Genetics and Epigenetics of Atopic Dermatitis: An Updated Systematic Review

 , , ,
, , ,
Abstract
:1. Introduction
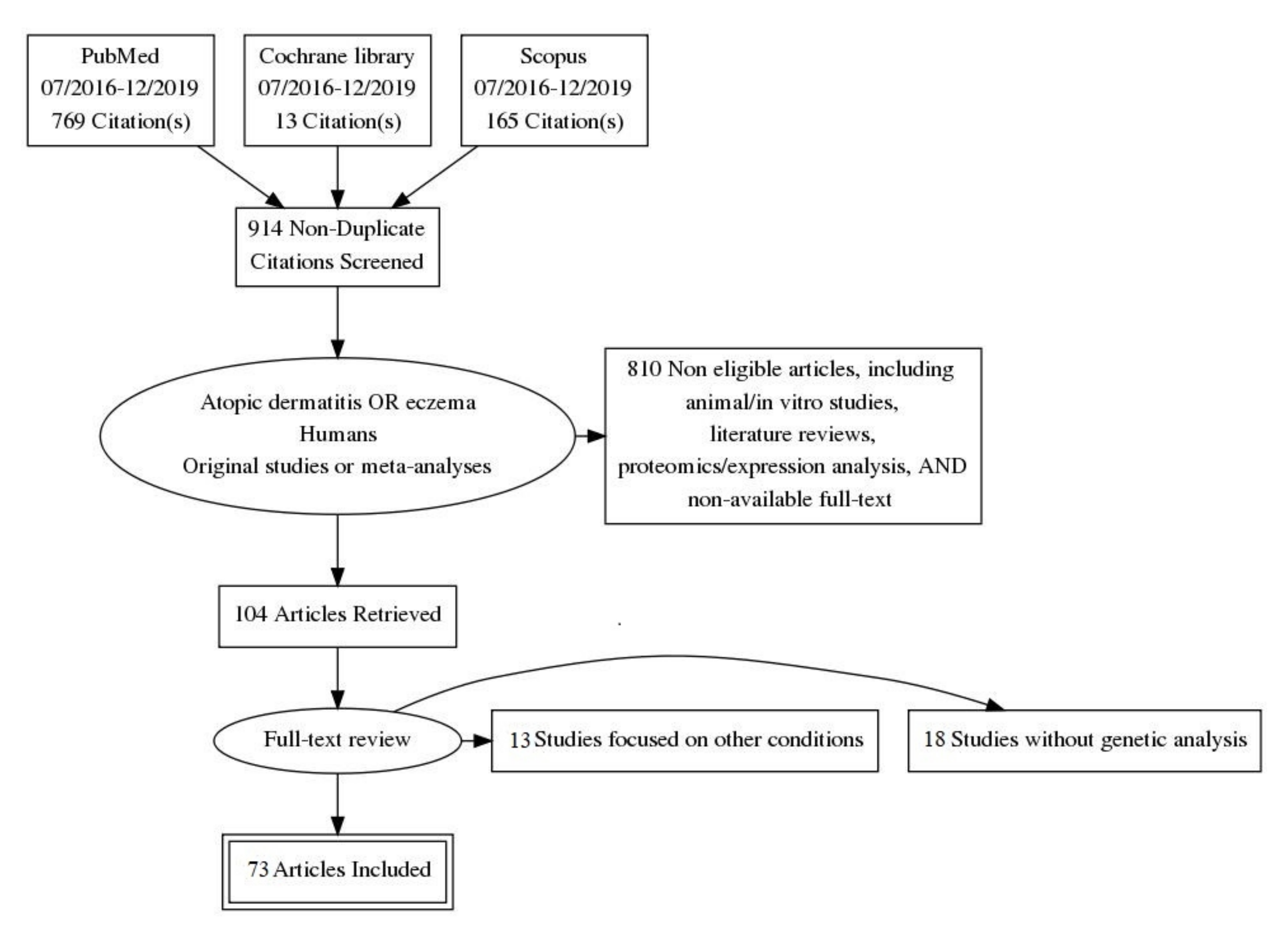
2. Materials and Methods
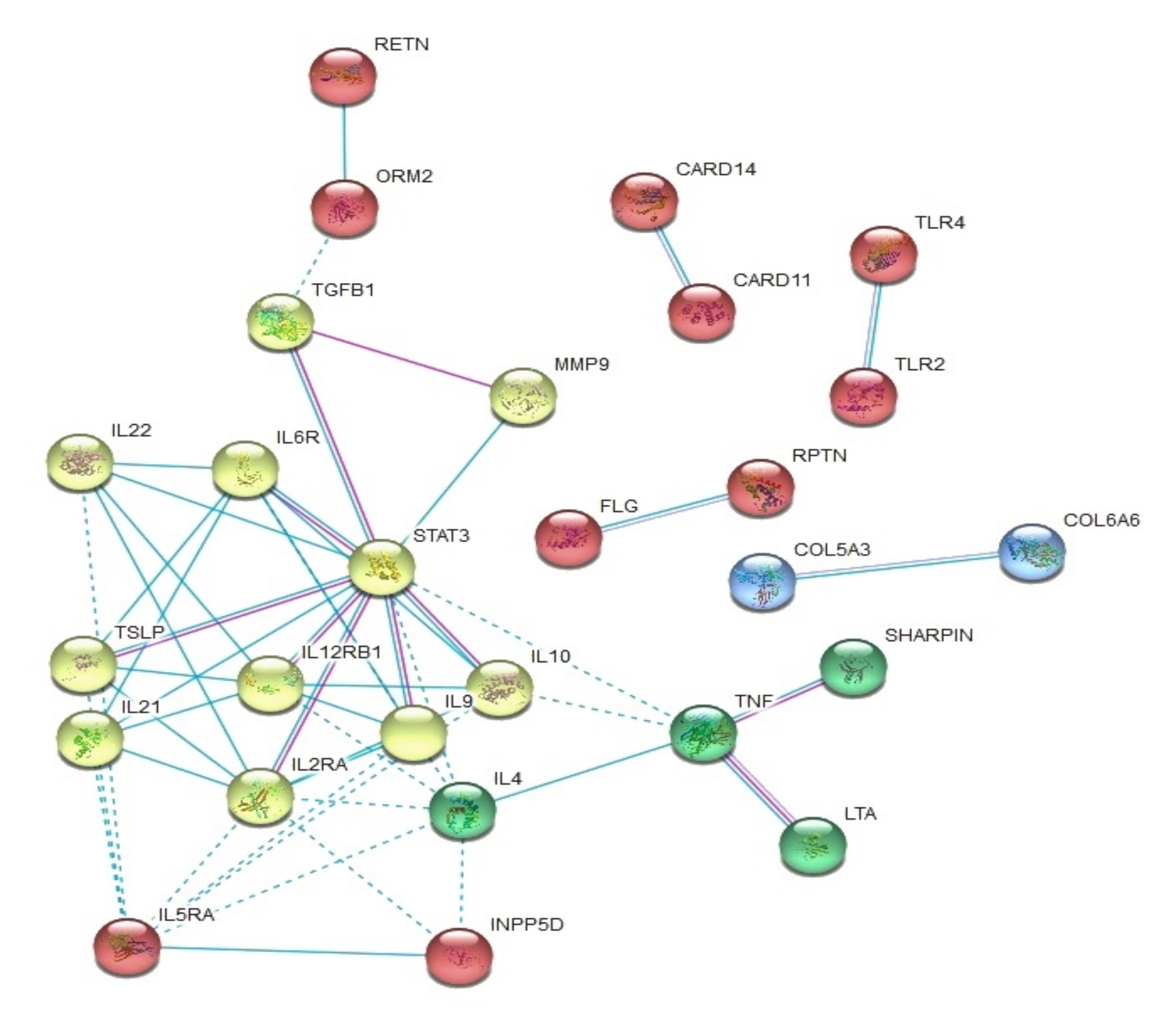
3. Results
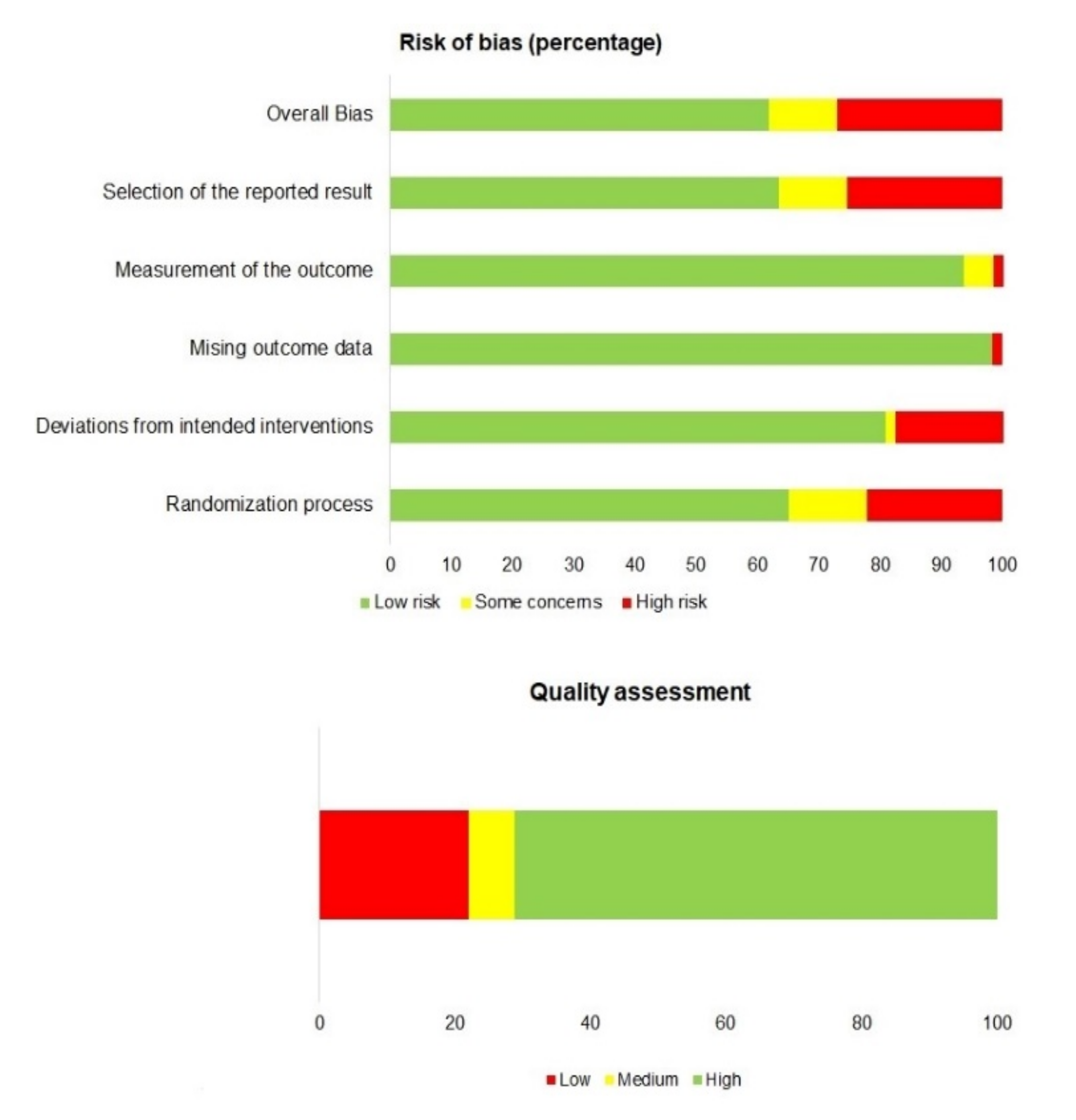
3.1. Selection, Bias and Quality of Articles
3.2. Genetic Studies
3.3. Filaggrin
3.3.1. Filaggrin Mutations and Other Allergic Diseases
3.3.2. Filaggrin Mutations and Early Onset of AD
3.3.3. Filaggrin Mutations and other Skin Diseases
3.3.4. Filaggrin Mutations and Eczema Severity
3.3.5. Filaggrin Mutations and Ethnicity Risk Factors
3.4. Other Genes
3.5. Epigenetic Studies
4. Discussion
4.1. Filaggrin
4.2. Other Genes
4.3. AD Epigenetics
5. Final Remarks
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Weidinger, S.; Beck, L.A.; Bieber, T.; Kabashima, K.; Irvine, A.D. Atopic dermatitis. Nat. Rev. Dis. Prim. 2018, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Silverberg, J.I. Atopic Dermatitis in Adults. Med. Clin. N. Am. 2020, 104, 157–176. [Google Scholar] [CrossRef] [PubMed]
- Dharmage, S.C.; Lowe, A.J.; Matheson, M.C.; Burgess, J.A.; Allen, K.J.; Abramson, M.J. Atopic dermatitis and the atopic march revisited. Allergy 2014, 69, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Bonamonte, D.; Filoni, A.; Vestita, M.; Romita, P.; Foti, C.; Angelini, G. The Role of the Environmental Risk Factors in the Pathogenesis and Clinical Outcome of Atopic Dermatitis. Biomed. Res. Int. 2019, 2019, 2450605. [Google Scholar] [CrossRef]
- Manousaki, D.; Paternoster, L.; Standl, M.; Moffatt, M.F.; Farrall, M.; Bouzigon, E.; Strachan, D.P.; Demenais, F.; Lathrop, M.; Cookson, W.O.C.M.; et al. Vitamin D levels and susceptibility to asthma, elevated immunoglobulin E levels, and atopic dermatitis: A Mendelian randomization study. PLoS Med. 2017, 14, e1002294. [Google Scholar] [CrossRef] [PubMed]
- Schram, M.E.; Tedja, A.M.; Spijker, R.; Bos, J.D.; Williams, H.C.; Spuls, P.I. Is there a rural/urban gradient in the prevalence of eczema? A systematic review. Br. J. Dermatol. 2010, 162, 964–973. [Google Scholar] [CrossRef]
- Romieu, I.; Torrent, M.; Garcia-Esteban, R.; Ferrer, C.; Ribas-Fitó, N.; Antó, J.M.; Sunyer, J. Maternal fish intake during pregnancy and atopy and asthma in infancy. Clin. Exp. Allergy 2007, 37, 518–525. [Google Scholar] [CrossRef]
- Leermakers, E.T.M.; Sonnenschein-van der Voort, A.M.M.; Heppe, D.H.M.; de Jongste, J.C.; Moll, H.A.; Franco, O.H.; Hofman, A.; Jaddoe, V.W.V.; Duijts, L. Maternal fish consumption during pregnancy and risks of wheezing and eczema in childhood: The Generation R Study. Eur. J. Clin. Nutr. 2013, 67, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Willers, S.M.; Devereux, G.; Craig, L.C.A.; McNeill, G.; Wijga, A.H.; Abou El-Magd, W.; Turner, S.W.; Helms, P.J.; Seaton, A. Maternal food consumption during pregnancy and asthma, respiratory and atopic symptoms in 5-year-old children. Thorax 2007, 62, 773–779. [Google Scholar] [CrossRef] [Green Version]
- Apfelbacher, C.J.; Diepgen, T.L.; Schmitt, J. Determinants of eczema: Population-based cross-sectional study in Germany. Allergy 2011, 66, 206–213. [Google Scholar] [CrossRef]
- Dalgard, F.J.; Gieler, U.; Tomas-Aragones, L.; Lien, L.; Poot, F.; Jemec, G.B.E.; Misery, L.; Szabo, C.; Linder, D.; Sampogna, F.; et al. The Psychological Burden of Skin Diseases: A Cross-Sectional Multicenter Study among Dermatological Out-Patients in 13 European Countries. J. Investig. Dermatol. 2015, 135, 984–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, Y.M.F.; Egeberg, A.; Skov, L.; Thyssen, J.P. Comorbidities of Atopic Dermatitis: Beyond Rhinitis and Asthma. Curr. Dermatol. Rep. 2017, 6, 35–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, M.A.; Vonk, J.M.; Baurecht, H.; Marenholz, I.; Tian, C.; Hoffman, J.D.; Helmer, Q.; Tillander, A.; Ullemar, V.; van Dongen, J.; et al. Shared genetic origin of asthma, hay fever and eczema elucidates allergic disease biology. Nat. Genet. 2017, 49, 1752–1757. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.A.R.R.; Vonk, J.M.; Baurecht, H.; Marenholz, I.; Tian, C.; Hoffman, J.D.; Helmer, Q.; Tillander, A.; Ullemar, V.; Lu, Y.; et al. Eleven loci with new reproducible genetic associations with allergic disease risk. J. Allergy Clin. Immunol. 2019, 143, 691–699. [Google Scholar] [CrossRef] [Green Version]
- Elmose, C.; Thomsen, S.F. Twin Studies of Atopic Dermatitis: Interpretations and Applications in the Filaggrin Era. J. Allergy 2015, 2015, 902359. [Google Scholar] [CrossRef] [Green Version]
- Barnes, K.C. An update on the genetics of atopic dermatitis: Scratching the surface in 2009. J. Allergy Clin. Immunol. 2010, 125, 16–29. [Google Scholar] [CrossRef]
- Bin, L.; Leung, D.Y.M. Genetic and epigenetic studies of atopic dermatitis. Allergy Asthma Clin. Immunol. 2016, 12, 52. [Google Scholar] [CrossRef] [Green Version]
- Guyatt, G.; Oxman, A.D.; Akl, E.A.; Kunz, R.; Vist, G.; Brozek, J.; Norris, S.; Falck-Ytter, Y.; Glasziou, P.; Debeer, H.; et al. GRADE guidelines: 1. Introduction—GRADE evidence profiles and summary of findings tables. J. Clin. Epidemiol. 2011, 64, 383–394. [Google Scholar] [CrossRef]
- Sterne, J.A.C.; Savović, J.; Page, M.J.; Elbers, R.G.; Blencowe, N.S.; Boutron, I.; Cates, C.J.; Cheng, H.-Y.; Corbett, M.S.; Eldridge, S.M.; et al. RoB 2: A revised Cochrane risk-of-bias tool for randomized trials. BMJ 2019, 366, l48981. [Google Scholar]
- Stang, A. Critical evaluation of the Newcastle-Ottawa scale for the assessment of the quality of nonrandomized studies in meta-analyses. Eur. J. Epidemiol. 2010, 25, 603–605. [Google Scholar] [CrossRef] [Green Version]
- Pathan, M.; Keerthikumar, S.; Ang, C.; Gangoda, L.; Quek, C.Y.J.; Williamson, N.A.; Mouradov, D.; Sieber, O.M.; Simpson, R.J.; Salim, A. FunRich: An open access standalone functional enrichment and interaction network analysis tool. Proteomics 2015, 15, 2597–2601. [Google Scholar] [CrossRef] [PubMed]
- Jassal, B.; Matthews, L.; Viteri, G.; Gong, C.; Lorente, P.; Fabregat, A.; Sidiropoulos, K.; Cook, J.; Gillespie, M.; Haw, R.; et al. The reactome pathway knowledgebase. Nucleic Acids Res. 2020, 48, D498–D503. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Shao, X.; Li, X.; Zhai, Y.; Zhang, Y.; Wang, S.; Fang, H. Identification of candidate genes in atopic dermatitis based on bioinformatic methods. Int. J. Dermatol. 2016, 55, 791–800. [Google Scholar] [CrossRef]
- Yang, Z.; Zeng, B.; Wang, C.; Wang, H.; Huang, P.; Pan, Y. MicroRNA-124 alleviates chronic skin inflammation in atopic eczema via suppressing innate immune responses in keratinocytes. Cell. Immunol. 2017, 319, 53–60. [Google Scholar] [CrossRef]
- Dissanayake, E.; Inoue, Y.; Ochiai, S.; Eguchi, A.; Nakano, T.; Yamaide, F.; Hasegawa, S.; Kojima, H.; Suzuki, H.; Mori, C.; et al. Hsa-mir-144-3p expression is increased in umbilical cord serum of infants with atopic dermatitis. J. Allergy Clin. Immunol. 2019, 143, 447–450. [Google Scholar] [CrossRef] [Green Version]
- Bao, L.; Chau, C.; Bao, J.; Tsoukas, M.M.; Chan, L.S. IL-4 dysregulates microRNAs involved in inflammation, angiogenesis and apoptosis in epidermal keratinocytes. Microbiol. Immunol. 2018, 62, 732–736. [Google Scholar] [CrossRef]
- Boorgula, M.P.; Taub, M.A.; Rafaels, N.; Daya, M.; Campbell, M.; Chavan, S.; Shetty, A.; Cheadle, C.; Barkataki, S.; Fan, J.; et al. Replicated methylation changes associated with eczema herpeticum and allergic response. Clin. Epigenetics 2019, 11, 122. [Google Scholar] [CrossRef] [Green Version]
- Carreras-Badosa, G.; Runnel, T.; Plaas, M.; Karner, J.; Ruckert, B.; Lattekivi, F.; Koks, S.; Akdis, C.A.; Kingo, K.; Rebane, A. microRNA-146a is linked to the production of IgE in mice but not in atopic dermatitis patients. Allergy 2018, 73, 2400–2403. [Google Scholar] [CrossRef]
- Chen, X.-F.; Zhang, L.-J.; Zhang, J.; Dou, X.; Shao, Y.; Jia, X.-J.; Zhang, W.; Yu, B. MiR-151a is involved in the pathogenesis of atopic dermatitis by regulating interleukin-12 receptor beta2. Exp. Dermatol. 2018, 27, 427–432. [Google Scholar] [CrossRef]
- Kumar, D.; Puan, K.J.; Andiappan, A.K.; Lee, B.; Westerlaken, G.H.A.; Haase, D.; Melchiotti, R.; Li, Z.; Yusof, N.; Lum, J.; et al. A functional SNP associated with atopic dermatitis controls cell type-specific methylation of the VSTM1 gene locus. Genome Med. 2017, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- Li, H.M.; Xiao, Y.J.; Min, Z.S.; Tan, C. Identification and interaction analysis of key genes and microRNAs in atopic dermatitis by bioinformatics analysis. Clin. Exp. Dermatol. 2019, 44, 257–264. [Google Scholar] [CrossRef]
- Thürmann, L.; Grützmann, K.; Klös, M.; Bieg, M.; Winter, M.; Polte, T.; Bauer, T.; Schick, M.; Bewerunge-Hudler, M.; Röder, S.; et al. Early-onset childhood atopic dermatitis is related to NLRP2 repression. J. Allergy Clin. Immunol. 2018, 141, 1482–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.-W.; Hojer, C.D.; Zhou, M.; Wu, X.; Wuster, A.; Lee, W.P.; Yaspan, B.L.; Chan, A.C. Regulation of T Cell Receptor Signaling by DENND1B in T H 2 Cells and Allergic Disease. Cell 2016, 164, 141–155. [Google Scholar] [CrossRef]
- Asad, S.; Winge, M.C.G.; Wahlgren, C.-F.; Bilcha, K.D.; Nordenskjöld, M.; Taylan, F.; Bradley, M. The tight junction gene Claudin-1 is associated with atopic dermatitis among Ethiopians. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 1939–1941. [Google Scholar] [CrossRef] [PubMed]
- Gimalova, G.F.; Karunas, A.S.; Fedorova, Y.Y.; Khusnutdinova, E.K. The study of filaggrin gene mutations and copy number variation in atopic dermatitis patients from Volga-Ural region of Russia. Gene 2016, 592, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Handa, S.; Khullar, G.; Pal, A.; Kamboj, P.; De, D. Filaggrin gene mutations in hand eczema patients in the Indian subcontinent: A prospective case-control study. Contact Dermat. 2019, 80, 359–364. [Google Scholar] [CrossRef]
- Jiang, X.-Y.; Zhao, J.-H.; Yu, C.-X.; Fang, L.; Zheng, X.-D.; Yin, X.-Y.; Wu, Y.-Y.; Tang, X.-F.; Zhou, F.-S.; Zhang, X.-J.; et al. Association analyses identify two susceptibility loci 5q31 and 5q22.1 for atopic dermatitis in Chinese Han population. Asian Pac. J. Allergy Immunol. 2018, 35, 196–202. [Google Scholar]
- Johansson, E.K.; Bergström, A.; Kull, I.; Lind, T.; Söderhäll, C.; van Hage, M.; Wickman, M.; Ballardini, N.; Wahlgren, C.-F. IgE sensitization in relation to preschool eczema and filaggrin mutation. J. Allergy Clin. Immunol. 2017, 140, 1572–1579. [Google Scholar] [CrossRef] [Green Version]
- Johansson, E.; Biagini Myers, J.M.; Martin, L.J.; He, H.; Pilipenko, V.; Mersha, T.; Weirauch, M.; Salomonis, N.; Ryan, P.; LeMasters, G.K.; et al. KIF3A genetic variation is associated with pediatric asthma in the presence of eczema independent of allergic rhinitis. J. Allergy Clin. Immunol. 2017, 140, 595–598. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Yoo, J.; Kim, J.; Park, J.; Han, E.; Jang, W.; Chae, H.; Lee, J.H.; Park, Y.M.; Kim, Y. Association of FLG single nucleotide variations with clinical phenotypes of atopic dermatitis. PLoS ONE 2017, 12, e0190077. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.S.; Choi, J.; Hahn, H.-J.; Lee, Y.-B.; Yu, D.-S.; Kim, J.-W. Association of Macrophage Migration Inhibitory Factor Polymorphisms with Total Plasma IgE Levels in Patients with Atopic Dermatitis in Korea. PLoS ONE 2016, 11, e0162477. [Google Scholar] [CrossRef]
- Ko, E.J.; Heo, W.I.; Park, K.Y.; Lee, M.-K.; Seo, S.J. Genetic polymorphism of thymic stromal lymphopoietin in Korean patients with atopic dermatitis and allergic march. J. Eur. Acad. Dermatol. Venereol. 2018, 32, e468–e470. [Google Scholar] [CrossRef] [PubMed]
- Leitch, C.S.; Natafji, E.; Yu, C.; Abdul-Ghaffar, S.; Madarasingha, N.; Venables, Z.C.; Chu, R.; Fitch, P.M.; Muinonen-Martin, A.J.; Campbell, L.E.; et al. Filaggrin-null mutations are associated with increased maturation markers on Langerhans cells. J. Allergy Clin. Immunol. 2016, 138, 482–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Oh, W.J.; Park, K.Y.; Kim, K.-H.; Seo, S.J. FLG mutations in the East Asian atopic dermatitis patients: Genetic and clinical implication. Exp. Dermatol. 2016, 25, 816–818. [Google Scholar] [CrossRef] [PubMed]
- Banihani, S.A.; Abu-Alia, K.F.; Khabour, O.F.; Alzoubi, K.H. Association between Resistin Gene Polymorphisms and Atopic Dermatitis. Biomolecules 2018, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Luukkonen, T.M.; Kiiski, V.; Ahola, M.; Mandelin, J.; Virtanen, H.; Poyhonen, M.; Kivirikko, S.; Surakka, I.; Reitamo, S.; Palotie, A.; et al. The Value of FLG Null Mutations in Predicting Treatment Response in Atopic Dermatitis: An Observational Study in Finnish Patients. Acta Derm. Venereol. 2017, 97, 456–463. [Google Scholar] [CrossRef] [Green Version]
- Manti, S.; Amorini, M.; Cuppari, C.; Salpietro, A.; Porcino, F.; Leonardi, S.; Del Giudice, M.M.; Marseglia, G.; Caimmi, D.P.; Salpietro, C. Filaggrin mutations and Molluscum contagiosum skin infection in patients with atopic dermatitis. Ann. Allergy. Asthma Immunol. 2017, 119, 446–451. [Google Scholar] [CrossRef]
- Morizane, S.; Ouchida, M.; Sunagawa, K.; Sugimoto, S.; Kobashi, M.; Sugihara, S.; Nomura, H.; Tsuji, K.; Sato, A.; Miura, Y.; et al. Analysis of All 34 Exons of the SPINK5 Gene in Japanese Atopic Dermatitis Patients. Acta Med. Okayama 2018, 72, 275–282. [Google Scholar]
- On, H.R.; Lee, S.E.; Kim, S.C.S.E.; Hong, W.J.; Kim, H.J.; Nomura, T.; Suzuki, S.; Shimizu, H.; Kim, S.C.S.E. Filaggrin Mutation in Korean Patients with Atopic Dermatitis. Yonsei Med. J. 2017, 58, 395–400. [Google Scholar] [CrossRef]
- Park, K.Y.; Li, K.; Seok, J.; Seo, S.J. An Analysis of the Filaggrin Gene Polymorphism in Korean Atopic Dermatitis Patients. J. Korean Med. Sci. 2016, 31, 1136–1142. [Google Scholar] [CrossRef] [Green Version]
- Ponińska, J.K.; Samoliński, B.; Tomaszewska, A.; Raciborski, F.; Samel-Kowalik, P.; Walkiewicz, A.; Lipiec, A.; Piekarska, B.; Krzych-Fałta, E.; Namysłowski, A.; et al. Haplotype dependent association of rs7927894 (11q13.5) with atopic dermatitis and chronic allergic rhinitis: A study in ECAP cohort. PLoS ONE 2017, 12, e0183922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roekevisch, E.; Leeflang, M.M.G.; Schram, M.E.; Campbell, L.E.; Irwin McLean, W.H.; Kezic, S.; Bos, J.D.; Spuls, P.I.; Middelkamp-Hup, M.A. Patients with atopic dermatitis with filaggrin loss-of-function mutations show good but lower responses to immunosuppressive treatment. Br. J. Dermatol. 2017, 177, 1745–1746. [Google Scholar] [CrossRef] [Green Version]
- Sekiya, A.; Kono, M.; Tsujiuchi, H.; Kobayashi, T.; Nomura, T.; Kitakawa, M.; Suzuki, N.; Yamanaka, K.; Sueki, H.; McLean, W.H.I.; et al. Compound heterozygotes for filaggrin gene mutations do not always show severe atopic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 158–162. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Wang, J.; Zhu, J.; Liang, Y. Down-regulated SHARPIN may accelerate the development of atopic dermatitis through activating interleukin-33/ST2 signalling. Exp. Dermatol. 2018, 27, 1328–1335. [Google Scholar] [CrossRef] [PubMed]
- Teye, K.; Numata, S.; Krol, R.P.; Ishii, N.; Matsuda, M.; Lee, J.-B.; Hamada, T.; Hashimoto, T. Prevalence of filaggrin gene mutations in patients with atopic dermatitis and ichthyosis vulgaris in Kyushu area of Japan and South Korea. J. Dermatol. Sci. 2017, 86, 174–177. [Google Scholar] [CrossRef]
- Behniafard, N.; Amirzargar, A.A.; Gharagozlou, M.; Delavari, F.; Hosseinverdi, S.; Sotoudeh, S.; Farhadi, E.; Mahmoudi, M.; Khaledi, M.; Moghaddam, Z.G.; et al. Single nucleotide polymorphisms of the genes encoding IL-10 and TGF-β1 in Iranian children with atopic dermatitis. Allergol. Immunopathol. 2018, 46, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, S.F.; Elmose, C.; Szecsi, P.B.; Stender, S.; Kyvik, K.O.; Backer, V.; Thyssen, J.P. Filaggrin gene loss-of-function mutations explain discordance of atopic dermatitis within dizygotic twin pairs. Int. J. Dermatol. 2016, 55, 1341–1344. [Google Scholar] [CrossRef]
- Thorsteinsdottir, S.; Stokholm, J.; Thyssen, J.P.; Norgaard, S.; Thorsen, J.; Chawes, B.L.; Bonnelykke, K.; Waage, J.; Bisgaard, H. Genetic, Clinical, and Environmental Factors Associated with Persistent Atopic Dermatitis in Childhood. JAMA Dermatol. 2019, 155, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Trzeciak, M.; Sakowicz-Burkiewicz, M.; Wesserling, M.; Glen, J.; Dobaczewska, D.; Bandurski, T.; Nowicki, R.; Pawelczyk, T. Altered Expression of Genes Encoding Cornulin and Repetin in Atopic Dermatitis. Int. Arch. Allergy Immunol. 2017, 172, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Tyurin, Y.A.; Shamsutdinov, A.F.; Kalinin, N.N.; Sharifullina, A.A.; Reshetnikova, I.D. Association of Toll-Like Cell Receptors TLR2 (p.Arg753GLN) and TLR4 (p.Asp299GLY) Polymorphisms with Indicators of General and Local Immunity in Patients with Atopic Dermatitis. J. Immunol. Res. 2017, 2017, 8493545. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Mitra, N.; Hoffstad, O.J.; Margolis, D.J. Influence of FLG mutations and TSLP polymorphisms on atopic dermatitis onset age. Ann. Allergy Asthma Immunol. 2017, 118, 737–738. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.-J.; Wang, S.-L.; Chen, P.-C.; Guo, Y.L. Prenatal perfluorooctanoic acid exposure and glutathione s-transferase T1/M1 genotypes and their association with atopic dermatitis at 2 years of age. PLoS ONE 2019, 14, e0210708. [Google Scholar] [CrossRef] [PubMed]
- Yoon, N.Y.; Wang, H.Y.; Jun, M.; Jung, M.; Kim, D.H.; Lee, N.R.; Hong, K.-W.; Seo, S.J.; Choi, E.H.; Lee, J.; et al. Simultaneous detection of barrier- and immune-related gene variations in patients with atopic dermatitis by reverse blot hybridization assay. Clin. Exp. Dermatol. 2018, 43, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Andersen, Y.M.F.; Egeberg, A.; Balslev, E.; Jørgensen, C.L.T.; Szecsi, P.B.; Stender, S.; Kaae, J.; Linneberg, A.; Gislason, G.; Skov, L.; et al. Filaggrin loss-of-function mutations, atopic dermatitis and risk of actinic keratosis: Results from two cross-sectional studies. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 1038–1043. [Google Scholar] [CrossRef]
- Asad, S.; Tapia-Páez, I.; Montano Montes, A.; Wahlgren, C.-F.; Bilcha, K.D.; Nordenskjöld, M.; Bradley, M. Evaluation of Single Nucleotide Variants in Ethiopian Patients with Atopic Dermatitis. Acta Derm. Venereol. 2019, 99, 101–102. [Google Scholar] [CrossRef]
- Song, Y.; Schwager, M.J.; Backer, V.; Guo, J.; Porsbjerg, C.; Khoo, S.-K.; Laing, I.A.; Moses, E.K.; LeSouëf, P.; Zhang, G. Environment Changes Genetic Effects on Respiratory Conditions and Allergic Phenotypes. Sci. Rep. 2017, 7, 6342. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.-Y.; Zheng, X.-D.; Fang, L.; Zhou, F.-S.; Sheng, Y.-J.; Wu, Y.-Y.; Yu, C.-X.; Zhu, J.; Xiao, F.-L. A variant on chromosome 2p13.3 is associated with atopic dermatitis in Chinese Han population. Gene 2017, 628, 281–285. [Google Scholar] [CrossRef]
- Can, C.; Yazicioglu, M.; Gurkan, H.; Tozkir, H.; Gorgulu, A.; Sut, N.H. Lack of Association between Toll-like Receptor 2 Polymorphisms (R753Q and A-16934T) and Atopic Dermatitis in Children from Thrace Region of Turkey. Balk. Med. J. 2017, 34, 232–238. [Google Scholar] [CrossRef]
- Chan, A.; Terry, W.; Zhang, H.; Karmaus, W.; Ewart, S.; Holloway, J.W.; Roberts, G.; Kurukulaaratchy, R.; Arshad, S.H. Filaggrin mutations increase allergic airway disease in childhood and adolescence through interactions with eczema and aeroallergen sensitization. Clin. Exp. Allergy 2018, 48, 147–155. [Google Scholar] [CrossRef]
- Chang, J.; Mitra, N.; Hoffstad, O.; Margolis, D.J. Association of Filaggrin Loss of Function and Thymic Stromal Lymphopoietin Variation with Treatment Use in Pediatric Atopic Dermatitis. JAMA Dermatol. 2017, 153, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Debinska, A.; Danielewicz, H.; Drabik-Chamerska, A.; Kalita, D.; Boznanski, A. Filaggrin loss-of-function mutations as a predictor for atopic eczema, allergic sensitization and eczema-associated asthma in Polish children population. Adv. Clin. Exp. Med. 2017, 26, 991–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbert, N.J.; Duijts, L.; den Dekker, H.T.; Jaddoe, V.W.V.; Sonnenschein-van der Voort, A.M.M.; de Jongste, J.C.; Pasmans, S.G.M.A. Role of environmental exposures and filaggrin mutations on associations of ethnic origin with risk of childhood eczema. The Generation R Study. Pediatr. Allergy Immunol. 2016, 27, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-Y.; Tang, J.-P.; Liu, Q.; Zheng, X.-D.; Fang, L.; Yin, X.-Y.; Jiang, X.-Y.; Zhou, F.-S.; Zhu, F.; Liang, B.; et al. Scanning indels in the 5q22.1 region and identification of the TMEM232 susceptibility gene that is associated with atopic dermatitis in the Chinese Han population. Gene 2017, 617, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Ziyab, A.H.; Ewart, S.; Lockett, G.A.; Zhang, H.; Arshad, H.; Holloway, J.W.; Karmaus, W. Expression of the filaggrin gene in umbilical cord blood predicts eczema risk in infancy: A birth cohort study. Clin. Exp. Allergy 2017, 47, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Paternoster, L.; Savenije, O.E.M.; Heron, J.; Evans, D.M.; Vonk, J.M.; Brunekreef, B.; Wijga, A.H.; Henderson, A.J.; Koppelman, G.H.; Brown, S.J. Identification of atopic dermatitis subgroups in children from 2 longitudinal birth cohorts. J. Allergy Clin. Immunol. 2018, 141, 964–971. [Google Scholar] [CrossRef] [Green Version]
- Al-Kzayer, L.F.Y.; Al-Aradi, H.M.H.; Shigemura, T.; Sano, K.; Tanaka, M.; Hamada, M.; Ali, K.H.; Aldaghir, O.M.; Nakazawa, Y.; Okuno, Y. DOCK8 mutation diagnosed using whole-exome sequencing of the dried blood spot-derived DNA: A case report of an Iraqi girl diagnosed in Japan. BMC Med. Genet. 2019, 20, 114. [Google Scholar] [CrossRef]
- Dadi, H.; Jones, T.A.; Merico, D.; Sharfe, N.; Ovadia, A.; Schejter, Y.; Reid, B.; Sun, M.; Vong, L.; Atkinson, A.; et al. Combined immunodeficiency and atopy caused by a dominant negative mutation in caspase activation and recruitment domain family member 11 (CARD11). J. Allergy Clin. Immunol. 2018, 141, 1818–1830. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.A.; Stinson, J.R.; Zhang, Y.; Abbott, J.K.; Weinreich, M.A.; Hauk, P.J.; Reynolds, P.R.; Lyons, J.J.; Nelson, C.G.; Ruffo, E.; et al. Germline hypomorphic CARD11 mutations in severe atopic disease. Nat. Genet. 2017, 49, 1192–1201. [Google Scholar] [CrossRef]
- Peled, A.; Sarig, O.; Sun, G.; Samuelov, L.; Ma, C.A.; Zhang, Y.; Dimaggio, T.; Nelson, C.G.; Stone, K.D.; Freeman, A.F.; et al. Loss-of-function mutations in caspase recruitment domain-containing protein 14 (CARD14) are associated with a severe variant of atopic dermatitis. J. Allergy Clin. Immunol. 2019, 143, 173–181. [Google Scholar] [CrossRef]
- Pigors, M.; Common, J.E.A.; Wong, X.F.C.C.; Malik, S.; Scott, C.A.; Tabarra, N.; Liany, H.; Liu, J.; Limviphuvadh, V.; Maurer-Stroh, S.; et al. Exome Sequencing and Rare Variant Analysis Reveals Multiple Filaggrin Mutations in Bangladeshi Families with Atopic Eczema and Additional Risk Genes. J. Investig. Dermatol. 2018, 138, 2674–2677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Makino, Y.; Nagata, M.; Furuta, J.; Enomoto, H.; Hirota, T.; Tamari, M.; Noguchi, E. A rare variant in CYP27A1 and its association with atopic dermatitis with high serum total IgE. Allergy 2016, 71, 1486–1489. [Google Scholar] [CrossRef]
- Heo, W.I.L.; Park, K.Y.; Jin, T.; Lee, M.-K.; Kim, M.; Choi, E.H.; Kim, H.-S.; Bae, J.M.; Moon, N.J.; Seo, S.J. Identification of novel candidate variants including COL6A6 polymorphisms in early-onset atopic dermatitis using whole-exome sequencing. BMC Med. Genet. 2017, 18, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaca, S.; Civelek, E.; Karaca, M.; Sahiner, U.M.; Ozgul, R.K.; Kocabas, C.N.; Polimanti, R.; Sekerel, B.E. Allergy-specific Phenome-Wide Association Study for Immunogenes in Turkish Children. Sci. Rep. 2016, 6, 33152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margolis, D.J.; Mitra, N.; Gochnauer, H.; Wubbenhorst, B.; D’Andrea, K.; Kraya, A.; Hoffstad, O.; Gupta, J.; Kim, B.; Yan, A.; et al. Uncommon Filaggrin Variants Are Associated with Persistent Atopic Dermatitis in African Americans. J. Investig. Dermatol. 2018, 138, 1501–1506. [Google Scholar] [CrossRef] [Green Version]
- Manz, J.; Rodriguez, E.; ElSharawy, A.; Oesau, E.-M.; Petersen, B.-S.; Baurecht, H.; Mayr, G.; Weber, S.; Harder, J.; Reischl, E.; et al. Targeted Resequencing and Functional Testing Identifies Low-Frequency Missense Variants in the Gene Encoding GARP as Significant Contributors to Atopic Dermatitis Risk. J. Investig. Dermatol. 2016, 136, 2380–2386. [Google Scholar] [CrossRef] [Green Version]
- Park, K.Y.; Park, M.K.; Seok, J.; Li, K.; Seo, S.J. Clinical characteristics of Korean patients with filaggrin-related atopic dermatitis. Clin. Exp. Dermatol. 2016, 41, 595–600. [Google Scholar] [CrossRef]
- Wong, X.F.C.C.C.; Denil, S.L.I.J.I.J.; Foo, J.N.; Chen, H.; Tay, A.S.L.; Haines, R.L.; Tang, M.B.Y.Y.; McLean, W.H.I.I.; Sandilands, A.; Smith, F.J.D.D.; et al. Array-based sequencing of filaggrin gene for comprehensive detection of disease-associated variants. J. Allergy Clin. Immunol. 2018, 141, 814–816. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Alvarez, M.R.; Jiang, W.; Jones, D.C.; Jayaraman, J.; Johnson, C.; Cookson, W.O.; Moffatt, M.F.; Trowsdale, J.; Traherne, J.A. LILRA6 copy number variation correlates with susceptibility to atopic dermatitis. Immunogenetics 2016, 68, 743–747. [Google Scholar] [CrossRef]
- Mathyer, M.E.; Quiggle, A.M.; Wong, X.F.C.C.; Denil, S.L.I.J.; Kumar, M.G.; Ciliberto, H.M.; Bayliss, S.J.; Common, J.E.; de Guzman Strong, C. Tiled array-based sequencing identifies enrichment of loss-of-function variants in the highly homologous filaggrin gene in African-American children with severe atopic dermatitis. Exp. Dermatol. 2018, 27, 989–992. [Google Scholar] [CrossRef]
- Elhaji, Y.; Sasseville, D.; Pratt, M.; Asai, Y.; Matheson, K.; McLean, W.H.I.; Hull, P.R. Filaggrin gene loss-of-function mutations constitute a factor in patients with multiple contact allergies. Contact Dermat. 2019, 80, 354–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.; Liu, Y.; Xue, R.; Chen, L.; Chen, H.; Shao, L.; Wang, J.; Zhang, X. Interleukin 4 -590C/T (rs2243250) Polymorphism Is Associated with Increased Risk of Atopic Dermatitis: Meta-Analysis of Case-Control Studies. Dermat. Contact Atopic Occup. Drug 2017, 28, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Margaritte-Jeannin, P.; Babron, M.-C.; Laprise, C.; Lavielle, N.; Sarnowski, C.; Brossard, M.; Moffatt, M.; Gagne-Ouellet, V.; Etcheto, A.; Lathrop, M.; et al. The COL5A3 and MMP9 genes interact in eczema susceptibility. Clin. Exp. Allergy 2018, 48, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Kong, J.; He, J. Genetic relationship between IL-10 gene polymorphisms and the risk of clinical atopic dermatitis. BMC Med. Genet. 2019, 20, 83. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, H.-C.; Feng, C.; Yan, M. Analysis of the Association of Polymorphisms rs5743708 in TLR2 and rs4986790 in TLR4 with Atopic Dermatitis Risk. Immunol. Investig. 2019, 48, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Chen, Z.-Y.; Li, L.-F. Association between the IL-10-1082G/A, IL-10-592A/C, and IL-10-819G/A Polymorphisms and Atopic Dermatitis Susceptibility: A Meta-Analysis. Genet. Test. Mol. Biomark. 2019, 23, 332–341. [Google Scholar] [CrossRef]
- Arkwright, P.D.; Chase, J.M.; Babbage, S.; Pravica, V.; David, T.J.; Hutchinson, I.V. Atopic dermatitis is associated with a low-producer transforming growth factor beta(1) cytokine genotype. J. Allergy Clin. Immunol. 2001, 108, 281–284. [Google Scholar] [CrossRef]
- Trzeciak, M.; Sakowicz-Burkiewicz, M.; Wesserling, M.; Dobaczewska, D.; Glen, J.; Nowicki, R.; Pawelczyk, T. Expression of Cornified Envelope Proteins in Skin and Its Relationship with Atopic Dermatitis Phenotype. Acta Derm. Venereol. 2017, 97, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Polcari, I.; Becker, L.; Stein, S.L.; Smith, M.S.; Paller, A.S. Filaggrin gene mutations in African Americans with both ichthyosis vulgaris and atopic dermatitis. Pediatr. Dermatol. 2014, 31, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Rebane, A. microRNA and Allergy. In Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2015; Volume 888, pp. 331–352. ISBN 9783319226705. [Google Scholar]
- Dissanayake, E.; Inoue, Y. MicroRNAs in Allergic Disease. Curr. Allergy Asthma Rep. 2016, 16, 67. [Google Scholar] [CrossRef] [PubMed]
- Alaskhar Alhamwe, B.; Khalaila, R.; Wolf, J.; von Bülow, V.; Harb, H.; Alhamdan, F.; Hii, C.S.; Prescott, S.L.; Ferrante, A.; Renz, H.; et al. Histone modifications and their role in epigenetics of atopy and allergic diseases. Allergy Asthma Clin. Immunol. 2018, 14, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamen, L.A.; Schlessinger, J.; Lowell, C.A. Pyk2 Is Required for Neutrophil Degranulation and Host Defense Responses to Bacterial Infection. J. Immunol. 2011, 186, 1656–1665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.R.; Novak, M.J. Beta2 integrin-dependent phosphorylation of protein-tyrosine kinase Pyk2 stimulated by tumor necrosis factor alpha and fMLP in human neutrophils adherent to fibrinogen. FEBS Lett. 1999, 451, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Calabresi, P.A.; Allie, R.; Mullen, K.M.; Yun, S.H.; Georgantas, R.W.; Whartenby, K.A. Kinetics of CCR7 expression differ between primary activation and effector memory states of T(H)1 and T(H)2 cells. J. Neuroimmunol. 2003, 139, 58–65. [Google Scholar] [CrossRef]
- Malaisse, J.; Bourguignon, V.; De Vuyst, E.; Lambert de Rouvroit, C.; Nikkels, A.F.; Flamion, B.; Poumay, Y. Hyaluronan metabolism in human keratinocytes and atopic dermatitis skin is driven by a balance of hyaluronan synthases 1 and 3. J. Investig. Dermatol. 2014, 134, 2174–2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, S.R.; O’Neill, L.A. A trio of microRNAs that control Toll-like receptor signalling. Int. Immunol. 2011, 23, 421–425. [Google Scholar] [CrossRef] [Green Version]
- Sonkoly, E.; Ståhle, M.; Pivarcsi, A. MicroRNAs and immunity: Novel players in the regulation of normal immune function and inflammation. Semin. Cancer Biol. 2008, 18, 131–140. [Google Scholar] [CrossRef]
- Renz, H.; Autenrieth, I.B.; Brandtzæg, P.; Cookson, W.O.; Holgate, S.; von Mutius, E.; Valenta, R.; Haller, D. Gene-environment interaction in chronic disease: A European Science Foundation Forward Look. J. Allergy Clin. Immunol. 2011, 128, S27–S49. [Google Scholar] [CrossRef]
- Von Mutius, E. Gene-environment interactions in asthma. J. Allergy Clin. Immunol. 2009, 123, 3–11. [Google Scholar] [CrossRef]
- Potaczek, D.P.; Harb, H.; Michel, S.; Alhamwe, B.A.; Renz, H.; Tost, J. Epigenetics and allergy: From basic mechanisms to clinical applications. Epigenomics 2017, 9, 539–571. [Google Scholar] [CrossRef]
- Candi, E.; Schmidt, R.; Melino, G. The cornified envelope: A model of cell death in the skin. Nat. Rev. Mol. Cell Biol. 2005, 6, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Irvine, A.D.; McLean, W.H.H.I.; Leung, D.Y.M.M. Filaggrin mutations associated with skin and allergic diseases. N. Engl. J. Med. 2011, 365, 1315–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kypriotou, M.; Huber, M.; Hohl, D. The human epidermal differentiation complex: Cornified envelope precursors, S100 proteins and the “fused genes” family. Exp. Dermatol. 2012, 21, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.J.; McLean, W.H.I. One remarkable molecule: Filaggrin. J. Investig. Dermatol. 2012, 132, 751–762. [Google Scholar] [CrossRef] [Green Version]
- Margolis, D.J.; Apter, A.J.; Gupta, J.; Hoffstad, O.; Papadopoulos, M.; Campbell, L.E.; Sandilands, A.; McLean, W.H.I.; Rebbeck, T.R.; Mitra, N. The persistence of atopic dermatitis and filaggrin (FLG) mutations in a US longitudinal cohort. J. Allergy Clin. Immunol. 2012, 130, 912–917. [Google Scholar] [CrossRef] [Green Version]
- Palmer, C.N.A.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.D.; et al. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef]
- Rogers, A.J.; Celedón, J.C.; Lasky-Su, J.A.; Weiss, S.T.; Raby, B.A. Filaggrin mutations confer susceptibility to atopic dermatitis but not to asthma. J. Allergy Clin. Immunol. 2007, 120, 1332–1337. [Google Scholar] [CrossRef]
- Smith, F.J.D.; Irvine, A.D.; Terron-Kwiatkowski, A.; Sandilands, A.; Campbell, L.E.; Zhao, Y.; Liao, H.; Evans, A.T.; Goudie, D.R.; Lewis-Jones, S.; et al. Loss-of-function mutations in the gene encoding filaggrin cause ichthyosis vulgaris. Nat. Genet. 2006, 38, 337–342. [Google Scholar] [CrossRef]
- Rodríguez, E.; Baurecht, H.; Herberich, E.; Wagenpfeil, S.; Brown, S.J.; Cordell, H.J.; Irvine, A.D.; Weidinger, S. Meta-analysis of filaggrin polymorphisms in eczema and asthma: Robust risk factors in atopic disease. J. Allergy Clin. Immunol. 2009, 123, 1361–1370. [Google Scholar] [CrossRef]
- Van Den Oord, R.A.H.M.; Sheikh, A. Filaggrin gene defects and risk of developing allergic sensitisation and allergic disorders: Systematic review and meta-analysis. BMJ 2009, 339, 86–88. [Google Scholar] [CrossRef] [Green Version]
- Marenholz, I.; Nickel, R.; Rüschendorf, F.; Schulz, F.; Esparza-Gordillo, J.; Kerscher, T.; Grüber, C.; Lau, S.; Worm, M.; Keil, T.; et al. Filaggrin loss-of-function mutations predispose to phenotypes involved in the atopic march. J. Allergy Clin. Immunol. 2006, 118, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Bønnelykke, K.; Pipper, C.B.; Tavendale, R.; Palmer, C.N.A.; Bisgaard, H. Filaggrin gene variants and atopic diseases in early childhood assessed longitudinally from birth. Pediatr. Allergy Immunol. 2010, 21, 954–961. [Google Scholar] [CrossRef]
- Weidinger, S.; O’Sullivan, M.; Illig, T.; Baurecht, H.; Depner, M.; Rodriguez, E.; Ruether, A.; Klopp, N.; Vogelberg, C.; Weiland, S.K.; et al. Filaggrin mutations, atopic eczema, hay fever, and asthma in children. J. Allergy Clin. Immunol. 2008, 121, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, S.; Rodríguez, E.; Stahl, C.; Wagenpfeil, S.; Klopp, N.; Illig, T.; Novak, N. Filaggrin mutations strongly predispose to early-onset and extrinsic atopic dermatitis. J. Investig. Dermatol. 2007, 127, 724–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, J.; Northstone, K.; Lee, S.P.; Liao, H.; Zhao, Y.; Pembrey, M.; Mukhopadhyay, S.; Smith, G.D.; Palmer, C.N.A.; McLean, W.H.I.; et al. The burden of disease associated with filaggrin mutations: A population-based, longitudinal birth cohort study. J. Allergy Clin. Immunol. 2008, 121, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Marenholz, I.; Kerscher, T.; Bauerfeind, A.; Esparza-Gordillo, J.; Nickel, R.; Keil, T.; Lau, S.; Rohde, K.; Wahn, U.; Lee, Y.A. An interaction between filaggrin mutations and early food sensitization improves the prediction of childhood asthma. J. Allergy Clin. Immunol. 2009, 123, 911–916. [Google Scholar] [CrossRef]
- Elias, P.M. Therapeutic Implications of a Barrier-based Pathogenesis of Atopic Dermatitis. Ann. Dermatol. 2010, 22, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Loo, E.X.L.; Shek, L.P.; Goh, A.; Teoh, O.H.; Chan, Y.H.; Soh, S.E.; Saw, S.M.; Kwek, K.; Gluckman, P.D.; Godfrey, K.M.; et al. Atopic Dermatitis in Early Life: Evidence for at Least Three Phenotypes? Results from the GUSTO Study. Int. Arch. Allergy Immunol. 2015, 166, 273–279. [Google Scholar] [CrossRef]
- Greisenegger, E.; Novak, N.; Maintz, L.; Bieber, T.; Zimprich, F.; Haubenberger, D.; Gleiss, A.; Stingl, G.; Kopp, T.; Zimprich, A. Analysis of four prevalent filaggrin mutations (R501X, 2282del4, R2447X and S3247X) in Austrian and German patients with atopic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 607–610. [Google Scholar] [CrossRef]
- Rupnik, H.; Rijavec, M.; Korošec, P. Filaggrin loss-of-function mutations are not associated with atopic dermatitis that develops in late childhood or adulthood. Br. J. Dermatol. 2015, 172, 455–461. [Google Scholar] [CrossRef]
- Flohr, C.; Johansson, S.G.O.; Wahlgren, C.-F.; Williams, H. How atopic is atopic dermatitis? J. Allergy Clin. Immunol. 2004, 114, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.J.; Sandilands, A.; Zhao, Y.; Liao, H.; Relton, C.L.; Meggitt, S.J.; Trembath, R.C.; Barker, J.N.W.N.; Reynolds, N.J.; Cordell, H.J.; et al. Prevalent and Low-Frequency Null Mutations in the Filaggrin Gene Are Associated with Early-Onset and Persistent Atopic Eczema. J. Investig. Dermatol. 2008, 128, 1591–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandilands, A.; Terron-Kwiatkowski, A.; Hull, P.R.; O’Regan, G.M.; Clayton, T.H.; Watson, R.M.; Carrick, T.; Evans, A.T.; Liao, H.; Zhao, Y.; et al. Comprehensive analysis of the gene encoding filaggrin uncovers prevalent and rare mutations in ichthyosis vulgaris and atopic eczema. Nat. Genet. 2007, 39, 650–654. [Google Scholar] [CrossRef] [PubMed]
- Barker, J.N.W.N.; Palmer, C.N.A.; Zhao, Y.; Liao, H.; Hull, P.R.; Lee, S.P.; Allen, M.H.; Meggitt, S.J.; Reynolds, N.J.; Trembath, R.C.; et al. Null mutations in the filaggrin gene (FLG) determine major susceptibility to early-onset atopic dermatitis that persists into adulthood. J. Investig. Dermatol. 2007, 127, 564–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stemmler, S.; Parwez, Q.; Petrasch-Parwez, E.; Epplen, J.T.; Hoffjan, S. Two common loss-of-function mutations within the filaggrin gene predispose for early onset of atopic dermatitis. J. Investig. Dermatol. 2007, 127, 722–724. [Google Scholar] [CrossRef] [PubMed]
- Ercan, H.; Ispir, T.; Kirac, D.; Baris, S.; Ozen, A.; Oztezcan, S.; Cengizlier, M.R. Predictors of atopic dermatitis phenotypes and severity: Roles of serum immunoglobulins and filaggrin gene mutation R501X. Allergol. Immunopathol. (Madr) 2013, 41, 86–93. [Google Scholar] [CrossRef]
- Weidinger, S.; Illig, T.; Baurecht, H.; Irvine, A.D.; Rodriguez, E.; Diaz-Lacava, A.; Klopp, N.; Wagenpfeil, S.; Zhao, Y.; Liao, H.; et al. Loss-of-function variations within the filaggrin gene predispose for atopic dermatitis with allergic sensitizations. J. Allergy Clin. Immunol. 2006, 118, 214–219. [Google Scholar] [CrossRef]
- Brunner, P.M.; Guttman-Yassky, E. Racial differences in atopic dermatitis. Ann. Allergy Asthma Immunol. 2019, 122, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, B.P.; Guttman-Yassky, E.; Alexis, A.F. Atopic dermatitis in diverse racial and ethnic groups-Variations in epidemiology, genetics, clinical presentation and treatment. Exp. Dermatol. 2018, 27, 340–357. [Google Scholar] [CrossRef] [Green Version]
- Margolis, D.J.; Gupta, J.; Apter, A.J.; Hoffstad, O.; Papadopoulos, M.; Rebbeck, T.R.; Wubbenhorst, B.; Mitra, N. Exome sequencing of filaggrin and related genes in African-American children with atopic dermatitis. J. Investig. Dermatol. 2014, 134, 2272–2274. [Google Scholar] [CrossRef] [Green Version]
- Shaw, T.E.; Currie, G.P.; Koudelka, C.W.; Simpson, E.L. Eczema Prevalence in the United States: Data from the 2003 National Survey of Children’s Health. J. Investig. Dermatol. 2011, 131, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanovic, N.; Flohr, C.; Irvine, A.D. The exposome in atopic dermatitis. Allergy 2020, 75, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Si, N.; Liu, Y.; Zhang, D.; Wang, R.; Zhang, Y.; Wang, S.; Liu, X.; Deng, X.; Ma, Y.; et al. Steroid sulfatase and filaggrin mutations in a boy with severe ichthyosis, elevated serum IgE level and moyamoya syndrome. Gene 2017, 628, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Imamura, Y.; Scott, I.C.; Greenspan, D.S. The pro-alpha3(V) collagen chain. Complete primary structure, expression domains in adult and developing tissues, and comparison to the structures and expression domains of the other types V and XI procollagen chains. J. Biol. Chem. 2000, 275, 8749–8759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, J.; Holden, P.; Hansen, U. The expanded collagen VI family: New chains and new questions. Connect. Tissue Res. 2013, 54, 345–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeBert, D.C.; Squirrell, J.M.; Rindy, J.; Broadbridge, E.; Lui, Y.; Zakrzewska, A.; Eliceiri, K.W.; Meijer, A.H.; Huttenlocher, A. Matrix metalloproteinase 9 modulates collagen matrices and wound repair. Development 2015, 142, 2136–2146. [Google Scholar] [CrossRef] [Green Version]
- Soldi, R.; Mitola, S.; Strasly, M.; Defilippi, P.; Tarone, G.; Bussolino, F. Role of alphavbeta3 integrin in the activation of vascular endothelial growth factor receptor-2. EMBO J. 1999, 18, 882–892. [Google Scholar] [CrossRef] [Green Version]
- Maegdefessel, L.; Azuma, J.; Toh, R.; Merk, D.R.; Deng, A.; Chin, J.T.; Raaz, U.; Schoelmerich, A.M.; Raiesdana, A.; Leeper, N.J.; et al. Inhibition of microRNA-29b reduces murine abdominal aortic aneurysm development. J. Clin. Investig. 2012, 122, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Salazar, K.D.; Lankford, S.M.; Brody, A.R. Mesenchymal stem cells produce Wnt isoforms and TGF-beta1 that mediate proliferation and procollagen expression by lung fibroblasts. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 297, L1002–L1011. [Google Scholar] [CrossRef] [Green Version]
- Rørvig, S.; Østergaard, O.; Heegaard, N.H.H.; Borregaard, N. Proteome profiling of human neutrophil granule subsets, secretory vesicles, and cell membrane: Correlation with transcriptome profiling of neutrophil precursors. J. Leukoc. Biol. 2013, 94, 711–721. [Google Scholar] [CrossRef]
- Foley, S.C.; Mogas, A.K.; Olivenstein, R.; Fiset, P.O.; Chakir, J.; Bourbeau, J.; Ernst, P.; Lemière, C.; Martin, J.G.; Hamid, Q. Increased expression of ADAM33 and ADAM8 with disease progression in asthma. J. Allergy Clin. Immunol. 2007, 119, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Huls, A.; Klumper, C.; MacIntyre, E.A.; Brauer, M.; Melen, E.; Bauer, M.; Berdel, D.; Bergstrom, A.; Brunekreef, B.; Chan-Yeung, M.; et al. Atopic dermatitis: Interaction between genetic variants of GSTP1, TNF, TLR2, and TLR4 and air pollution in early life. Pediatr. Allergy Immunol. 2018, 29, 596–605. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Hirota, T.; Obara, K.; Shimizu, M.; Jodo, A.; Kameda, M.; Doi, S.; Fujita, K.; Shirakawa, T.; Enomoto, T.; et al. An association study of asthma and related phenotypes with polymorphisms in negative regulator molecules of the TLR signaling pathway. J. Hum. Genet. 2006, 51, 284–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kormann, M.S.D.; Depner, M.; Hartl, D.; Klopp, N.; Illig, T.; Adamski, J.; Vogelberg, C.; Weiland, S.K.; von Mutius, E.; Kabesch, M. Toll-like receptor heterodimer variants protect from childhood asthma. J. Allergy Clin. Immunol. 2008, 122, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.-Y.; Schumann, R.R.; Hamann, L.; Neumann, K.; Worm, M.; Heine, G. Association of the toll-like receptor 2 A-16934T promoter polymorphism with severe atopic dermatitis. Allergy 2009, 64, 1608–1615. [Google Scholar] [CrossRef]
- Potaczek, D.; Nastalek, M.; Okumura, K.; Wojas-Pelc, A.; Undas, A.; Nishiyama, C. An association of TLR2-16934A > T polymorphism and severity/phenotype of atopic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 715–721. [Google Scholar] [CrossRef]
- Li, H.-F.; Yan, L.-P.; Wang, K.; Li, X.-T.; Liu, H.-X.; Tan, W. Association between ADAM33 polymorphisms and asthma risk: A systematic review and meta-analysis. Respir. Res. 2019, 20, 38. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Yan, F.; Yang, Z.; Zhou, J.; Chen, Y.; Ding, Z. Association between ADAM33 S2 and V4 polymorphisms and susceptibility to allergic rhinitis: A meta-analysis. Allergol. Immunopathol. 2016, 44, 170–176. [Google Scholar] [CrossRef]
- Bedsaul, J.R.; Carter, N.M.; Deibel, K.E.; Hutcherson, S.M.; Jones, T.A.; Wang, Z.; Yang, C.; Yang, Y.-K.; Pomerantz, J.L. Mechanisms of Regulated and Dysregulated CARD11 Signaling in Adaptive Immunity and Disease. Front. Immunol. 2018, 9, 2105. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Chertov, O.; Bykovskaia, S.N.; Chen, Q.; Buffo, M.J.; Shogan, J.; Anderson, M.; Schröder, J.M.; Wang, J.M.; Howard, O.M.; et al. Beta-defensins: Linking innate and adaptive immunity through dendritic and T cell CCR6. Science 1999, 286, 525–528. [Google Scholar] [CrossRef]
- Jurevic, R.J.; Bai, M.; Chadwick, R.B.; White, T.C.; Dale, B.A. Single-nucleotide polymorphisms (SNPs) in human beta-defensin 1: High-throughput SNP assays and association with Candida carriage in type I diabetics and nondiabetic controls. J. Clin. Microbiol. 2003, 41, 90–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertin, J.; Wang, L.; Guo, Y.; Jacobson, M.D.; Poyet, J.L.; Srinivasula, S.M.; Merriam, S.; DiStefano, P.S.; Alnemri, E.S. CARD11 and CARD14 are novel caspase recruitment domain (CARD)/membrane-associated guanylate kinase (MAGUK) family members that interact with BCL10 and activate NF-kappa B. J. Biol. Chem. 2001, 276, 11877–11882. [Google Scholar] [CrossRef]
- Guo, X.; Zhang, Y.; Wang, P.; Li, T.; Fu, W.; Mo, X.; Shi, T.; Zhang, Z.; Chen, Y.; Ma, D.; et al. VSTM1-v2, a novel soluble glycoprotein, promotes the differentiation and activation of Th17 cells. Cell. Immunol. 2012, 278, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Hirayasu, K.; Arase, H. Leukocyte Immunoglobulin-Like Receptor (LILR). In Encyclopedia of Signaling Molecules; Springer International Publishing: Cham, Switzerland, 2018; pp. 2854–2861. [Google Scholar]
- Su, H.C.; Jing, H.; Angelus, P.; Freeman, A.F. Insights into immunity from clinical and basic science studies of DOCK8 immunodeficiency syndrome. Immunol. Rev. 2019, 287, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Minkiewicz, J.; de Rivero Vaccari, J.P.; Keane, R.W. Human astrocytes express a novel NLRP2 inflammasome. Glia 2013, 61, 1113–1121. [Google Scholar] [CrossRef]
- Tschopp, J.; Martinon, F.; Burns, K. NALPs: A novel protein family involved in inflammation. Nat. Rev. Mol. Cell Biol. 2003, 4, 95–104. [Google Scholar] [CrossRef]
- Walne, A.J.; Vulliamy, T.; Kirwan, M.; Plagnol, V.; Dokal, I. Constitutional mutations in RTEL1 cause severe dyskeratosis congenita. Am. J. Hum. Genet. 2013, 92, 448–453. [Google Scholar] [CrossRef] [Green Version]
- Gray, P.W.; Aggarwal, B.B.; Benton, C.V.; Bringman, T.S.; Henzel, W.J.; Jarrett, J.A.; Leung, D.W.; Moffat, B.; Ng, P.; Svedersky, L.P. Cloning and expression of cDNA for human lymphotoxin, a lymphokine with tumour necrosis activity. Nature 1984, 312, 721–724. [Google Scholar] [CrossRef]
- Trzeciak, M.; Wesserling, M.; Bandurski, T.; Glen, J.; Nowicki, R.; Pawelczyk, T. Association of a Single Nucleotide Polymorphism in a Late Cornified Envelope-like Proline-rich 1 Gene (LELP1) with Atopic Dermatitis. Acta Derm. Venereol. 2016, 96, 459–463. [Google Scholar] [CrossRef] [Green Version]
- Knüppel, S.; Esparza-Gordillo, J.; Marenholz, I.; Holzhütter, H.-G.; Bauerfeind, A.; Ruether, A.; Weidinger, S.; Lee, Y.-A.; Rohde, K. Multi-locus stepwise regression: A haplotype-based algorithm for finding genetic associations applied to atopic dermatitis. BMC Med. Genet. 2012, 13, 8. [Google Scholar] [CrossRef] [Green Version]
- Margolis, D.J.; Gupta, J.; Apter, A.J.; Ganguly, T.; Hoffstad, O.; Papadopoulos, M.; Rebbeck, T.R.; Mitra, N. Filaggrin-2 variation is associated with more persistent atopic dermatitis in African American subjects. J. Allergy Clin. Immunol. 2014, 133, 784–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Xiao, L.; Jia, J.; Li, F.; Wang, X.; Duan, Q.; Jing, H.; Yang, P.; Chen, C.; Wang, Q.; et al. Cornulin Is Induced in Psoriasis Lesions and Promotes Keratinocyte Proliferation via Phosphoinositide 3-Kinase/Akt Pathways. J. Investig. Dermatol. 2019, 139, 71–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turksen, K.; Troy, T.-C. Permeability barrier dysfunction in transgenic mice overexpressing claudin 6. Development 2002, 129, 1775–1784. [Google Scholar] [PubMed]
- Yamakoshi, T.; Makino, T.; Ur Rehman, M.; Yoshihisa, Y.; Sugimori, M.; Shimizu, T. Trichohyalin-like 1 protein, a member of fused S100 proteins, is expressed in normal and pathologic human skin. Biochem. Biophys. Res. Commun. 2013, 432, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Kantor, R.; Silverberg, J.I. Environmental risk factors and their role in the management of atopic dermatitis. Expert Rev. Clin. Immunol. 2017, 13, 15–26. [Google Scholar] [CrossRef]
- Isidoro-García, M.; Dávila-González, I.; Pascual de Pedro, M.; Sanz-Lozano, C.; Lorente-Toledano, F. Interactions between genes and the environment. Epigenetics in allergy. Allergol. Immunopathol. 2007, 35, 254–258. [Google Scholar] [CrossRef] [Green Version]
- Makeyev, E.V.; Maniatis, T. Multilevel Regulation of Gene Expression by MicroRNAs. Science 2008, 319, 1789–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Maes, T.; Cobos, F.A.; Schleich, F.; Sorbello, V.; Henket, M.; De Preter, K.; Bracke, K.R.; Conickx, G.; Mesnil, C.; Vandesompele, J.; et al. Asthma inflammatory phenotypes show differential microRNA expression in sputum. J. Allergy Clin. Immunol. 2016, 137, 1433–1446. [Google Scholar] [CrossRef] [Green Version]
- Sinha, A.; Yadav, A.K.; Chakraborty, S.; Kabra, S.K.; Lodha, R.; Kumar, M.; Kulshreshtha, A.; Sethi, T.; Pandey, R.; Malik, G.; et al. Exosome-enclosed microRNAs in exhaled breath hold potential for biomarker discovery in patients with pulmonary diseases. J. Allergy Clin. Immunol. 2013, 132, 219–222. [Google Scholar] [CrossRef]
- Bauer, S.M. Atopic Eczema: Genetic Associations and Potential Links to Developmental Exposures. Int. J. Toxicol. 2017, 36, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Rebane, A.; Runnel, T.; Aab, A.; Maslovskaja, J.; Rückert, B.; Zimmermann, M.; Plaas, M.; Kärner, J.; Treis, A.; Pihlap, M.; et al. MicroRNA-146a alleviates chronic skin inflammation in atopic dermatitis through suppression of innate immune responses in keratinocytes. J. Allergy Clin. Immunol. 2014, 134, 836–847. [Google Scholar] [CrossRef] [PubMed]
- Deaton, A.M.; Bird, A. CpG islands and the regulation of transcription. Genes Dev. 2011, 25, 1010–1022. [Google Scholar] [CrossRef] [Green Version]
- Takai, D.; Jones, P.A. Comprehensive analysis of CpG islands in human chromosomes 21 and 22. Proc. Natl. Acad. Sci. USA 2002, 99, 3740–3745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schübeler, D. Function and information content of DNA methylation. Nature 2015, 517, 321–326. [Google Scholar] [CrossRef]
- Nestor, C.E.; Barrenäs, F.; Wang, H.; Lentini, A.; Zhang, H.; Bruhn, S.; Jörnsten, R.; Langston, M.A.; Rogers, G.; Gustafsson, M.; et al. DNA Methylation Changes Separate Allergic Patients from Healthy Controls and May Reflect Altered CD4+ T-Cell Population Structure. PLoS Genet. 2014, 10, e1004059. [Google Scholar] [CrossRef] [Green Version]
- Pascual, M.; Suzuki, M.; Isidoro-Garcia, M.; Padrón, J.; Turner, T.; Lorente, F.; Dávila, I.; Greally, J.M. Epigenetic changes in B lymphocytes associated with house dust mite allergic asthma. Epigenetics 2011, 6, 1131–1137. [Google Scholar] [CrossRef]
- Joubert, B.R.; Felix, J.F.; Yousefi, P.; Bakulski, K.M.; Just, A.C.; Breton, C.; Reese, S.E.; Markunas, C.A.; Richmond, R.C.; Xu, C.-J.J.; et al. DNA Methylation in Newborns and Maternal Smoking in Pregnancy: Genome-wide Consortium Meta-analysis. Am. J. Hum. Genet. 2016, 98, 680–696. [Google Scholar] [CrossRef] [Green Version]
- Joehanes, R.; Just, A.C.; Marioni, R.E.; Pilling, L.C.; Reynolds, L.M.; Mandaviya, P.R.; Guan, W.; Xu, T.; Elks, C.E.; Aslibekyan, S.; et al. Epigenetic Signatures of Cigarette Smoking. Circ. Cardiovasc. Genet. 2016, 9, 436–447. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Chang, C.; Lu, Q. The Genetics and Epigenetics of Atopic Dermatitis-Filaggrin and Other Polymorphisms. Clin. Rev. Allergy Immunol. 2016, 51, 315–328. [Google Scholar] [CrossRef]
- Fontalba, A.; Gutierrez, O.; Fernandez-Luna, J.L. NLRP2, an inhibitor of the NF-kappaB pathway, is transcriptionally activated by NF-kappaB and exhibits a nonfunctional allelic variant. J. Immunol. 2007, 179, 8519–8524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruey, J.M.; Bruey-Sedano, N.; Newman, R.; Chandler, S.; Stehlik, C.; Reed, J.C. PAN1/NALP2/PYPAF2, an inducible inflammatory mediator that regulates NF-kappaB and caspase-1 activation in macrophages. J. Biol. Chem. 2004, 279, 51897–51907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steevels, T.A.M.; van Avondt, K.; Westerlaken, G.H.A.; Stalpers, F.; Walk, J.; Bont, L.; Coffer, P.J.; Meyaard, L. Signal inhibitory receptor on leukocytes-1 (SIRL-1) negatively regulates the oxidative burst in human phagocytes. Eur. J. Immunol. 2013, 43, 1297–1308. [Google Scholar] [CrossRef] [PubMed]
- Steevels, T.A.M.; Lebbink, R.J.; Westerlaken, G.H.A.; Coffer, P.J.; Meyaard, L. Signal inhibitory receptor on leukocytes-1 is a novel functional inhibitory immune receptor expressed on human phagocytes. J. Immunol. 2010, 184, 4741–4748. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Study Type | Population/Country | Objective | Sample Size | Genes | SNP/Mutation | Results/Conclusion |
|---|---|---|---|---|---|---|---|
| [77] | WES (Whole-exome sequencing) | Iraq | To determine DOCK8 deficiency | 1 child | DOCK8 | c.3332delT, | Mutation present in hyperimmunoglobulin E syndrome (HIES) and non-Hodgkin lymphoma patient. |
| Phe1113Leufs *2 (rs140392509) | |||||||
| [65] | Candidate gene | Denmark | To determine association of atopic dermatitis (AD) with ichthyosis vulgaris (IV) and actinic keratosis (AK) | 481 AK patients, 9112 Healthy controls (HC) | FLG | 1537C>T(R501X) (rs61816761) | FLG homozygous loss of function and AK (in 0.8% of AK studied vs. 0.2% in control population) |
| 2318_2321del (2282del4) | |||||||
| 7375C>T (R2447X; rs138726443) | |||||||
| [66] | Candidate gene | Ethiopia | To elucidate SNVs associated with AD | 184 patients of AD and 186 HC | SPINK5 | rs2303063; | Significant association with AD. |
| rs2303067 | |||||||
| [35] | Candidate gene, WES | Ethiopia | To elucidate SNVs associated with AD To establish the role of CLDN1 variants in Ethiopian AD patients | 22 patients for WES; 159 AD patients and 192 HC for genotyping | CLDN1 | rs17501010 | rs893051 is associated with development of AD in early life. |
| rs9290927 | |||||||
| rs9290929 | |||||||
| rs893051 | |||||||
| [46] | Candidate gene | Jordan | To study the association between resistin gene polymorphisms and AD | 162 AD patients, 161 HC | RETN | SNP +299 G>A (rs3745367) | rs3745367 associated with AD in a gender- and age-specific manner (male, less than 10 y) |
| SNP +157 C>T (rs3219177) | |||||||
| [57] | Candidate gene | Iran | To identify association of SNPs in IL-10 and TGF-β1 and AD in Iranian patients | 89 children with AD, 138 HC | TGF-β1 | cdn 10 | cdn10/C allele, CC genotype associated with AD |
| cdn 25 | cdn 25/C allele associated with AD | ||||||
| [68] | Candidate gene | Chinese Han | To identify variants in Chinese Han population associated to AD | 4619 AD patients and 10789 HC | CD207/VAX2 | rs112111458 (allele G/A) | Association of rs112111458 and AD |
| [69] | Candidate gene | Turkish children | To evaluate if some TLR2 gene polymorphisms are associated with AD | 70 children with AD, 69 HC | TLR2 | rs5743708 (R753Q) | None |
| rs4696480 (A-16934T) | |||||||
| [70] | Candidate gene | Isle of Wight | To study the association of FLG loss of function with atopic march | 1150 participants of the Isle of Wight birth cohort | FLG | R501X (rs61816761) | FLG loss of function mutations are associated with early life eczema at age 1, 2 and 4 years and was consistently associated with rhinitis from 4 years onwards |
| 2282del | |||||||
| S3247X (rs150597413) | |||||||
| [71] | Candidate genes | USA (mixed population of children) | To determine whether variations in FLG and TSLP genotype corresponded to differences in therapeutic treatment use over time | 842 children with AD | FLG | R501X (rs61816761) | Variations in FLG and TSLP genotype were associated with differences in self-reported skin clearance, TCI usage, and steroid usage |
| 2282del4 | |||||||
| R2447X (rs138726443) | |||||||
| S3247X (rs150597413) | |||||||
| TSLP | rs1898671 | ||||||
| 2282del4 | |||||||
| [78] | WES | Canada | To identify the genetic aberration in 4 related patients with combined immunodeficiency, early-onset asthma, eczema, and food allergies, as well as autoimmunity | 4 related patients | CARD11 | hg19:chr7:2987341:G>A NM_032415:exon3:c.C88T:p.R30W (rs145474800) | CARD11- R30W is associated with recurrent infections, autoimmunity, and severe atopy. The novel R30W mutations described abrogate the NF-κB pathway and lead to decreased IL-2 and IFN-γ secretion and lymphocyte proliferation |
| [72] | Candidate gene | Poland | To investigate the importance of 4 common FLG null mutations in the susceptibility to eczema in Polish children population | 50 children with AD, 37 children with non-atopic eczema and 71 HC children | FLG | R501X (rs61816761) | FLG null mutations and AD are associated but explain only a part of AD cases (13.8%) |
| 2282del4 | |||||||
| R2447X (rs138726443) | |||||||
| S3247X (rs150597413) | |||||||
| [73] | Candidate gene | Different ethnic origins (Dutch, Cape Verdean, Dutch Antillean, Moroccan, Surinamese-Creole, Surinamese-Hindustani, Turkish children) | To study the association of known genetic factors and ethnic origin with the development of eczema | 3096 children | FLG | 2282del4 | Carrier frequencies of FLG mutations in children of non-Dutch origins were low. |
| R2447X (rs138726443) | |||||||
| R501X (rs61816761) | |||||||
| S3247X (rs150597413) | |||||||
| [91] | Meta-analysis | Canada (Caucasian) | Study the effect of FLG mutations on contact dermatitits (CD) | 165 patients with CD, 891 HC | FLG | 2282del4 | Association between FLG loss of function mutations and contact polysensitivity, especially in R501X polymorphism. |
| R2447X (rs138726443) | |||||||
| S3247X (rs150597413) | |||||||
| R501X (rs61816761) | |||||||
| [13] | GWAS (Genome-wide association study) | European ancestry | To identify shared risk variants of a broad allergic disease phenotype that considers the presence of asthma, hay fever and eczema | 180,129 cases with asthma and/or hay fever and/or eczema, and 180,709 HC | FLG | R501X (rs61816761) | This SNP is a stronger risk factor for eczema than for hay fever or asthma. |
| RPTN-[ ]-HRNR | rs12123821 | This SNP is a stronger risk factor for eczema than for hay fever or asthma. | |||||
| IL1R2-[ ]-IL18R1 | rs12470864 | This SNP is a stronger risk factor for eczema than for hay fever or asthma. | |||||
| WDR36-[ ]-CAMK4 | rs6594499 | This SNP is a stronger risk factor for hay fever than for eczema or asthma. | |||||
| IL2RA | rs61839660 | This SNP is a stronger risk factor for eczema than for hay fever or asthma. | |||||
| GSDMB | rs921650 | This SNP is a stronger risk factor for eczema than for hay fever or asthma. | |||||
| [36] | Candidate gene | Russia | To explore the frequency FLG mutations and CNVs in AD patients and control subjects of Russian and Tatar ethnic origin living in Volga-Ural region of Russia | 177 Russian, 126 Tatar AD patients; and 152 Russian, 109 Tatar HC | FLG | 2282del4 R501X (rs61816761) R2447X (rs138726443) | Significant differences in 2282del4 frequency were found between Tatar AD patients and HC. The allelic frequency of the R501X mutation in AD patients was 0.85% and in HC -0.47%. The allelic frequency of R2447X was 1.75% in patients, and 1.33% in HC. |
| [37] | Candidate gene | India | To investigate the personal consequences of having atopic dermatitis and/or hand eczema and FLG mutations | 163 patients and 86 HC | FLG | S2889X (rs782477344) | Mutations in S2889X constituted 96.4% of all FLG mutations. No carrier of R501X and Q2417X mutations was identified. FLG mutations are associated with irritant contact dermatitis with or without atopy, allergic contact dermatitis without atopy, and idiopathic subtypes. FLG mutations were associated with more severe hand eczema. |
| 2282del4 | |||||||
| R501X (rs61816761) | |||||||
| Q2417X (rs528722713) | |||||||
| 2282del4 | |||||||
| R2447X (rs138726443) | |||||||
| [83] | WES | Korea | To identify family-specific candidate genetic variants associated with early-onset AD in Koreans. | 3 families (2 affected AD and 2 unaffected individuals) for WES. 112 AD and 61 HC for validation studies. | COL6A6 | rs16830494 | COL6A6 variants may be risk factors for AD because the minor allele (AA) in rs16830494 and the rs59021909 (TT) allele and the rs200963433 heterozygous (CT) frequency were all higher in AD cases compared to controls, but no significant association was reached. |
| rs59021909 | |||||||
| rs200963433 | |||||||
| [38] | Candidate gene | Chinese Han | To study in the Chinese Ham population the association AD with previously reported SNPs | 3013 AD patients, 5483 HC | TLR1-TLR6 | rs2101521 | SNPs rs2158177 and rs1837253 are associated with AD in Chinese Han population |
| WDR36-CAMK4 | rs1438673 | ||||||
| PTGER4 | rs7720838 | ||||||
| NFATC2 | rs6021270 | ||||||
| IL1RL1-IL18R1 | rs3771175 | ||||||
| THSD7B | rs1469621 | ||||||
| RAD50/IL13 | rs2158177 | ||||||
| TSLP | rs1837253 | ||||||
| [39] | Candidate gene | Sweden | To explore the longitudinal relation between preschool eczema, FLG mutation, or both and IgE sensitization in childhood. | 1890 children | FLG | 2282del4 | Preschool eczema is associated with IgE sensitization to both food allergens and aeroallergens up to 16 years of age. FLG mutation is associated with IgE sensitization to peanut but not to other allergens. Sensitized children with preceding PSE are more often polysensitized. |
| R501X (rs61816761) | |||||||
| R2447X (rs138726443) | |||||||
| [40] | Candidate gene | USA | To elucidate the associations between KIF3A SNPs and asthma, eczema, and AR, alone and in combination. | 7000 children and 1020 HC. Results were replicated in 762 children with atopy. | KIF3A | rs9784600 | KIF3A is associated with asthma + eczema. The presence of AR comorbidity did not increase the genetic association of KIF3A with asthma or even with asthma+ eczema. |
| rs9784675 | |||||||
| rs11740584 | |||||||
| rs7737031 | |||||||
| rs17691077 | |||||||
| rs2299007 | |||||||
| rs3798130 | |||||||
| rs12186803 | |||||||
| rs1468216 | |||||||
| rs2023822 | |||||||
| rs2237059 | |||||||
| rs2023823 | |||||||
| [84] | Phenome-WAS (Phenome-wide association study) | Turkey | To dissect the role of immunogenetics in allergy and asthma. | 974 children | ADAM33 | rs2787094 | rs2280090 was associated with reduced MEF240s (i.e., the ratio of mean expiratory flow after 240 s of hypertonic saline inhalation with respect to the age- and ancestry-matched reference value) and with an increased risk of allergic bronchitis; rs3918396 was associated with wheezing and eczema comorbidity. |
| rs543749 | |||||||
| rs2280090 | |||||||
| rs2280091 | |||||||
| rs3918396 | |||||||
| rs6127096 | |||||||
| rs511898 | |||||||
| rs2280090 | |||||||
| rs3918396 | |||||||
| IL4 | rs2243250 | rs2243250 is associated with increased FEV240 (forced expiratory flow volume after 240 s of hypertonic saline inhalation). | |||||
| rs2070874 | |||||||
| CD14 | rs2569190 | Associated with asthma. | |||||
| ADRB2 | rs1042713 | No association | |||||
| rs1042714 | |||||||
| IL13 | rs1800925 | No association | |||||
| rs1295686 | |||||||
| rs20541 | |||||||
| IL4R | rs1805015 | No association | |||||
| rs1801275 | |||||||
| MS4A2 | rs1441586 | No association | |||||
| rs569108 | |||||||
| SERPINE1 | rs1799768 | No association | |||||
| TNF | rs1800629 | No association | |||||
| [42] | Candidate gene | Korean | To study MIF promoter polymorphisms and total plasma IgE in AD Korean patients | 178 AD patients, 80 HC | MIF | rs755622 (−173 G to C) | MIF promoter polymorphisms in the −173 C allele and the MIF C/5-CATT and MIF C/7-CATT haplotypes were significantly associated with an increased risk for AD. |
| [41] | Candidate gene | Korea | To identify FLG SNP variations and evaluated its association with clinical phenotypes, including AD and other parameters. | 81 patients | FLG | rs71626704 | rs71626704 and rs76413899 were significantly associated with a history of asthma and cheilitis. rs62623409 and rs71625199 were associated with sensitization to environmental allergens. |
| rs76413899 | |||||||
| rs62623409 | |||||||
| rs71625199 | |||||||
| [43] | Candidate gene | Korea | To investigate the association between four possible TSLP polymorphisms and atopic disease in a Korean population. | - | TSLP | rs2289276 | rs3806932, rs2289276, and rs2289278 are associated with susceptibility of AD. rs3806932, rs3806933, and rs2289276 form one linkage disequilibrium block. The GTT haplotype strongly contributes to atopic march. |
| rs2289278 | |||||||
| rs3806932 | |||||||
| rs3806933 | |||||||
| [44] | Candidate gene | UK | 224 patients and 40 HC | FLG | 2282del4 | Subjects with FLG-null mutations have more mature Langerhans cells in non-lesional skin irrespective of whether they have AD. | |
| R501X (rs61816761) | |||||||
| S3247X (rs150597413) | |||||||
| R2447X (rs138726443) | |||||||
| [45] | Candidate gene | Chinese Han/ Singapore, Chinese Han/ Shanghai, Chinese Han/Shanxi, Korean, Japanese/ Kyushu and Japanese/ mainland | To assess the significance of FLG mutations as clinical biomarkers in East Asian populations. | 1384 patients and 1031 HC | FLG | 3321delA | c.3321delA was found in all populations. Some mutations showed south-to-north (or north-to-south) distribution gradient: p.K4022X, the most prevalent FLG mutation in northern China and Korea, declined in frequency moving southward; in contrast, c.6950del8 (e.g., p.Q2417X, p.E2422X) showed the reverse. p.S2554X/p.S2889X/p.S3296X/Q1701X mutations were Japanese-specific. |
| K4022X (rs146466242) | |||||||
| 6950del8 | |||||||
| Q2417X (rs528722713) | |||||||
| E2422X (rs374588791) | |||||||
| S2554X (rs121909626) | |||||||
| S2889X (rs782477344) | |||||||
| S3296 (rs760426769) | |||||||
| Q1701X (rs4547271) | |||||||
| [92] | Meta-analysis | China, Taiwan, Japan and Saudi Arabia (Asian population) and Poland, Czech R., Macedonia, Egypt (Caucasian) | To study the association between IL-4-590C/T polymorphism and AD susceptibility. | 923 patients and 1215 HC | IL-4 | -590C/T | The IL-4 -590C/T polymorphism may contribute to AD susceptibility in the overall population and children, especially for Asian children. |
| [89] | CNV analysis | UK | To assess the contribution of LILR and LILRA3 genes CNV to AD | 1482 patients from 378 families | LILR, LILRA3 | The transmission of one copy of LILRA6 within families was potentially related to the development of AD. | |
| [47] | Candidate gene | Finland | To test the association of the 4 most prevalent European FLG null mutations, the 2 Finnish enriched FLG null mutations, the FLG 12-repeat allele, and 50 additional epidermal barrier gene variants, with risk of AD, disease severity, clinical features, risk of other atopic diseases, age of onset, and treatment response. | 501 patients with AD and 1710 HC | FLG | R501X (rs61816761) | 2822del4 was significantly associated with early-onset AD and asthma. R501X was associated with early-onset and suggestively with keratosis pilaris. R2447X showed suggestive association with early-onset AD. Baseline IgE values were higher in patients with FLG null mutations, but the association was not significant; FLG null mutations were not associated with atopic hand eczema, dermographism, or HSV infections. |
| R2447X (rs138726443) | |||||||
| S3247X (rs150597413) | |||||||
| S1020X (rs200360684) | |||||||
| V603M (rs2306942) | |||||||
| rs12730241 | |||||||
| CLDN1 | No significant association with AD | ||||||
| CLDN4 | |||||||
| CLDN20 | |||||||
| CLDN23 | |||||||
| OCLN | |||||||
| IVL | |||||||
| FLG2 | |||||||
| LOR | |||||||
| JAM-1 | |||||||
| TJP1 | |||||||
| [79] | WES | USA | To study CARD11 mutations in four families with recalcitrant, severe atopic disease. | 8 patients | CARD11 | L194P | The study describes rare hypomorphic dominant negative mutations in CARD11 in 4 unrelated families, which lead to dominantly inherited, severe atopy, with variable infection beyond the skin. |
| R975W (rs1064795307) | |||||||
| E57D | |||||||
| dup183_196 | |||||||
| [5] | GWAS | UK | To test whether genetically lowered vitamin D levels were associated with risk of asthma, atopic dermatitis, or elevated serum IgE levels. | 33996 children | DHCR7 | rs12785878 | No association |
| CYP2R1 | rs10741657 | ||||||
| CYP24A1 | rs6013897 | ||||||
| [48] | Candidate gene | Italy | To evaluate the role of FLG polymorphisms expression and risk of developing a concomitant Molluscum contagiosum sustained skin infection in the pediatric population with AD. | 100 children with AD and 97 healthy children | FLG | rs79808464 | FLG mutations are associated with early onset of AD, more severe clinical course of disease, and a significantly increased risk of M. contagiosum sustained skin infection |
| rs116222149 | |||||||
| rs11584340 | |||||||
| rs113136594 | |||||||
| rs145828067 | |||||||
| rs374910442 | |||||||
| rs747005144 | |||||||
| rs145627745 | |||||||
| rs144209313 | |||||||
| rs74129443 | |||||||
| rs192455877 | |||||||
| rs150957860 | |||||||
| rs138055273 | |||||||
| rs147472105 | |||||||
| rs183942200 | |||||||
| rs558269137 | |||||||
| [86] | NGS (Next-generation sequencing) | German | To identify disease association in the locus 11q13.5 using combination of sequencing and functional annotation. | 31 AD patients | LRRC32 | A407T | Association of low-frequency and rare missense mutations within the LRRC32 gene with AD. |
| R518W (rs142940671) | |||||||
| R312 | |||||||
| S411R (rs201424816) | |||||||
| R414W (rs201431152) | |||||||
| R652C | |||||||
| [93] | Meta-analysis | French, French- Canadian and UK | To detect new interacting genes involved in eczema | 388 French families. Replication in 253 French-Canadian and 207 UK family datasets. | COL5A3 | rs2287807 | Identified significant interaction between two new genes, COL5A3 and MMP9, which may be accounted for by a degradation of COL5A3 by MMP9 influencing eczema susceptibility. |
| MMP9 | rs17576 | ||||||
| [85] | NGS | USA | To evaluate FLG LoF variation in children of African ancestry and the association with AD and AD persistence. | 262 African American children and 133 Caucasians | FLG | R501X (rs61816761) | Rare FLG LoF variants in African American children are associated with AD and more persistent AD. In contrast to Europeans, no FLG LoF variants predominate in African American children. The most common variants were R501X, S3316X, and R826X. |
| S3316X (rs149484917) | |||||||
| R826X (rs115746363) | |||||||
| R2447X (rs138726443) | |||||||
| Q570X (rs192402912) | |||||||
| R3409X (rs201356558) | |||||||
| S3247X (rs150597413) | |||||||
| Q3818X (rs148606936) | |||||||
| H440fs | |||||||
| [90] | CNV (Copy number variations) analysis | African American (USA) | To study FLG LoF and CNV in African American population | 39 children with AD | FLG | R501X (rs61816761) | rs149484917 is a population-specific FLG LoF unique to several populations of African Ancestry. Two new FLG LoF were identified (488delG and S3101X) |
| R826X (rs115746363) | |||||||
| S3316X (rs149484917) | |||||||
| 488delG | |||||||
| S3101X | |||||||
| [49] | Candidate gene | Japan | To study polymorphisms of SPINK5 gene in Japanese AD patients | 57 patients, 50 HC | SPINK5 | Q267R (rs6892205) | Only S368N frequency differed between Japanese patients and data from Human Genetic Variation Database. Algorithms predicting functional effects of amino acids substitution showed significant scores for R654H |
| A335V (rs34482796) | |||||||
| S368N (rs230306) | |||||||
| D386N (rs2303064) | |||||||
| R711Q (rs3777134) | |||||||
| E825D (rs2303070) | |||||||
| [50] | Candidate gene | Korea | To examine the spectrum of null-mutations and compare with other Asian countries | 70 patients | FLG | R501X (rs61816761) | Only 11 AD patients had FLG mutations. This frequency was lower than that described for other Asian populations (Chinese, Japanese, Singaporean) |
| 3321delA | |||||||
| Y1767X (rs1222103354) | |||||||
| S1695X (rs772851618) | |||||||
| Q1701X (rs145738429) | |||||||
| Q1745X (rs1209640261) | |||||||
| Q1790X (rs200622741) | |||||||
| S2554X (rs121909626) | |||||||
| S2889X | |||||||
| S3296X (rs761212672) | |||||||
| K4022X (rs146466242) | |||||||
| 3222del4 | |||||||
| S1515X (rs180768115) | |||||||
| Q2417X (rs528722713) | |||||||
| [51] | Candidate gene | Korea | To investigate the genetic polymorphisms of FLG in Korean AD patients | 9 ichtyosis vulgaris patients 50 AD patients 55 HC | FLG | K4022X (rs146466242) | This loss-of-function mutation was only found in AD patients. 62 new SNPs were identified |
| [87] | NGS | Korea | To investigate clinical characteristics of AD patients with FLG mutations. To determine differences between patients with and without FLG mutations | 1110 patients, 68 with mutations in FLG gene | FLG | K4022X (rs146466242) | Null alleles were associated with early onset of AD and higher risk of developing the disease by age 2 years. |
| 3321delA | EASI score was also higher in these patients | ||||||
| [76] | GWAS | UK, Netherlands | To investigate longitudinal phenotypes of AD in two independent cohorts | UK: 9894 individuals (ALSPAC) NL: 3652 individuals (PIAMA) | INPP5D | rs1057258-c | Six classes based on temporal trajectories of rash were identified: persistent, early-onset/late resolving, early-onset/early-resolving, medium-onset/resolved, late-onset/resolved. FLG null mutations were strongly associated with early-onset and late-onset of AD (p < 0.00001: ALSPAC) and early-onset/late-resolving (p = 0.0006: PIAMA) |
| PRR5L | rs12295535-t | ||||||
| STAT3 | rs17881320-t | ||||||
| PPP2R3D | rs2143950-t | ||||||
| ACTL9 | rs2164983-a | ||||||
| IL6R | rs2228145-c | ||||||
| KIF3A | rs2897442-c | ||||||
| OVOL1 | rs479844-g | ||||||
| C11orf30 | rs7927894-t | ||||||
| IL22 | rs2227482-t | ||||||
| IL21 | rs17389644-a | ||||||
| IL2RA | rs6602364-g | ||||||
| [80] | WES | USA (Hispanic, Caucasian, African-American) | To identify rare DNA variants conferring significant risk for AD | 3 patients | CARD14 | c.1778T>C, I593T | Downregulation of CARD14 led to severe AD and reduced skin protection against infection and dysregulated cutaneous inflammation pathways |
| c.2206A>C,N737H (rs535171797) | |||||||
| [81] | WES, rare enrichment analysis | Bangladeshi | To analyze the genetic architecture of patients with AD from a Bangladeshi community in London, UK | 53 cases and 42 HC from 70 families | SCAND3 | Some rare sequence variations of candidate genes have been identified. FLG loss-of-function variations were carried by almost 50% of AD-affected individuals. | |
| TCHHL1 | |||||||
| ADCY10 | |||||||
| MTF1 | |||||||
| MCM10 | |||||||
| ORM2 | |||||||
| CUX2 | |||||||
| MAST2 | |||||||
| PHLDB1 | |||||||
| FLG | |||||||
| [52] | Candidate gene | Poland | To explore the role of different SNPs at 11q13.5 in predisposing to allergic phenotypes | 270 AD patients, 540 HC | rs7927894 | The haplotype TATG in these SNPs fully explained the association with AD (p = 0.00021)The TG haplotype in the last two SNPs was also related to allergic rhinitis | |
| rs2513517 | |||||||
| rs7930794 | |||||||
| rs7125552 | |||||||
| [94] | Meta-analysis | Asian, Caucasian | To assess the genetic relationship between Il-10 polymorphisms and susceptibility to AD. | 16 case-control studies | IL-10 | IL-10 -1082a/G | These polymorphisms showed a weak association with AD susceptibility |
| IL-10 -819T/C | |||||||
| IL-10 -592a/C | |||||||
| [53] | Candidate gene | Netherlands (Caucasians, Asians, Afro- Caribbean) | To investigate whether FLG mutations influence the outcome of immunosuppressive therapy | 42 patients with severe AD: 3 Asians, 1 Afro-Caribbean, 38 Caucasians | FLG | R501X (rs61816761) | FLG mutation group showed a trend towards less improvement in the course of 24 weeks of treatment with methotrexate and azothiopine |
| 2282del4 | |||||||
| R2447X (rs138726443) | |||||||
| S3247X (rs150597413) | |||||||
| 5321delA | |||||||
| [54] | Candidate gene | Japan | To elucidate the effect of bi-allelic FLG mutation on AD incidence and severity | 6 individuals from 3 families | FLG | Q1790X+S3296X | The most severe AD was associated with c.3321delA+S2889X bi-allelic combination. By contrast, individuals with S2889X+S3296X did not develop AD |
| Q1790X+S2889X | |||||||
| S2889X+S3296X | |||||||
| Q1701X+S2889X | |||||||
| 3321delA+S2889X | |||||||
| [67] | Candidate gene | Inuit | To study the effect of environment on genotype-phenotype association in a genetically homogeneous population, living in two separate areas | 615 Greenlandic Inuit individuals 643 Danish Inuit individuals | ADAM | rs612709 | LTα rs2844484 was associated with AD in Greenland patients (p = 0.035) The risk of AD was related to the genotype distribution of this SNP, with a significant interaction with the place of residence |
| rs528557 | |||||||
| rs44707 | |||||||
| rs2787094 | |||||||
| ALOX5 | rs4986832 | ||||||
| rs892690 | |||||||
| rs2115819 | |||||||
| LT-α | rs2844484 | ||||||
| rs909253 | |||||||
| rs1041981 | |||||||
| LTC4S | rs730012 | ||||||
| NOS I | rs7977109 | ||||||
| ORMDL3 | rs4065275 | ||||||
| rs12603332 | |||||||
| TBXA2R | rs4523 | ||||||
| TNF-α | rs1799964 | ||||||
| rs1800630 | |||||||
| rs1800629 | |||||||
| [82] | WES | Japan | To investigate rare genetic variants associated with AD | 469 AD patients, 935 HC | APOB | rs145862664 | Gene polymorphism in CYP27A1, a gene involved in vitamin D3 metabolism, was related to AD |
| CYP27A1 | rs199691576 | ||||||
| C3orf15 | rs193146105 | ||||||
| GAK | rs142107211 | ||||||
| VNN2 | rs200230703 | ||||||
| USP35 | rs200193128 | ||||||
| ZNF749 | rs76428401 | ||||||
| [55] | Candidate gene | China | To investigate the potential role of SHARPIN in the pathogenesis of AD | 65 AD patients, 100 HC | SHARPIN | g.480G>A | SNPs g.4320, g.4334, g.4343, g.4344, g.5363 were present both in patients and controls. The mutations in SHARPIN were only present in AD patients, decreasing the expression in AD lesions. |
| g.4576A>T | |||||||
| g.5070C>T | |||||||
| rs200698932 | |||||||
| rs769935596 | |||||||
| rs760937092 | |||||||
| rs1280524235 | |||||||
| g.4320 | |||||||
| g.4334 | |||||||
| g.4343 | |||||||
| g.4344 | |||||||
| g.5363 | |||||||
| [56] | Candidate gene | Japanese, Korean | To determine prevalence of FLG mutation in AD and IV patients in Japan and South Korea | Japan: 26 IV patients, 91 AD patients South Korea: 76 AD patients | FLG | R501X (rs61816761) | Mutation S3296X only appeared in Japanese AD patients. R501X and R826X only appeared in IV patientsThe rest of the mutations were found in both Korean and Japanese patients. |
| S2554X (rs121909626) | |||||||
| S2889X | |||||||
| G1109EfsX (rs133912394) | |||||||
| K4022X (rs146466242) | |||||||
| S1695X (rs772851618) | |||||||
| Q1701X (rs145738429) | |||||||
| R826X (rs115746363) | |||||||
| [58] | Candidate gene | Denmark | To examine the association between loss-of-function mutations in FLG and AD and asthma in adult twins | 575 adults twins with asthma | FLG | R501X (rs61816761) | Within the dizygotic twin population, 11 pairs were discordant for FLG mutation. The risk of AD increased in the twin with FLG mutation. No significant association was found with FLG mutations and asthma |
| 2282del4 | |||||||
| R2447X (rs138726443) | |||||||
| [59] | Candidate gene | Denmark | To explore heritable, environmental, and clinical factors related to persistent AD | 417 children: 186 patients (40 with persistent AD), 231 HC. Follow-up study from birth to age 13 y | FLG | R501X (rs61816761) | 29% of patients with persistent AD had FLG mutations. The higher AD genetic risk score, the higher risk of persistent AD. |
| 2282del4 | |||||||
| R2447X (rs138726443) | |||||||
| S3247X (rs150597413) | |||||||
| FLG2 | |||||||
| SPRR3 | |||||||
| [60] | Candidate gene | Poland | To identify new potential markers of AD | 159 AD patients, 108 HC | RPTN | rs284544 | In FLG WT patients, RPTN rs3001978CC was significantly associated with AD early age onset (p = 0.033), pruritus (p = 0.021), severity of AD (p = 0.045) and concomitant asthma (p = 0.041) rs 941934 allele A was more frequent in AD patients (p = 0.007), the homozygous AA only appeared in AD patients (p = 0.019). The association depended on FLG mutations. |
| rs28441202 | |||||||
| rs3001978 | |||||||
| rs12117644 | |||||||
| CRNN | rs941934 | ||||||
| FLG | R2447X (rs138726443) | ||||||
| S3247X (rs150597413) | |||||||
| [61] | Candidate gene | Russia | To determine the relationship of TLRs polymorphisms with AD | 25 AD patients, 25 AD and rhinitis/asthma patients, 100 HC | TLR2 | rs55743708 (G>A) | Increased levels of IL-4 and IL-10. Dysfunction of cell activation. |
| TLR4 | rs4986790(A>G) | Increased levels of IL-4 and IL-1. Weaker cell response to microbial antigens. | |||||
| [62] | Candidate gene | USA | To examine the effect of FLG mutations and TSLP (thymic stromal lymphopoietin) polymorphisms on the age of AD onset | 822 children (age 2–17 y) | FLG | R501X (rs61816761) | FLG null mutations were associated with early onset of AD. Number of mutations was associated with timing of onset. No association was found between TSLP polymorphism and timing of onset. |
| 2282del4 | |||||||
| R2447X (rs138726443) | |||||||
| S3247X (rs150597413) | |||||||
| TSLP | rs1898671 | ||||||
| [63] | Candidate gene | Taiwan | To investigate the association between gene–environmental interaction and childhood AD | 839 mother–child pairs | GST | GST-T1/M1 mutants | GST null genotypes in association with high levels of perfluoroalkyl substances in blood increased the risk of developing AD |
| [88] | NGS | Singapore population: Chinese, Indian, Malay | To sequence the entire FLG coding region in Singaporeans from different ethnicities | 334 patients with AD and/or IV | FLG | S1515X (rs180768115) | A new technology that improved accuracy and cost-effectiveness is described. New mutations have been identified |
| E2422X (rs374588791) | |||||||
| S406X (rs189114758) | |||||||
| c.6950-6957del8 | |||||||
| c.1640delG | |||||||
| Q368X (rs746899204) | |||||||
| 3321delA | |||||||
| 7945delA | |||||||
| Q2417X (rs528722713) | |||||||
| 2952delC | |||||||
| 9040-9058dup19 | |||||||
| Q1790X (rs200622741) | |||||||
| S1302X (rs754812742) | |||||||
| S1515X (rs180768115) | |||||||
| 4004del2 | |||||||
| 2282del4 | |||||||
| R2447X (rs138726443) | |||||||
| 477insA | |||||||
| 678delA | |||||||
| S378X (rs755134998) | |||||||
| 3036delT | |||||||
| 10866delA | |||||||
| rs10067777 | |||||||
| [74] | GWAS | China | To identify AD susceptibility genes in 5q22.1 and observe expression in AD tissues | 3031 cases, 5075 HC | TMEM232 | rs7701890 | rs11357450 had the strongest association with risk of AD (OR = 1.20; p = 0.04) |
| rs13360927 | |||||||
| rs13361382 | |||||||
| rs5870408 | |||||||
| rs1400764268 | |||||||
| rs35639206 | |||||||
| rs137936676 | |||||||
| rs10617471 | |||||||
| rs11357450 | |||||||
| [64] | Candidate gene | Korea | To identify mutations and SNPs in barrier- or immune-related genes | 279 AD patients, 224 HC | KLK7 | KLK7 MT | More frequent in AD patients (p = 0.04). No differences between AD groups |
| FLG | 3321delA | More frequent in moderate/sever AD patients (p = 0.038) | |||||
| K4022X (rs14646429) | More frequent in patients but not statistically different | ||||||
| SPINK5 | 1156 | More frequent in AD vs HC (p < 0.001). No differences between AD groups | |||||
| 1188 | |||||||
| 2475 | |||||||
| DEFB1 | rs5743399 | More frequent in AD vs HC (p < 0.002). No differences between AD groups | |||||
| KDR | rs2305948 | More frequent in AD vs HC (p = 0.03). No differences between AD groups | |||||
| IL5RA | rs334809 | More frequent in AD vs HC (p < 0.001). More frequent in mild AD vs moderate/severe | |||||
| IL9 | rs31563 | More frequent in AD vs HC (p < 0.001). No differences between mild AD and HC; and between moderate AD vs. HC | |||||
| IL12RB1 | rs393548 | More frequent in AD vs HC (p = 0.02). | |||||
| rs436857 | More frequent in AD vs HC (p = 0.01). More frequent in mild AD vs. HC. No differences between moderate AD and HC | ||||||
| IL13 | rs20541 | Heterozygous less frequent in patients vs. HC | |||||
| [95] | Meta-analysis | Germany, Turkey, Italy, Finland, Ukraine, Russia | To assess whether TLRs polymorphisms are associate with risk of AD | TLR2: 9 studies; 733 cases, 807 HC | TLR2 | rs5743708 | Increased risk of AD for GA heterozygous |
| TLR4: 6 studies; 646 cases, 601 HC | TLR4 | rs4986790 | AG showed some correlation with risk of AD, but was non-conclusive. | ||||
| [96] | Meta-analysis | Saudi Arabia, Iran, India, Netherlands, Italy, Poland, Macedonia, Czech Republic, China, Korea, Taiwan, Germany, UK | To summarize the current evidence on association between IL-10 polymorphisms and susceptibility to AD | 15 case-control studies, 1647 patients, 2031 HC | IL-10 | IL-10-1092 G/A | Increased risk of AD associated with IL-10-819 G/A mutation in Caucasian subjects and with IL-10-1092 G/A in Asian patients |
| IL-10-592 A/C | |||||||
| IL-10-819 G/A | |||||||
| [75] | GWAS | UK | To assess whether FLG expression in umbilical cord blood associates with and predicts AD | 94 infants | FLG | S3247X (rs150597413) | FLG expression in cord blood correlated with AD risk. 2.94-increased risk for mutated FLG variants |
| 2282del4 |
| Cell/Ttissue Types | Epigenetic Assay | Significant Findings | Reference |
|---|---|---|---|
| Primary adult human keratocytes | miFinder miRNA PCR Array | Broad dysregulation of miRNAs upon IL-4 treatment | [27] |
| Whole blood samples | DNA methylation profiling | Identifying CpG methylation sites in IL4 and IL13 associated with AD phenotype | [28] |
| Serum | Real-time PCR for miR-146a | miR-146a levels are unaltered in AD patients | [29] |
| Serum | RNA sequencing | miR-151a and miR-409 overexpressed in Chinese AD patients | [30] |
| AD lesioned skin | Microarray expression data from GSE32924 | Regulatory network comprising 182 miRNAS | [24] |
| Umbilical cord serum | Exiqon Serum/Plasma Focus micro- RNA PCR Panel (179 miRNAs) | miR-144 levels are higher in the umbilical cord of AD children | [26] |
| Whole blood samples | DNA methylation profiling | Association between smoking and the methylation state of PITPNM2 | [13] |
| Monocytes and neutrophils | VSTM1 methylation | Polymorphism dependent VSTM1 methylation | [31] |
| AD lesioned skin | Microarray miRNA expression (GSE31408) | Hsa-let-7a-5p, has-miR-26a-5p and has-miR-143-3p differentially expressed in lesioned tissues | [32] |
| Whole blood samples | NLRP2 methylation | NLRP2 methylation is associated both to the environment and SNP | [33] |
| AD lesioned skin | Real-time PCR assay | miR-124 downregulated in AD lesional tissue | [25] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin, M.J.; Estravís, M.; García-Sánchez, A.; Dávila, I.; Isidoro-García, M.; Sanz, C. Genetics and Epigenetics of Atopic Dermatitis: An Updated Systematic Review. Genes 2020, 11, 442. https://doi.org/10.3390/genes11040442
Martin MJ, Estravís M, García-Sánchez A, Dávila I, Isidoro-García M, Sanz C. Genetics and Epigenetics of Atopic Dermatitis: An Updated Systematic Review. Genes. 2020; 11(4):442. https://doi.org/10.3390/genes11040442
Chicago/Turabian StyleMartin, Maria J, Miguel Estravís, Asunción García-Sánchez, Ignacio Dávila, María Isidoro-García, and Catalina Sanz. 2020. "Genetics and Epigenetics of Atopic Dermatitis: An Updated Systematic Review" Genes 11, no. 4: 442. https://doi.org/10.3390/genes11040442
APA StyleMartin, M. J., Estravís, M., García-Sánchez, A., Dávila, I., Isidoro-García, M., & Sanz, C. (2020). Genetics and Epigenetics of Atopic Dermatitis: An Updated Systematic Review. Genes, 11(4), 442. https://doi.org/10.3390/genes11040442