.; Lynch, S.; McGinnity, P.; Prodöhl, P.A. Genetic evidence supports recolonisation by Mya arenaria of western Europe from North America. Mar. Ecol. Prog. Ser. 2016, 549, 99–112. [Google Scholar] [CrossRef] [Green Version]
- Krapal, A.M.; Popa, O.P.; Iorgu, E.I.; Costache, M.; Popa, L.O. Isolation and characterization of new microsatellite markers for the invasive softshell clam, Mya arenaria (L.)(Bivalvia: Myidae). Int. J. Mol. Sci. 2012, 13, 2515–2520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Zhang, H.; Wang, A.; Lu, M.; Shen, Z.; Lian, C. Phenotypic plasticity accounts for most of the variation in leaf manganese concentrations in Phytolacca americana growing in manganese-contaminated environments. Plant Soil 2015, 396, 215–227. [Google Scholar] [CrossRef]
- Zieritz, A.; Hoffman, J.I.; Amos, W.; Aldridge, D.C. Phenotypic plasticity and genetic isolation-by-distance in the freshwater mussel Unio pictorum (Mollusca: Unionoida). Evol. Ecol. 2010, 24, 923–938. [Google Scholar] [CrossRef]
- Telesca, L.; Michalek, K.; Sanders, T.; Peck, L.S.; Thyrring, J.; Harper, E.M. Blue mussel shell shape plasticity and natural environments: A quantitative approach. Sci. Rep. 2018, 8, 2865. [Google Scholar] [CrossRef] [Green Version]
- Bonhomme, V.; Picq, S.; Gaucherel, C.; Claude, J. Momocs: Outline analysis using R. J. Stat. Softw. 2014, 56, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Schone, B.R.; Dunca, E.; Fiebig, J.; Pfeiffer, M. Mutvei’s solution: An ideal agent for resolving microgrowth structures of biogenic carbonates. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 228, 149–166. [Google Scholar] [CrossRef]
- Hoffman, J.I.; Peck, L.S.; Hillyard, G.; Zieritz, A.; Clark, M.S. No evidence for genetic differentiation between Antarctic limpet Nacella concinna morphotypes. Mar. Biol. 2010, 157, 765–778. [Google Scholar] [CrossRef]
- Trussell, G.C.; Etter, R.J. Integrating genetic and environmental forces that shape the evolution of geographic variation in a marine snail. In Microevolution rate, Pattern, Process; Springer: Dordrecht, The Netherlands, 2011; pp. 321–337. [Google Scholar]
- Vendrami, D.L.; Telesca, L.; Weigand, H.; Weiss, M.; Fawcett, K.; Lehman, K.; Hoffman, J.I. RAD sequencing resolves fine-scale population structure in a benthic invertebrate: Implications for understanding phenotypic plasticity. R. Soc. Open Sci. 2017, 4, 160548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shields, J.L.; Barnes, P.; Heath, D.D. Growth and survival differences among native, introduced and hybrid blue mussels (Mytilus spp.): Genotype, environment and interaction effects. Mar. Biol. 2008, 154, 919–928. [Google Scholar] [CrossRef]
- Fuentes, J.; Reyero, I.; Zapata, C.; Alvarez, G. Influence of stock and culture site on growth rate and mortality of mussels (Mytilus galloprovincialis Lmk.) in Galicia, Spain. Aquaculture 1992, 105, 131–142. [Google Scholar] [CrossRef]
- Dickie, L.M.; Boudreau, P.R.; Freeman, K.R. Influences of stock and site on growth and mortality in the blue mussel (Mytilus edulis). Can. J. Fish. Aquat. Sci. 1984, 41, 134–140. [Google Scholar] [CrossRef]
- Fitzer, S.C.; Vittert, L.; Bowman, A.; Kamenos, N.A.; Phoenix, V.R.; Cusack, M. Ocean acidification and temperature increase impact mussel shell shape and thickness: Problematic for protection? Ecol. Evol. 2015, 5, 4875–4884. [Google Scholar] [CrossRef] [Green Version]
- Illesca, A.; Oyarzún, P.A.; Toro, J.E.; Gardner, J.P.A. Morphometric variability of smooth-shelled blue mussels from the Pacific coast of South America. Biol. J. Linn. Soc. 2018, 20, 1–16. [Google Scholar] [CrossRef]
- Brönmark, C.; Lakowitz, T.; Hollander, J. Predator-induced morphological plasticity across local populations of a freshwater snail. PLoS ONE 2011, 6, e21773. [Google Scholar] [CrossRef]
- Swan, E.F. The growth of the clam Mya arenaria as affected by the substratum. Ecology 1852, 33, 530–534. [Google Scholar] [CrossRef]
- Emerson, C.W. Influence of sediment disturbance and water flow on the growth of the soft-shell clam, Mya arenaria L. Can. J. Fish. Aquat. Sci. 1990, 47, 1655–1663. [Google Scholar] [CrossRef]
- Bieler, R.; Mikkelsen, P.M.; Collins, T.M.; Glover, E.A.; González, V.L.; Graf, D.L.; Harper, E.M.; Healy, J.; Kawauchi, G.Y.; Sharma, P.P.; et al. Investigating the Bivalve Tree of Life—An exemplar-based approach combining molecular and novel morphological characters. Invertebr. Syst. 2014, 28, 32. [Google Scholar] [CrossRef] [Green Version]
- Palmer, A.R. Calcification in marine molluscs: How costly is it? Proc. Natl. Acad. Sci. USA 1992, 89, 1379–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, S.-A.; Morley, S.A.; Peck, L.S. Latitudinal trends in shell production cost from the tropics to the poles. Sci. Adv. 2017, 3, e1701362. [Google Scholar] [CrossRef] [Green Version]
- Leung, J.Y.S.; Russell, B.D.; Connell, S.D. Mineralogical plasticity acts as a compensatory mechanism to the impacts of ocean acidification. Environ. Sci. Technol. 2017, 51, 2652–2659. [Google Scholar] [CrossRef]
- Telesca, L.; Peck, L.S.; Sanders, T.; Thyrring, J.; Sejr, M.K.; Harper, E.M. Biomineralization plasticity and environmental heterogeneity predict geographical resilience patterns of foundation species to future change. Glob. Chang. Biol. 2019, 25, 4179–4193. [Google Scholar] [CrossRef] [PubMed]
- Filippenko, D.; Naumenko, E. Patterns of the growth of soft-shell clam Mya arenaria L. (Bivalvia) in shallow water estuaries of the southern Baltic Sea. Ecohydrol. Hydrobiol. 2014, 14, 157–165. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Suffolk County, NY, USA, 1989. [Google Scholar]
- Paradis, E. pegas: An R package for population genetics with an integrated–modular approach. Bioinformatics 2010, 26, 419–420. [Google Scholar] [CrossRef] [Green Version]
- Raymond, M. GENEPOP (version 1.2): Population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Rousset, F. genepop’007: A complete re-implementation of the genepop software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Keenan, K.; McGinnity, P.; Cross, T.F.; Crozier, W.W.; Prodöhl, P.A. diveRsity: An R package for the estimation and exploration of population genetics parameters and their associated errors. Methods Ecol. Evol. 2013, 4, 782–788. [Google Scholar] [CrossRef] [Green Version]
- Stoffel, M.A.; Esser, M.; Kardos, M.; Humble, E.; Nichols, H.; David, P.; Hoffman, J.I. inbreedR: An R package for the analysis of inbreeding based on genetic markers. Methods Ecol. Evol. 2016, 7, 1331–1339. [Google Scholar] [CrossRef]
- Schneider, S.; Roessli, D.; Excoffier, L. Arlequin version 2.000; A Software for Population Genetics Data Analysis; Genetics and Biometry Laboratory, University of Geneva: Geneva, Switzerland, 2000. [Google Scholar]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T.; Ahmed, I. adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Adams, D.C.; Rohlf, F.J.; Slice, D.E. Geometric morphometrics: Ten years of progress following the “revolution”. Ital. J. Zool. 2004, 71, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Kuhl, F.P.; Giardina, C.R. Elliptic Fourier features of a closed contour. Comput. Graph. Image Process. 1982, 18, 236–258. [Google Scholar] [CrossRef]
- Rohlf, F.J.; Archie, J.W. A comparison of Fourier methods for the description of wing shape in mosquitoes (Diptera: Culicidae). Syst. Zool. 1984, 33, 302. [Google Scholar] [CrossRef]
- Crampton, J.S. Elliptic Fourier shape analysis of fossil bivalves: Some practical considerations. Lethaia 1995, 28, 179–186. [Google Scholar] [CrossRef]
- Claude, J. Morphometrics with R; Springer: New York, NY, USA, 2008. [Google Scholar]
- Bookstein, F.L. Morphometric Tools for Landmark Data: Geometry and Biology; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Schielzeth, H. Simple means to improve the interpretability of regression coefficients. Methods Ecol. Evol. 2010, 1, 103–113. [Google Scholar] [CrossRef]
- Bolker, B.M. Linear and generalized linear mixed models. In Ecological Statistics; Fox, G.A., Negrete-Yankelevich, S., Sosa, V.J., Eds.; Oxford University Press: Oxford, UK, 2015; pp. 309–333. [Google Scholar]
- Côrte-Real, H.B.S.M.; Hawkins, S.J.; Thorpe, J.P. Population differentiation and taxonomic status of the exploited limpet Patella candei in the Macaronesian islands (Azores, Madeira, Canaries). Mar. Biol. 1996, 125, 141–152. [Google Scholar] [CrossRef]
- Almaça, C. Evolutionary and zoogeographical remarks on the Mediterranean fauna of brachyuran crabs. In Mediterranean Marine Ecosystems; Springer: Boston, MA, USA, 1985; pp. 347–366. [Google Scholar]
- Krakau, M.; Jacobsen, S.; Jensen, K.T.; Reise, K. The cockle Cerastoderma edule at Northeast Atlantic shores: Genetic signatures of glacial refugia. Mar. Biol. 2012, 159, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Maggs, C.A.; Castilho, R.; Foltz, D.; Henzler, C.; Jolly, M.T.; Kelly, J.; Viard, F. Evaluating signatures of glacial refugia for North Atlantic benthic marine taxa. Ecology 2008, 89, S108–S122. [Google Scholar] [CrossRef] [Green Version]
- Gardner, J.P.A.; Thompson, R.J. Influence of genotype and geography on shell shape and morphometric trait variation among North Atlantic blue mussel (Mytilus spp.) populations. Biol. J. Linn. Soc. 2009, 96, 875–897. [Google Scholar] [CrossRef]
- Aronson, R.B.; Thatje, S.; Clarke, A.; Peck, L.S.; Blake, D.B.; Wilga, C.D.; Seibel, B.A. Climate change and invasibility of the Antarctic benthos. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 129–154. [Google Scholar] [CrossRef] [Green Version]
- Harper, E.M.; Peck, L.S. Latitudinal and depth gradients in marine predation pressure. Glob. Ecol. Biogeogr. 2016, 25, 670–678. [Google Scholar] [CrossRef]
- Clarke, A. Temperature and extinction in the sea: A physiologist’s view. Paleobiology 1993, 19, 499–518. [Google Scholar] [CrossRef]
- Watson, S.-A.; Peck, L.S.; Tyler, P.A.; Southgate, P.C.; Tan, K.S.; Day, R.W.; Morley, S.A. Marine invertebrate skeleton size varies with latitude, temperature and carbonate saturation: Implications for global change and ocean acidification. Glob. Change Biol. 2012, 18, 3026–3038. [Google Scholar] [CrossRef]
- Zwarts, L.; Wanink, J. Siphon size and burying depth in deposit- and suspension-feeding benthic bivalves. Mar. Biol. 1989, 100, 227–240. [Google Scholar] [CrossRef]
- Zwarts, L.; Wanink, J.H. How oystercatchers and curlews successively deplete clams. In Coastal Waders and Wildfowl in Winter; Evans, P., Goss-Custard, J., Hale, W., Eds.; Cambridge University Press: Cambridge, UK, 1984; pp. 69–83. [Google Scholar]
- Watson, S.-A. Latitudinal Gradients in Marine Invertebrate Shell Morphology: Production Costs and Predation Pressure; University of Southampton: Southampton, UK, 2009. [Google Scholar]
- Harper, E.M.; Clark, M.S.; Hoffman, J.I.; Philipp, E.R.E.; Peck, L.S.; Morley, S.A. Iceberg scour and shell damage in the Antarctic bivalve Laternula elliptica. PLoS ONE 2012, 7, e46341. [Google Scholar] [CrossRef] [Green Version]
- Hinch, S.G.; Bailey, R.C.; Green, R.H. Growth of Lampsilis radiata (Bivalvia: Unionidae) in sand and mud: A reciprocal transplant experiment. Can. J. Fish. Aquat. Sci. 1986, 43, 548–552. [Google Scholar] [CrossRef]
- Moore, H.B. The relation of shell growth to environment in Patella vulgata. J. Molluscan Stud. 1934, 21, 217–222. [Google Scholar]
- Borrell, Y.J.; Pineda, H.; McCarthy, I.; Vazquez, E.; Sanchez, J.A.; Lizana, G.B. Correlations between fitness and heterozygosity at allozyme and microsatellite loci in the Atlantic salmon, Salmo salar L. Heredity 2004, 92, 585. [Google Scholar] [CrossRef] [Green Version]
- Yezerinac, S.M.; Lougheed, S.C.; Handford, P. Morphological variability and enzyme heterozygosity: Individual and population level correlations. Evolution 1992, 46, 1959–1964. [Google Scholar] [CrossRef] [PubMed]
- Mitton, J.B. Relationship between heterozygosity for enzyme loci and variation of morphological characters in natural populations. Nature 1978, 273, 661–662. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population ID | Location | Samples Used for Genetic Analysis | Samples Used for Shape Analysis | Samples Used for Thickness Analysis | Ho | He | Na | Ar |
|---|---|---|---|---|---|---|---|---|
| ITA | Comacchio (Italy) | 30 | 28 | 18 | 0.75 (0.13) | 0.76 (0.07) | 8.23 (2.37) | 6.24 (1.6) |
| LIS | Lisbon (Portugal) | 28 | 29 | 21 | 0.67 (0.19) | 0.74 (0.18) | 8.36 (2.24) | 6.56 (1.73) |
| BRE | Brest (France) | 32 | 35 | 19 | 0.76 (0.19) | 0.77 (0.12) | 9.54 (3.32) | 7.08 (2.56) |
| PLY | Plymouth (UK) | 27 | 30 | 19 | 0.66 (0.13) | 0.75 (0.09) | 8.63 (3.44) | 6.6 (2.24) |
| SAN | Saint Andrews (UK) | 23 | 22 | 20 | 0.74 (0.12) | 0.72 (0.1) | 5.91 (1.64) | 5.16 (1.45) |
| LCR | Le Crotoy (France) | 31 | 40 | 21 | 0.71 (0.14) | 0.75 (0.08) | 8.18 (2.44) | 6.36 (1.54) |
| TXL | Balgzand (Netherlands) | 30 | 30 | 21 | 0.71 (0.15) | 0.76 (0.1) | 8.72 (2.76) | 6.36 (1.65) |
| SYL | Sylt (Germany) | 12 | 13 | 10 | 0.62 (0.14) | 0.73 (0.13) | 6.36 (2.65) | 6.63 (2.65) |
| KIE | Kiel (Germany) | 34 | 35 | 18 | 0.59 (0.15) | 0.74 (0.11) | 8.91 (2.38) | 6.29 (1.52) |
| Total | 247 | 262 | 167 | 0.69 (0.06) | 0.75 (0.02) | 8.09 (1.19) | 6.36 (0.52) |
| Coefficient | Estimate | SE | 95% CI | t | p-Value |
|---|---|---|---|---|---|
| Shell shape GLMM † | |||||
| (Intercept) | 0.002 | 0.11 | −0.37; 0.37 | 0.01 | 0.99 |
| Shape (PC2) | −0.01 | 0.17 | −0.52; 0.49 | −0.07 | 0.94 |
| Shape (PC3) | 0.02 | 0.19 | −0.51; 0.53 | 0.12 | 0.91 |
| Latitude | −0.004 | 0.10 | −0.23; 0.22 | −0.03 | 0.97 |
| Length × Shape (PC1) | −0.11 | 0.08 | −0.35; 0.13 | −1.27 | 0.20 |
| Length × Shape (PC2) | −0.07 | 0.10 | −0.27; 0.13 | −0.70 | 0.49 |
| Length × Shape (PC3) | 0.27 | 0.10 | 0.07; 0.45 | 2.70 | 0.0072 |
| gPC1 | 0.03 | 0.06 | −0.09; 0.15 | 0.47 | 0.64 |
| gPC2 | −0.01 | 0.06 | −0.13; 0.12 | −0.15 | 0.88 |
| sMLH | 0.02 | 0.04 | −0.06; 0.11 | 0.55 | 0.58 |
| Shell layers thickness GLMM * | |||||
| (Intercept) | 274.60 | 7.15 | 257.38; 291.44 | 38.41 | <0.0001 |
| Layer (CL) | −181.29 | 7.30 | −204.95; −158.30 | −24.82 | <0.0001 |
| Layer (CCL) | −19.45 | 14.63 | −45.03; 5.82 | −1.33 | 0.18 |
| Latitude × Layer (PR) | 24.30 | 7.36 | 9.02; 39.27 | 3.30 | 0.0010 |
| Latitude × Layer (CL) | −2.68 | 3.40 | −17.10; 11.27 | −0.79 | 0.43 |
| Latitude × Layer (CCL) | −23.72 | 13.36 | −37.43; −9.69 | −1.78 | 0.076 |
| Length × Layer (PR) | 25.36 | 6.92 | 12.29; 38.93 | 3.66 | 0.0003 |
| Length × Layer (CL) | 6.74 | 2.67 | −7.63; 21.44 | 2.53 | 0.012 |
| Length × Layer (CCL) | 30.71 | 9.79 | 14.88; 46.79 | 3.14 | 0.0018 |
| gPC1 | −0.98 | 2.65 | −9.80; 7.82 | −0.37 | 0.71 |
| gPC2 | −1.62 | 2.45 | −10.62; 7.44 | −0.66 | 0.51 |
| sMLH | 0.82 | 2.14 | −6.80; 8.64 | 0.38 | 0.70 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Noia, M.; Telesca, L.; Vendrami, D.L.J.; Gokalp, H.K.; Charrier, G.; Harper, E.M.; Hoffman, J.I. Population Genetic Structure Is Unrelated to Shell Shape, Thickness and Organic Content in European Populations of the Soft-Shell Clam Mya Arenaria. Genes 2020, 11, 298. https://doi.org/10.3390/genes11030298
De Noia M, Telesca L, Vendrami DLJ, Gokalp HK, Charrier G, Harper EM, Hoffman JI. Population Genetic Structure Is Unrelated to Shell Shape, Thickness and Organic Content in European Populations of the Soft-Shell Clam Mya Arenaria. Genes. 2020; 11(3):298. https://doi.org/10.3390/genes11030298
Chicago/Turabian StyleDe Noia, Michele, Luca Telesca, David L. J. Vendrami, Hatice K. Gokalp, Grégory Charrier, Elizabeth M. Harper, and Joseph I. Hoffman. 2020. "Population Genetic Structure Is Unrelated to Shell Shape, Thickness and Organic Content in European Populations of the Soft-Shell Clam Mya Arenaria" Genes 11, no. 3: 298. https://doi.org/10.3390/genes11030298
APA StyleDe Noia, M., Telesca, L., Vendrami, D. L. J., Gokalp, H. K., Charrier, G., Harper, E. M., & Hoffman, J. I. (2020). Population Genetic Structure Is Unrelated to Shell Shape, Thickness and Organic Content in European Populations of the Soft-Shell Clam Mya Arenaria. Genes, 11(3), 298. https://doi.org/10.3390/genes11030298