Genetic Diversity and Population Structure of Llamas (Lama glama) from the Camelid Germplasm Bank—Quimsachata

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
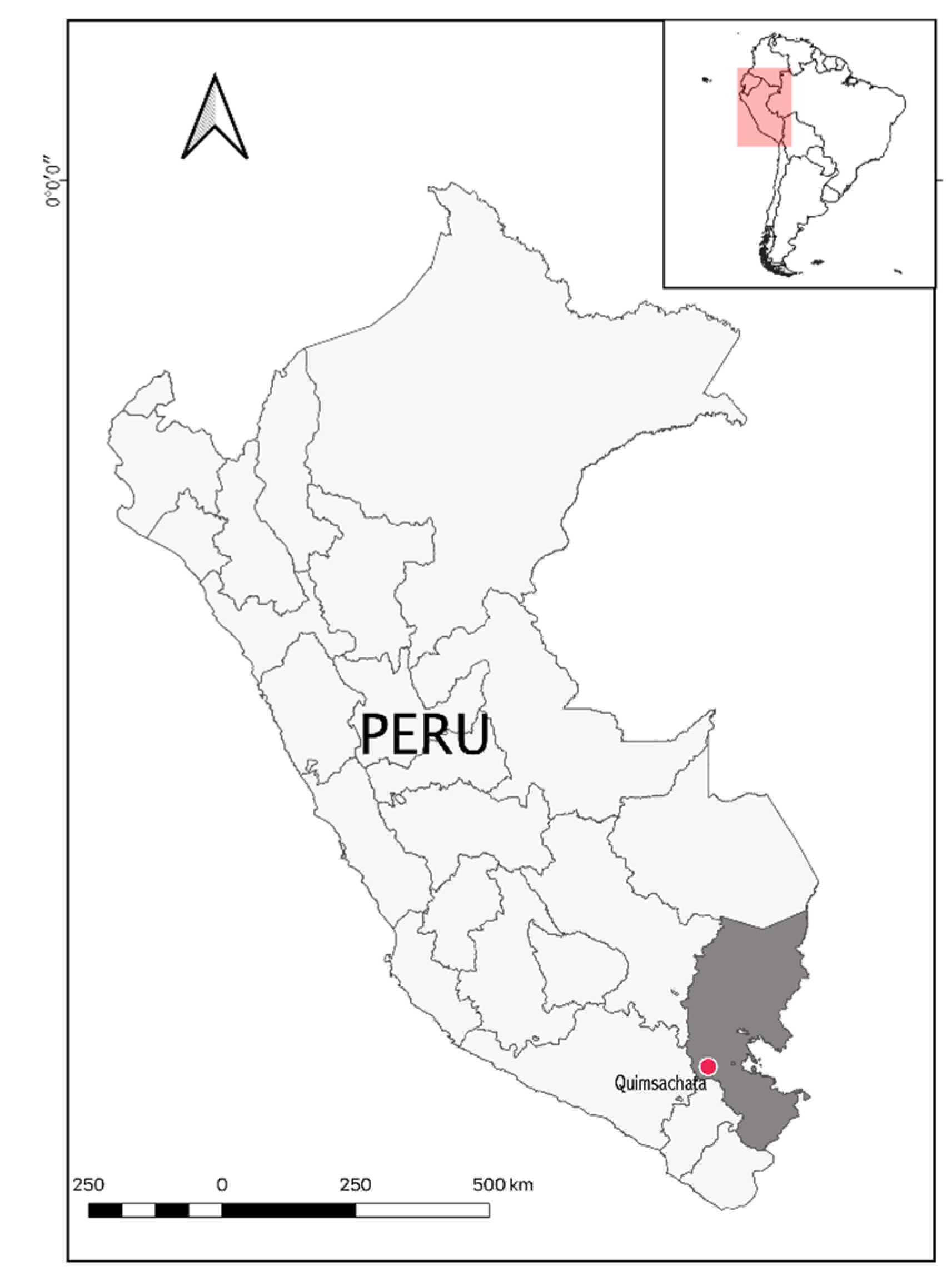
2.1. Sample Collection
2.2. DNA Amplification and Microsatellite Genotyping
2.3. Statistical Analysis
3. Results
3.1. Genetic Diversity Assessment
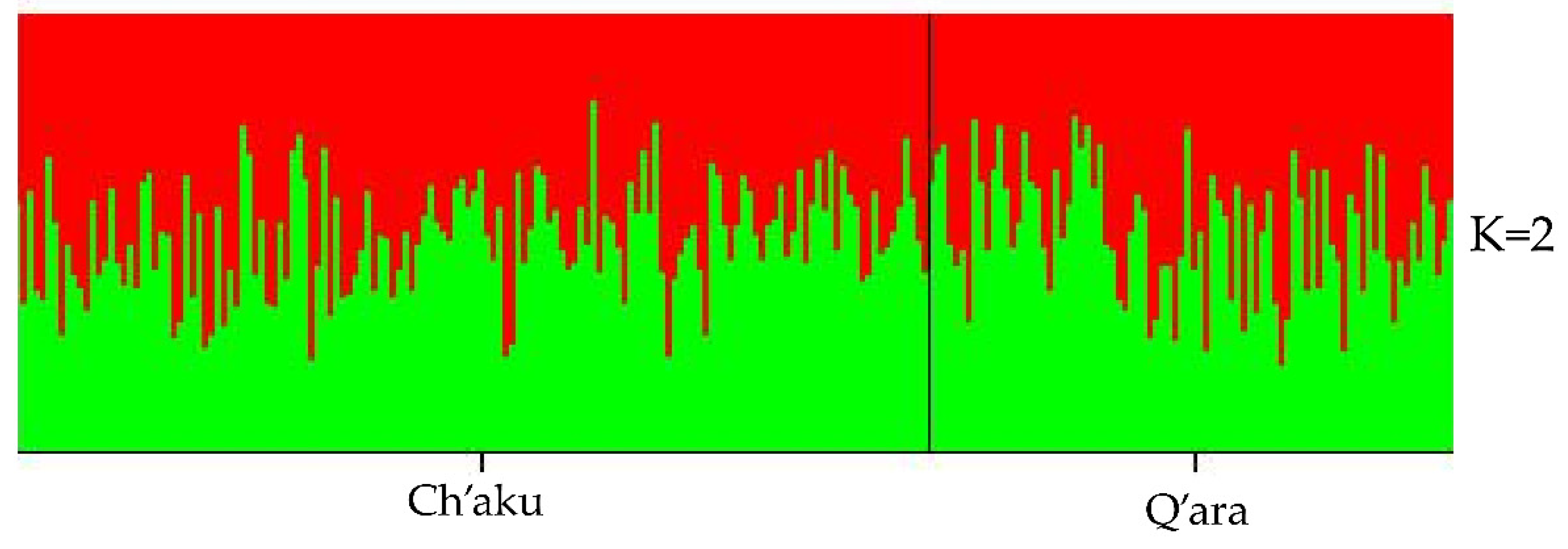
3.2. Genetic Differentiation between Ch’aku and Q’ara
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bustamante, A.V.; Maté, M.L.; Zambelli, A.; Vidal-Rioja, L.; Lamas, H.E.; Giovambattista, G. Análisis de diversidad genética en tres poblaciones de llamas (Lama glama) del noroeste argentino. Rev. Chil. Hist. Nat. 2006, 79, 175–184. [Google Scholar] [CrossRef]
- Barreta, J.; Gutiérrez-Gil, B.; Iñiguez, V.; Romero, F.; Saavedra, V.; Chiri, R.; Rodríguez, T.; Arranz, J.J. Analysis of genetic diversity in Bolivian llama populations using microsatellites. J. Anim. Breed. Genet. 2013, 130, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Fernández, B.S. Situación Actual de los Camélidos Sudamericanos en Perú. Proyecto de Cooperación Técnica en Apoyo de la Crianza y Aprovechamiento de los Camélidos Sudamericanos en la Región Andina Tcp/Rla/2914; Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO): Roma, Italy, 2005. [Google Scholar]
- Llacsa, J. Determinación de Indicadores Biométricos para la Selección de Llamas (Lama glama) Productoras de Carne. Ph.D. Thesis, Tesis de Médico Veterinario y Zootecnista, Universidad Nacional del Altiplano, Puno, Peru, 2006. [Google Scholar]
- Wilson, D.E.; Reeder, D.M. Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2005; Volume 1, p. 645, ISBN 0801882214, 9780801882210. [Google Scholar]
- Wheeler, J.C. Evolution and present situation of the South American Camelidae. Biol. J. Linn. Soc. 1995, 54, 271–295. [Google Scholar] [CrossRef]
- Iñiguez, L.C.; Alem, R.; Wauer, A.; Mueller, J. Fleece types, fiber characteristics and production system of an outstanding llama population from Southern Bolivia. Small Rumin. Res. 1998, 30, 57–65. [Google Scholar] [CrossRef]
- Stemmer, A.; Wurzinger, M.; Soelkner, J.; Valle Zárate, A.; Nuernberg, M.; Delgado, J. La llama de Ayopaya: Descripción de un recurso genético autóctono. Arch. Zootec. 2005, 54, 253–259. [Google Scholar]
- Maquera, F.E. Caracterización y Persistencia Fenotípica en Llamas Karas y Lanudas del Centro Experimental La Raya-Puno. Master’s Thesis, Universidad Nacional Agraria La Molina, Lima, Peru, 1991. [Google Scholar]
- INEI (Instituto Nacional de Estadística e Informática). IV Censo Nacional Agropecuario 2012, Lima, Peru. Available online: http://proyectos.inei.gob.pe/web/DocumentosPublicos/ResultadosFinalesIVCENAGRO.pdf (accessed on 25 July 2019).
- INEI (Instituto Nacional de Estadística e Informática). III Censo Nacional Agropecuario 1994, Lima, Peru. Available online: http://censos.inei.gob.pe/bcoCuadros/IIIcenagro.htm (accessed on 10 July 2019).
- Gutierrez, G.A.; Gutierrez, J.P.; Huanca, T.; Wurzinger, M. Challenges and opportunities of genetic improvement in alpacas and llamas in Peru. In Proceedings of the World Congress on Genetics Applied to Livestock Production, Auckland, New Zealand, 12–16 February 2018; p. 762. [Google Scholar]
- Huanca, T.; Apaza, N.; Gonzales, M. Experiencia del INIA en el fortalecimiento del Banco de germoplasma de camélidos domésticos. Arch. Latinoam. Prod. Anim. 2007, 15, 186–194. [Google Scholar]
- Wurzinger, M.; Gutiérrez, G. Analysis of a multi-stakeholder process during the start-up phase of two community-based llama breeding programs in Peru. Livest. Res. Rural. Dev. 2017, 29, 10. [Google Scholar]
- Hammond, K. Conservation of domestic animal diversity: Global overview. In Proceedings of the 5th World Congress on Genetics Applied to Livestock Production, Guelph, Canada, 7–12 August 1994; Volume 21, pp. 423–439. [Google Scholar]
- Luikart, G.; Gielly, L.; Excoffier, L.; Vigne, J.D.; Bouvet, J.; Taberlet, P. Multiple maternal origins and weak phylogeographic structure in domestic goats. Proc. Natl. Acad. Sci. USA 2001, 98, 5927–5932. [Google Scholar] [CrossRef] [Green Version]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics, 4th ed.; Logmans Green: Harlow, UK, 1996; ISBN-13 978-0582243026. [Google Scholar]
- Ihara, N.; Takasuga, A.; Mizoshita, K.; Takeda, H.; Sugimoto, M.; Mizoguchi, Y.; Hirano, T.; Itoh, T.; Watanabe, T.; Reed, K.M.; et al. A comprehensive genetic map of the cattle genome based on 3802 microsatellites. Genome Res. 2004, 14, 1987–1998. [Google Scholar] [CrossRef] [Green Version]
- Slatkin, M. A measure of population subdivision based on microsatellite allele frequencies. Genetics 1995, 139, 457–462. [Google Scholar]
- Maté, M.L.; Bustamante, A.; Giovambattista, G.; De Lamo, D.; Von Thüngen, J.; Zambelli, A.; Vidal-Rioja, L. Genetic diversity and differentiation of guanaco populations from Argentina inferred from microsatellite data. Anim. Genet. 2005, 36, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Paredes, M.; Machaca, J.; Azor, P.J.; Alonso-Moraga, A.; Membrillo, A.; Muñoz-Serrano, A. Genetic Differentiation of Six Peruvian Alpaca Populations. In Fibre Production in South American Camelids and Other Fibre Animals; Wageningen Academic Publishers: Wageningen, The Netherlands, 2011; pp. 161–166. [Google Scholar]
- Lang, K.D.M.; Wang, Y.; Plante, Y. Fifteen polymorphic dinucleotide microsatellites in llamas and alpacas. Anim. Genet. 1996, 27, 293. [Google Scholar] [CrossRef]
- Penedo, M.C.T.; Caetano, A.R.; Cordova, K.I. Microsatellite markers for South American camelids. Anim. Genet. 1998, 29, 411–412. [Google Scholar] [PubMed]
- Obreque, V.; Coogle, L.; Henney, P.J.; Bailey, E.; Mancilla, R.; García-Huidobro, J.; Hinrichsen, P.; Cothran, E.G. Characterization of 10 polymorphic alpaca dinucleotide microsatellites. Anim. Genet. 1998, 29, 461–462. [Google Scholar] [PubMed]
- Obreque, V.; Mancilla, R.; García-Huidobro, J.; Cothran, E.G.; Hinrichsen, P. Thirteen new dinucleotide microsatellites in Alpaca. Anim. Genet. 1999, 30, 397–398. [Google Scholar] [CrossRef] [PubMed]
- Penedo, M.C.T.; Caetano, A.R.; Cordova, K. Eight microsatellite markers for South American camelids. Anim. Genet. 1999, 30, 166–167. [Google Scholar] [CrossRef]
- Penedo, M.C.T.; Caetano, A.R.; Cordova, K.I. Six microsatellite markers for South American camelids. Anim. Genet. 1999, 30, 399. [Google Scholar] [CrossRef]
- Sarno, R.J.; David, V.A.; Franklin, W.L.; O’Brien, S.J.; Johnson, W.E. Development of microsatellite markers in the guanaco, Lama guanicoe: Utility for South American camelids. Mol. Ecol. 2000, 9, 1922–1924. [Google Scholar] [CrossRef]
- Wheeler, J.C.; Chikhi, L.; Bruford, M.W. Genetic analysis of the origins of domestic South American camelids. In Archaeology and Animal Domestication: New Genetic and Archaeological Paradigms: New Genetic and Archaeological Paradigms; Zeder, M., Ed.; University of California Press: Berkeley, CA, USA, 2006; pp. 329–341. [Google Scholar]
- Rodríguez, J.B.; Wheeler, J.C.; Dodd, C.S.; Bruford, M.W.; Rosadio, R.A. Determinación de parentesco en alpacas (Vicugna pacos) por medio del análisis de ADN microsatélite. Rev. Investig. Vet. del Peru 2004, 15, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Marín, J.C.; Casey, C.S.; Kadwell, M.; Yaya, K.; Hoces, D.; Olazabal, J.; Rosadio, R.; Rodriguez, J.; Spotorno, A.; Bruford, M.W.; et al. Mitochondrial phylogeography and demographic history of the Vicuña: Implications for conservation. Heredity 2007, 99, 70–80. [Google Scholar] [CrossRef] [Green Version]
- Melo, C.; Manunza, A.; Melo, M.; Olivera, L.; Amills, M. Analysis of the mitochondrial diversity of alpacas in eight farming areas of the south of Peru. In Fibre Production in South American Camelids and Other Fibre Animals; Wageningen Academic Publishers: Wageningen, The Netherlands, 2011; pp. 87–91. [Google Scholar] [CrossRef]
- Kadwell, M.; Fernandez, M.; Stanley, H.F.; Baldi, R.; Wheeler, J.C.; Rosadio, R.; Bruford, M.W. Genetic analysis reveals the wild ancestors of the llama and the alpaca. Proc. R. Soc. B Biol. Sci. 2001, 268, 2575–2584. [Google Scholar] [CrossRef] [Green Version]
- Cano, Z.L.; Rosadio, A.R.; Maturrano, H.L.; Dávalos, R.R.; Wheeler, J.C. Caracterización fenotípica y análisis de ADN mitocondrial de llamas de marcapomacocha, Peru. Rev. Investig. Vet. Peru. 2012, 23, 388–398. [Google Scholar] [CrossRef] [Green Version]
- Heath, D.; Smith, P.; Williams, D.; Harris, P.; Arias-Stella, J.; Krüger, H. The heart and pulmonary vasculature of the llama (Lama glama). Thorax 1974, 29, 463–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cristofanelli, S.; Antonini, M.; Torres, D.; Polidori, P.; Renieri, C. Meat and carcass quality from Peruvian llama (Lama glama) and alpaca (Lama pacos). Meat Sci. 2004, 66, 589–593. [Google Scholar] [CrossRef]
- Cristofanelli, S.; Antonini, M.; Torres, D.; Polidori, P.; Renieri, C. Carcass characteristics of peruvian llama (Lama glama) and alpaca (Lama pacos) reared in the Andean highlands. Small Rumin. Res. 2005, 58, 219–222. [Google Scholar] [CrossRef]
- Quispe, E.C.; Rodríguez, T.C.; Iñiguez, L.R.; Mueller, J.P. Producción de fibra de alpaca, llama, vicuña y guanaco en Sudamérica. Anim. Genet. Resour. 2009, 45, 1–14. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Bustamante, A.V.; Maté, M.L.; Zambelli, A.; Vidal-Rioja, L. Isolation and characterization of 10 polymorphic dinucleotide microsatellite markers for llama and guanaco. Mol. Ecol. Notes 2003, 3, 68–69. [Google Scholar] [CrossRef]
- De Arruda, M.P.; Gonçalves, E.C.; Schneider, M.P.C.; Da Costa Da Silva, A.L.; Morielle-Versute, E. An alternative genotyping method using dye-labeled universal primer to reduce unspecific amplifications. Mol. Biol. Rep. 2010, 37, 2031–2036. [Google Scholar] [CrossRef]
- Marshall, T.C.; Slate, J.; Kruuk, L.E.B.; Pemberton, J.M. Statistical confidence for likelihood-based paternity inference in natural populations. Mol. Ecol. 1998, 7, 639–655. [Google Scholar] [CrossRef] [Green Version]
- Belkhir, K.; Borsa, P.; Chikhi, L.; Raufaste, N.; Bonhomme, F. GENETIX 4.05, Logiciel sous WindowsTM pour la génétique des populations. In Laboratoire Génome, Populations, Interactions, CNRS UMR 5171; Université de Montpellier II: Montpellier, France, 2004. [Google Scholar]
- Raymond, M.; Rousset, F. GENEPOP (Version 1.2): Population Genetics Software for Exact Tests and Ecumenicism. J. Hered 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinforma 2005, 1. [Google Scholar] [CrossRef] [Green Version]
- Wright, S. Evolution and the Genetics of Populations, Volume 4, Variability within and among Natural Populations; University of Chicago Press: Chicago, IL, USA, 1984; p. 590, ISBN-13: 978-0226910529. [Google Scholar]
- FSTAT A Program to Estimate and Test Gene Diversities and Fixation Indices (Version 2.9.3). 2001. 149, 507–526. Available online: https://www2.unil.ch/popgen/softwares/fstat.htm (accessed on 20 February 2019).
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Moritz, C. Conservation units and translocations: Strategies for conserving evolutionary processes. Hereditas 1999, 130, 217–228. [Google Scholar] [CrossRef]
- Aranguren-Méndez, J.; Jordana, J.; Gomez, M. Genetic diversity in Spanish donkey breeds using microsatellite DNA markers. Genet. Sel. Evol. 2001, 33, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, A.V.; Zambelli, A.; De Lamo, D.A.; Von Thungen, J.; Vidal-Rioja, L. Genetic variability of guanaco and llama populations in Argentina. Small Rumin. Res. 2002, 44, 97–101. [Google Scholar] [CrossRef]
- Callen, D.F.; Thompson, A.D.; Shen, Y.; Phillips, H.A.; Richards, R.I.; Mulley, J.C.; Sutherland, G.R. Incidence and origin of “null” alleles in the (AC)n microsatellite markers. Am. J. Hum. Genet. 1993, 52, 922. [Google Scholar] [CrossRef]
- Franklin, W.L. Biology, ecology, and relationship to man of the South American camelids. Mamm. Biol. S. Am. 1982, 6, 457–489. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Micro Satellite | Ch’aku | Q’ara | Global Population | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | Ho | He | PIC | A | Ho | He | PIC | A | Ho | He | PIC | |
| GLM4 | 9 | 0.66 | 0.78 | 0.75 | 10 | 0.76 | 0.78 | 0.66 | 10 | 0.72 | 0.78 | 0.71 |
| LAB1 | 13 | 0.72 | 0.79 | 0.53 | 13 | 0.70 | 0.73 | 0.5 | 14 | 0.71 | 0.76 | 0.52 |
| LCA54 | 4 | 0.6 | 0.61 | 0.75 | 8 | 0.53 | 0.56 | 0.75 | 8 | 0.56 | 0.59 | 0.75 |
| LCA65 | 8 | 0.61 | 0.65 | 0.82 | 8 | 0.61 | 0.64 | 0.77 | 9 | 0.61 | 0.64 | 0.79 |
| LCA77 | 8 | 0.66 | 0.64 | 0.86 | 10 | 0.67 | 0.68 | 0.86 | 11 | 0.66 | 0.67 | 0.86 |
| LCA82 | 9 | 0.68 | 0.79 | 0.82 | 7 | 0.52 | 0.72 | 0.81 | 9 | 0.57 | 0.75 | 0.82 |
| LCA83 | 8 | 0.79 | 0.78 | 0.77 | 7 | 0.72 | 0.74 | 0.71 | 8 | 0.75 | 0.76 | 0.74 |
| LCA85 | 8 | 0.76 | 0.85 | 0.90 | 7 | 0.74 | 0.81 | 0.86 | 8 | 0.75 | 0.82 | 0.88 |
| LGU76 | 14 | 0.83 | 0.84 | 0.83 | 14 | 0.75 | 0.79 | 0.78 | 15 | 0.78 | 0.81 | 0.8 |
| YWLL08 | 17 | 0.90 | 0.91 | 0.59 | 17 | 0.83 | 0.87 | 0.63 | 19 | 0.86 | 0.89 | 0.62 |
| YWLL44 | 16 | 0.84 | 0.89 | 0.58 | 17 | 0.85 | 0.87 | 0.59 | 17 | 0.85 | 0.875 | 0.59 |
| YWLL59 | 15 | 0.73 | 0.84 | 0.74 | 17 | 0.75 | 0.84 | 0.70 | 19 | 0.74 | 0.84 | 0.73 |
| VOLP03 | 9 | 0.74 | 0.68 | 0.62 | 8 | 0.59 | 0.64 | 0.57 | 10 | 0.64 | 0.66 | 0.59 |
| Average | 10.5 | 0.73 | 0.77 | 0.74 | 10.9 | 0.69 | 0.74 | 0.71 | 12.08 | 0.71 | 0.76 | 0.72 |
| Global Population | Ch’aku Population | Q’ara Population | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Microsatellite | Deficit of Heterozygotes (p-Value) | Standard Deviation | FIS | Microsatellite | Deficit of Heterozygotes (p-Value) | Standard Deviation | FIS | Microsatellite | Deficit of Heterozygotes (p-Value) | Standard Deviation | FIS |
| GLM4 | 0.0062 | 0.0014 | 0.073 | GLM4 | 0.0016 | 0.0004 | 0.145 | GLM4 | 0.2851 | 0.012 | 0.0335 |
| LAB1 | 0.0002 | 0.0002 | 0.056 | LAB1 | 0 | 0 | 0.087 | LAB1 | 0.176 | 0.0128 | 0.0383 |
| LCA54 | 0.0217 | 0.0051 | 0.04 | LCA54 | 0.0831 | 0 | 0.0211 | LCA54 | 0.0475 | 0.0054 | 0.0512 |
| LCA65 | 0.054 | 0.0134 | 0.054 | LCA65 | 0.2489 | 0.0167 | 0.0676 | LCA65 | 0.1858 | 0.0127 | 0.0467 |
| LCA77 | 0.259 | 0.015 | 0.004 | LCA77 | 0.6392 | 0.0144 | −0.0216 | LCA77 | 0.2113 | 0.0206 | 0.0171 |
| LCA82 | 0.0001 | 0.0002 | 0.223 | LCA82 | 0.0142 | 0.0018 | 0.1327 | LCA82 | 0.0024 | 0.0006 | 0.2788 |
| LCA83 | 0.0463 | 0.0066 | 0.01 | LCA83 | 0.0302 | 0.0025 | −0.0152 | LCA83 | 0.2907 | 0.0089 | 0.0251 |
| LCA85 | 0.0002 | 0.0001 | 0.089 | LCA85 | 0.0038 | 0.0006 | 0.1191 | LCA85 | 0.0155 | 0.0013 | 0.0788 |
| LGU76 | 0.139 | 0.0227 | 0.039 | LGU76 | 0.1939 | 0.0128 | 0.0149 | LGU76 | 0.2682 | 0.0155 | 0.0537 |
| YWLL08 | 0.182 | 0.0458 | 0.034 | YWLL08 | 0.4779 | 0.0205 | 0.0127 | YWLL08 | 0.1105 | 0.0113 | 0.0473 |
| YWLL44 | 0.118 | 0.0133 | 0.033 | YWLL44 | 0.1722 | 0.0133 | 0.0372 | YWLL44 | 0.2247 | 0.0153 | 0.0335 |
| YWLL59 | 0.0001 | 0.0001 | 0.118 | YWLL59 | 0 | 0 | 0.1255 | YWLL59 | 0.0373 | 0.0068 | 0.109 |
| VOLP03 | 0.434 | 0.0511 | 0.027 | VOLP03 | 0.9412 | 0.0091 | −0.0785 | VOLP03 | 0.1508 | 0.0113 | 0.0905 |
| Microsatellite | Ch’aku | Q’ara |
|---|---|---|
| GLM4 | * | 214 |
| LAB1 | 177 | 203 |
| LCA54 | * | 159, 171, 173, 179 |
| LCA65 | 202 | 200 |
| LCA77 | 264 | 256, 262, 278 |
| LCA82 | 126, 132 | * |
| LCA83 | 216 | * |
| LCA85 | 214 | * |
| LGU76 | 285 | 279 |
| YWLL08 | 181, 201 | 157, 189 |
| YWLL44 | 127 | 133, 143 |
| YWLL59 | 102, 114 | 138, 142, 148, 150 |
| VOLP03 | 153, 171 | 191 |
| Total | 15 | 20 |
| Microsatellites | FST |
|---|---|
| GLM4 | 0.005 |
| LAB1 | 0.009 |
| LCA54 | 0.033 |
| LCA65 | 0.001 |
| LCA77 | 0.005 |
| LCA82 | 0.017 |
| LCA83 | 0.022 |
| LCA85 | 0.009 |
| LGU76 | 0.009 |
| YWLL08 | 0.016 |
| YWLL44 | 0.005 |
| YWLL59 | 0.001 |
| VOLP03 | 0.002 |
| Average | 0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paredes, G.F.; Yalta-Macedo, C.E.; Gutierrez, G.A.; Veli-Rivera, E.A. Genetic Diversity and Population Structure of Llamas (Lama glama) from the Camelid Germplasm Bank—Quimsachata. Genes 2020, 11, 541. https://doi.org/10.3390/genes11050541
Paredes GF, Yalta-Macedo CE, Gutierrez GA, Veli-Rivera EA. Genetic Diversity and Population Structure of Llamas (Lama glama) from the Camelid Germplasm Bank—Quimsachata. Genes. 2020; 11(5):541. https://doi.org/10.3390/genes11050541
Chicago/Turabian StyleParedes, Gabriela F., Claudia E. Yalta-Macedo, Gustavo A. Gutierrez, and Eudosio A. Veli-Rivera. 2020. "Genetic Diversity and Population Structure of Llamas (Lama glama) from the Camelid Germplasm Bank—Quimsachata" Genes 11, no. 5: 541. https://doi.org/10.3390/genes11050541
APA StyleParedes, G. F., Yalta-Macedo, C. E., Gutierrez, G. A., & Veli-Rivera, E. A. (2020). Genetic Diversity and Population Structure of Llamas (Lama glama) from the Camelid Germplasm Bank—Quimsachata. Genes, 11(5), 541. https://doi.org/10.3390/genes11050541