The Oncogenic Potential of the Centromeric Border Protein FAM84B of the 8q24.21 Gene Desert


Abstract
:1. Introduction
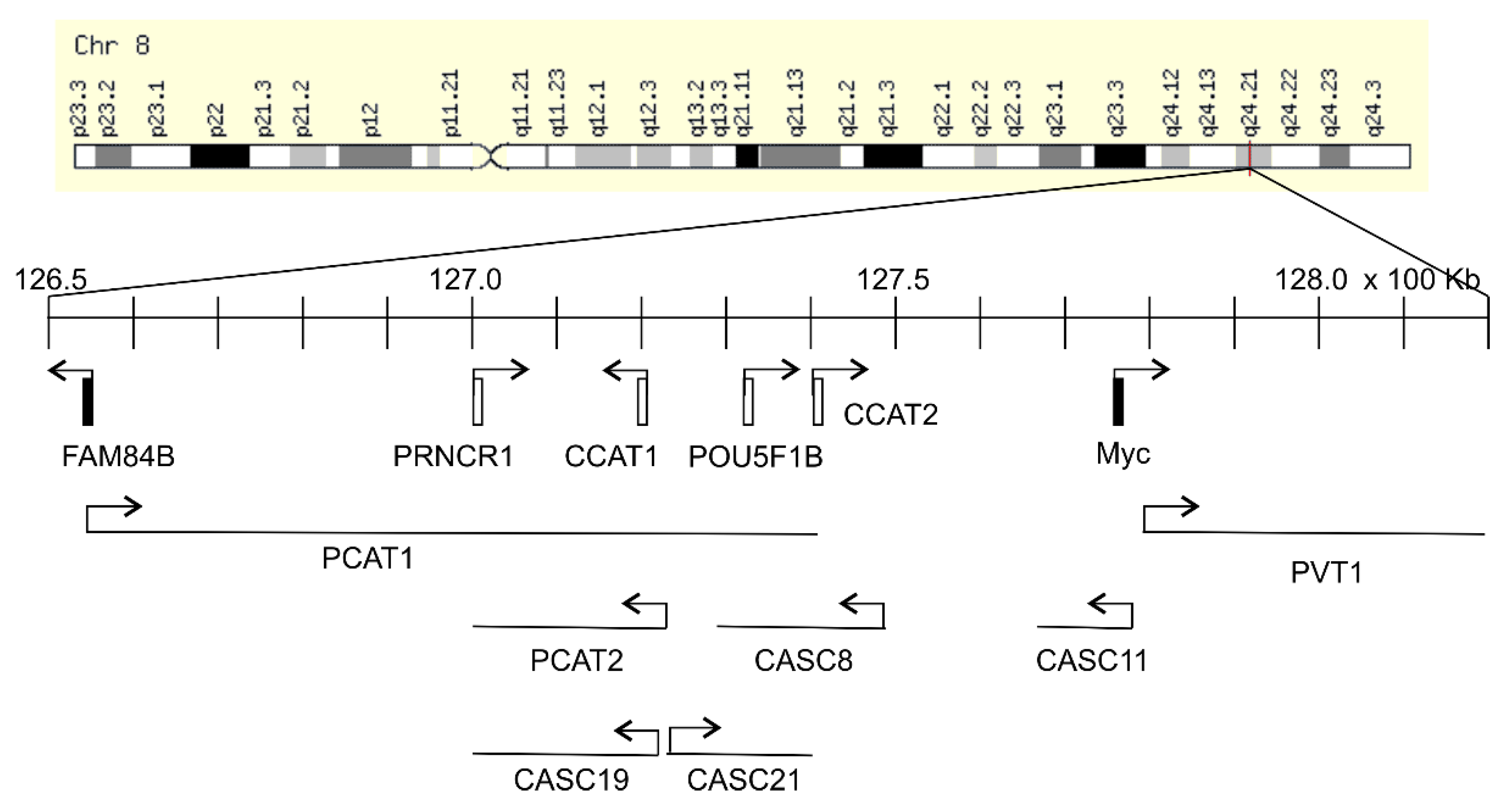
2. Function of the 8q24.21 Gene Desert in Cancers
2.1. Association of the 8q24.21 Gene Desert with Oncogenesis
2.2. Upregulation of Myc as A Mechanism underlying the Gene Desert-Derived Oncogenic Activities
3. The Contributions of FAM84B to Oncogenesis
3.1. FAM84B Facilitates Esophageal Cancer
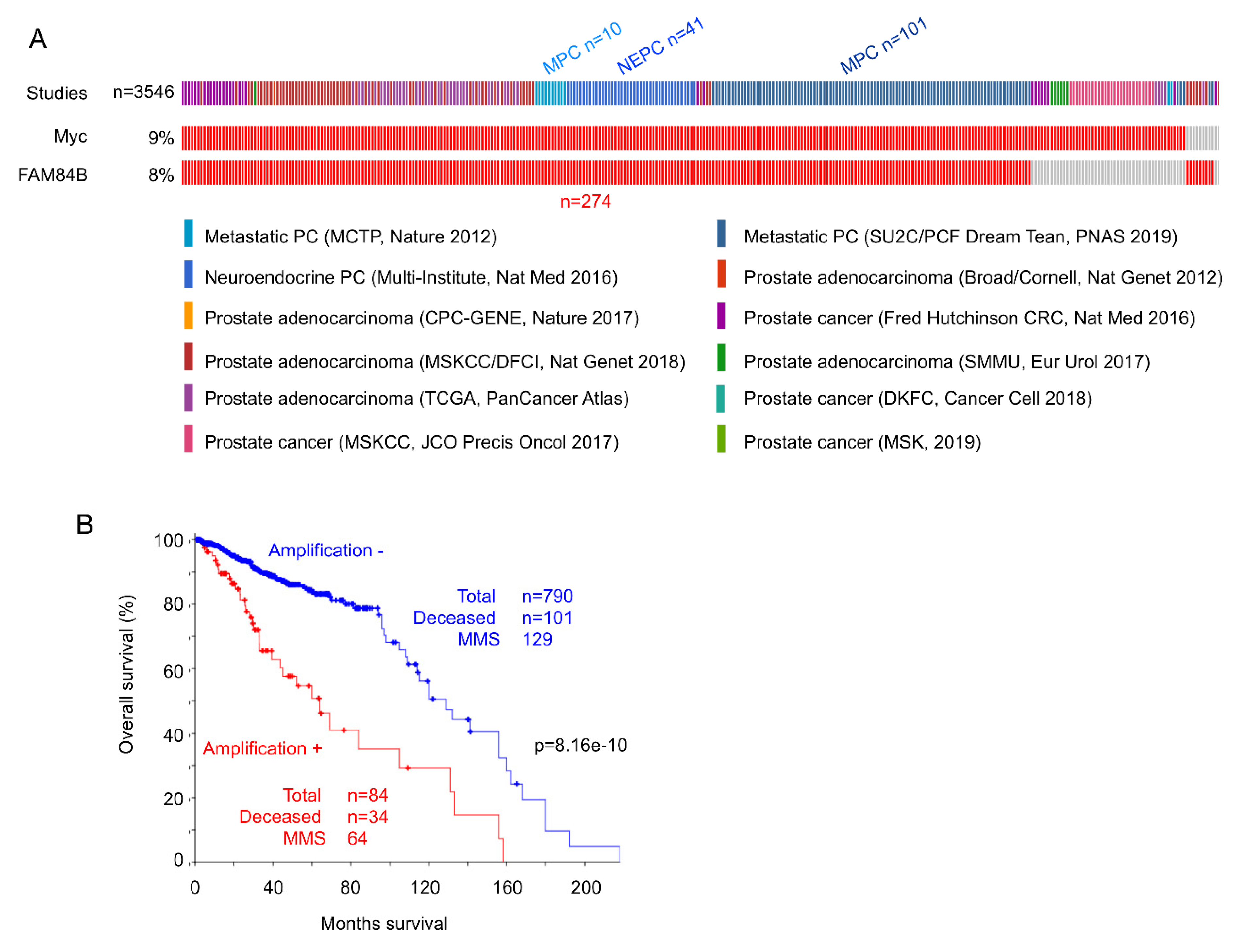
3.2. FAM84B-Mediated Enhancement of Prostate Cancer
4. Potential Collaboration between FAM84B and MYC during Tumorigenesis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kalkat, M.; De Melo, J.; Hickman, K.A.; Lourenco, C.; Redel, C.; Resetca, D.; Tamachi, A.; Tu, W.B.; Penn, L.Z. Myc deregulation in primary human cancers. Genes 2017, 8, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.M. The myc family of nuclear proto-oncogenes. Cancer Treat. Res. 1989, 47, 37–71. [Google Scholar] [PubMed]
- Beroukhim, R.; Mermel, C.H.; Porter, D.; Wei, G.; Raychaudhuri, S.; Donovan, J.; Barretina, J.; Boehm, J.S.; Dobson, J.; Urashima, M.; et al. The landscape of somatic copy-number alteration across human cancers. Nature 2010, 463, 899–905. [Google Scholar] [CrossRef] [PubMed]
- Haupt, Y.; Alexander, W.S.; Barri, G.; Klinken, S.P.; Adams, J.M. Novel zinc finger gene implicated as myc collaborator by retrovirally accelerated lymphomagenesis in E mu-myc transgenic mice. Cell 1991, 65, 753–763. [Google Scholar] [CrossRef]
- Van Lohuizen, M.; Verbeek, S.; Scheijen, B.; Wientjens, E.; Van der Gulden, H.; Berns, A. Identification of cooperating oncogenes in E mu-myc transgenic mice by provirus tagging. Cell 1991, 65, 737–752. [Google Scholar] [CrossRef]
- Huang, R.; Cheung, N.K.; Vider, J.; Cheung, I.Y.; Gerald, W.L.; Tickoo, S.K.; Holland, E.C.; Blasberg, R.G. MYCN and MYC regulate tumor proliferation and tumorigenesis directly through BMI1 in human neuroblastomas. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2011, 25, 4138–4149. [Google Scholar] [CrossRef] [Green Version]
- Waldron, T.; De Dominici, M.; Soliera, A.R.; Audia, A.; Iacobucci, I.; Lonetti, A.; Martinelli, G.; Zhang, Y.; Martinez, R.; Hyslop, T.; et al. c-Myb and its target Bmi1 are required for p190BCR/ABL leukemogenesis in mouse and human cells. Leukemia 2012, 26, 644–653. [Google Scholar] [CrossRef]
- Wang, H.B.; Liu, G.H.; Zhang, H.; Xing, S.; Hu, L.J.; Zhao, W.F.; Xie, B.; Li, M.Z.; Zeng, B.H.; Li, Y.; et al. Sp1 and c-Myc regulate transcription of BMI1 in nasopharyngeal carcinoma. FEBS J. 2013, 280, 2929–2944. [Google Scholar] [CrossRef]
- Land, H.; Parada, L.F.; Weinberg, R.A. Tumorigenic conversion of primary embryo fibroblasts requires at least two cooperating oncogenes. Nature 1983, 304, 596–602. [Google Scholar] [CrossRef]
- Leone, G.; DeGregori, J.; Sears, R.; Jakoi, L.; Nevins, J.R. Myc and Ras collaborate in inducing accumulation of active cyclin E/Cdk2 and E2F. Nature 1997, 387, 422–426. [Google Scholar] [CrossRef]
- Wang, C.; Lisanti, M.P.; Liao, D.J. Reviewing once more the c-myc and Ras collaboration: Converging at the cyclin D1-CDK4 complex and challenging basic concepts of cancer biology. Cell Cycle 2011, 10, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Wong, N.; Gu, Y.; Kapoor, A.; Lin, X.; Ojo, D.; Wei, F.; Yan, J.; De Melo, J.; Major, P.; Wood, G.; et al. Upregulation of FAM84B during prostate cancer progression. Oncotarget 2017, 8, 19218–19235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huppi, K.; Pitt, J.J.; Wahlberg, B.M.; Caplen, N.J. The 8q24 gene desert: An oasis of non-coding transcriptional activity. Front. Genet. 2012, 3, 69. [Google Scholar] [CrossRef] [Green Version]
- Sur, I.; Tuupanen, S.; Whitington, T.; Aaltonen, L.A.; Taipale, J. Lessons from functional analysis of genome-wide association studies. Cancer Res. 2013, 73, 4180–4184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derderian, C.; Orunmuyi, A.T.; Olapade-Olaopa, E.O.; Ogunwobi, O.O. PVT1 signaling is a mediator of cancer progression. Front. Oncol. 2019, 9, 502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, T.; Farina, L.; Macino, G.; Paci, P. PVT1: A rising star among oncogenic long noncoding RNAs. Biomed. Res. Int. 2015, 2015, 304208. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.F.; Yang, L.; Chen, L.L. The long noncoding RNA regulation at the MYC locus. Curr. Opin. Genet. Dev. 2015, 33, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Shamseer, L.; Moher, D.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A.; Group, P.-P. Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015: Elaboration and explanation. BMJ 2015, 350, g7647. [Google Scholar] [CrossRef] [Green Version]
- Moher, D.; Shamseer, L.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A.; Group, P.-P. Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015 statement. Syst. Rev. 2015, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Gudmundsson, J.; Sulem, P.; Gudbjartsson, D.F.; Blondal, T.; Gylfason, A.; Agnarsson, B.A.; Benediktsdottir, K.R.; Magnusdottir, D.N.; Orlygsdottir, G.; Jakobsdottir, M.; et al. Genome-wide association and replication studies identify four variants associated with prostate cancer susceptibility. Nat. Genet. 2009, 41, 1122–1126. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.C.; Hsing, A.W.; Edward, Y.; Biritwum, R.; Tettey, Y.; Adjei, A.; Cook, M.B.; De Marzo, A.; Netto, G.; Tay, E.; et al. A comprehensive resequence-analysis of 250 kb region of 8q24.21 in men of African ancestry. Prostate 2014, 74, 579–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bensen, J.T.; Xu, Z.; McKeigue, P.M.; Smith, G.J.; Fontham, E.T.; Mohler, J.L.; Taylor, J.A. Admixture mapping of prostate cancer in African Americans participating in the North Carolina-Louisiana Prostate Cancer Project (PCaP). Prostate 2014, 74, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasserman, N.F.; Aneas, I.; Nobrega, M.A. An 8q24 gene desert variant associated with prostate cancer risk confers differential in vivo activity to a MYC enhancer. Genome Res. 2010, 20, 1191–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grisanzio, C.; Freedman, M.L. Chromosome 8q24-associated cancers and MYC. Genes Cancer 2010, 1, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Ghoussaini, M.; Song, H.; Koessler, T.; Al Olama, A.A.; Kote-Jarai, Z.; Driver, K.E.; Pooley, K.A.; Ramus, S.J.; Kjaer, S.K.; Hogdall, E.; et al. Multiple loci with different cancer specificities within the 8q24 gene desert. J. Natl. Cancer Inst. 2008, 100, 962–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiltie, A.E. Common predisposition alleles for moderately common cancers: Bladder cancer. Curr. Opin. Genet. Dev. 2010, 20, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Braem, M.G.; Schouten, L.J.; Peeters, P.H.; Van den Brandt, P.A.; Onland-Moret, N.C. Genetic susceptibility to sporadic ovarian cancer: A systematic review. Biochim. Biophys. Acta 2011, 1816, 132–146. [Google Scholar] [CrossRef]
- Haerian, M.S.; Baum, L.; Haerian, B.S. Association of 8q24.21 loci with the risk of colorectal cancer: A systematic review and meta-analysis. J. Gastroenterol. Hepatol. 2011; 26, 1475–1484. [Google Scholar] [CrossRef]
- Haiman, C.A.; Le Marchand, L.; Yamamato, J.; Stram, D.O.; Sheng, X.; Kolonel, L.N.; Wu, A.H.; Reich, D.; Henderson, B.E. A common genetic risk factor for colorectal and prostate cancer. Nat. Genet. 2007, 39, 954–956. [Google Scholar] [CrossRef] [Green Version]
- Broderick, P.; Carvajal-Carmona, L.; Pittman, A.M.; Webb, E.; Howarth, K.; Rowan, A.; Lubbe, S.; Spain, S.; Sullivan, K.; Fielding, S.; et al. A genome-wide association study shows that common alleles of SMAD7 influence colorectal cancer risk. Nat. Genet. 2007, 39, 1315–1317. [Google Scholar] [CrossRef]
- Tomlinson, I.; Webb, E.; Carvajal-Carmona, L.; Broderick, P.; Kemp, Z.; Spain, S.; Penegar, S.; Chandler, I.; Gorman, M.; Wood, W.; et al. A genome-wide association scan of tag SNPs identifies a susceptibility variant for colorectal cancer at 8q24.21. Nat. Genet. 2007, 39, 984–988. [Google Scholar] [CrossRef]
- Hindorff, L.A.; Gillanders, E.M.; Manolio, T.A. Genetic architecture of cancer and other complex diseases: Lessons learned and future directions. Carcinogenesis 2011, 32, 945–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, A.C.; Lakhani, S.R.; Simpson, P.T. Pleomorphic lobular carcinoma of the breast: Molecular pathology and clinical impact. Future Oncol. 2009, 5, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Closas, M.; Chanock, S. Genetic susceptibility loci for breast cancer by estrogen receptor status. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2008, 14, 8000–8009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Easton, D.F.; Pooley, K.A.; Dunning, A.M.; Pharoah, P.D.; Thompson, D.; Ballinger, D.G.; Struewing, J.P.; Morrison, J.; Field, H.; Luben, R.; et al. Genome-wide association study identifies novel breast cancer susceptibility loci. Nature 2007, 447, 1087–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, F.R.; Feigelson, H.S.; Cox, D.G.; Haiman, C.A.; Albanes, D.; Buring, J.; Calle, E.E.; Chanock, S.J.; Colditz, G.A.; Diver, W.R.; et al. A common 8q24 variant in prostate and breast cancer from a large nested case-control study. Cancer Res. 2007, 67, 2951–2956. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, H.; Akamatsu, S.; Takata, R.; Takahashi, A.; Kubo, M.; Nakamura, Y. Prostate cancer genomics, biology, and risk assessment through genome-wide association studies. Cancer Sci. 2012, 103, 607–613. [Google Scholar] [CrossRef]
- Amundadottir, L.T.; Sulem, P.; Gudmundsson, J.; Helgason, A.; Baker, A.; Agnarsson, B.A.; Sigurdsson, A.; Benediktsdottir, K.R.; Cazier, J.B.; Sainz, J.; et al. A common variant associated with prostate cancer in European and African populations. Nat. Genet. 2006, 38, 652–658. [Google Scholar] [CrossRef]
- Freedman, M.L.; Haiman, C.A.; Patterson, N.; McDonald, G.J.; Tandon, A.; Waliszewska, A.; Penney, K.; Steen, R.G.; Ardlie, K.; John, E.M.; et al. Admixture mapping identifies 8q24 as a prostate cancer risk locus in African-American men. Proc. Natl. Acad. Sci. USA 2006, 103, 14068–14073. [Google Scholar] [CrossRef] [Green Version]
- Gudmundsson, J.; Sulem, P.; Manolescu, A.; Amundadottir, L.T.; Gudbjartsson, D.; Helgason, A.; Rafnar, T.; Bergthorsson, J.T.; Agnarsson, B.A.; Baker, A.; et al. Genome-wide association study identifies a second prostate cancer susceptibility variant at 8q24. Nat. Genet. 2007, 39, 631–637. [Google Scholar] [CrossRef]
- Haiman, C.A.; Patterson, N.; Freedman, M.L.; Myers, S.R.; Pike, M.C.; Waliszewska, A.; Neubauer, J.; Tandon, A.; Schirmer, C.; McDonald, G.J.; et al. Multiple regions within 8q24 independently affect risk for prostate cancer. Nat. Genet. 2007, 39, 638–644. [Google Scholar] [CrossRef] [Green Version]
- Yeager, M.; Orr, N.; Hayes, R.B.; Jacobs, K.B.; Kraft, P.; Wacholder, S.; Minichiello, M.J.; Fearnhead, P.; Yu, K.; Chatterjee, N.; et al. Genome-wide association study of prostate cancer identifies a second risk locus at 8q24. Nat. Genet. 2007, 39, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Van Duin, M.; Van Marion, R.; Vissers, K.; Watson, J.E.; Van Weerden, W.M.; Schroder, F.H.; Hop, W.C.; Van der Kwast, T.H.; Collins, C.; Van Dekken, H. High-resolution array comparative genomic hybridization of chromosome arm 8q: Evaluation of genetic progression markers for prostate cancer. Genes Chromosomes Cancer 2005, 44, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Ramnarine, V.R.; Kobelev, M.; Gibb, E.A.; Nouri, M.; Lin, D.; Wang, Y.; Buttyan, R.; Davicioni, E.; Zoubeidi, A.; Collins, C.C. The evolution of long noncoding RNA acceptance in prostate cancer initiation and progression, and its clinical utility in disease management. Eur. Urol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Urrutia, E.; Bustamante Montes, L.P.; Ladron de Guevara Cervantes, D.; Perez-Plasencia, C.; Campos-Parra, A.D. Crosstalk between long non-coding RNAs, micro-RNAs and mRNAs: Deciphering molecular mechanisms of master regulators in cancer. Front. Oncol. 2019, 9, 669. [Google Scholar] [CrossRef]
- Chung, S.; Nakagawa, H.; Uemura, M.; Piao, L.; Ashikawa, K.; Hosono, N.; Takata, R.; Akamatsu, S.; Kawaguchi, T.; Morizono, T.; et al. Association of a novel long non-coding RNA in 8q24 with prostate cancer susceptibility. Cancer Sci. 2011, 102, 245–252. [Google Scholar] [CrossRef]
- Kastler, S.; Honold, L.; Luedeke, M.; Kuefer, R.; Moller, P.; Hoegel, J.; Vogel, W.; Maier, C.; Assum, G. POU5F1P1, a putative cancer susceptibility gene, is overexpressed in prostatic carcinoma. Prostate 2010, 70, 666–674. [Google Scholar] [CrossRef]
- Hayashi, H.; Arao, T.; Togashi, Y.; Kato, H.; Fujita, Y.; De Velasco, M.A.; Kimura, H.; Matsumoto, K.; Tanaka, K.; Okamoto, I.; et al. The OCT4 pseudogene POU5F1B is amplified and promotes an aggressive phenotype in gastric cancer. Oncogene 2015, 34, 199–208. [Google Scholar] [CrossRef]
- Pan, Y.; Zhan, L.; Chen, L.; Zhang, H.; Sun, C.; Xing, C. POU5F1B promotes hepatocellular carcinoma proliferation by activating AKT. Biomed. Pharmacother. Biomed. Pharmacother. 2018, 100, 374–380. [Google Scholar] [CrossRef]
- Prensner, J.R.; Iyer, M.K.; Balbin, O.A.; Dhanasekaran, S.M.; Cao, Q.; Brenner, J.C.; Laxman, B.; Asangani, I.A.; Grasso, C.S.; Kominsky, H.D.; et al. Transcriptome sequencing across a prostate cancer cohort identifies PCAT-1, an unannotated lincRNA implicated in disease progression. Nat. Biotechnol. 2011, 29, 742–749. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Rand, K.A.; Hazelett, D.J.; Ingles, S.A.; Kittles, R.A.; Strom, S.S.; Rybicki, B.A.; Nemesure, B.; Isaacs, W.B.; Stanford, J.L.; et al. Prostate cancer susceptibility in men of African ancestry at 8q24. J. Natl. Cancer Inst. 2016, 108. [Google Scholar] [CrossRef]
- Bawa, P.; Zackaria, S.; Verma, M.; Gupta, S.; Srivatsan, R.; Chaudhary, B.; Srinivasan, S. Integrative analysis of normal long intergenic non-coding RNAs in prostate cancer. PLoS ONE 2015, 10, e0122143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.L.; Liu, H.M.; Han, N.; Li, F.H.; Sun, F.; Fan, D.M.; Xu, Q. PCAT1 promotes the proliferative and migratory potentials of ovarian cancer via targeting NEK2. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8239–8248. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Chen, Y.; Liao, X.; Liu, D.; Li, F.; Ruan, H.; Jia, W. Overexpression of long noncoding RNA PCAT-1 is a novel biomarker of poor prognosis in patients with colorectal cancer. Med. Oncol. 2013, 30, 588. [Google Scholar] [CrossRef] [PubMed]
- Kalmar, A.; Nagy, Z.B.; Galamb, O.; Csabai, I.; Bodor, A.; Wichmann, B.; Valcz, G.; Bartak, B.K.; Tulassay, Z.; Igaz, P.; et al. Genome-wide expression profiling in colorectal cancer focusing on lncRNAs in the adenoma-carcinoma transition. BMC Cancer 2019, 19, 1059. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, T.; Matsuyama, T.; Toiyama, Y.; Takahashi, N.; Ishikawa, T.; Uetake, H.; Yamada, Y.; Kusunoki, M.; Calin, G.; Goel, A. CCAT1 and CCAT2 long noncoding RNAs, located within the 8q.24.21 ‘gene desert’, serve as important prognostic biomarkers in colorectal cancer. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. ESMO 2017, 28, 1882–1888. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, C.; Chang, Y.; Zhang, Z.; Hu, Y.; Zhang, F.; Lu, Y.; Zheng, L.; Zhang, W.; Li, X.; et al. Long non-coding RNA CASC11 interacts with hnRNP-K and activates the WNT/beta-catenin pathway to promote growth and metastasis in colorectal cancer. Cancer Lett. 2016, 376, 62–73. [Google Scholar] [CrossRef]
- Zhang, L.; Kang, W.; Lu, X.; Ma, S.; Dong, L.; Zou, B. LncRNA CASC11 promoted gastric cancer cell proliferation, migration and invasion in vitro by regulating cell cycle pathway. Cell Cycle 2018, 17, 1886–1900. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.G.; Wang, C.H.; He, R.Q.; Xu, R.Y.; Ji, C.B. LncRNA CASC11 promotes the development of esophageal carcinoma by regulating KLF6. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8878–8887. [Google Scholar] [CrossRef]
- Zheng, Y.; Nie, P.; Xu, S. Long noncoding RNA CASC21 exerts an oncogenic role in colorectal cancer through regulating miR-7-5p/YAP1 axis. Biomed. Pharmacother. Biomed. Pharmacother. 2020, 121, 109628. [Google Scholar] [CrossRef]
- Durst, M.; Gissmann, L.; Ikenberg, H.; zur Hausen, H. A papillomavirus DNA from a cervical carcinoma and its prevalence in cancer biopsy samples from different geographic regions. Proc. Natl. Acad. Sci. USA 1983, 80, 3812–3815. [Google Scholar] [CrossRef] [Green Version]
- Clifford, G.M.; Smith, J.S.; Plummer, M.; Munoz, N.; Franceschi, S. Human papillomavirus types in invasive cervical cancer worldwide: A meta-analysis. Br. J. Cancer 2003, 88, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Shen, Y.T.; Kapoor, A.; Ojo, D.; Wei, F.; De Melo, J.; Lin, X.; Wong, N.; Yan, J.; Tao, L.; et al. CYB5D2 displays tumor suppression activities towards cervical cancer. Biochim. Biophys. Acta 2016, 1862, 556–565. [Google Scholar] [CrossRef] [PubMed]
- Peter, M.; Rosty, C.; Couturier, J.; Radvanyi, F.; Teshima, H.; Sastre-Garau, X. MYC activation associated with the integration of HPV DNA at the MYC locus in genital tumors. Oncogene 2006, 25, 5985–5993. [Google Scholar] [CrossRef] [Green Version]
- Kraus, I.; Driesch, C.; Vinokurova, S.; Hovig, E.; Schneider, A.; Von Knebel Doeberitz, M.; Durst, M. The majority of viral-cellular fusion transcripts in cervical carcinomas cotranscribe cellular sequences of known or predicted genes. Cancer Res. 2008, 68, 2514–2522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferber, M.J.; Thorland, E.C.; Brink, A.A.; Rapp, A.K.; Phillips, L.A.; McGovern, R.; Gostout, B.S.; Cheung, T.H.; Chung, T.K.; Fu, W.Y.; et al. Preferential integration of human papillomavirus type 18 near the c-myc locus in cervical carcinoma. Oncogene 2003, 22, 7233–7242. [Google Scholar] [CrossRef] [Green Version]
- Diao, M.K.; Liu, C.Y.; Liu, H.W.; Li, J.T.; Li, F.; Mehryar, M.M.; Wang, Y.J.; Zhan, S.B.; Zhou, Y.B.; Zhong, R.G.; et al. Integrated HPV genomes tend to integrate in gene desert areas in the CaSki, HeLa, and SiHa cervical cancer cell lines. Life Sci. 2015, 127, 46–52. [Google Scholar] [CrossRef]
- Li, W.; Qi, Y.; Cui, X.; Huo, Q.; Zhu, L.; Zhang, A.; Tan, M.; Hong, Q.; Yang, Y.; Zhang, H.; et al. Characteristic of HPV integration in the genome and transcriptome of cervical cancer tissues. Biomed. Res. Int. 2018, 2018, 6242173. [Google Scholar] [CrossRef]
- Hu, Z.; Zhu, D.; Wang, W.; Li, W.; Jia, W.; Zeng, X.; Ding, W.; Yu, L.; Wang, X.; Wang, L.; et al. Genome-wide profiling of HPV integration in cervical cancer identifies clustered genomic hot spots and a potential microhomology-mediated integration mechanism. Nat. Genet. 2015, 47, 158–163. [Google Scholar] [CrossRef]
- Ling, H.; Spizzo, R.; Atlasi, Y.; Nicoloso, M.; Shimizu, M.; Redis, R.S.; Nishida, N.; Gafa, R.; Song, J.; Guo, Z.; et al. CCAT2, a novel noncoding RNA mapping to 8q24, underlies metastatic progression and chromosomal instability in colon cancer. Genome Res. 2013, 23, 1446–1461. [Google Scholar] [CrossRef] [Green Version]
- Ahmadiyeh, N.; Pomerantz, M.M.; Grisanzio, C.; Herman, P.; Jia, L.; Almendro, V.; He, H.H.; Brown, M.; Liu, X.S.; Davis, M.; et al. 8q24 prostate, breast, and colon cancer risk loci show tissue-specific long-range interaction with MYC. Proc. Natl. Acad. Sci. USA 2010, 107, 9742–9746. [Google Scholar] [CrossRef] [Green Version]
- Du, M.; Yuan, T.; Schilter, K.F.; Dittmar, R.L.; Mackinnon, A.; Huang, X.; Tschannen, M.; Worthey, E.; Jacob, H.; Xia, S.; et al. Prostate cancer risk locus at 8q24 as a regulatory hub by physical interactions with multiple genomic loci across the genome. Hum. Mol. Genet. 2015, 24, 154–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotelo, J.; Esposito, D.; Duhagon, M.A.; Banfield, K.; Mehalko, J.; Liao, H.; Stephens, R.M.; Harris, T.J.; Munroe, D.J.; Wu, X. Long-range enhancers on 8q24 regulate c-Myc. Proc. Natl. Acad. Sci. USA 2010, 107, 3001–3005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sur, I.K.; Hallikas, O.; Vaharautio, A.; Yan, J.; Turunen, M.; Enge, M.; Taipale, M.; Karhu, A.; Aaltonen, L.A.; Taipale, J. Mice lacking a Myc enhancer that includes human SNP rs6983267 are resistant to intestinal tumors. Science 2012, 338, 1360–1363. [Google Scholar] [CrossRef] [PubMed]
- Dave, K.; Sur, I.; Yan, J.; Zhang, J.; Kaasinen, E.; Zhong, F.; Blaas, L.; Li, X.; Kharazi, S.; Gustafsson, C.; et al. Mice deficient of Myc super-enhancer region reveal differential control mechanism between normal and pathological growth. eLife 2017, 6. [Google Scholar] [CrossRef]
- Xiang, J.F.; Yin, Q.F.; Chen, T.; Zhang, Y.; Zhang, X.O.; Wu, Z.; Zhang, S.; Wang, H.B.; Ge, J.; Lu, X.; et al. Human colorectal cancer-specific CCAT1-L lncRNA regulates long-range chromatin interactions at the MYC locus. Cell Res. 2014, 24, 513–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.; Cui, R.; Jeon, Y.J.; Lee, J.H.; Lee, J.H.; Sim, H.; Park, J.K.; Fadda, P.; Tili, E.; Nakanishi, H.; et al. Long-range interaction and correlation between MYC enhancer and oncogenic long noncoding RNA CARLo-5. Proc. Natl. Acad. Sci. USA 2014, 111, 4173–4178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, K.B.; Maia, A.T.; O’Reilly, M.; Ghoussaini, M.; Prathalingam, R.; Porter-Gill, P.; Ambs, S.; Prokunina-Olsson, L.; Carroll, J.; Ponder, B.A. A functional variant at a prostate cancer predisposition locus at 8q24 is associated with PVT1 expression. PLoS Genet. 2011, 7, e1002165. [Google Scholar] [CrossRef]
- Tseng, Y.Y.; Moriarity, B.S.; Gong, W.; Akiyama, R.; Tiwari, A.; Kawakami, H.; Ronning, P.; Reuland, B.; Guenther, K.; Beadnell, T.C.; et al. PVT1 dependence in cancer with MYC copy-number increase. Nature 2014, 512, 82–86. [Google Scholar] [CrossRef]
- Zhang, Y.; Tan, Y.; Wang, H.; Xu, M.; Xu, L. Long non-coding RNA plasmacytoma variant translocation 1 (PVT1) enhances proliferation, migration, and epithelial-mesenchymal transition (EMT) of pituitary adenoma cells by activating beta-catenin, c-Myc, and cyclin D1 expression. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 7652–7659. [Google Scholar] [CrossRef]
- Huang, X.P.; Rong, T.H.; Wang, J.Y.; Tang, Y.Q.; Li, B.J.; Xu, D.R.; Zhao, M.Q.; Zhang, L.J.; Fang, Y.; Su, X.D.; et al. Negative implication of C-MYC as an amplification target in esophageal cancer. Cancer Genet. Cytogenet. 2006, 165, 20–24. [Google Scholar] [CrossRef]
- Cheng, C.; Cui, H.; Zhang, L.; Jia, Z.; Song, B.; Wang, F.; Li, Y.; Liu, J.; Kong, P.; Shi, R.; et al. Genomic analyses reveal FAM84B and the NOTCH pathway are associated with the progression of esophageal squamous cell carcinoma. GigaScience 2016, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, F.M.; Cheng, J.C.; Chang, Y.L.; Lee, J.M.; Koong, A.C.; Chuang, E.Y. Circulating mRNA profiling in esophageal squamous cell carcinoma identifies FAM84B as a biomarker in predicting pathological response to neoadjuvant chemoradiation. Sci. Rep. 2015, 5, 10291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parisi, F.; Ariyan, S.; Narayan, D.; Bacchiocchi, A.; Hoyt, K.; Cheng, E.; Xu, F.; Li, P.; Halaban, R.; Kluger, Y. Detecting copy number status and uncovering subclonal markers in heterogeneous tumor biopsies. BMC Genom. 2011, 12, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Duin, M.; Van Marion, R.; Vissers, K.J.; Hop, W.C.; Dinjens, W.N.; Tilanus, H.W.; Siersema, P.D.; Van Dekken, H. High-resolution array comparative genomic hybridization of chromosome 8q: Evaluation of putative progression markers for gastroesophageal junction adenocarcinomas. Cytogenet. Genome Res. 2007, 118, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Q.; Yu, S.; Zhu, C.; Zhang, Z.; Cao, H.; Xu, J. Long non-coding RNA FAM84B-AS promotes resistance of gastric cancer to platinum drugs through inhibition of FAM84B expression. Biochem. Biophys. Res. Commun. 2019, 509, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Camp, N.J.; Farnham, J.M.; Allen-Brady, K.; Cannon-Albright, L.A. Statistical recombinant mapping in extended high-risk Utah pedigrees narrows the 8q24 prostate cancer locus to 2.0 Mb. Prostate 2007, 67, 1456–1464. [Google Scholar] [CrossRef]
- Ting, M.C.; Liao, C.P.; Yan, C.; Jia, L.; Groshen, S.; Frenkel, B.; Roy-Burman, P.; Coetzee, G.A.; Maxson, R. An enhancer from the 8q24 prostate cancer risk region is sufficient to direct reporter gene expression to a subset of prostate stem-like epithelial cells in transgenic mice. Dis. Models Mech. 2012, 5, 366–374. [Google Scholar] [CrossRef] [Green Version]
- Mei, W.; Lin, X.; Kapoor, A.; Gu, Y.; Zhao, K.; Tang, D. The contributions of prostate cancer stem cells in prostate cancer initiation and metastasis. Cancers 2019, 11, 434. [Google Scholar] [CrossRef] [Green Version]
- Bagnall, P. Diagnosis and treatment of prostate cancer. Nurs. Times 2014, 110, 12–15. [Google Scholar]
- Heinlein, C.A.; Chang, C. Androgen receptor in prostate cancer. Endocr. Rev. 2004, 25, 276–308. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Gu, Y.; Kapoor, A.; Wei, F.; Aziz, T.; Ojo, D.; Jiang, Y.; Bonert, M.; Shayegan, B.; Yang, H.; et al. Overexpression of MUC1 and genomic alterations in its network associate with prostate cancer progression. Neoplasia 2017, 19, 857–867. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Lin, X.; Kapoor, A.; He, L.; Wei, F.; Gu, Y.; Mei, W.; Zhao, K.; Yang, H.; Tang, D. FAM84B promotes prostate tumorigenesis through a network alteration. Ther. Adv. Med. Oncol. 2019, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mardian, E.B.; Bradley, R.M.; Duncan, R.E. The HRASLS (PLA/AT) subfamily of enzymes. J. Biomed. Sci. 2015, 22, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, X.; Lin, J.; Jin, C.; Xia, B. Solution structure of the N-terminal catalytic domain of human H-REV107—A novel circular permutated NlpC/P60 domain. FEBS Lett. 2010, 584, 4222–4226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akiyama, H.; Hiraki, Y.; Noda, M.; Shigeno, C.; Ito, H.; Nakamura, T. Molecular cloning and biological activity of a novel Ha-Ras suppressor gene predominantly expressed in skeletal muscle, heart, brain, and bone marrow by differential display using clonal mouse EC cells, ATDC5. J. Biol. Chem. 1999, 274, 32192–32197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shyu, R.Y.; Hsieh, Y.C.; Tsai, F.M.; Wu, C.C.; Jiang, S.Y. Cloning and functional characterization of the HRASLS2 gene. Amino Acids 2008, 35, 129–137. [Google Scholar] [CrossRef]
- Hajnal, A.; Klemenz, R.; Schafer, R. Subtraction cloning of H-rev107, a gene specifically expressed in H-ras resistant fibroblasts. Oncogene 1994, 9, 479–490. [Google Scholar]
- Wang, C.H.; Shyu, R.Y.; Wu, C.C.; Tsai, T.C.; Wang, L.K.; Chen, M.L.; Jiang, S.Y.; Tsai, F.M. Phospholipase A/Acyltransferase enzyme activity of H-rev107 inhibits the H-RAS signaling pathway. J. Biomed. Sci. 2014, 21, 36. [Google Scholar] [CrossRef] [Green Version]
- Han, C.W.; Jeong, M.S.; Jang, S.B. Molecular interaction between K-Ras and H-REV107 in the Ras signaling pathway. Biochem. Biophys. Res. Commun. 2017, 491, 257–264. [Google Scholar] [CrossRef]
- Tsai, F.M.; Shyu, R.Y.; Jiang, S.Y. RIG1 suppresses Ras activation and induces cellular apoptosis at the Golgi apparatus. Cell. Signal. 2007, 19, 989–999. [Google Scholar] [CrossRef]
- Morales, M.; Arenas, E.J.; Urosevic, J.; Guiu, M.; Fernandez, E.; Planet, E.; Fenwick, R.B.; Fernandez-Ruiz, S.; Salvatella, X.; Reverter, D.; et al. RARRES3 suppresses breast cancer lung metastasis by regulating adhesion and differentiation. EMBO Mol. Med. 2014, 6, 865–881. [Google Scholar] [CrossRef] [PubMed]
- Homer-Bouthiette, C.; Zhao, Y.; Shunkwiler, L.B.; Van Peel, B.; Garrett-Mayer, E.; Baird, R.C.; Rissman, A.I.; Guest, S.T.; Ethier, S.P.; John, M.C.; et al. Deletion of the murine ortholog of the 8q24 gene desert has anti-cancer effects in transgenic mammary cancer models. BMC Cancer 2018, 18, 1233. [Google Scholar] [CrossRef] [PubMed]
- Kamath, A.; Tara, H.; Xiang, B.; Bajaj, R.; He, W.; Li, P. Double-minute MYC amplification and deletion of MTAP, CDKN2A, CDKN2B, and ELAVL2 in an acute myeloid leukemia characterized by oligonucleotide-array comparative genomic hybridization. Cancer Genet. Cytogenet. 2008, 183, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.; Bothma, H.; Veale, R.; Willem, P. Genomic imbalances in esophageal carcinoma cell lines involve Wnt pathway genes. World J. Gastroenterol. 2011, 17, 2909–2923. [Google Scholar] [CrossRef]
- Camps, J.; Nguyen, Q.T.; Padilla-Nash, H.M.; Knutsen, T.; McNeil, N.E.; Wangsa, D.; Hummon, A.B.; Grade, M.; Ried, T.; Difilippantonio, M.J. Integrative genomics reveals mechanisms of copy number alterations responsible for transcriptional deregulation in colorectal cancer. Genes Chromosomes Cancer 2009, 48, 1002–1017. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Carroll, P.A.; Freie, B.W.; Mathsyaraja, H.; Eisenman, R.N. The MYC transcription factor network: Balancing metabolism, proliferation and oncogenesis. Front. Med. 2018, 12, 412–425. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| lnRNA | PC | GC | ESC | HCC | OVC | CRC | Ref |
|---|---|---|---|---|---|---|---|
| PRNCR1 | Exp + Cell prolif + AR sig + | NA | NA | NA | NA | [46] | |
| POU5F1B | Exp + | Prom | NA | Prom | NA | NA | [47,48,49] |
| PCAT1 | Exp + | NA | NA | NA | Cell prolif + | Poor OS | [50,51,52,53,54] |
| PCAT2 | Exp + | NA | NA | NA | NA | NA | [50,51,52] |
| CCAT1 | NA | NA | NA | NA | NA | Exp + Poor OS | [55,56] |
| CCAT2 | NA | NA | NA | NA | NA | Exp + Poor OS | [55,56] |
| CASC11 | NA | Cell prolif + | Prom | Met + | [59] | ||
| CASC19 | NA | NA | NA | NA | NA | Exp + | [55,56] |
| CASC21 | NA | NA | NA | NA | NA | Prom | [60] |
| Member | Function | Refs |
|---|---|---|
| HRASLS1 | Inhibition of NIH3 Ras cell proliferation | [96] |
| HRASLS2 | Reduction of Ras-GTP level Reduction of HCT116 and HeLa cell colony number Downregulation of Ras expression in HtTA cervical cancer cells | [97] |
| HRASLS3 | Inhibition of Ras ability to transform rat fibroblasts Inhibition of Ras signalling Inhibition of K-Ras via binding to K-Ras | [98] [99] [100] |
| HRASLS4 | Suppression of Ras activation Inhibition of breast cancer metastasis to the lung | [101] [102] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, Y.; Lin, X.; Kapoor, A.; Chow, M.J.; Jiang, Y.; Zhao, K.; Tang, D. The Oncogenic Potential of the Centromeric Border Protein FAM84B of the 8q24.21 Gene Desert. Genes 2020, 11, 312. https://doi.org/10.3390/genes11030312
Gu Y, Lin X, Kapoor A, Chow MJ, Jiang Y, Zhao K, Tang D. The Oncogenic Potential of the Centromeric Border Protein FAM84B of the 8q24.21 Gene Desert. Genes. 2020; 11(3):312. https://doi.org/10.3390/genes11030312
Chicago/Turabian StyleGu, Yan, Xiaozeng Lin, Anil Kapoor, Mathilda Jing Chow, Yanzhi Jiang, Kuncheng Zhao, and Damu Tang. 2020. "The Oncogenic Potential of the Centromeric Border Protein FAM84B of the 8q24.21 Gene Desert" Genes 11, no. 3: 312. https://doi.org/10.3390/genes11030312
APA StyleGu, Y., Lin, X., Kapoor, A., Chow, M. J., Jiang, Y., Zhao, K., & Tang, D. (2020). The Oncogenic Potential of the Centromeric Border Protein FAM84B of the 8q24.21 Gene Desert. Genes, 11(3), 312. https://doi.org/10.3390/genes11030312