VviWRKY40, a WRKY Transcription Factor, Regulates Glycosylated Monoterpenoid Production by VviGT14 in Grape Berry

 ,
, 

Abstract
:1. Introduction
2. Methods and Materials
2.1. Plant Materials and ABA Treatments
2.2. Physicochemical Analysis
2.3. Cloning of the VviGT14 Promoter and VviWRKY40 Coding Sequence (CDS)
2.4. RNA Extraction and Quantitative Real-Time PCR Analysis
2.5. DNA Pull-Down Assay
2.6. Yeast One-Hybrid Assay
2.7. Electrophoretic Mobility Shift Assay
2.8. VviGT14 Promoter Activity Assay
2.9. Transient Overexpression of VviWRKY40 in Tobacco Leaves
2.10. Subcellular Localization of VviWRKY40
2.11. Dual Luciferase Assay
2.12. Extraction and Determination of Monoterpenoids
2.13. Extraction and Determination of ABA
2.14. Accession Numbers
3. Results
3.1. Characterization of the VviGT14 Promoter Region
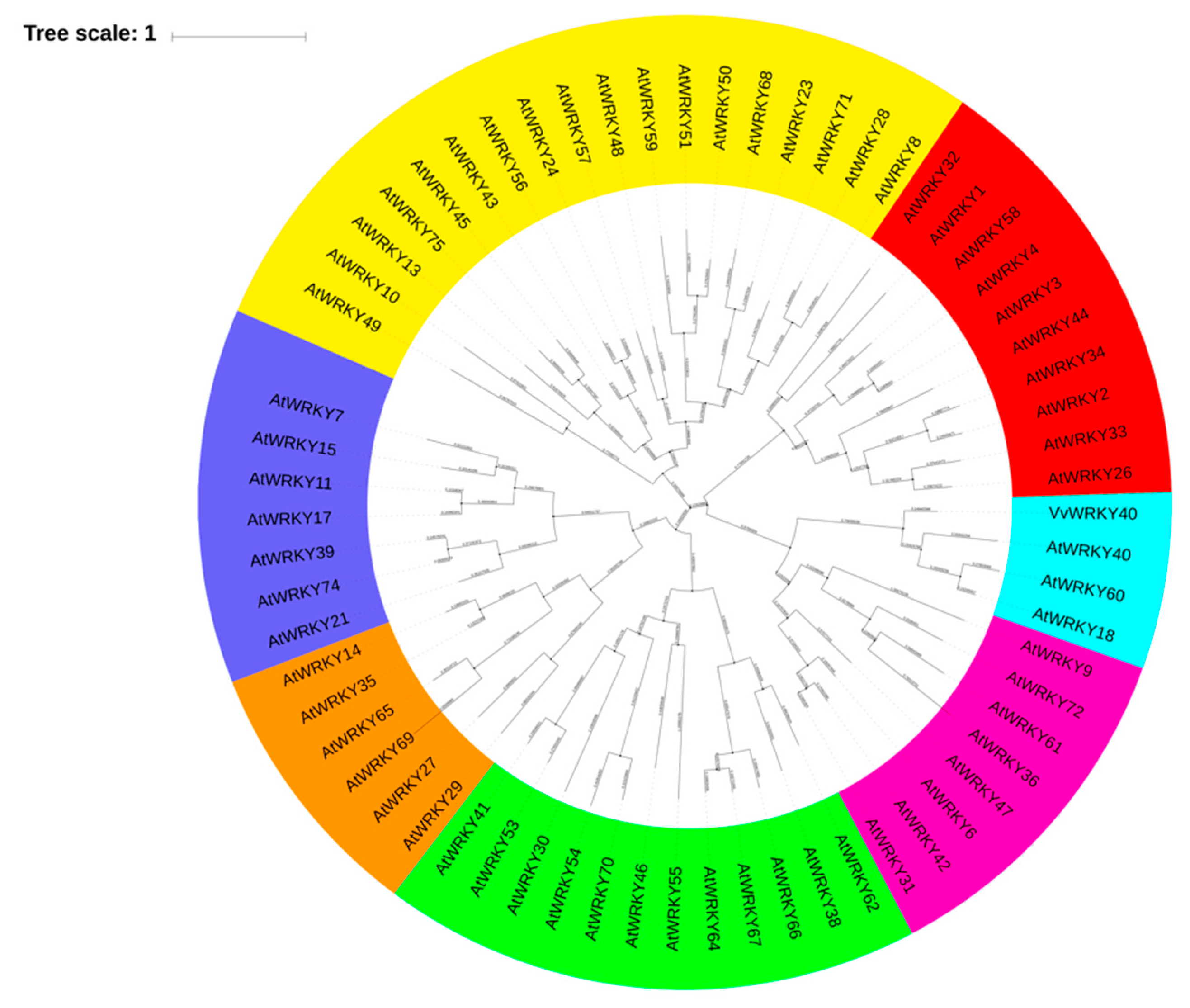
3.2. Screening of TFs Binding to the VviGT14 Promoter
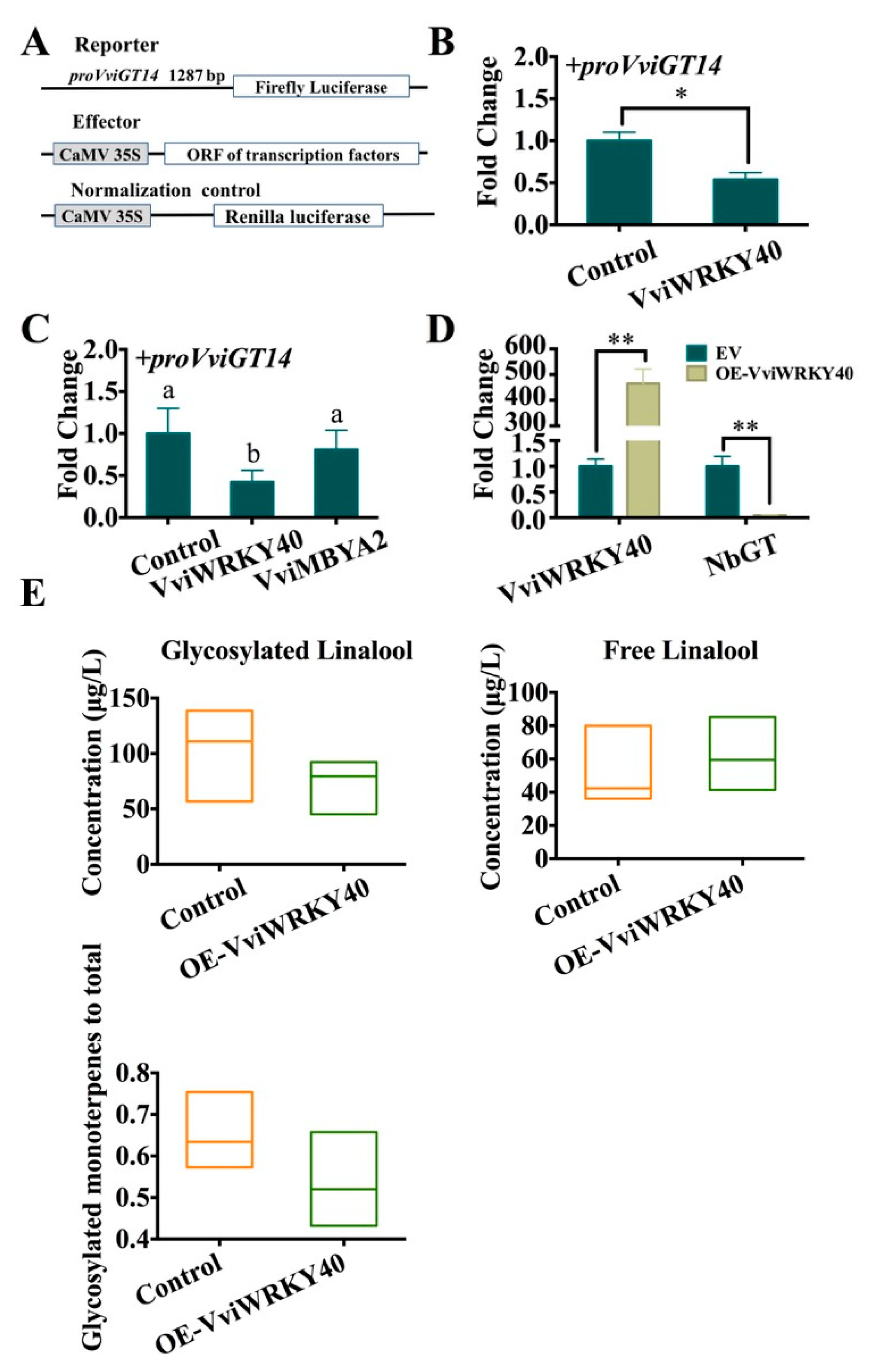
3.3. Validation of VviWRKY40 Interaction with the VviGT14 Promoter
3.4. VviWRKY40 Localizing in the Cell Nucleus and Lacking Transcriptional Activation Function
3.5. Negative Regulation of VviWRKY40 on VviGT14
3.6. Negative Correlation between VviWRKY40 Expression and VviGT14 Expression during Grape Berry Ripening
3.7. ABA Downregulates VviWRKY40 and Upregulates VviGT14 in Grapevine Cell Suspension
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Strauss, C.R.; Wilson, B.; Gooley, P.R.; Williams, P.J. Role of monoterpenes in grape and wine flavor. In Biogeneration of Aromas; ACS Publications: Washington, DC, USA, 1986. [Google Scholar]
- Bonisch, F.; Frotscher, J.; Stanitzek, S.; Ruhl, E.; Wust, M.; Bitz, O.; Schwab, W. Activity-based profiling of a physiologic aglycone library reveals sugar acceptor promiscuity of family 1 UDP-glucosyltransferases from grape. Plant Physiol. 2014, 166, 23–39. [Google Scholar] [CrossRef] [Green Version]
- Schieberle, P. Primary odorants of pale lager beer. Eur. Food Res. Technol. 1991, 193, 558–565. [Google Scholar] [CrossRef]
- Grimaldi, A.; Bartowsky, E.; Jiranek, V.; Schieberle, P. A survey of glycosidase activities of commercial wine strains of Oenococcus oeni. Int. J. Food Microbiol. 2005, 105, 233–234. [Google Scholar] [CrossRef]
- Gunata, Y.Z.; Bayonove, C.L.; Baumes, R.L.; Cordonnier, R.E. Stability of free and bound fractions of some aroma components of grapes cv. Muscat during the wine processing: Preliminary results. Am. J. Enol. Vitic. 1986, 37, 112–114. [Google Scholar]
- Tholl, D. Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Curr. Opin. Plant Biol. 2006, 9, 297–304. [Google Scholar] [CrossRef]
- Lichtenthaler, H. The 1 deoxy D xylulose 5 phosphate pathway of isoprenoid biosynthesis in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 47–65. [Google Scholar] [CrossRef]
- Muhlemann, J.K.; Klempien, A.; Dudareva, N. Floral volatiles: From biosynthesis to function. Plant Cell Environ. 2014, 37, 1936–1949. [Google Scholar] [CrossRef]
- Lim, E.K. Plant glycosyltransferases: Their potential as novel biocatalysts. Chem. Eur. J. 2005, 11, 5486–5494. [Google Scholar] [CrossRef]
- Ross, J.; Li, Y.; Lim, E.-K. Higher plant glycosyltransferases. Genome Biol. 2001, 2, 3001–3006. [Google Scholar] [CrossRef]
- Zhu, B.Q.; Cai, J.; Wang, Z.Q.; Xu, X.Q.; Duan, C.Q.; Pan, Q.H. Identification of a plastid-localized bifunctional nerolidol/linalool synthase in relation to linalool biosynthesis in young grape berries. Int. J. Mol. Sci. 2014, 15, 21992–22010. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.M.; Aubourg, S.P.; Schouwey, M.B.; Daviet, L.; Schalk, M.; Toub, O.; Lund, S.T.; Bohlmann, J. Functional annotation, genome organization and phylogeny of the grapevine (Vitis vinifera) terpene synthase gene family based on genome assembly, FLcDNA cloning, and enzyme assays. BMC Plant Biol. 2010, 10, 226. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.M.; Bohlmann, J. Identification of Vitis vinifera (-)-alpha-terpineol synthase by in silico screening of full-length cDNA ESTs and functional characterization of recombinant terpene synthase. Phytochemistry 2004, 65, 1223–1229. [Google Scholar] [CrossRef]
- Lücker, J.; Bowen, P.; Bohlmann, J. Vitis vinifera terpenoid cyclases: Functional identification of two sesquiterpene synthase cDNAs encoding (+)-valencene synthase and (-)-germacrene D synthase and expression of mono- and sesquiterpene synthases in grapevine flowers and berries. Phytochemistry 2004, 65, 2649–2659. [Google Scholar] [CrossRef]
- Bonisch, F.; Frotscher, J.; Stanitzek, S.; Ruhl, E.; Wust, M.; Bitz, O.; Schwab, W. A UDP-Glucose: Monoterpenol glucosyltransferase adds to the chemical diversity of the grapevine metabolome. Plant Physiol. 2014, 165, 561–581. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Y.; Wen, Y.Q.; Meng, N.; Qian, X.; Pan, Q.H. Monoterpenyl glycosyltransferases differentially contribute to production of monoterpenyl glycosides in two aromatic vitis vinifera varieties. Front. Plant Sci. 2017, 8, 1226. [Google Scholar] [CrossRef]
- Wen, Y.Q.; Zhong, G.Y.; Gao, Y.; Lan, Y.B.; Duan, C.Q.; Pan, Q.H. Using the combined analysis of transcripts and metabolites to propose key genes for differential terpene accumulation across two regions. BMC Plant Biol. 2015, 15, 240. [Google Scholar] [CrossRef] [Green Version]
- Lois, L.C.; Diéguez, S.C.; Peña, M.L.G.D.L.; Gómez, E.F. SPE-GC determination of aromatic compounds in two varieties of white grape during ripening. Chromatographia 2001, 53, S350–S355. [Google Scholar] [CrossRef]
- Coelho, E.; Rocha, S.M.; Barros, A.S.; Delgadillo, I.; Coimbra, M.A. Screening of variety and prefermentation-related volatile compounds during ripening of white grapes to define their evolution profile. Anal. Chim. Acta 2007, 597, 257–264. [Google Scholar] [CrossRef]
- Zhang, E.; Chai, F.; Zhang, H.; Li, S.; Liang, Z.; Fan, P. Effects of sunlight exclusion on the profiles of monoterpene biosynthesis and accumulation in grape exocarp and mesocarp. Food Chem. 2017, 237, 379–389. [Google Scholar] [CrossRef]
- Mandaokar, A.; Thines, B.; Shin, B.; Lange, B.M.; Choi, G.; Koo, Y.J.; Yoo, Y.J.; Choi, Y.; Choi, G.; Browse, J. Transcriptional regulators of stamen development in Arabidopsis identified by transcriptional profiling. Plant J. Cell Mol. Biol. 2010, 46, 984–1008. [Google Scholar] [CrossRef]
- Bedon, F.; Bomal, C.; Caron, S.; Levasseur, C.; Boyle, B.; Mansfield, S.D.; Schmidt, A.; Gershenzon, J.; Grimapettenati, J.; Séguin, A. Subgroup 4 R2R3-MYBs in conifer trees: Gene family expansion and contribution to the isoprenoid- and flavonoid-oriented responses. J. Exp. Bot. 2010, 61, 3847–3864. [Google Scholar] [CrossRef] [PubMed]
- Skibbe, M.; Qu, N.; Galis, I.; Baldwin, I.T. Induced plant defenses in the natural environment: Nicotiana attenuata WRKY3 and WRKY6 coordinate responses to herbivory. Plant Cell 2008, 20, 1984–2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, G.-J.; Xue, X.Y.; Mao, Y.-B.; Wang, L.-J.; Chen, X. Arabidopsis MYC2 interacts with DELLA proteins in regulating sesquiterpene synthase gene expression. Plant Cell 2012, 24, 2635–2648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, K.; Matsumoto, T.; Okada, A.; Komiyama, K.; Chujo, T.; Yoshikawa, H.; Nojiri, H.; Yamane, H.; Okada, K. Identification of target genes of the bZIP transcription factor OsT- GAP1, whose overexpression causes elicitor induced hyperaccumulation of diterpenoid phytoalexins in rice cells. PLoS ONE 2014, 9, e105823. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, K.; Nishizawa, Y.; Minami, E.; Nojiri, H.; Yamane, H.; Okada, K. Overexpression of the bZIP transcription factor OsbZIP79 sup- presses the production of diterpenoid phytoalexin in rice cells. J. Plant Physiol. 2015, 173, 19–27. [Google Scholar] [CrossRef]
- Zhang, A.; Hu, X.; Kuang, S.; Ge, H.; Yin, X.; Chen, K. Isolation, classification and transcription profiles of the Ethylene Response Factors (ERFs) in ripening kiwifruit. Sci. Hortic. 2016, 199, 209–215. [Google Scholar] [CrossRef]
- Nieuwenhuizen, N.J.; Allan, A.C.; Atkinson, R.G. The Genetics of Kiwifruit Flavor and Fragrance; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Reddy, V.A.; Wang, Q.; Dhar, N.; Kumar, N.; Venkatesh, P.N.; Rajan, C.; Panicker, D.; Sridhar, V.; Mao, H.Z.; Sarojam, R. Spearmint R2R3-MYB transcription factor MsMYB negatively regulates monoterpene production and suppresses the expression of geranyl diphosphate synthase large subunit (MsGPPS.LSU). Plant Biotehnol. J. 2017, 15, 1105–1119. [Google Scholar] [CrossRef] [Green Version]
- Chuang, Y.-C.; Hung, Y.-C.; Tsai, W.-C.; Chen, W.-H.; Chen, H.-H. PbbHLH4 regulates floral monoterpene biosynthesis in Phalaenopsis orchids. J. Exp. Bot. 2018, 69, 4363–4377. [Google Scholar] [CrossRef] [Green Version]
- Cramer, G.R.; Ghan, R.; Schlauch, K.A.; Tillett, R.L.; Heymann, H.; Ferrarini, A.; Delledonne, M.; Zenoni, S.; Fasoli, M.; Pezzotti, M. Transcriptomic analysis of the late stages of grapevine (Vitis vinifera cv. Cabernet Sauvignon) berry ripening reveals significant induction of ethylene signaling and flavor pathways in the skin. BMC Plant Biol. 2014, 14, 370. [Google Scholar] [CrossRef] [Green Version]
- Czemmel, S.; Stracke, R.; Weisshaar, B.; Cordon, N.; Harris, N.N.; Walker, A.R.; Robinson, S.P.; Bogs, J. The grapevine R2R3-MYB transcription factor VvMYBF1 regulates flavonol synthesis in developing grape berries. Plant Physiol. 2009, 151, 1513–1530. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Cloning: A Laboratory Manual; Cold Spring Harbor Protocols/Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Mu, L.; He, F.; Pan, Q.-H.; Zhou, L.; Duan, C.-Q. Screening and verification of late embryogenesis abundant protein interacting with anthocyanidin reductase in grape berries. Vitis J. Grapevine Res. 2014, 53, 81–87. [Google Scholar]
- Sun, R.Z.; Pan, Q.H.; Duan, C.Q.; Wang, J. Light response and potential interacting proteins of a grape flavonoid 3’-hydroxylase gene promoter. Plant Physiol. Biochem. 2015, 97, 70–81. [Google Scholar] [CrossRef]
- Vinatzer, B.A.; Liu, D.; Shi, L.; Han, C.; Yu, J.; Li, D.; Zhang, Y. Validation of reference genes for gene expression studies in virus-infected nicotiana benthamiana using quantitative real-time PCR. PLoS ONE 2012, 7, e46451. [Google Scholar]
- Zhao, R.; Wang, X.F.; Zhang, D.P. CPK12: A Ca2+-dependent protein kinase balancer in abscisic acid signaling. Plant. Signal. Behav. 2011, 6, 1687–1690. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsi mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2013. [Google Scholar] [CrossRef] [Green Version]
- Holl, J.; Vannozzi, A.; Czemmel, S.; D’Onofrio, C.; Walker, A.R.; Rausch, T.; Lucchin, M.; Boss, P.K.; Dry, I.B.; Bogs, J. The R2R3-MYB transcription factors MYB14 and MYB15 regulate stilbene biosynthesis in Vitis vinifera. Plant Cell 2013, 25, 4135–4149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.; Bausewein, A.; Greiner, S.; Dauchot, N.; Harms, K.; Rausch, T. CiMYB17, a stress-induced chicory R2R3-MYB transcription factor, activates promoters of genes involved in fructan synthesis and degradation. New Phytol. 2017, 215, 281–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogs, J.; Jaffé, F.W.; Takos, A.M.; Walker, A.R.; Robinson, S.P. The grapevine transcription factor VvMYBPA1 regulates proanthocyanidin synthesis during fruit development. Plant Physiol. 2007, 143, 1347–1361. [Google Scholar] [CrossRef] [Green Version]
- Lan, Y.B.; Qian, X.; Yang, Z.J.; Xiang, X.F.; Yang, W.X.; Liu, T.; Zhu, B.Q.; Pan, Q.H.; Duan, C.Q. Striking changes in volatile profiles at sub-zero temperatures during over-ripening of ‘Beibinghong’ grapes in Northeastern China. Food Chem. 2016, 212, 172–182. [Google Scholar] [CrossRef]
- Gao, Y.; Li, X.X.; Han, M.M.; Yang, X.F.; Li, Z.; Wang, J.; Pan, Q.H. Rain-shelter cultivation modifies carbon allocation in the polyphenolic and volatile metabolism of Vitis vinifera L. chardonnay grapes. PLoS ONE 2016, 11, e0156117. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.-Q.; Liu, B.; Zhu, B.-Q.; Lan, Y.-B.; Gao, Y.; Wang, D.; Reeves, M.J.; Duan, C.-Q. Differences in volatile profiles of Cabernet Sauvignon grapes grown in two distinct regions of China and their responses to weather conditions. Plant Physiol. Biochem. 2015, 89, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhang, M.; Ren, J.; Qi, J.; Zhang, G.; Leng, P. Reciprocity between abscisic acid and ethylene at the onset of berry ripening and after harvest. BMC Plant Biol. 2010, 10, 257. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.J.; Li, M.; Chen, J.; Liu, P.; Li, Z. Effects of MeJA on arabidopsis metabolome under endogenous JA deficiency. Sci. Rep. 2016, 6, 37674. [Google Scholar] [CrossRef] [Green Version]
- Fenoll, J.; Manso, A.; Hellín, P.; Ruiz, L.; Flores, P. Changes in the aromatic composition of the Vitis vinifera grape Muscat Hamburg during ripening. Food Chem. 2009, 114, 420–428. [Google Scholar] [CrossRef]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef]
- Chen, H.; Lai, Z.; Shi, J.; Xiao, Y.; Chen, Z.X.; Xu, X.P. Roles of arabidopsis WRKY18, WRKY40 and WRKY60 transcription factors in plant responses to abscisic acid and abiotic stress. BMC Plant Biol. 2010, 10, 281. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhu, W.; Fang, L.; Sun, X.; Su, L.; Liang, Z.; Wang, N.; Londo, J.P.; Li, S.H.; Xin, H.P. Genome-wide identification of WRKY family genes and their response to cold stress in Vitis vinifera. BMC Plant Biol. 2014, 14, 103. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Guo, R.; Xu, X.; Gao, M.; Li, X.; Song, J.; Zheng, Y.; Wang, X. Evolution and expression analysis of the grape (Vitis vinifera L.) WRKY gene family. J. Exp. Bot. 2014, 65, 1513–1528. [Google Scholar] [CrossRef]
- Zhu, D.; Hou, L.; Xiao, P.; Guo, Y.; Deyholos, M.K.; Liu, X. VvWRKY30, a grape WRKY transcription factor, plays a positive regulatory role under salinity stress. Plant Sci. 2018, 280, 132–142. [Google Scholar] [CrossRef]
- Jiang, J.; Xi, H.; Dai, Z.; Lecourieux, F.; Yuan, L.; Liu, X.; Patra, B.; Wei, Y.; Li, S.; Wang, L. VvWRKY8 represses stilbene synthase genes through direct interaction with VvMYB14 to control resveratrol biosynthesis in grapevine. J. Exp. Bot. 2019, 70, 715–729. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.H.; Wang, J.W.; Wang, S.; Wang, J.Y.; Chen, X.Y. Characterization of GaWRKY1, a cotton transcription factor that regulates the sesquiterpene synthase gene (+)-delta-cadinene synthase A. Plant Physiol. 2004, 135, 507–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Y.; Yan, L.; Liu, Z.Q.; Cao, Z.; Mei, C.; Xin, Q.; Wu, F.Q.; Wang, X.F.; Du, S.Y.; Jiang, T.; et al. The Mg-chelatase H subunit of Arabidopsis antagonizes a group of WRKY transcription repressors to relieve ABA-responsive genes of inhibition. Plant Cell 2010, 22, 1909–1935. [Google Scholar] [CrossRef] [Green Version]
- Young, P.R.; Lashbrooke, J.G.; Alexandersson, E.; Jacobson, D.; Moser, C.; Velasco, R.; Vivier, M.A. The genes and enzymes of the carotenoid metabolic pathway inVitis viniferaL. BMC Genom. 2012, 13, 243. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Wang, S.; Lin, H.; Satio, T.; Ampa, K.; Todoroki, Y.; Kondo, S. Effects of abscisic acid agonist or antagonist applications on aroma volatiles and anthocyanin biosynthesis in grape berries. J. Hortic. Sci. Biotechnol. 2017, 93, 392–399. [Google Scholar] [CrossRef]
- Portnoy, V.; Bar, E.; Shimoni-Shor, E.; Feder, A.; Saar, Y.; Saar, U.; Baumkoler, F.; Lewinsohn, E.; Schaffer, A.A.; Katzir, N.; et al. Genetic and chemical characterization of an EMS induced mutation in Cucumis melo CRTISO gene. Arch. Biochem. Biophys. 2013, 539, 117–125. [Google Scholar]
- Liu, Z.Q.; Yan, L.; Wu, Z.; Mei, C.; Lu, K.; Yu, Y.T.; Liang, S.; Zhang, X.F.; Wang, X.F.; Zhang, D.P. Cooperation of three WRKY-domain transcription factors WRKY18, WRKY40 and WRKY60 in repressing two ABA-responsive genes ABI4 and ABI5 in Arabidopsis. J. Exp. Bot. 2012, 18, 63. [Google Scholar] [CrossRef] [Green Version]
- Rushton, D.L.; Tripathi, P.; Rabara, R.C.; Lin, J.; Ringler, P.; Boken, A.K.; Langum, T.J.; Smidt, L.; Boomsma, D.D.; Emme, N.J.; et al. WRKY transcription factors: Key components in abscisic acid signaling. J. Plant. Biotech. 2012, 10, 2–11. [Google Scholar] [CrossRef]
- Fraser, L.G.; Seal, A.G.; Montefiori, M.; McGhie, T.K.; Allan, A.C. An R2R3 MYB transcription factor determines red petal colour in an Actinidia (kiwifruit) hybrid population. BMC Genom. 2013, 14, 28. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.L.; Xia, X.J.; Kuang, S.; Zhang, X.L.; Yin, X.R.; Yu, J.Q.; Chen, K.S. A novel ethylene responsive factor CitERF13 plays a role in photosynthesis regulation. Plant Sci. 2017, 256, 112–119. [Google Scholar] [CrossRef]
- Min, T.; Yin, X.-R.; Shi, Y.-N.; Luo, Z.-R.; Yao, Y.-C.; Grierson, D.; Ferguson, I.B.; Chen, K.-S. Ethylene-responsive transcription factors interact with promoters of ADH and PDC involved in persimmon (Diospyros kaki) fruit de-astringency. J. Exp. Bot. 2012, 63, 6393–6405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabi, J.P.; Broetto, S.G.; Da Silva, S.L.G.L.; Zhong, S.; Lajolo, F.M.; Nascimento, J.R.O.D. Analysis of Papaya Cell Wall-Related Genes during Fruit Ripening Indicates a Central Role of Polygalacturonases during Pulp Softening. PLoS ONE 2014, 9, e105685. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Id | Function |
|---|---|
| VIT_204s0008g06000 | PREDICTED: ethylene-responsive transcription factor ERF003 [Vitis vinifera] |
| VIT_206s0004g00490 | PREDICTED: ethylene-responsive transcription factor RAP2-11 [Vitis vinifera] |
| VIT_209s0002g00470 | PREDICTED: ethylene-responsive transcription factor RAP2-12-like [Vitis vinifera] |
| VIT_202s0234g00130 | PREDICTED: ethylene-responsive transcription factor2-like [Vitis vinifera] |
| VIT_216s0013g01090 | PREDICTED: ethylene-responsive transcription factor5-like [Vitis vinifera] |
| VIT_204s0008g05760 | PREDICTED: WRKY transcription factor 40 [Vitis vinifera] |
| VIT_207s0129g00030 | PREDICTED: protein SHORT-ROOT-like isoform 1 [Vitis vinifera] |
| VIT_206s0009g01790 | PREDICTED: ABSCISIC ACID-INSENSITIVE 5-like protein 2 [Vitis vinifera] |
| VIT_205s0077g01120 | hypothetical protein VITISV_039159 [Vitis vinifera] |
| VIT_208s0007g03900 | PREDICTED: heat stress transcription factor A-3-like [Vitis vinifera] |
| Gene Id | Function |
|---|---|
| XM_010660100.2 | PREDICTED: probable WRKY transcription factor 22 [Vitis vinifera] |
| XM_002272004.4 | PREDICTED: probable WRKY transcription factor 24 [Vitis vinifera] |
| XM_002276158.4 | PREDICTED: probable WRKY transcription factor 32 [Vitis vinifera] |
| XM_010649972.2 | PREDICTED: probable WRKY transcription factor 40 [Vitis vinifera] |
| XM_019221865.1 | PREDICTED: probable WRKY transcription factor 26 [Vitis vinifera] |
| XM_002272684.3 | PREDICTED: probable WRKY transcription factor 41 [Vitis vinifera] |
| XM_010662802.2 | PREDICTED: probable WRKY transcription factor 4 [Vitis vinifera] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; He, L.; An, X.; Yu, K.; Meng, N.; Duan, C.; Pan, Q.-H. VviWRKY40, a WRKY Transcription Factor, Regulates Glycosylated Monoterpenoid Production by VviGT14 in Grape Berry. Genes 2020, 11, 485. https://doi.org/10.3390/genes11050485
Li X, He L, An X, Yu K, Meng N, Duan C, Pan Q-H. VviWRKY40, a WRKY Transcription Factor, Regulates Glycosylated Monoterpenoid Production by VviGT14 in Grape Berry. Genes. 2020; 11(5):485. https://doi.org/10.3390/genes11050485
Chicago/Turabian StyleLi, Xiangyi, Lei He, Xiaohui An, Keji Yu, Nan Meng, Changqing Duan, and Qiu-Hong Pan. 2020. "VviWRKY40, a WRKY Transcription Factor, Regulates Glycosylated Monoterpenoid Production by VviGT14 in Grape Berry" Genes 11, no. 5: 485. https://doi.org/10.3390/genes11050485
APA StyleLi, X., He, L., An, X., Yu, K., Meng, N., Duan, C., & Pan, Q. -H. (2020). VviWRKY40, a WRKY Transcription Factor, Regulates Glycosylated Monoterpenoid Production by VviGT14 in Grape Berry. Genes, 11(5), 485. https://doi.org/10.3390/genes11050485