Genome-Wide Diversity, Population Structure and Demographic History of Dromedaries in the Central Desert of Iran

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
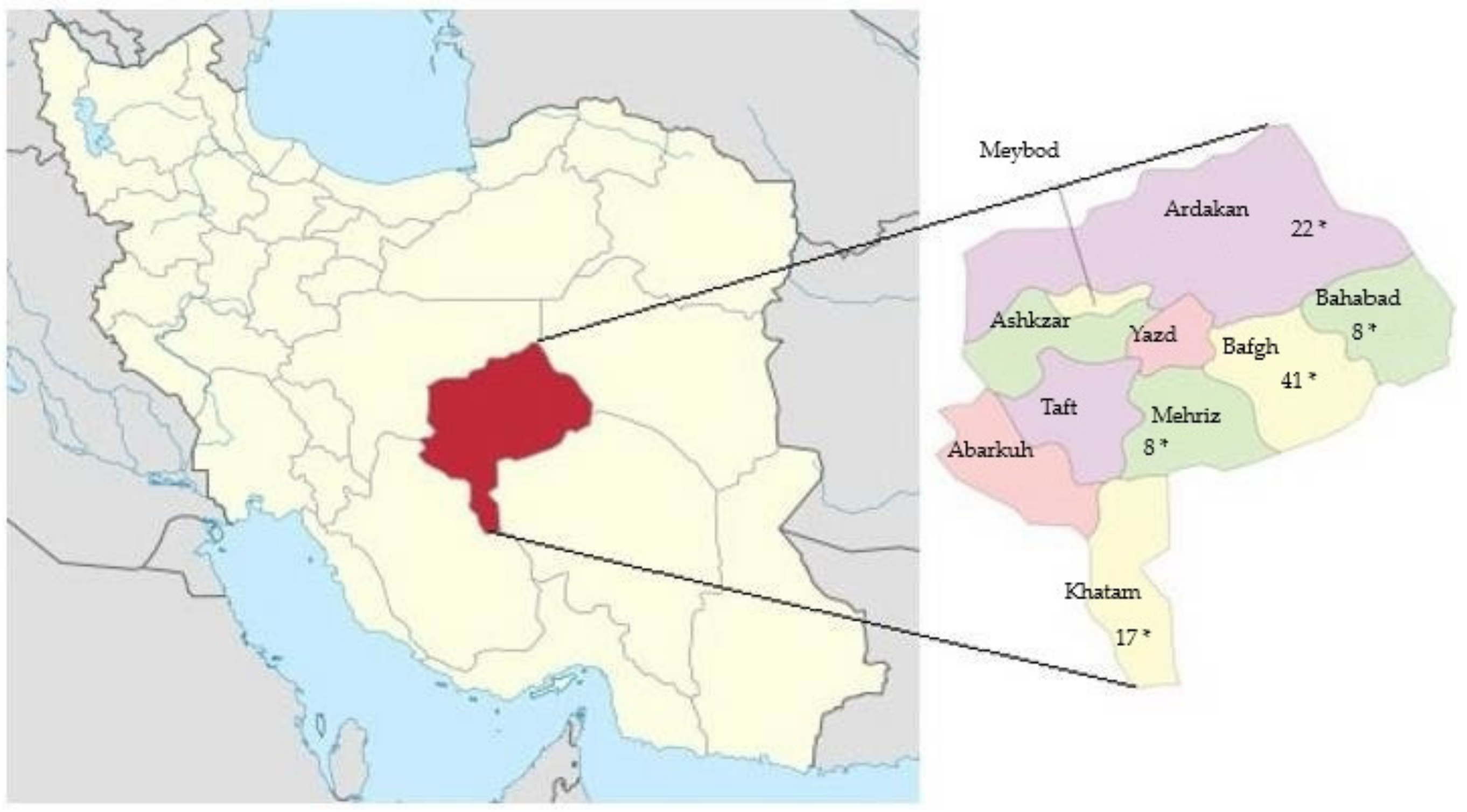
2.1. Animal Resources
2.2. Genotyping-by-Sequencing (GBS) Data
2.3. SNP Calling
2.4. Population Structure, Linkage Disequilibrium and Genome-Wide Diversity
2.5. Effective Population Size and Tajima’s D
3. Results
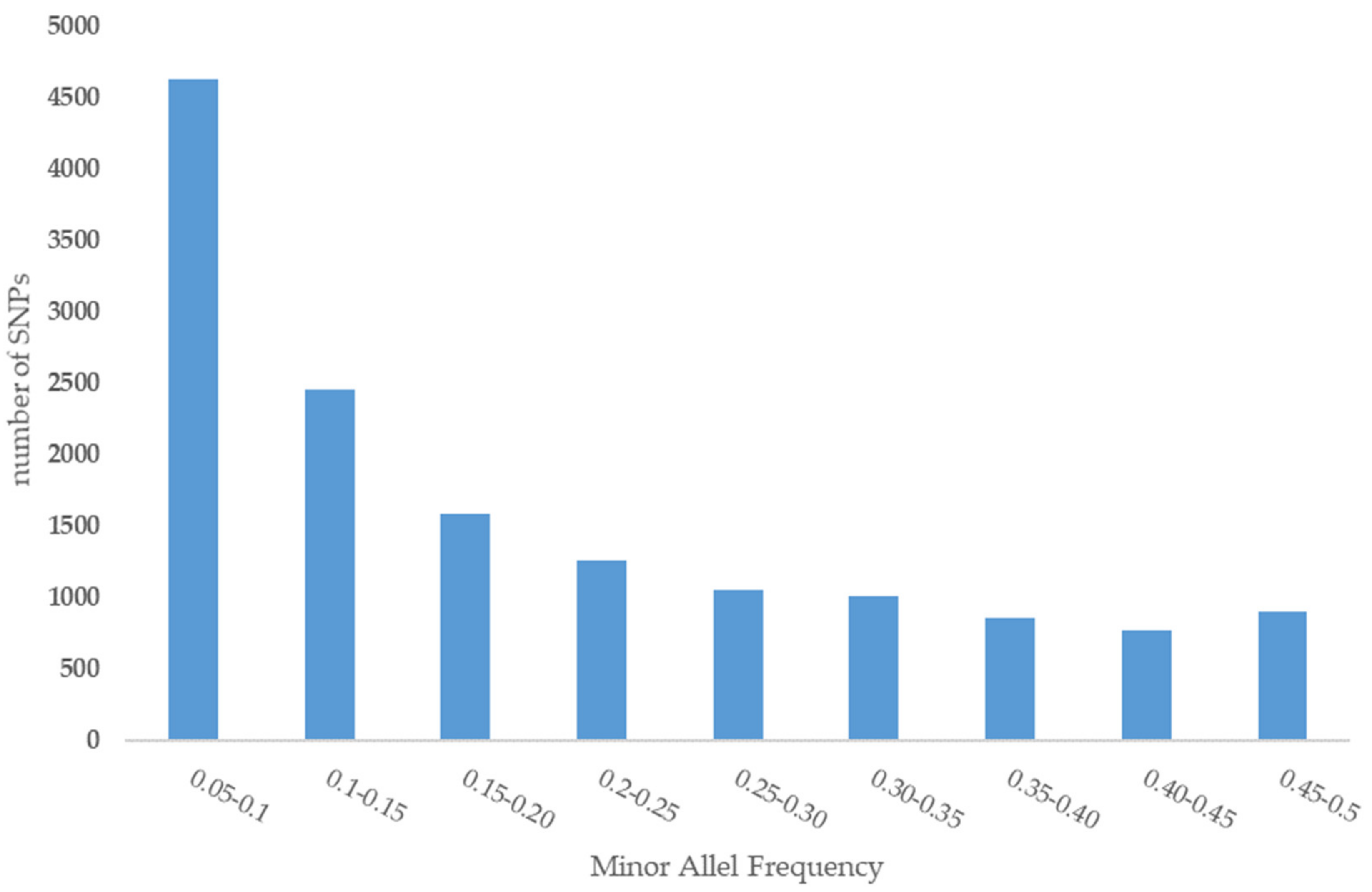
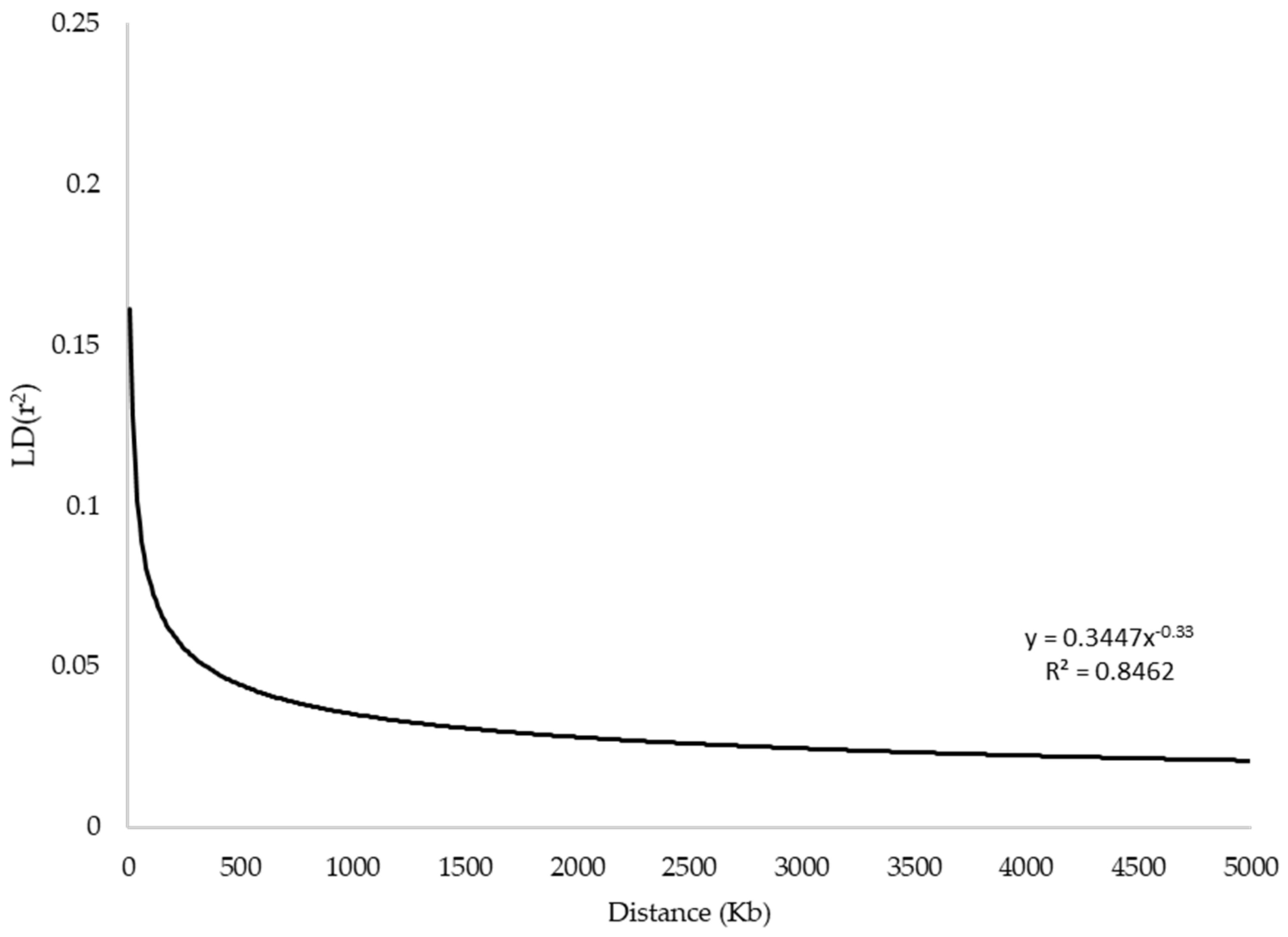
3.1. Genomic Diversity and Linkage Disequilibrium
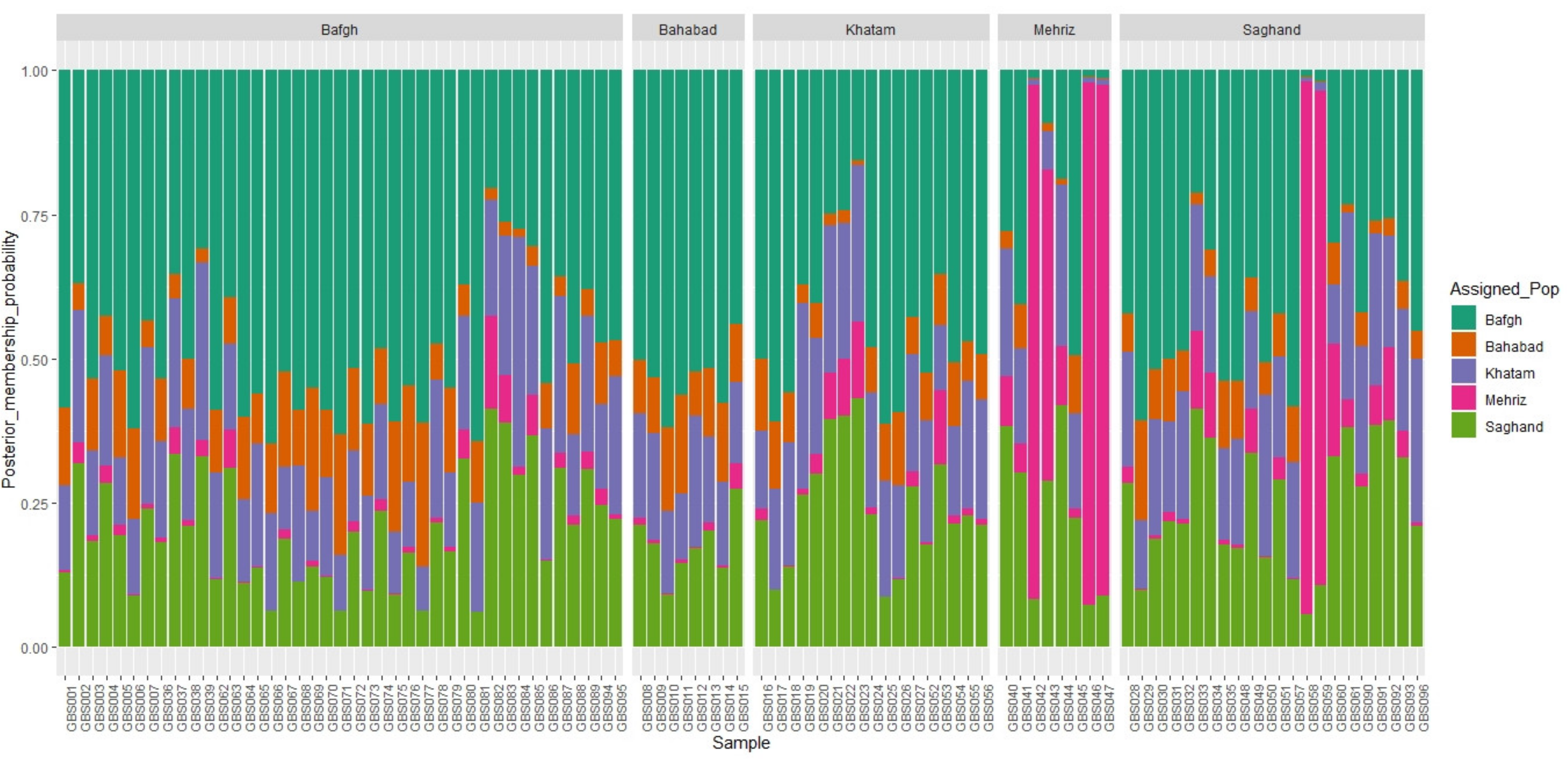
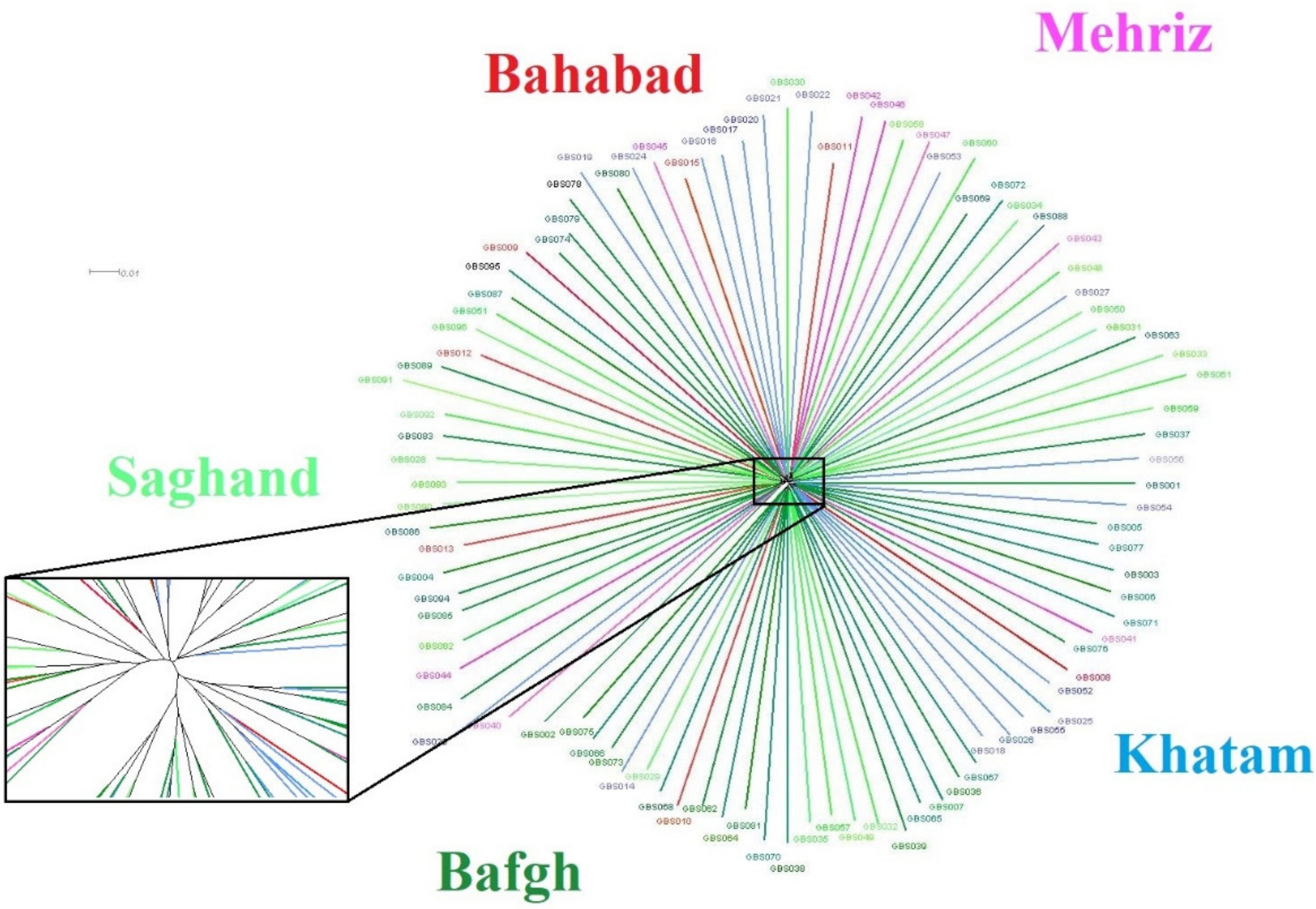
3.2. Cluster Analysis and Genomic Relationship
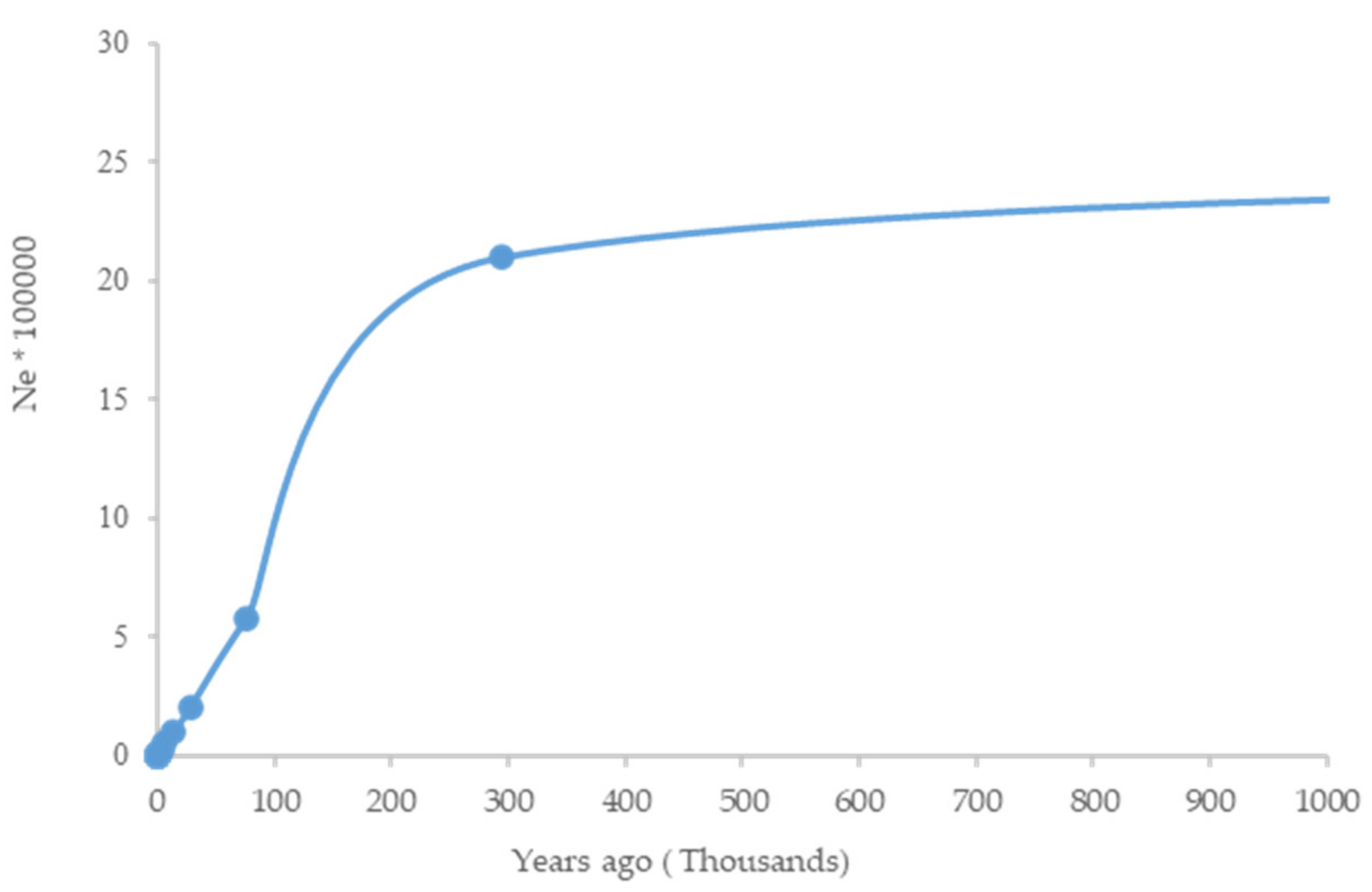
3.3. Demographic History (Effective Population Size)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilson, R.T. The Tropical Agriculturalist: Camels; CTA/Macmillan: Basingstoke, London, UK, 1998. [Google Scholar]
- Burger, P.A.; Ciani, E.; Faye, B. Old World camels in a modern world—A balancing act between conservation and genetic improvement. Anim. Genet. 2019, 50, 598–612. [Google Scholar] [CrossRef] [PubMed]
- Burren, A.; Signer-Hasler, H.; Neuditschko, M.; Tetens, J.; Kijas, J.; Drögemüller, C.; Flury, C. Fine-scale population structure analysis of seven local Swiss sheep breeds using genome-wide SNP data. Anim. Genet. Resour. 2014, 55, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Hashim, W.M.; Galal, M.Y.; Ali, A.M.; Khalafalla-Abdelmalik, A.I.; Hamid, S.A.; Mohamed, K.A. Dromedary camels in Sudan, types and sub types, distribution and movement. Int. J. Pharm. Res. Anal. 2015, 5, 8–12. [Google Scholar]
- Emami Meybodi, M.; Afshar, H.; Niyasari, A.; Arab, M.A.; Khaki, M. Identification and Conservation of Iranian Bactrian Camels. In International Information System for the Agricultural Sciences and Technology; Animal Science Research Institute: Karaj, Iran, 2008. [Google Scholar]
- Al-Hashmi, H. Combating Desertification in Asia, Africa and the Middle East; Springer Science and Business Media LLC: Cham, Switzerland, 2013. [Google Scholar]
- Lehane, S. The Iranian Water Crisis; Strategic Analysis Paper; Future Directions International Pty Ltd.: Perth, Australia, 2014. [Google Scholar]
- Almathen, F.; Charruau, P.; Mohandesan, E.; Mwacharo, J.M.; Orozco-terWengel, P.; Pitt, D.; Abdussamad, A.M.; Uerpmann, M.; Uerpmann, H.P.; De Cupere, B.; et al. Ancient and modern DNA reveal dynamics of domestication and cross-continental dispersal of the dromedary. Proc. Natl. Acad. Sci. USA 2016, 113, 6707–6712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mburu, D.N.; Ochieng, J.W.; Kuria, S.G.; Jianlin, H.; Kaufmann, B.; Rege, J.E.O.; Hanotte, O. Genetic diversity and relationships of indigenous Kenyan camel (Camelus dromedarius) populations: Implications for their classification. Anim. Genet. 2003, 34, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Babar, M.; Hussain, T.; Wajid, A.; Nawaz, A.; Nadeem, A.; Shah, S.A.; Shahid, M.A.; Ahmad, N.; Javed, K.; Abdullah, A. Mitochondrial cytochrome-b and D-loop sequence based genetic diversity in Mareecha and Bareela camel breeds of Pakistan. J. Anim. Plant Sci. 2015, 25, 591–594. [Google Scholar]
- Bahbahani, H.; Musa, H.H.; Wragg, D.; Shuiep, E.S.; Almathen, F.; Hanotte, O. Genome Diversity and Signatures of Selection for Production and Performance Traits in Dromedary Camels. Front. Genet. 2019, 10, 893. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Kim, J.-J. Effective Population Size and Signatures of Selection Using Bovine 50K SNP Chips in Korean Native Cattle (Hanwoo). Evol. Bioinform. 2015, 11, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Guang, X.; Al-Fageeh, M.B.; Cao, J.; Pan, S.; Zhou, H.; Zhang, L.; AbuTarboush, M.H.; Xing, Y.; Xie, Z.; et al. Camelid genomes reveal evolution and adaptation to desert environments. Nat. Commun. 2014, 5, 5188. [Google Scholar] [CrossRef] [Green Version]
- Elbers, J.P.; Rogers, M.F.; Perelman, P.L.; Proskuryakova, A.A.; Serdyukova, N.A.; Johnson, W.E.; Horin, P.; Corander, J.; Murphy, D.; Burger, P.A. Improving Illumina assemblies with Hi-C and long reads: An example with the North African dromedary. Mol. Ecol. Resour. 2019, 19, 1015–1026. [Google Scholar] [CrossRef]
- Ruvinskiy, D.; Larkin, D.M.; Farré, M. A Near Chromosome Assembly of the Dromedary Camel Genome. Front. Genet. 2019, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Khalkhali-Evrigh, R.; Hafezian, S.H.; Hedayat-Evrigh, N.; Farhadi, A.; Bakhtiarizadeh, M.R. Genetic variants analysis of three dromedary camels using whole genome sequencing data. PLoS ONE 2018, 13, e0204028. [Google Scholar] [CrossRef] [PubMed]
- Javanrouh, A.; Banabazi, M.H.; Esmaeilkhanian, S.; Amirinia, C.; Seyedabadi, H.R.; Emrani, H. Optimization on salting out method for DNA extraction from animal and poultry blood cells. In Proceedings of the 57th Ann Meet European Assoc Anim Prod, Antalya, Turkey, 17–20 September 2006. [Google Scholar]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Pembleton, L.W.; O I Cogan, N.; Forster, J.W. StAMPP: An R package for calculation of genetic differentiation and structure of mixed-ploidy level populations. Mol. Ecol. Resour. 2013, 13, 946–952. [Google Scholar] [CrossRef]
- Hill, W.G.; Robertson, A. Linkage disequilibrium in finite populations. Theor. Appl. Genet. 1968, 38, 226–231. [Google Scholar] [CrossRef]
- Zhang, C.; Dong, S.-S.; Xu, J.-Y.; He, W.-M.; Yang, T.-L. PopLDdecay: A fast and effective tool for linkage disequilibrium decay analysis based on variant call format files. Bioinformatics 2018, 35, 1786–1788. [Google Scholar] [CrossRef]
- Barbato, M.; Orozco-Terwengel, P.; Tapio, M.; Bruford, M.W. SNeP: A tool to estimate trends in recent effective population size trajectories using genome-wide SNP data. Front. Genet. 2015, 6, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Sved, J. Linkage disequilibrium and homozygosity of chromosome segments in finite populations. Theor. Popul. Boil. 1971, 2, 125–141. [Google Scholar] [CrossRef]
- Weir, B.S.; Hill, W.G. Effect of Mating Structure on Variation in Linkage Disequilibrium. Genetics 1980, 95, 477–488. [Google Scholar] [PubMed]
- Hayes, B.; Visscher, P.M.; McPartlan, H.C.; E Goddard, M. Novel Multilocus Measure of Linkage Disequilibrium to Estimate Past Effective Population Size. Genome Res. 2003, 13, 635–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohta, T.; Kimura, M. Linkage Disequilibrium between Two Segregating Nucleotide Sites under the Steady Flux of Mutations in a Finite Population. Genetics 1971, 68, 571–580. [Google Scholar] [PubMed]
- Zhang, Q.; Tyler-Smith, C.; Long, Q. An extended Tajima’s D neutrality test incorporating SNP calling and imputation uncertainties. Stat. Interface 2015, 8, 447–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, F.; Ming, L.; Si, R.; Yi, L.; He, J.; Ji, R. A Genome-Wide Association Study Identifies Quantitative Trait Loci Affecting Hematological Traits in Camelus bactrianus. Animals 2020, 10, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Abri, M.; Holl, H.M.; Miller, D.; Abdalla, S.; Shykind, B.; Malek, J.; Mohamoud, Y.; Pasha, K.; Khalili, A.; Antczak, D.F.; et al. Selection of SNP markers for a dromedary camel genotyping array. In Proceedings of the 36th International Society for Animal Genetics Conference, Dublin, Ireland, 16–21 July 2017. [Google Scholar]
- King, R.C.; Stansfield, W.D.; Mulligan, P.K. A Dictionary of Genetics; Oxford University Press: New York, NY, USA, 2006. [Google Scholar]
- Nosrati, M. The Pattern of Linkage Disequilibrium in Livestock Genome. Iran. J. Appl. Anim. Sci. 2017, 7, 1–7. [Google Scholar]
- Mastrangelo, S.; Di Gerlando, R.; Tolone, M.; Tortorici, L.; Sardina, M.T.; Portolano, B. International Sheep Genomics Consortium Genome wide linkage disequilibrium and genetic structure in Sicilian dairy sheep breeds. BMC Genet. 2014, 15, 108. [Google Scholar] [CrossRef] [Green Version]
- Corbin, L.; Blott, S.; Swinburne, J.E.; Vaudin, M.; Bishop, S.C.; Woolliams, J.A. Linkage disequilibrium and historical effective population size in the Thoroughbred horse. Anim. Genet. 2010, 41, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Uimari, P.; Tapio, M. Extent of linkage disequilibrium and effective population size in Finnish Landrace and Finnish Yorkshire pig breeds1. J. Anim. Sci. 2011, 89, 609–614. [Google Scholar] [CrossRef]
- Zhan, B.; Fadista, J.; Thomsen, B.; Hedegaard, J.; Panitz, F.; Bendixen, C. Global assessment of genomic variation in cattle by genome resequencing and high-throughput genotyping. BMC Genom. 2011, 12, 557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorjanc, G.; A Cleveland, M.; Houston, R.D.; Hickey, J.M. Potential of genotyping-by-sequencing for genomic selection in livestock populations. Genet. Sel. Evol. 2015, 47, 12. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chr | Number of SNPs | Mean MAF | Chr | Number of SNPs | Mean MAF |
|---|---|---|---|---|---|
| 1 | 150 | 0.23 | 19 | 1655 | 0.18 |
| 2 | 412 | 0.22 | 20 | 51 | 0.27 |
| 3 | 78 | 0.25 | 21 | 305 | 0.21 |
| 4 | 113 | 0.21 | 22 | 20 | 0.33 |
| 5 | 125 | 0.20 | 23 | 16 | 0.21 |
| 6 | 413 | 0.22 | 24 | 84 | 0.21 |
| 7 | 372 | 0.22 | 25 | 1113 | 0.19 |
| 8 | 281 | 0.22 | 26 | 51 | 0.24 |
| 9 | 1829 | 0.19 | 27 | 45 | 0.19 |
| 10 | 353 | 0.27 | 28 | 30 | 0.17 |
| 11 | 1442 | 0.20 | 29 | 84 | 0.25 |
| 12 | 231 | 0.20 | 30 | 101 | 0.21 |
| 13 | 167 | 0.24 | 31 | 195 | 0.20 |
| 14 | 1062 | 0.19 | 32 | 22 | 0.25 |
| 15 | 150 | 0.25 | 33 | 144 | 0.22 |
| 16 | 378 | 0.19 | 34 | 55 | 0.16 |
| 17 | 379 | 0.18 | 35 | 376 | 0.20 |
| 18 | 1301 | 0.18 | 36 | 194 | 0.19 |
| X | 745 | 0.18 |
| Distance | N | Mean | SD | % r2 > 0.2 | % r2 > 0.3 |
|---|---|---|---|---|---|
| 0–40 kb | 7,020,569 | 0.0899 | 0.234 | 98.02 | 94.50 |
| 40–60 kb | 613,045 | 0.0199 | 0.034 | 0.15 | 0.52 |
| 60–100 kb | 735,773 | 0.0208 | 0.039 | 0.36 | 0.77 |
| 100–250 kb | 1,082,002 | 0.0177 | 0.030 | 0.13 | 0.66 |
| 250–500 kb | 789,072 | 0.0198 | 0.035 | 0.28 | 0.76 |
| 0.5–1 Mb | 650,753 | 0.0175 | 0.030 | 0.14 | 0.30 |
| 1–2 Mb | 436,200 | 0.0181 | 0.033 | 0.10 | 0.28 |
| 2–5 Mb | 808,181 | 0.0194 | 0.0351 | 0.33 | 0.59 |
| 5–10 Mb | 719,173 | 0.0188 | 0.033 | 0.21 | 0.49 |
| 10–20 Mb | 809,000 | 0.017 | 0.029 | 0.08 | 0.45 |
| 20–50 Mb | 841,882 | 0.0187 | 0.031 | 0.10 | 0.58 |
| 50–92 Mb | 204,580 | 0.0164 | 0.032 | 0.11 | 0.10 |
| Bafgh | Bahabad | Khatam | Saghand | Mehriz | |
|---|---|---|---|---|---|
| Bafgh | 0 | - | - | - | - |
| Bahabad | 0.003 | 0 | - | - | - |
| Khatam | 0.005 | 0.004 | 0 | - | - |
| Saghand | 0.004 | 0.002 | 0.004 | 0 | - |
| Mehriz | 0.011 | 0.010 | 0.011 | 0.008 | 0 |
| Generations Ago | Ne | Dist. | r2 | r2SD |
|---|---|---|---|---|
| 1 | 89 | 18,735,020 | 0.010774 | 0.015864 |
| 2 | 108 | 16,440,742 | 0.010643 | 0.015069 |
| 2 | 136 | 14,311,335 | 0.010036 | 0.013526 |
| 3 | 155 | 11,950,567 | 0.011016 | 0.016526 |
| 4 | 186 | 10,407,033 | 0.010833 | 0.015233 |
| 4 | 220 | 9,096,109 | 0.010719 | 0.014915 |
| 6 | 282 | 7,360,504 | 0.010603 | 0.015451 |
| 7 | 324 | 6,387,962 | 0.010813 | 0.014915 |
| 8 | 396 | 5,401,410 | 0.010624 | 0.014839 |
| 10 | 487 | 4,410,026 | 0.010759 | 0.015362 |
| 13 | 618 | 3,561,115 | 0.010637 | 0.015982 |
| 16 | 799 | 2,844,204 | 0.010428 | 0.01519 |
| 21 | 1001 | 2,241,547 | 0.010652 | 0.014796 |
| 25 | 1237 | 1,901,651 | 0.010217 | 0.014037 |
| 35 | 1694 | 1,375,758 | 0.010399 | 0.014926 |
| 44 | 2116 | 1,097,526 | 0.010477 | 0.014596 |
| 60 | 2883 | 811,669 | 0.010442 | 0.015188 |
| 80 | 3680 | 617,213 | 0.010787 | 0.015267 |
| 113 | 5344 | 438785 | 0.01048 | 0.01491 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bitaraf Sani, M.; Harofte, J.Z.; Bitaraf, A.; Esmaeilkhanian, S.; Banabazi, M.H.; Salim, N.; Teimoori, A.; Shafei Naderi, A.; Faghihi, M.A.; Burger, P.A.; et al. Genome-Wide Diversity, Population Structure and Demographic History of Dromedaries in the Central Desert of Iran. Genes 2020, 11, 599. https://doi.org/10.3390/genes11060599
Bitaraf Sani M, Harofte JZ, Bitaraf A, Esmaeilkhanian S, Banabazi MH, Salim N, Teimoori A, Shafei Naderi A, Faghihi MA, Burger PA, et al. Genome-Wide Diversity, Population Structure and Demographic History of Dromedaries in the Central Desert of Iran. Genes. 2020; 11(6):599. https://doi.org/10.3390/genes11060599
Chicago/Turabian StyleBitaraf Sani, Morteza, Javad Zare Harofte, Ahmad Bitaraf, Saeid Esmaeilkhanian, Mohammad Hossein Banabazi, Nader Salim, Abbas Teimoori, Ali Shafei Naderi, Mohammad Ali Faghihi, Pamela Anna Burger, and et al. 2020. "Genome-Wide Diversity, Population Structure and Demographic History of Dromedaries in the Central Desert of Iran" Genes 11, no. 6: 599. https://doi.org/10.3390/genes11060599
APA StyleBitaraf Sani, M., Harofte, J. Z., Bitaraf, A., Esmaeilkhanian, S., Banabazi, M. H., Salim, N., Teimoori, A., Shafei Naderi, A., Faghihi, M. A., Burger, P. A., Silawi, M., & Taghipour Sheshdeh, A. (2020). Genome-Wide Diversity, Population Structure and Demographic History of Dromedaries in the Central Desert of Iran. Genes, 11(6), 599. https://doi.org/10.3390/genes11060599