Controlling Apomixis: Shared Features and Distinct Characteristics of Gene Regulation


Abstract
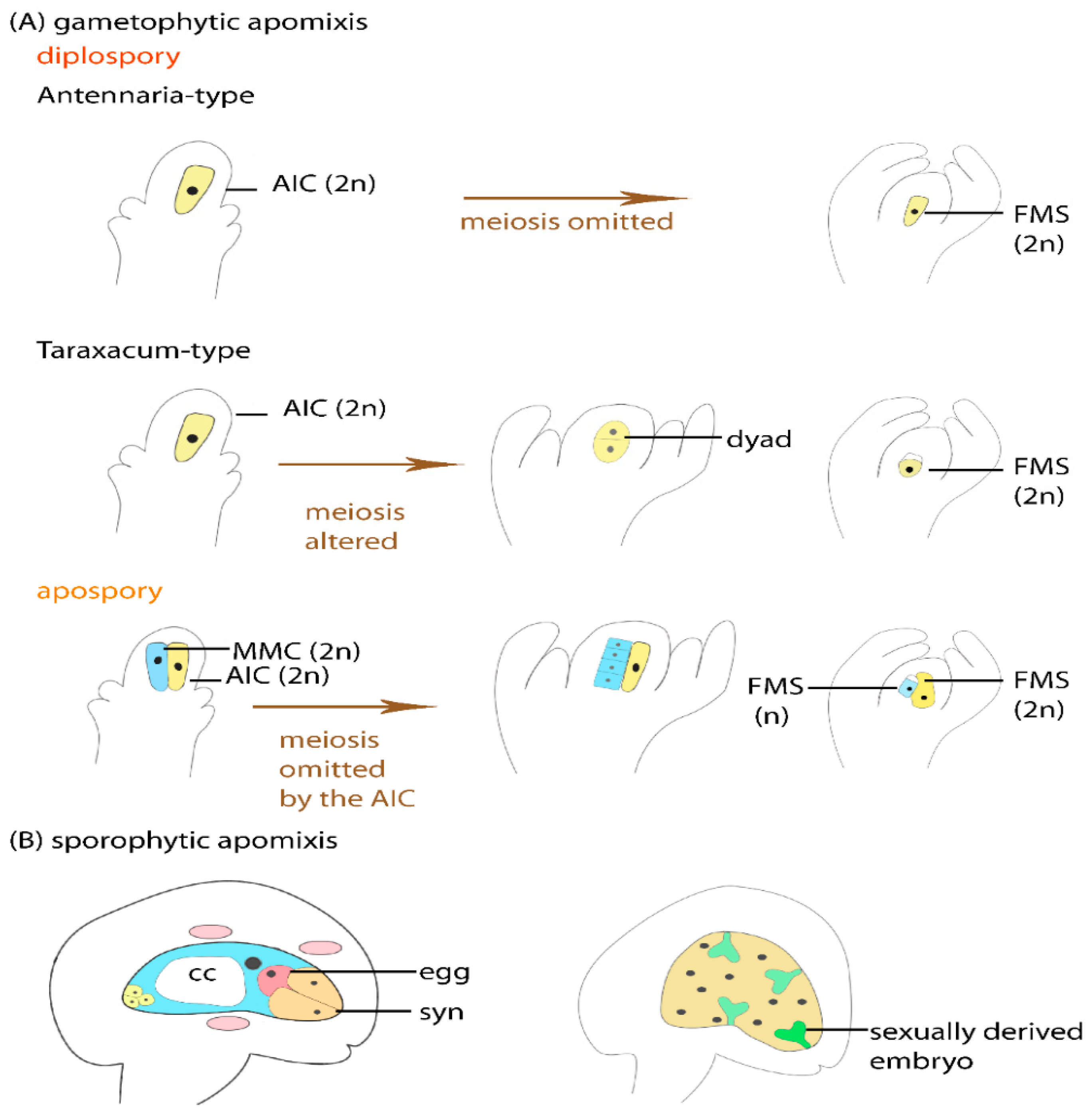
:1. Plant Reproduction Is Characterized by Developmental Flexibility Including Sexual and Asexual Formation of Seeds (Apomixis)
2. The Developmental Flexibility of Plant Reproduction might Hold Evolutionary Advantages
3. Genetic Loci Linked to Apomixis Typically Represent Hemizygous Heterochromatic Regions
Genes Located on Apomixis Loci Suggest That Different Regulatory Pathways Are Involved in Controlling Apomixis
4. Transcriptional Analysis Identifies Genes Differentially Regulated during Sexual and Apomictic Reproduction
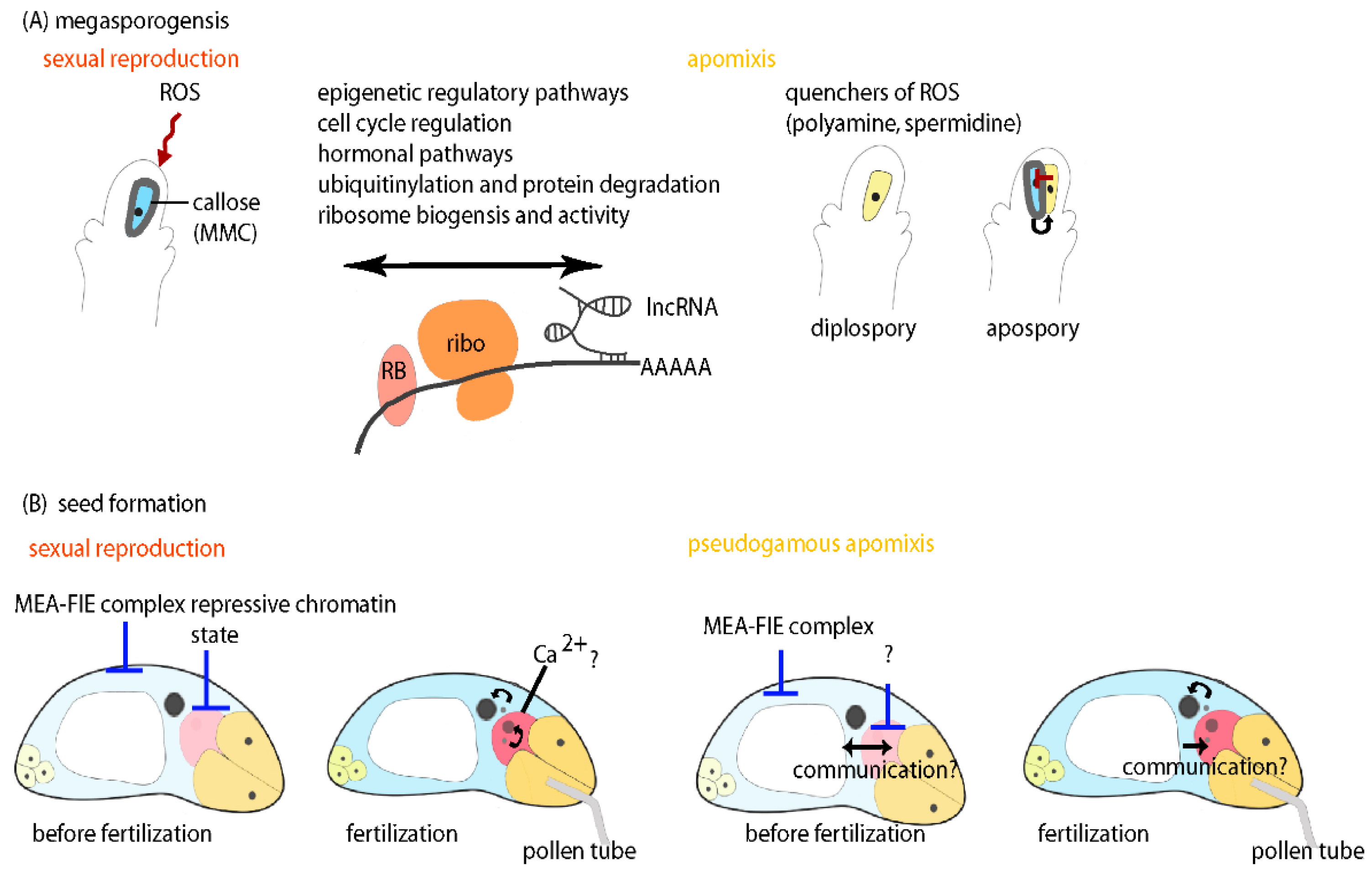
5. Different Layers of Regulation Are in Place to Control Development during Sexual and Apomictic Megasporogenesis
5.1. Specialized Ribosomes and Associated Factors Emerge as Novel Players in Gene Regulation
5.2. Epigenetic Regulatory Pathways Are Involved in Regulation of Germline Development
5.3. Cell-Cycle Control and Regulation of Meiosis is Differentially Regulated during Meiosis and Apomeiosis
5.4. Signal Transduction, Cell-Cell Communication and Hormonal Pathways Appear to Be Involved in Regulation of Apomixis
5.5. Regulation of Seed Development in Apomicts Might Require both Repression and Activation of Gene Activity
6. Are Stress Signal and Nutritional State Triggering the Determination for Sexual Reproduction or Apomixis?
7. Brief Summary and Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Hörandl, E.; Hojsgaard, D. The evolution of apomixis in angiosperms: A reappraisal. Plant Biosyst 2012, 146, 681–693. [Google Scholar] [CrossRef]
- Whitton, J.; Sears, C.; Baack, E.; Otto, S. The Dynamic Nature of Apomixis in the Angiosperms. Int. J. Plant Sci. 2008, 169, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Hojsgaard, D.; Klatt, S.; Baier, R.; Carman, J.G.; Hörandl, E. Taxonomy and Biogeography of Apomixis in Angiosperms and Associated Biodiversity Characteristics. CRC Crit. Rev. Plant Sci. 2014, 33, 414–427. [Google Scholar] [CrossRef] [Green Version]
- Carman, J.G. Asynchronous expression of duplicate genes in angiosperms may cause apomixis, bispory, tetraspory, and polyembryony. Biol. J. Linn. Soc. 1997, 61, 51–94. [Google Scholar] [CrossRef]
- Wang, X.; Xu, Y.; Zhang, S.; Cao, L.; Huang, Y.; Cheng, J. Genomic analyses of primitive, wild and cultivated citrus provide insights into asexual reproduction. Nat. Genet. 2017, 49, 765–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hojsgaard, D.; Horandl, E. The Rise of Apomixis in Natural Plant Populations. Front. Plant Sci. 2019, 10, 358. [Google Scholar] [CrossRef] [PubMed]
- Fei, X.; Shi, J.; Liu, Y.; Jinshuang, N.; Wei, A. The steps from sexual reproduction to apomixis. Planta 2019, 249, 1715–1730. [Google Scholar] [CrossRef] [Green Version]
- Spillane, C.; Steimer, A.; Grossniklaus, U. Apomixis in agriculture: The quest for clonal seeds. Sex. Plant Reprod. 2001, 14, 179–187. [Google Scholar] [CrossRef]
- Koltunow, A.M.; Bicknell, R.A.; Chaudhury, A.M. Apomixis: Molecular Strategies for the Generation of Genetically Identical Seeds without Fertilization. Plant Physiol. 1995, 108, 1345–1352. [Google Scholar] [CrossRef] [Green Version]
- Barcaccia, G.; Albertini, E. Apomixis in plant reproduction: A novel perspective on an old dilemma. Plant Reprod. 2013, 26, 159–179. [Google Scholar] [CrossRef] [Green Version]
- Hisanaga, T.; Yamaoka, S.; Kawashima, T.; Higo, A.; Nakajima, K.; Araki, T.; Kohchi, T.; Berger, F. Building new insights in plant gametogenesis from an evolutionary perspective. Nat. Plants 2019, 5, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, K. Be my baby: Patterning toward plant germ cells. Curr. Opin. Plant Biol. 2018, 41, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Erbasol Serbes, I.; Palovaara, J.; Gross-Hardt, R. Development and function of the flowering plant female gametophyte. Curr. Top. Dev. Biol. 2019, 131, 401–434. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.W.; Schmidt, A.; Grossniklaus, U. The female gametophyte: An emerging model for cell type-specific systems biology in plant development. Front. Plant Sci. 2015, 6, 907. [Google Scholar] [CrossRef] [Green Version]
- Koltunow, A.M.; Grossniklaus, U. Apomixis: A developmental perspective. Annu. Rev. Plant Biol. 2003, 54, 547–574. [Google Scholar] [CrossRef] [Green Version]
- Nakano, M.; Shimada, T.; Endo, T.; Fujii, H.; Nesumi, H.; Kita, M.; Ebinac, M.; Shimizua, T.; Omurabd, M. Characterization of genomic sequence showing strong association with polyembryony among diverse Citrus species and cultivars, and its synteny with Vitis and Populus. Plant Sci. Int. J. Exp. Plant Biol. 2012, 183, 131–142. [Google Scholar] [CrossRef]
- Bicknell, R.A.; Koltunow, A.M. Understanding apomixis: Recent advances and remaining conundrums. Plant Cell 2004, 16 (Suppl. 1), S228–S245. [Google Scholar] [CrossRef] [Green Version]
- Albertini, E.; Barcaccia, G.; Carman, J.G.; Pupilli, F. Did apomixis evolve from sex or was it the other way around? J. Exp. Bot. 2019, 70, 2951–2964. [Google Scholar] [CrossRef]
- Nogler, G.A. Gametophytic Apomixis. In Embryology of Angiosperms; Johri, B.M., Ed.; Springer: Berlin/Heidelberg, Germany, 1984; pp. 475–518. [Google Scholar]
- Leblanc, O.; Mazzucato, A. Screening procedures to identify and quantify apomixis. In The Flowering of Apomixis: From Mechanisms to Genetic Engineering; Savidan, Y., Carman, J.G., Dresselhaus, T., Eds.; CIMMYT: Texcoco, Mexico, 2001. [Google Scholar]
- Van Dijk, P.J.; Bakx-Schotman, J.M.T. Formation of Unreduced Megaspores (Diplospory) in Apomictic Dandelions (Taraxacum officinale, s.l.) Is Controlled by a Sex-Specific Dominant Locus. Genetics 2004, 166, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Koltunow, A.M.; Johnson, S.D.; Rodrigues, J.C.; Okada, T.; Hu, Y.; Tsuchiya, T.; Wilson, S.; Fletcher, P.; Ito, K.; Suzuku, G. Sexual reproduction is the default mode in apomictic Hieracium subgenus Pilosella, in which two dominant loci function to enable apomixis. Plant J. 2011, 66, 890–902. [Google Scholar] [CrossRef] [Green Version]
- Mandáková, T.; Ashby, K.; Price, B.; Windham, M.; Carman, J.; Lysak, M. Genome structure and apomixis in Phoenicaulis (Brassicaceae; Boechereae). J. Syst. Evol. 2019. [Google Scholar] [CrossRef]
- Van Dijk, P.; Vijverberg, K. The significance of apomixis in the evolution of the angiosperms: A reappraisal. In Plant Species-Level Systematics. New Perspectives on Pattern and Process; Regnum Vegetabile 143; Bakker, F.T., Chatrou, L.W., Gravendeel, B., Pelzer, P.B., Eds.; Koeltz Scientific Books: Ruggell, Liechtenstein, 2005; pp. 101–116. [Google Scholar]
- Muller, H.J. The Relation of Recombination to Mutational Advance. Mutat. Res. 1964, 106, 2–9. [Google Scholar] [CrossRef]
- R Marshall, D.; Brown, A. The evolution of apomixis. Heredity 1981, 47. [Google Scholar] [CrossRef] [Green Version]
- Brukhin, V.; Osadtchiy, J.V.; Florez-Rueda, A.M.; Smetanin, D.; Bakin, E.; Nobre, M.S.; Bakin, E.; Grossniklaus, U. The Boechera Genus as a Resource for Apomixis Research. Front. Plant Sci. 2019, 10, 392. [Google Scholar] [CrossRef] [Green Version]
- Lovell, J.T.; Aliyu, O.M.; Mau, M.; Schranz, M.E.; Koch, M.; Kiefer, C.; Song, B.-H.; Mitchell-Olds, T.; Sharbel, T.F. On the origin and evolution of apomixis in Boechera. Plant Reprod. 2013, 26, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Podio, M.; Cáceres, M.E.; Samoluk, S.S.; Seijo, J.G.; Pessino, S.C.; Ortiz, J.P.A.; Pupilli, F. A methylation status analysis of the apomixis-specific region in Paspalum spp. suggests an epigenetic control of parthenogenesis. J. Exp. Bot. 2014, 65, 6411–6424. [Google Scholar] [CrossRef] [Green Version]
- Hojsgaard, D.H.; Martinez, E.J.; Quarin, C.L. Competition between meiotic and apomictic pathways during ovule and seed development results in clonality. New Phytol. 2013, 197, 336–347. [Google Scholar] [CrossRef]
- Hojsgaard, D.; Hörandl, E. A little bit of sex matters for genome evolution in asexual plants. Front. Plant Sci. 2015, 6, 82. [Google Scholar] [CrossRef] [Green Version]
- Hodač, L.; Klatt, S.; Hojsgaard, D.; Sharbel, T.F.; Hörandl, E. A little bit of sex prevents mutation accumulation even in apomictic polyploid plants. BMC Evol. Biol. 2019, 19, 170. [Google Scholar] [CrossRef] [Green Version]
- Carman, J.G.; Mateo de Arias, M.; Gao, L.; Zhao, X.; Kowallis, B.M.; Sherwood, D.A.; Srivastava, M.K.; Dwivedi, K.K.; Price, B.J.; Watts, L. Apospory and Diplospory in Diploid Boechera (Brassicaceae) May Facilitate Speciation by Recombination-Driven Apomixis-to-Sex Reversals. Front. Plant Sci. 2019, 10, 724. [Google Scholar] [CrossRef]
- Juranic, M.; Johnson, S.D.; Koltunow, A.M. Phenotypic plasticity of aposporous embryo sac development in Hieracium praealtum. Plant Signal. Behav. 2019, 14, 1622981. [Google Scholar] [CrossRef]
- Hand, M.L.; Koltunow, A.M.G. The Genetic Control of Apomixis: Asexual Seed Formation. Genetics 2014, 197, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Ozias-Akins, P.; Roche, D.; Hanna, W.W. Tight clustering and hemizygosity of apomixis-linked molecular markers in Pennisetum squamulatum implies genetic control of apospory by a divergent locus that may have no allelic form in sexual genotypes. Proc. Natl. Acad. Sci. USA 1998, 95, 5127–5132. [Google Scholar] [CrossRef] [Green Version]
- Grimanelli, D.; Leblanc, O.; Espinosa, E.; Perotti, E.; Gonzalez de Leon, D.; Savidan, Y. Mapping diplosporous apomixis in tetraploid Tripsacum: One gene or several genes? Heredity 1998, 80, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Grossniklaus, U.; Nogler, G.A.; van Dijk, P.J. How to Avoid Sex. Genet. Control Gametophytic Apomixis 2001, 13, 1491–1498. [Google Scholar] [CrossRef] [Green Version]
- Grimanelli, D.; Leblanc, O.; Perotti, E.; Grossniklaus, U. Developmental genetics of gametophytic apomixis. Trends Genet. TIG 2001, 17, 597–604. [Google Scholar] [CrossRef]
- Ozias-Akins, P.; Akiyama, Y.; Hanna, W.W. Molecular characterization of the genomic region linked with apomixis in Pennisetum/Cenchrus. Funct. Integr. Genom. 2003, 3, 94–104. [Google Scholar] [CrossRef]
- Zappacosta, D.; Gallardo, J.; Carballo, J.; Meier, M.; Rodrigo, J.M.; Gallo, C.A.; Selva, J.P.; Stein, J.; Ortiz, J.P.A.; Albertini, E. A High-Density Linkage Map of the Forage Grass Eragrostis curvula and Localization of the Diplospory Locus. Front. Plant Sci. 2019, 10, 918. [Google Scholar] [CrossRef] [Green Version]
- Ozias-Akins, P.; Dijk, P. Mendelian Genetics of Apomixis in Plants. Annu. Rev. Genet. 2007, 41, 509–537. [Google Scholar] [CrossRef]
- Pupilli, F.; Barcaccia, G. Cloning plants by seeds: Inheritance models and candidate genes to increase fundamental knowledge for engineering apomixis in sexual crops. J. Biotechnol. 2012, 159, 291–311. [Google Scholar] [CrossRef]
- Schallau, A.; Arzenton, F.; Johnston, A.J.; Hahnel, U.; Koszegi, D.; Blattner, F.R.; Altschmied, L.; Haberer, G.; Barcaccia, G.; Bäumlein, H. Identification and genetic analysis of the APOSPORY locus in Hypericum perforatum L. Plant J. 2010, 62, 773–784. [Google Scholar] [CrossRef]
- Noyes, R.D.; Rieseberg, L.H. Two Independent Loci Control Agamospermy (Apomixis) in the Triploid Flowering Plant Erigeron annuus. Genetics 2000, 155, 379–390. [Google Scholar]
- Vasut, R.J.; Vijverberg, K.; van Dijk, P.J.; de Jong, H. Fluorescent in situ hybridization shows DIPLOSPOROUS located on one of the NOR chromosomes in apomictic dandelions (Taraxacum) in the absence of a large hemizygous chromosomal region. Genome 2014, 57, 609–620. [Google Scholar] [CrossRef]
- Vijverberg, K.; Milanovic-Ivanovic, S.; Bakx-Schotman, T.; van Dijk, P.J. Genetic fine-mapping of DIPLOSPOROUS in Taraxacum (dandelion; Asteraceae) indicates a duplicated DIP-gene. BMC Plant Biol. 2010, 10, 154. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, Y.; Hanna, W.W.; Ozias-Akins, P. High-resolution physical mapping reveals that the apospory-specific genomic region (ASGR) in Cenchrus ciliaris is located on a heterochromatic and hemizygous region of a single chromosome. TAG Theor. Appl. Genet. Theor. Angew. Genet. 2005, 111, 1042–1051. [Google Scholar] [CrossRef]
- Conner, J.A.; Goel, S.; Gunawan, G.; Cordonnier-Pratt, M.M.; Johnson, V.E.; Liang, C.; Wang, H.; Pratt, L.H.; Mullet, J.E.; DeBarry, J. Sequence analysis of bacterial artificial chromosome clones from the apospory-specific genomic region of Pennisetum and Cenchrus. Plant Physiol. 2008, 147, 1396–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goel, S.; Chen, Z.; Akiyama, Y.; Conner, J.A.; Basu, M.; Gualtieri, G.; Hanna, W.W.; Ozias-Akins, P. Comparative physical mapping of the apospory-specific genomic region in two apomictic grasses: Pennisetum squamulatum and Cenchrus ciliaris. Genetics 2006, 173, 389–400. [Google Scholar] [CrossRef] [Green Version]
- Galla, G.; Siena, L.A.; Ortiz, J.P.A.; Baumlein, H.; Barcaccia, G.; Pessino, S.C.; Belluci, M.; Pupilli, F. A Portion of the Apomixis Locus of Paspalum Simplex is Microsyntenic with an Unstable Chromosome Segment Highly Conserved Among Poaceae. Sci. Rep. 2019, 9, 3271. [Google Scholar] [CrossRef] [Green Version]
- Catanach, A.S.; Erasmuson, S.K.; Podivinsky, E.; Jordan, B.R.; Bicknell, R. Deletion mapping of genetic regions associated with apomixis in Hieracium. Proc. Natl. Acad. Sci. USA 2006, 103, 18650–18655. [Google Scholar] [CrossRef] [Green Version]
- Kotani, Y.; Henderson, S.; Suzuki, G.; Johnson, S.; Okada, T.; Siddons, H.; Mukai, Y.; Koltunow, A.M. The LOSS OF APOMEIOSIS (LOA) locus in Hieracium praealtum can function independently of the associated large-scale repetitive chromosomal structure. New Phytol. 2014, 201, 973–981. [Google Scholar] [CrossRef]
- Okada, T.; Ito, K.; Johnson, S.D.; Oelkers, K.; Suzuki, G.; Houben, A.; Mukai, Y.; Koltunow, A.M. Chromosomes carrying meiotic avoidance loci in three apomictic eudicot Hieracium subgenus Pilosella species share structural features with two monocot apomicts. Plant Physiol. 2011, 157, 1327–1341. [Google Scholar] [CrossRef] [Green Version]
- Sharbel, T.F.; Voigt, M.L.; Mitchell-Olds, T.; Kantama, L.; de Jong, H. Is the aneuploid chromosome in an apomictic Boechera holboellii a genuine B chromosome? Cytogenet. Genome Res. 2004, 106, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Sharbel, T.F.; Mitchell-Olds, T.; Dobes, C.; Kantama, L.; de Jong, H. Biogeographic distribution of polyploidy and B chromosomes in the apomictic Boechera holboellii complex. Cytogenet. Genome Res. 2005, 109, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Mandakova, T.; Schranz, M.E.; Sharbel, T.F.; de Jong, H.; Lysak, M.A. Karyotype evolution in apomictic Boechera and the origin of the aberrant chromosomes. Plant J. 2015, 82, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Kantama, L.; Sharbel, T.F.; Schranz, M.E.; Mitchell-Olds, T.; de Vries, S.; de Jong, H. Diploid apomicts of the Boechera holboellii complex display large-scale chromosome substitutions and aberrant chromosomes. Proc. Natl. Acad. Sci. USA 2007, 104, 14026–14031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schranz, M.E.; Kantama, L.; de Jong, H.; Mitchell-Olds, T. Asexual reproduction in a close relative of Arabidopsis: A genetic investigation of apomixis in Boechera (Brassicaceae). New Phytol. 2006, 171, 425–438. [Google Scholar] [CrossRef] [PubMed]
- Dechaud, C.; Volff, J.-N.; Schartl, M.; Naville, M. Sex and the TEs: Transposable elements in sexual development and function in animals. Mob. DNA 2019, 10, 42. [Google Scholar] [CrossRef]
- Taylor, D.; Chu, E.; Spektor, R.; Soloway, P. Long Non-Coding RNA Regulation of Reproduction and Development. Mol. Reprod. Dev. 2015, 82, 932–956. [Google Scholar] [CrossRef] [Green Version]
- Galla, G.; Basso, A.; Grisan, S.; Bellucci, M.; Pupilli, F.; Barcaccia, G. Ovule Gene Expression Analysis in Sexual and Aposporous Apomictic Hypericum perforatum L. (Hypericaceae) Accessions. Front. Plant Sci. 2019, 10, 654. [Google Scholar] [CrossRef] [Green Version]
- Corral, J.M.; Vogel, H.; Aliyu, O.M.; Hensel, G.; Thiel, T.; Kumlehn, J.; Sharbel, T.F. A Conserved Apomixis-Specific Polymorphism Is Correlated with Exclusive Exonuclease Expression in Premeiotic Ovules of Apomictic Boechera Species. Plant Physiol. 2013, 163, 1660–1672. [Google Scholar] [CrossRef] [Green Version]
- Mau, M.; Lovell, J.T.; Corral, J.M.; Kiefer, C.; Koch, M.A.; Aliyu, O.M.; Sharbel, T.F. Hybrid apomicts trapped in the ecological niches of their sexual ancestors. Proc. Natl. Acad. Sci. USA 2015, 112, E2357–E2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mau, M.; Corral, J.M.; Vogel, H.; Melzer, M.; Fuchs, J.; Kuhlmann, M.; de Storme, N.; Geelen, D.; Sharbel, T.F. The conserved chimeric transcript UPGRADE2 is associated with unreduced pollen formation and is exclusively found in apomictic Boechera species. Plant Physiol. 2013, 163, 1640–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, M.A. A new chromosome was born: Comparative chromosome painting in Boechera. Trends Plant Sci. 2015, 20, 533–535. [Google Scholar] [CrossRef] [PubMed]
- Mancini, M.; Permingeat, H.; Colono, C.; Siena, L.; Pupilli, F.; Azzaro, C.; de Alencar Dusi, D.M.; de Campos Carneiro, V.T.; Podio, M.; Seijo, J.G. The MAP3K-Coding QUI-GON JINN (QGJ) Gene Is Essential to the Formation of Unreduced Embryo Sacs in Paspalum. Front. Plant Sci. 2018, 9, 1547. [Google Scholar] [CrossRef] [Green Version]
- Siena, L.A.; Ortiz, J.P.; Calderini, O.; Paolocci, F.; Caceres, M.E.; Kaushal, P.; Grisan, S.; Pessino, S.C.; Pupilli, F. An apomixis-linked ORC3-like pseudogene is associated with silencing of its functional homolog in apomictic Paspalum simplex. J. Exp. Bot. 2016, 67, 1965–1978. [Google Scholar] [CrossRef] [Green Version]
- Conner, J.A.; Gunawan, G.; Ozias-Akins, P. Recombination within the apospory specific genomic region leads to the uncoupling of apomixis components in Cenchrus ciliaris. Planta 2013, 238, 51–63. [Google Scholar] [CrossRef]
- Conner, J.A.; Ozias-Akins, P. Apomixis: Engineering the Ability to Harness Hybrid Vigor in Crop Plants. Methods Mol. Biol. 2017, 1669, 17–34. [Google Scholar] [CrossRef]
- Horstman, A.; Li, M.; Heidmann, I.; Weemen, M.; Chen, B.; Muino, J.M.; Angenent, G.C.; Boutilier, K. The BABY BOOM Transcription Factor Activates the LEC1-ABI3-FUS3-LEC2 Network to Induce Somatic Embryogenesis. Plant Physiol. 2017, 175, 848–857. [Google Scholar] [CrossRef] [Green Version]
- Boutilier, K.; Offringa, R.; Sharma, V.K.; Kieft, H.; Ouellet, T.; Zhang, L.; Hattori, J.; Liu, C.-M.; van Lammeren, A.A.M. Ectopic Expression of BABY BOOM Triggers a Conversion from Vegetative to Embryonic Growth. Plant Cell 2002, 14, 1737–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conner, J.A.; Mookkan, M.; Huo, H.; Chae, K.; Ozias-Akins, P. A parthenogenesis gene of apomict origin elicits embryo formation from unfertilized eggs in a sexual plant. Proc. Natl. Acad. Sci. USA 2015, 112, 11205–11210. [Google Scholar] [CrossRef] [Green Version]
- Worthington, M.; Heffelfinger, C.; Bernal, D.; Quintero, C.; Zapata, Y.P.; Perez, J.G.; De Vega, J.; Miles, J.; Dellaporta, S.; Tohme, J. A Parthenogenesis Gene Candidate and Evidence for Segmental Allopolyploidy in Apomictic Brachiaria decumbens. Genetics 2016, 203, 1117–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanday, I.; Skinner, D.; Yang, B.; Mercier, R.; Sundaresan, V. A male-expressed rice embryogenic trigger redirected for asexual propagation through seeds. Nature 2019, 565, 91–95. [Google Scholar] [CrossRef]
- Shimada, T.; Endo, T.; Fujii, H.; Nakano, M.; Sugiyama, A.; Daido, G.; Ohta, S.; Yoshioka, T.; Omura, M. MITE insertion-dependent expression of CitRKD1 with a RWP-RK domain regulates somatic embryogenesis in citrus nucellar tissues. BMC Plant Biol. 2018, 18, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tedeschi, F.; Rizzo, P.; Rutten, T.; Altschmied, L.; Bäumlein, H. RWP-RK domain-containing transcription factors control cell differentiation during female gametophyte development in Arabidopsis. New Phytol. 2017, 213, 1909–1924. [Google Scholar] [CrossRef] [PubMed]
- Koszegi, D.; Johnston, A.J.; Rutten, T.; Czihal, A.; Altschmied, L.; Kumlehn, J.; Wüst, S.E.; Kiroukhova, O.; Gheyselinck, J.; Grossniklaus, U. Members of the RKD transcription factor family induce an egg cell-like gene expression program. Plant J. 2011, 67, 280–291. [Google Scholar] [CrossRef] [Green Version]
- Wuest, S.E.; Vijverberg, K.; Schmidt, A.; Weiss, M.; Gheyselinck, J.; Lohr, M.; Wellmer, F.; Rahnenführer, J.; von Mering, C.; Grossniklaus, U. Arabidopsis Female Gametophyte Gene Expression Map Reveals Similarities between Plant and Animal Gametes. Curr. Biol. 2010, 20, 506–512. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.; Schmid, M.W.; Klostermeier, U.C.; Qi, W.; Guthörl, D.; Sailer, C.; Waller, M.; Rosenstiel, P.; Grossniklaus, U. Apomictic and Sexual Germline Development Differ with Respect to Cell Cycle, Transcriptional, Hormonal and Epigenetic Regulation. PLoS Genet. 2014, 10, e1004476. [Google Scholar] [CrossRef] [Green Version]
- Koi, S.; Hisanaga, T.; Sato, K.; Shimamura, M.; Yamato Katsuyuki, T.; Ishizaki, K.; Kohchi, T.; Nakajama, K. An Evolutionarily Conserved Plant RKD Factor Controls Germ Cell Differentiation. Curr. Biol. 2016, 26, 1775–1781. [Google Scholar] [CrossRef] [Green Version]
- Rovekamp, M.; Bowman, J.L.; Grossniklaus, U. Marchantia MpRKD Regulates the Gametophyte-Sporophyte Transition by Keeping Egg Cells Quiescent in the Absence of Fertilization. Curr. Biol. 2016, 26, 1782–1789. [Google Scholar] [CrossRef] [Green Version]
- Vielle-Calzada, J.P.; Nuccio, M.L.; Budiman, M.A.; Thomas, T.L.; Burson, B.L.; Hussey, M.A.; Wing, R.A. Comparative gene expression in sexual and apomictic ovaries of Pennisetum ciliare (L.) Link. Plant Mol. Biol. 1996, 32, 1085–1092. [Google Scholar] [CrossRef]
- Sahu, P.P.; Gupta, S.; Malaviya, D.R.; Roy, A.K.; Kaushal, P.; Prasad, M. Transcriptome Analysis of Differentially Expressed Genes During Embryo Sac Development in Apomeiotic Non-Parthenogenetic Interspecific Hybrid of Pennisetum glaucum. Mol. Biotechnol. 2012, 51, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Miyazaki, C.; Kojimai, A.; Saito, A.; Adachi, T. Isolation and Characterization of a Gene Expressed during Early Embryo Sac Development in Apomictic Guinea Grass (Panicum maximum). J. Plant Physiol. 1999, 154, 55–62. [Google Scholar] [CrossRef]
- Radhakrishna, A.; Dwivedi, K.K.; Srivastava, M.K.; Roy, A.K.; Malaviya, D.R.; Kaushal, P. Transcriptomic data of pre-meiotic stage of floret development in apomictic and sexual types of guinea grass (Panicum maximum Jacq.). Data Brief 2018, 18, 590–593. [Google Scholar] [CrossRef]
- Albertini, E.; Marconi, G.; Barcaccia, G.; Raggi, L.; Falcinelli, M. Isolation of candidate genes for apomixis in Poa pratensis L. Plant Mol. Biol. 2004, 56, 879–894. [Google Scholar] [CrossRef] [PubMed]
- Albertini, E.; Marconi, G.; Reale, L.; Barcaccia, G.; Porceddu, A.; Ferranti, F.; Falcinelli, M. SERK and APOSTART. Candidate Genes for Apomixis in Poa pratensis. Plant Physiol. 2005, 138, 2185–2199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leblanc, O.; Armstead, I.; Pessino, S.; Ortiz, J.P.A.; Evans, C.; do Valle, C.; Hayward, M.D. Non-radioactive mRNA fingerprinting to visualise gene expression in mature ovaries of Brachiaria hybrids derived from B. brizantha, an apomictic tropical forage. Plant Sci. 1997, 126, 49–58. [Google Scholar] [CrossRef]
- Rodrigues, J.C.M.; Cabral, G.B.; Dusi, D.M.A.; de Mello, L.V.; Rigden, D.J.; Carneiro, V.T.C. Identification of differentially expressed cDNA sequences in ovaries of sexual and apomictic plants of Brachiaria brizantha. Plant Mol. Biol. 2003, 53, 745–757. [Google Scholar] [CrossRef]
- Pessino, S.C.; Espinoza, F.; Martínez, E.J.; Ortiz, J.P.A.; Valle, E.M.; Quarín, C.L. Isolation of cDNA Clones Differentially Expressed in Flowers of Apomictic and Sexual Paspalum Notatum. Hereditas 2001, 134, 35–42. [Google Scholar] [CrossRef]
- Polegri, L.; Calderini, O.; Arcioni, S.; Pupilli, F. Specific expression of apomixis-linked alleles revealed by comparative transcriptomic analysis of sexual and apomictic Paspalum simplex Morong flowers. J. Exp. Bot. 2010, 61, 1869–1883. [Google Scholar] [CrossRef] [Green Version]
- Ochogavía, A.C.; Seijo, J.G.; González, A.M.; Podio, M.; Duarte Silveira, E.; Machado Lacerda, A.L.; de Campos Carneiro, V.T.; Ortiz, J.P.A.; Pessino, S.C. Characterization of retrotransposon sequences expressed in inflorescences of apomictic and sexual Paspalum notatum plants. Sex. Plant Reprod. 2011, 24, 231–246. [Google Scholar] [CrossRef]
- Ortiz, J.P.A.; Revale, S.; Siena, L.A.; Podio, M.; Delgado, L.; Stein, J.; Leblanc, O.; Pessino, S.C. A reference floral transcriptome of sexual and apomictic Paspalum notatum. BMC Genom. 2017, 18, 318. [Google Scholar] [CrossRef] [Green Version]
- Garbus, I.; Romero, J.R.; Selva, J.P.; Pasten, M.C.; Chinestra, C.; Carballo, J.; Zappacosta, D.C.; Echenique, V. De novo transcriptome sequencing and assembly from apomictic and sexual Eragrostis curvula genotypes. PLoS ONE 2017, 12, e0185595. [Google Scholar] [CrossRef] [PubMed]
- Barcaccia, G.; Varotto, S.; Meneghetti, S.; Albertini, E.; Porceddu, A.; Parrini, P.; Lucchin, M. Analysis of gene expression during flowering in apomeiotic mutants of Medicago spp.: Cloning of ESTs and candidate genes for 2n eggs. Sex. Plant Reprod. 2001, 14, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Zang, G.; Cheng, C.; Luan, M.; Dai, Z.; Xu, Y.; Yang, Z.; Zhao, L.; Su, J. Diplosporous development in Boehmeria tricuspis: Insights from de novo transcriptome assembly and comprehensive expression profiling. Sci. Rep. 2017, 7, 46043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galla, G.; Zenoni, S.; Avesani, L.; Altschmied, L.; Rizzo, P.; Sharbel, T.F.; Barcaccia, G. Pistil Transcriptome Analysis to Disclose Genes and Gene Products Related to Aposporous Apomixis in Hypericum perforatum L. Front. Plant Sci. 2017, 8, 79. [Google Scholar] [CrossRef] [Green Version]
- Tucker, M.R.; Araujo, A.-C.G.; Paech, N.A.; Hecht, V.; Schmidt, E.D.L.; Rossell, J.-B.; de Vries, S.C.; Koltunow, A.M.G. Sexual and Apomictic Reproduction in Hieracium subgenus Pilosella Are Closely Interrelated Developmental Pathways. Plant Cell 2003, 15, 1524–1537. [Google Scholar] [CrossRef] [Green Version]
- Brauning, S.; Catanach, A.; Lord, J.M.; Bicknell, R.; Macknight, R.C. Comparative transcriptome analysis of the wild-type model apomict Hieracium praealtum and its loss of parthenogenesis (lop) mutant. BMC Plant Biol. 2018, 18, 206. [Google Scholar] [CrossRef] [Green Version]
- Sharbel, T.F.; Voigt, M.-L.; Corral, J.M.; Galla, G.; Kumlehn, J.; Klukas, C.; Schreiber, F.; Vogel, H.; Rotter, B. Apomictic and Sexual Ovules of Boechera Display Heterochronic Global Gene Expression Patterns. Plant Cell 2010, 22, 655–671. [Google Scholar] [CrossRef] [Green Version]
- Sharbel, T.F.; Voigt, M.-L.; Corral, J.M.; Thiel, T.; Varshney, A.; Kumlehn, J.; Vogel, H.; Rotter, B. Molecular signatures of apomictic and sexual ovules in the Boechera holboellii complex. Plant J. 2009, 58, 870–882. [Google Scholar] [CrossRef]
- Zühl, L.; Volkert, C.; Ibberson, D.; Schmidt, A. Differential activity of F-box genes and E3 ligases distinguishes sexual versus apomictic germline specification in Boechera. J. Exp. Bot. 2019, 70, 5643–5657. [Google Scholar] [CrossRef] [Green Version]
- Florez Rueda, A.M.; Grossniklaus, U.; Schmidt, A. Laser-assisted Microdissection (LAM) as a Tool for Transcriptional Profiling of Individual Cell Types. J. Vis. Exp. JoVE 2016, 111, 53916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.; Schmid, M.W.; Grossniklaus, U. Analysis of plant germline development by high-throughput RNA profiling: Technical advances and new insights. Plant J. 2012, 70, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Schmid, M.W.; Schmidt, A.; Klostermeier, U.C.; Barann, M.; Rosenstiel, P.; Grossniklaus, U. A Powerful Method for Transcriptional Profiling of Specific Cell Types in Eukaryotes: Laser-Assisted Microdissection and RNA Sequencing. PLoS ONE 2012, 7, e29685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.; Wuest, S.E.; Vijverberg, K.; Baroux, C.; Kleen, D.; Grossniklaus, U. Transcriptome Analysis of the Arabidopsis Megaspore Mother Cell Uncovers the Importance of RNA Helicases for Plant Germline Development. PLoS Biol. 2011, 9, e1001155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, T.; Hu, Y.; Tucker, M.R.; Taylor, J.M.; Johnson, S.D.; Spriggs, A.; Tsuchiya, T.; Oelkers, K.; Rodrigures, J.C.; Koltunow, A.M. Enlarging Cells Initiating Apomixis in Hieracium praealtum Transition to an Embryo Sac Program prior to Entering Mitosis. Plant Physiol. 2013, 163, 216–231. [Google Scholar] [CrossRef] [Green Version]
- Juranic, M.; Tucker, M.R.; Schultz, C.J.; Shirley, N.J.; Taylor, J.M.; Spriggs, A.; Johnson, S.D.; Bulone, V.; Koltunow, A.M. Asexual Female Gametogenesis Involves Contact with a Sexually-Fated Megaspore in Apomictic Hieracium. Plant Physiol. 2018, 177, 1027–1049. [Google Scholar] [CrossRef] [Green Version]
- Yamada-Akiyama, H.; Akiyama, Y.; Ebina, M.; Xu, Q.; Tsuruta, S.-I.; Yazaki, J.; Kishimoto, N.; Kikuchi, S.; Takahara, M.; Takamizo, T. Analysis of expressed sequence tags in apomictic guineagrass (Panicum maximum). J. Plant Physiol. 2008, 166, 750–761. [Google Scholar] [CrossRef]
- Laspina, N.; Vega, T.; Seijo, G.; Gonzalez, A.; Martelotto, L.; Stein, J.; Podio, M.; Ortiz, J.P.A.; Echenique, V.C.; Quarin, C.L. Gene expression analysis at the onset of aposporous apomixis in Paspalum notatum. Plant Mol. Biol. 2008, 67, 615–628. [Google Scholar] [CrossRef]
- Cervigni, G.; Paniego, N.; Pessino, S.; Selva, J.P.; Diaz, M.; Spangenberg, G.; Echenique, V. Gene expression in diplosporous and sexual Eragrostis curvula genotypes with differing ploidy levels. Plant Mol. Biol. 2008, 67, 11–23. [Google Scholar] [CrossRef]
- Galla, G.; Vogel, H.; Sharbel, T.F.; Barcaccia, G. De novo sequencing of the Hypericum perforatum L. flower transcriptome to identify potential genes that are related to plant reproduction sensu lato. BMC Genom. 2015, 16, 254. [Google Scholar] [CrossRef] [Green Version]
- Rabiger, D.S.; Taylor, J.M.; Spriggs, A.; Hand, M.L.; Henderson, S.T.; Johnson, S.D.; Oelkers, K.; Hrmova, M.; Saito, K.; Suzuki, G. Generation of an integrated Hieracium genomic and transcriptomic resource enables exploration of small RNA pathways during apomixis initiation. BMC Biol. 2016, 14, 86. [Google Scholar] [CrossRef] [PubMed]
- Long, J.M.; Liu, Z.; Wu, X.M.; Fang, Y.N.; Jia, H.H.; Xie, Z.Z.; Deng, X.X.; Guo, W.W. Genome-scale mRNA and small RNA transcriptomic insights into initiation of citrus apomixis. J. Exp. Bot. 2016, 67, 5743–5756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.; Schmid, M.W.; Grossniklaus, U. Plant germline formation: Common concepts and developmental flexibility in sexual and asexual reproduction. Development 2015, 142, 229–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackenzie, A.; Heslop-Harrison, J.; Dickinson, H.G. Elimination of Ribosomes during Meiotic Prophase. Nature 1967, 215, 997–999. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Conner, J.; Ozias-Akins, P. Identification of ovule transcripts from the Apospory-Specific Genomic Region (ASGR)-carrier chromosome. BMC Genom. 2011, 12, 206. [Google Scholar] [CrossRef] [Green Version]
- Xue, S.; Barna, M. Specialized ribosomes: A new frontier in gene regulation and organismal biology. Nat. Rev. Mol. Cell Biol. 2012, 13, 355–369. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Zhu, C.; Guang, S.; Feng, X. The Functions of Non-coding RNAs in rRNA Regulation. Front. Genet. 2019, 10, 290. [Google Scholar] [CrossRef] [Green Version]
- Dalla Venezia, N.; Vincent, A.; Marcel, V.; Catez, F.; Diaz, J.-J. Emerging Role of Eukaryote Ribosomes in Translational Control. Int. J. Mol. Sci. 2019, 20, 1226. [Google Scholar] [CrossRef] [Green Version]
- Bazin, J.; Baerenfaller, K.; Gosai, S.J.; Gregory, B.D.; Crespi, M.; Bailey-Serres, J. Global analysis of ribosome-associated noncoding RNAs unveils new modes of translational regulation. Proc. Natl. Acad. Sci. USA 2017, 114, E10018–E10027. [Google Scholar] [CrossRef] [Green Version]
- Ding, D.-Q.; Haraguchi, T.; Hiraoka, Y. The role of chromosomal retention of noncoding RNA in meiosis. Chromosome Res. 2013, 21, 665–672. [Google Scholar] [CrossRef] [Green Version]
- Flórez-Zapata, N.M.V.; Reyes-Valdés, M.H.; Martínez, O. Long non-coding RNAs are major contributors to transcriptome changes in sunflower meiocytes with different recombination rates. BMC Genom. 2016, 17, 490. [Google Scholar] [CrossRef] [Green Version]
- Hafidh, S.; Čapková, V.; Honys, D. Safe Keeping the Message: mRNP Complexes Tweaking after Transcription. In RNA Infrastructure and Networks; Collins, L.J., Ed.; Springer: New York, NY, USA, 2011; pp. 118–136. [Google Scholar]
- Liu, Y.; Imai, R. Function of Plant DExD/H-Box RNA Helicases Associated with Ribosomal RNA Biogenesis. Front. Plant Sci. 2018, 9, 125. [Google Scholar] [CrossRef] [PubMed]
- Owttrim, G. RNA helicases and abiotic stress. Nucleic Acids Res. 2006, 34, 3220–3230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silveira, É.D.; Guimarães, L.A.; de Alencar Dusi, D.M.; da Silva, F.R.; Martins, N.F.; do Carmo Costa, M.M.; Alves-Ferreira, M.; de Campos Carneiro, V.T. Expressed sequence-tag analysis of ovaries of Brachiaria brizantha reveals genes associated with the early steps of embryo sac differentiation of apomictic plants. Plant Cell Rep. 2012, 31, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Gehring, M. Epigenetic dynamics during flowering plant reproduction: Evidence for reprogramming? New Phytol. 2019, 224, 91–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Aguilar, M.; Michaud, C.; Leblanc, O.; Grimanelli, D. Inactivation of a DNA Methylation Pathway in Maize Reproductive Organs Results in Apomixis-Like Phenotypes. Plant Cell 2010, 22, 3249–3267. [Google Scholar] [CrossRef] [Green Version]
- Olmedo-Monfil, V.; Durán-Figueroa, N.; Arteaga-Vandázquez, M.; Demesa-Arévalo, E.; Autran, D.; Grimanelli, D.; Grimanelli, D.; Slotkin, K.; Martienssen, R.A.; Vielle-Calzada, J.-P. Control of female gamete formation by a small RNA pathway in Arabidopsis. Nature 2010, 464, 628–632. [Google Scholar] [CrossRef]
- Singh, M.; Goel, S.; Meeley, R.B.; Dantec, C.; Parrinello, H.; Michaud, C.; Leblanc, O.; Grimanelli, D. Production of Viable Gametes without Meiosis in Maize Deficient for an ARGONAUTE Protein. Plant Cell 2011, 23, 443–458. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Lagana, E.; Rodriguez-Leal, D.; Lua, J.; Vielle-Calzada, J.P. A Multigenic Network of ARGONAUTE4 Clade Members Controls Early Megaspore Formation in Arabidopsis. Genetics 2016, 204, 1045–1056. [Google Scholar] [CrossRef] [Green Version]
- Meister, G. Argonaute proteins: Functional insights and emerging roles. Nat. Rev. Genet. 2013, 14, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Garbus, I.; Selva, J.P.; Pasten, M.C.; Bellido, A.M.; Carballo, J.; Albertini, E.; Echenique, V. Characterization and discovery of miRNA and miRNA targets from apomictic and sexual genotypes of Eragrostis curvula. BMC Genom. 2019, 20, 839. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, J.P.A.; Leblanc, O.; Rohr, C.; Grisolia, M.; Siena, L.A.; Podio, M.; Colono, C.; Azzaro, C.; Pessino, S.C. Small RNA-seq reveals novel regulatory components for apomixis in Paspalum notatum. BMC Genom. 2019, 20, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amiteye, S.; Corral, J.M.; Vogel, H.; Kuhlmann, M.; Mette, M.F.; Sharbel, T.F. Novel microRNAs and microsatellite-like small RNAs in sexual and apomictic Boechera species. MicroRNA 2013, 2, 45–62. [Google Scholar] [CrossRef] [PubMed]
- Amiteye, S.; Corral, J.M.; Vogel, H.; Sharbel, T.F. Analysis of conserved microRNAs in floral tissues of sexual and apomictic Boechera species. BMC Genom. 2011, 12, 500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravi, M.; Marimuthu, M.P.A.; Siddiqi, I. Gamete formation without meiosis in Arabidopsis. Nature 2008, 451, 1121. [Google Scholar] [CrossRef] [PubMed]
- Pawlowski, W.P.; Wang, C.-J.R.; Golubovskaya, I.N.; Szymaniak, J.M.; Shi, L.; Hamant, O.; Zhu, T.; Harper, L.; Sheridan, W.F.; Cande, W.Z. Maize AMEIOTIC1 is essential for multiple early meiotic processes and likely required for the initiation of meiosis. Proc. Natl. Acad. Sci. USA 2009, 106, 3603–3608. [Google Scholar] [CrossRef] [Green Version]
- D’Erfurth, I.; Cromer, L.; Jolivet, S.; Girard, C.; Horlow, C.; Sun, Y.; To, J.P.C.; Berchowitz, L.E.; Copenhaver, G.P.; Mercier, R. The CYCLIN-A CYCA1;2/TAM Is Required for the Meiosis I to Meiosis II Transition and Cooperates with OSD1 for the Prophase to First Meiotic Division Transition. PLoS Genet. 2010, 6, e1000989. [Google Scholar] [CrossRef] [Green Version]
- D’Erfurth, I.; Jolivet, S.; Froger, N.; Catrice, O.; Novatchkova, M.; Mercier, R. Turning Meiosis into Mitosis. PLoS Biol. 2009, 7, e1000124. [Google Scholar] [CrossRef] [Green Version]
- Mieulet, D.; Jolivet, S.; Rivard, M.; Cromer, L.; Vernet, A.; Mayonove, P.; Pereira, L.; Droc, G.; Courtois, B.; Guiderdoni, E. Turning rice meiosis into mitosis. Cell Res. 2016, 26, 1242–1254. [Google Scholar] [CrossRef]
- Marimuthu, M.P.A.; Jolivet, S.; Ravi, M.; Pereira, L.; Davda, J.N.; Cromer, L.; Wang, L.; Nogue, F.; Chan, S.W.L.; Siddiqu, I. Synthetic Clonal Reproduction Through Seeds. Science 2011, 331, 876. [Google Scholar] [CrossRef]
- Wang, C.; Liu, Q.; Shen, Y.; Hua, Y.; Wang, J.; Lin, J.; Wu, M.; Sun, T.; Cheng, Z.; Mercier, R. Clonal seeds from hybrid rice by simultaneous genome engineering of meiosis and fertilization genes. Nat. Biotechnol. 2019, 37, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Kelliher, T.; Starr, D.; Richbourg, L.; Chintamanani, S.; Delzer, B.; Nuccio, M.L.; Green, J.; Chen, Z.; McCuiston, J.; Wang, W. MATRILINEAL, a sperm-specific phospholipase, triggers maize haploid induction. Nature 2017, 542, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Pires, N.D. Seed evolution: Parental conflicts in a multi-generational household. Biomol. Concepts 2014, 5, 71–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprunck, S. Twice the fun, double the trouble: Gamete interactions in flowering plants. Curr. Opin. Plant Biol. 2020, 53, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Hands, P.; Rabiger, D.S.; Koltunow, A. Mechanisms of endosperm initiation. Plant Reprod. 2016, 29, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Vijverberg, K.; Ozias-Akins, P.; Schranz, M.E. Identifying and Engineering Genes for Parthenogenesis in Plants. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derkacheva, M.; Hennig, L. Variations on a theme: Polycomb group proteins in plants. J. Exp. Bot. 2013, 65, 2769–2784. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, J.C.M.; Tucker, M.R.; Johnson, S.D.; Hrmova, M.; Koltunow, A.M.G. Sexual and apomictic seed formation in Hieracium requires the plant polycomb-group gene FERTILIZATION INDEPENDENT ENDOSPERM. Plant Cell 2008, 20, 2372–2386. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Nolan, T.M.; Jiang, H.; Yin, Y. AP2/ERF Transcription Factor Regulatory Networks in Hormone and Abiotic Stress Responses in Arabidopsis. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Nic-Can, G.I.; López-Torres, A.; Barredo-Pool, F.; Wrobel, K.; Loyola-Vargas, V.M.; Rojas-Herrera, R.; De-la-Pena, C. New Insights into Somatic Embryogenesis: LEAFY COTYLEDON1, BABY BOOM1 and WUSCHEL-RELATED HOMEOBOX4 Are Epigenetically Regulated in Coffea canephora. PLoS ONE 2013, 8, e72160. [Google Scholar] [CrossRef] [Green Version]
- Kirioukhova, O.; Shah, J.N.; Larsen, D.S.; Tayyab, M.; Mueller, N.E.; Govind, G.; Baroux, C.; Federer, M.; Gheyselinck, J.; Barrell, P.J. Aberrant imprinting may underlie evolution of parthenogenesis. Sci. Rep. 2018, 8, 10626. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, J.C.; Ferree, P.M. Genome Silencing and Elimination: Insights from a “Selfish” B Chromosome. Front Genet. 2017, 8, 50. [Google Scholar] [CrossRef] [Green Version]
- Hörandl, E.; Speijer, D. How oxygen gave rise to eukaryotic sex. Proc. R. Soc. B Biol. Sci. 2018, 285, 20172706. [Google Scholar] [CrossRef] [Green Version]
- Mirzaghaderi, G.; Horandl, E. The evolution of meiotic sex and its alternatives. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161221. [Google Scholar] [CrossRef]
- Horandl, E.; Hadacek, F. The oxidative damage initiation hypothesis for meiosis. Plant Reprod. 2013, 26, 351–367. [Google Scholar] [CrossRef] [Green Version]
- Kelliher, T.; Walbot, V. Hypoxia Triggers Meiotic Fate Acquisition in Maize. Science 2012, 337, 345–348. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Bramsiepe, J.; Van Durme, M.; Komaki, S.; Prusicki, M.A.; Maruyama, D.; Forner, J.; Medzihradszky, A.; Wijnker, E.; Harashima, H. RETINOBLASTOMA RELATED1 mediates germline entry in Arabidopsis. Science 2017, 356, eaaf6532. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ullah, F.; Zhou, D.-X.; Yi, M.; Zhao, Y. Mechanisms of ROS Regulation of Plant Development and Stress Responses. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Ha, H.C.; Sirisoma, N.S.; Kuppusamy, P.; Zweier, J.L.; Woster, P.M.; Casero, R.A. The natural polyamine spermine functions directly as a free radical scavenger. Proc. Natl. Acad. Sci. USA 1998, 95, 11140–11145. [Google Scholar] [CrossRef] [Green Version]
- Hong, Z.; Zhang, Z.; Olson, J.M.; Verma, D.P.S. A Novel UDP-Glucose Transferase Is Part of the Callose Synthase Complex and Interacts with Phragmoplastin at the Forming Cell Plate. Plant Cell 2001, 13, 769–779. [Google Scholar] [CrossRef]
- Luna, E.; Pastor, V.; Robert, J.; Flors, V.; Mauch-Mani, B.; Ton, J. Callose deposition: A multifaceted plant defense response. Mol. Plant Microbe Interact. MPMI 2011, 24, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Rodkiewicz, B. Callose in cell walls during mega-sporogenesis in Angiosperms. Planta 1970, 93, 39–47. [Google Scholar] [CrossRef]
- Koltunow, A.M. Apomixis: Embryo Sacs and Embryos Formed without Meiosis or Fertilization in Ovules. Plant Cell 1993, 5, 1425–1437. [Google Scholar] [CrossRef] [Green Version]
- De Storme, N.; Geelen, D. The impact of environmental stress on male reproductive development in plants: Biological processes and molecular mechanisms. Plant Cell Environ. 2014, 37, 1–18. [Google Scholar] [CrossRef]
- Otsubo, Y.; Nakashima, A.; Yamamoto, M.; Yamashita, A. TORC1-Dependent Phosphorylation Targets in Fission Yeast. Biomolecules 2017, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Sherwood, D.A. A Simple Metabolic Switch May Activate Apomixis in Arabidopsis thaliana. Ph.D. Thesis, Utah University, Salt Lake City, UT, USA, 2018. [Google Scholar]
- Pfeiffer, A.; Janocha, D.; Dong, Y.; Medzihradszky, A.; Schone, S.; Daum, G.; Suzaki, T.; Forner, J.; Langenecker, T.; Rempel, E. Integration of light and metabolic signals for stem cell activation at the shoot apical meristem. Elife 2016, 5, e17023. [Google Scholar] [CrossRef]
- Mayer, C.; Grummt, I. Ribosome biogenesis and cell growth: mTOR coordinates transcription by all three classes of nuclear RNA polymerases. Oncogene 2006, 25, 6384–6391. [Google Scholar] [CrossRef] [Green Version]
- Grossmann, J.; Fernández, H.; Chaubey, P.M.; Valdés, A.E.; Gagliardini, V.; Cañal, M.J.; Russo, G.; Grossniklaus, U. Proteogenomic Analysis Greatly Expands the Identification of Proteins Related to Reproduction in the Apogamous Fern Dryopteris affinis ssp. affinis. Front. Plant Sci. 2017, 8, 336. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene | Type of Apomixis | Element of Apomixis | Locus | Plant Family | Species | Publication |
|---|---|---|---|---|---|---|
| ARI7 | gametophytic | apospory | HAPPY | Hypericaceae | Hypericum perforatum | [44,62] |
| APOLLO | gametophytic | female apomeiosis | - | Brassicaceae | Boechera ssp. | [63,64] |
| UPG2 | gametophytic | male apomeiosis | - | Brassicaceae | Boechera ssp. | [65] |
| QGJ | gametophytic | apospory | - | Poaceae | Paspalum notatum | [67] |
| ORC3 | gametophytic | endosperm formation | ACL | Poaceae | Paspalum simplex | [51,68] |
| BBM(L) | gametophytic | parthenogensis | ASGR | Poaceae | Pennisetum squamulatum Cenchrus ciliaris Brachiara decumbens | [49,69,70,73,74] |
| RKD | sporophytic | somatic embryogenesis | - | Rutaceae | Citrus | [5,76] |
| Plant Family | Species | Type of Apomixis | Tissues Profiled | Methods of Analysis | References |
|---|---|---|---|---|---|
| Poaceae | Pennisetum ciliare | gametophytic | unpollinated ovaries | modified differential display | [83] |
| Poaceae | Pennisetum glaucum | gametophytic | spikelets at 4 developmental stages (pre-meiosis, meiocyte, gametogenesis, mature gametophyte) | suppression subtractive hybridization | [84] |
| Poaceae | Panicum maximum | gametophytic | flower buds | cDNA library | [85] |
| Poaceae | Panicum maximum | gametophytic | spikelets (pre-meiosis) | RNA-Seq (Illumina HiSeq2500) | [86] |
| Poaceae | Panicum maximum | gametophytic | immature pistils | custom microarray | [110] |
| Poaceae | Paspalum notatum | gametophytic | inflorescences at 4 developmental stages (early premeiosis; late premeiosis/ meiosis; postmeiosis; anthesis) | RNA-Seq (Roche 454) | [94] |
| Poaceae | Paspalum notatum | gametophytic | florets at different developmental stages | cDNA-AFLP | [92] |
| Poaceae | Paspalum notatum | gametophytic | inflorescences | differential display analysis | [91,111] |
| Poaceae | Eragostris curvula | gametophytic | panicles | differential display analysis | [112] |
| Poaceae | Eragostris curvula | gametophytic | spikelets with embryo sacs at all developmental stages | RNA-Seq (Roche 454) | [95] |
| Poaceae | Poa pratensis | gametophytic | florets at 4 developmental stages (pre-meiosis; meiosis; post-meiosis; anthesis) | cDNA-AFLP | [87,88] |
| Hypericaceae | Hypericum perforatum | gametophytic | nucellus tissues harboring MMC or AIC before (apo)meiosis | RNA-Seq (Illumina NextSeq500) | [62] |
| Hypericaceae | Hypericum perforatum | gametophytic | pistils | custom microarray | [98] |
| Hypericaceae | Hypericum perforatum | gametophytic | whole flowers at range of developmental stages | cDNA libraries | [113] |
| Asteraceae | Hieracium praealtum Hieracium aurantiacum; parthenogenesis incapable accession lop138 | gametophytic | ovules and ovaries at different developmental stages isolated by manual microdissection | RNA-Seq (Illumina HiSeq2000) | [100] |
| Asteraceae | Hieracium praealtum | gametophytic | AIC, developing female gametophytes (2–4 nucleate), and somatic ovule cells isolated by LAM | RNA-Seq (Roche 454; Illumina HiSeq2000) | [108,109] |
| Asteraceae | Hieracium praealtum | gametophytic | ovaries | RNA-Seq (Illumina HiSeq2000) | [114] |
| Urticaceae | Boehmeria tricuspis | gametophytic apomixis | Flowers at 4 developmental stages (MMC; FMS, embryo sac, mature embryo) | RNA-Seq (Illumina HiSeq4000) | [88] |
| Brassicaceae | Boechera | gametophytic | nucellus tissues harboring MMC or AIC isolated by LAM | RNA-Seq (Illumina NextSeq500) | [103] |
| Brassicaceae | Boechera gunnisoniana | gametophytic | AIC, egg cell, central cell, synergids isolated by LAM | ATH1 microarray, RNA-Seq (SOLiD V4) | [81] |
| Brassicaceae | Boechera | gametophytic | ovules isolated by manual microdissection | RNA-Seq (Roche 454); custom microarray | [63] |
| Brassicaceae | Boechera | gametophytic | antherheads at pollen mother cell stage | custom microarray | [65] |
| Brassicaceae | Boechera | gametophytic | ovules isolated by manual microdissection at 4 developmental stages (early premeiosis; late premeiosis; FMS, gametophyte) | SuperSAGE | [101] |
| Brassicaceae | Boechera | gametophytic | ovules isolated by manual microdissection pooled flower stages | SuperSAGE; RNA-Seq (Roche 454) | [102] |
| Fabaceae | Medicago sativa | apomeiotic mutant | flower buds at 4 developmental stages (pre-meiosis, initial meiosis, final meiosis, and post-meiosis) | cDNA-AFLP | [96] |
| Rutaceae | Citrus | somatic embryogensis | fruits 15, 30, 45, and 60 d after flowering | custom microarray | [76] |
| Rutaceae | Citrus | somatic embryogensis | leaves, ovules, seeds, fruits | RNA-Seq (Illumina Genome Analyzer) | [5] |
| Rutaceae | Citrus | somatic embryogensis | ovaries at anthesis and at 3, 7, 17, 21, and 28 d after flowering | RNA-Seq (Illumina Genome Analyzer) | [115] |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmidt, A. Controlling Apomixis: Shared Features and Distinct Characteristics of Gene Regulation. Genes 2020, 11, 329. https://doi.org/10.3390/genes11030329
Schmidt A. Controlling Apomixis: Shared Features and Distinct Characteristics of Gene Regulation. Genes. 2020; 11(3):329. https://doi.org/10.3390/genes11030329
Chicago/Turabian StyleSchmidt, Anja. 2020. "Controlling Apomixis: Shared Features and Distinct Characteristics of Gene Regulation" Genes 11, no. 3: 329. https://doi.org/10.3390/genes11030329
APA StyleSchmidt, A. (2020). Controlling Apomixis: Shared Features and Distinct Characteristics of Gene Regulation. Genes, 11(3), 329. https://doi.org/10.3390/genes11030329