Cytogenetic Characterization of Seven Novel satDNA Markers in Two Species of Spined Loaches (Cobitis) and Their Clonal Hybrids

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Specimens and Ethics Statement
2.2. Bioinformatics Analyses
2.3. Preparation of Mitotic Chromosomes
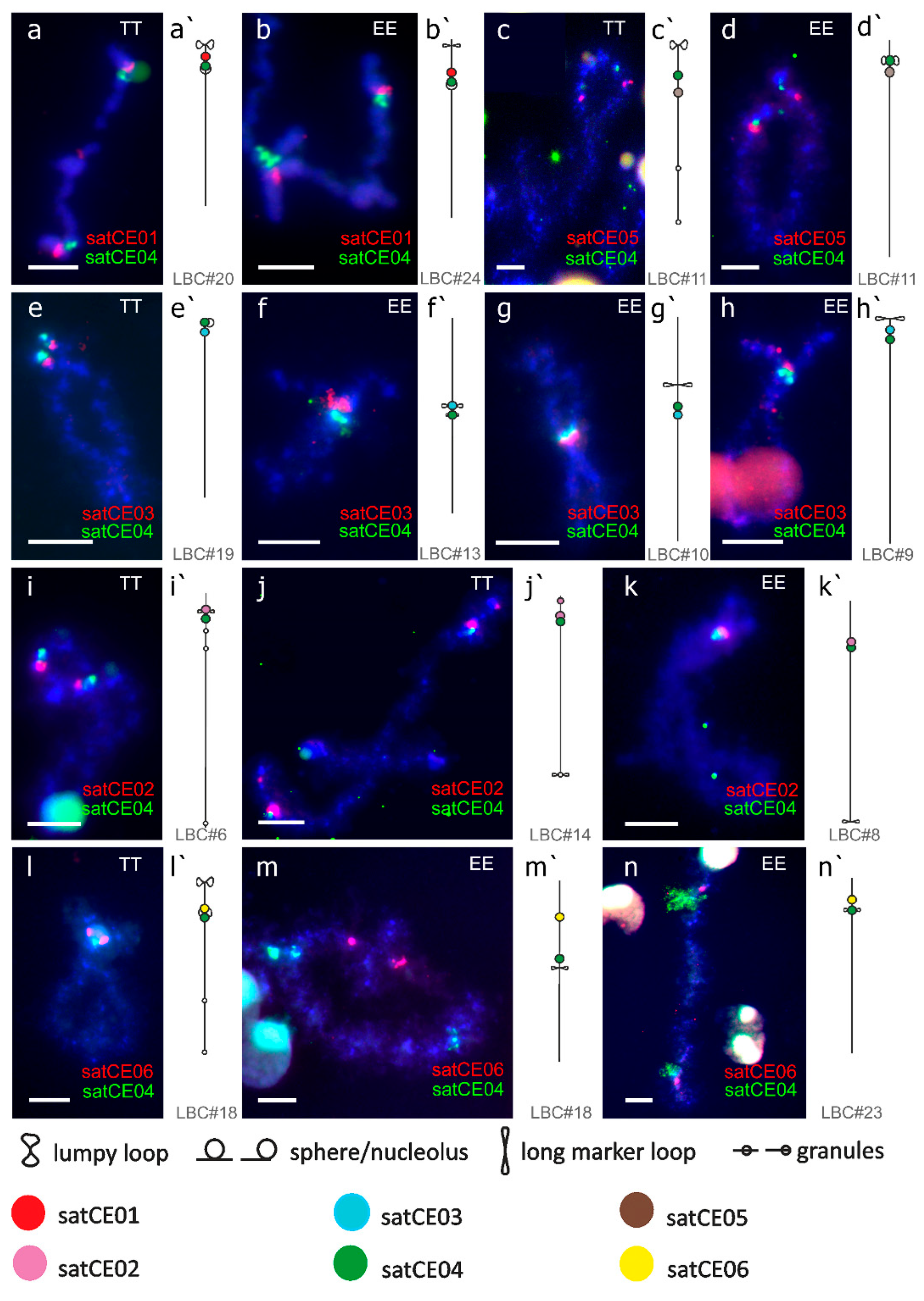
2.4. Preparation of Lampbrush Chromosome
2.5. DNA Probes and Fluorescent In Situ Hybridization (FISH)
2.6. Image Processing
3. Results
3.1. Identification and Mapping of satDNA Repeats
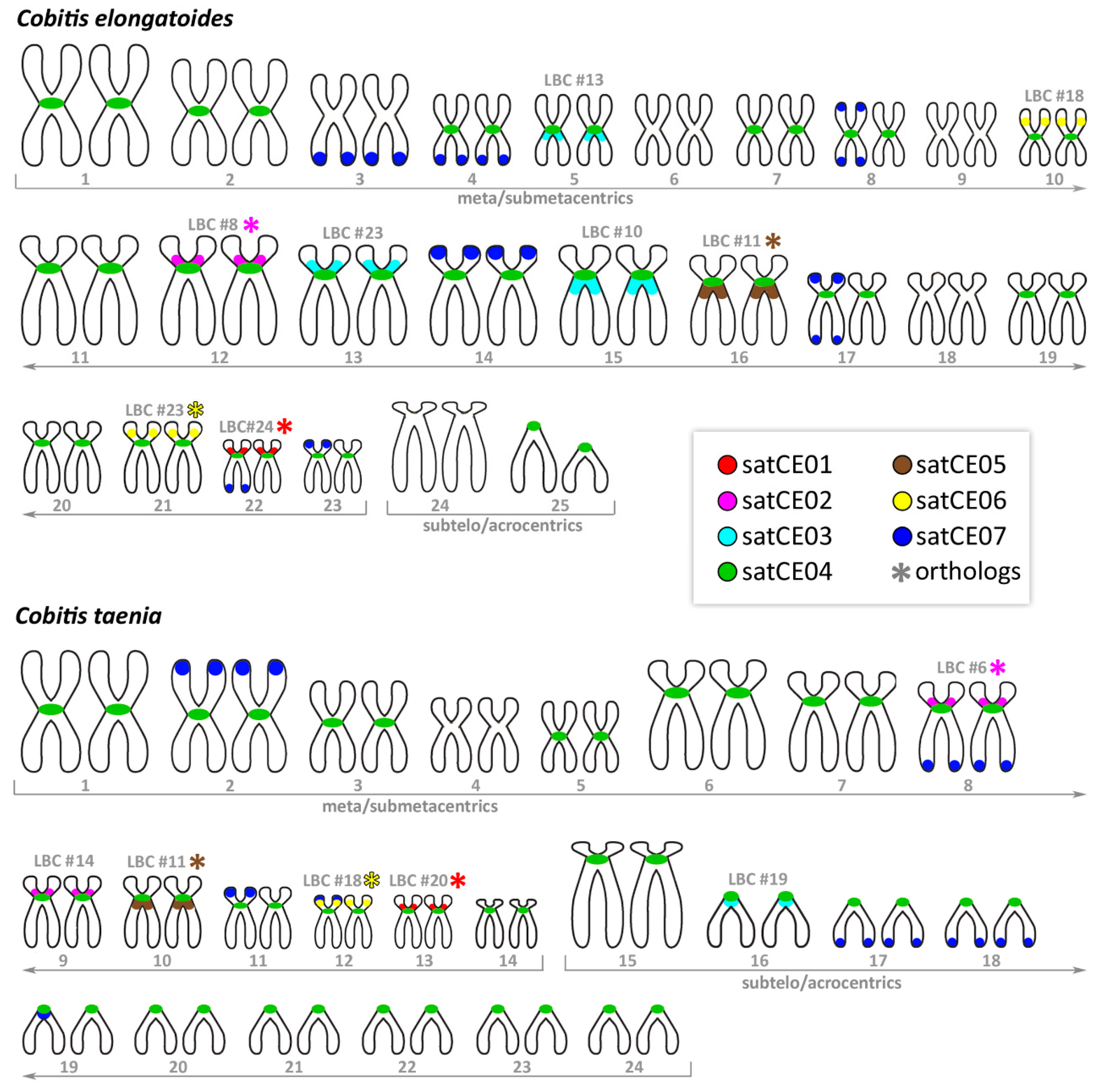
3.2. Karyotypes
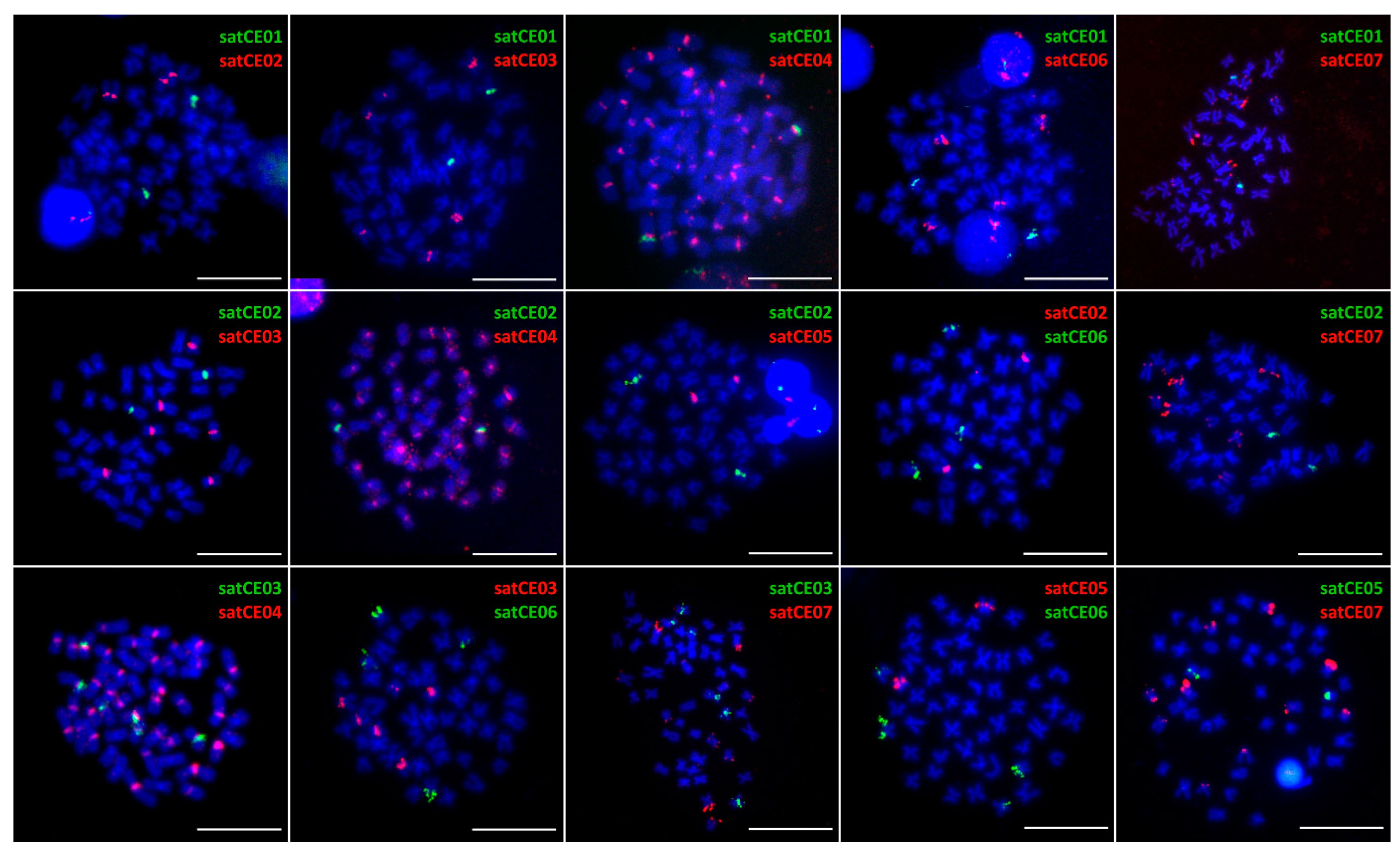
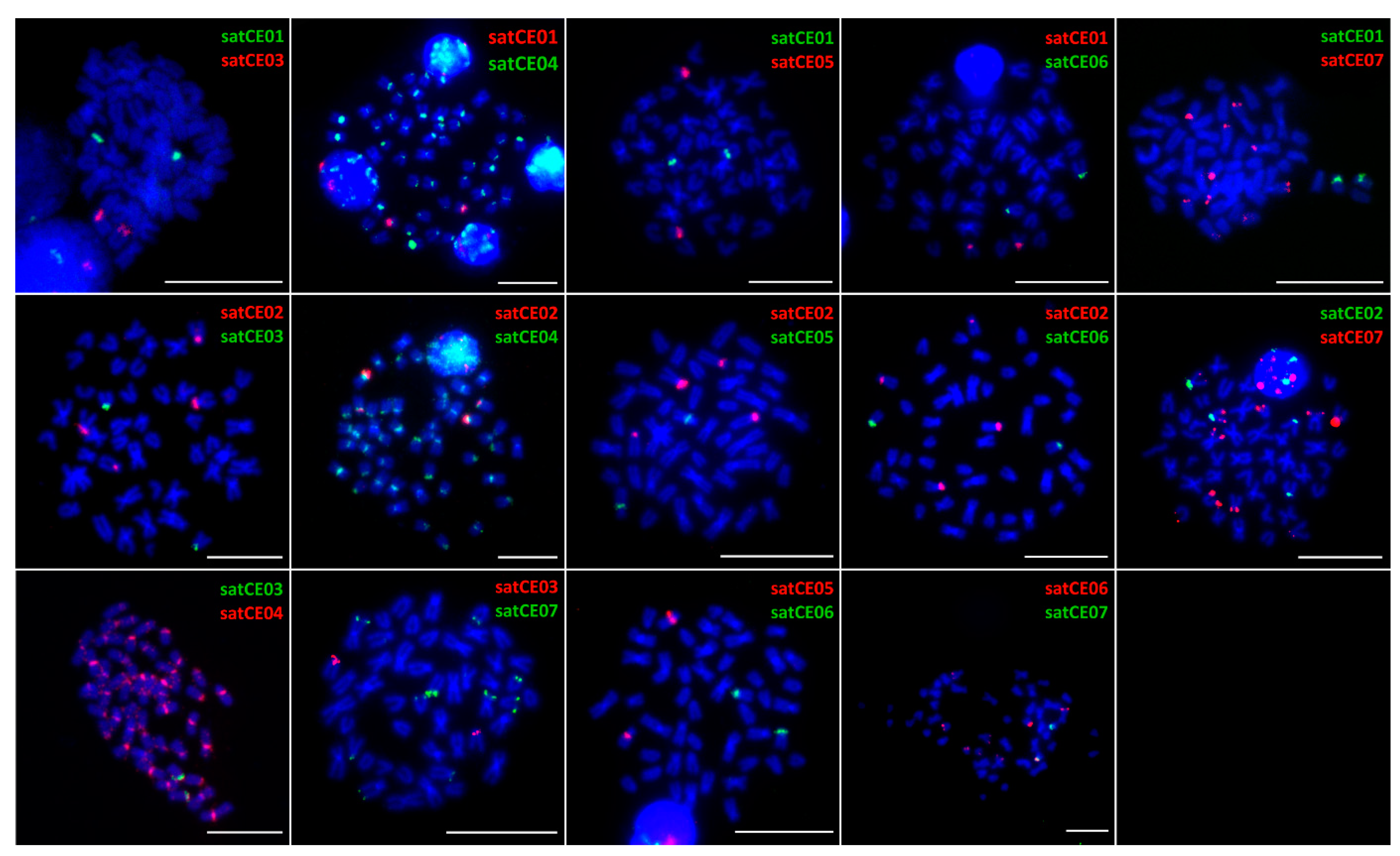
3.3. Mapping of satDNA Markers
4. Discussion
4.1. Karyotype Evolution within Spined loaches of the Central European Hybrid Complex
4.2. Interspecific Variability of Satellite DNAs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bullini, L. Origin and evolution of animal hybrid species. Trends Ecol. Evol. 1994, 9, 422–426. [Google Scholar] [CrossRef]
- Abbott, R.; Albach, D.; Ansell, S.; Arntzen, J.W.; Baird, S.J.E.; Bierne, N.; Boughman, J.; Brelsford, A.; Buerkle, C.A.; Buggs, R.; et al. Hybridization and speciation. J. Evol. Biol. 2013, 26, 229–246. [Google Scholar] [CrossRef] [Green Version]
- Janko, K.; Pačes, J.; Wilkinson-Herbots, H.; Costa, R.J.; Roslein, J.; Drozd, P.; Iakovenko, N.; Rídl, J.; Hroudová, M.; Kočí, J.; et al. Hybrid asexuality as a primary postzygotic barrier between nascent species: On the interconnection between asexuality, hybridization and speciation. Mol. Ecol. 2018, 27, 248–263. [Google Scholar] [CrossRef] [Green Version]
- Choleva, L.; Janko, K.; Gelas, K.D.; Bohlen, J.; Šlechtová, V.; Rábová, M.; Ráb, P. Synthesis of Clonality and Polyploidy in Vertebrate Animals by Hybridization between Two Sexual Species. Evolution 2012, 66, 2191–2203. [Google Scholar] [CrossRef] [PubMed]
- Stenberg, P.; Saura, A. Meiosis and Its Deviations in Polyploid Animals. Cytogenet. Genome Res. 2013, 140, 185–203. [Google Scholar] [CrossRef] [PubMed]
- Ohara, K.; Dong, S.; Taniguchi, N. High Proportion of Heterozygotes in Microsatellite DNA Loci of Wild Clonal Silver Crucian Carp, Carassius langsdorfii. Zool. Sci. 1999, 16, 909–913. [Google Scholar] [CrossRef]
- Morishima, K.; Horie, S.; Yamaha, E.; Arai, K. A Cryptic Clonal Line of the Loach Misgurnus anguillicaudatus (Teleostei: Cobitidae) Evidenced by Induced Gynogenesis, Interspecific Hybridization, Microsatellite Genotyping and Multilocus DNA Fingerprinting. Zool. Sci. 2002, 19, 565–575. [Google Scholar] [CrossRef] [Green Version]
- Janko, K.; Culling, M.A.; Ráb, P.; Kotlík, P. Ice age cloning – comparison of the Quaternary evolutionary histories of sexual and clonal forms of spiny loaches (Cobitis; Teleostei) using the analysis of mitochondrial DNA variation. Mol. Ecol. 2005, 14, 2991–3004. [Google Scholar] [CrossRef]
- Janko, K.; Flajšhans, M.; Choleva, L.; Bohlen, J.; ŠLechtová, V.; Rábová, M.; Lajbner, Z.; ŠLechta, V.; Ivanova, P.; Dobrovolov, I.; et al. Diversity of European spined loaches (genus Cobitis L.): An update of the geographic distribution of the Cobitis taenia hybrid complex with a description of new molecular tools for species and hybrid determination. J. Fish Biol. 2007, 71, 387–408. [Google Scholar] [CrossRef]
- Tóth, B.; Várkonyi, E.; Hidas, A.; Meleg, E.E.; Váradi, L. Genetic analysis of offspring from intra- and interspecific crosses of Carassius auratus gibelio by chromosome and RAPD analysis. J. Fish Biol. 2005, 66, 784–797. [Google Scholar] [CrossRef]
- Angers, B.; Schlosser, I.J. The origin of Phoxinus eos-neogaeus unisexual hybrids. Mol. Ecol. 2007, 16, 4562–4571. [Google Scholar] [CrossRef] [PubMed]
- Collares-Pereira, M.J.; Coelho, M.M. Reconfirming the hybrid origin and generic status of the Iberian cyprinid complex Squalius alburnoides. J. Fish Biol. 2010, 76, 707–715. [Google Scholar] [CrossRef]
- Kottelat, M. Conspectus Cobitidum: An Inventory of the Loaches of the World (Teleostei: Syprinformes: Cobitoidei); National University of Singapore: Singapore, 2012. [Google Scholar]
- Vasil’ev, V.P.; Lebedeva, E.B.; Vasil’eva, E.D.; Ryskov, A.P. Monoclonal and de novo arising tetraploid forms of the genus Cobitis (Cobitidae) from different clonal-bisexual complexes. Dokl. Biol. Sci. 2007, 416, 360. [Google Scholar] [CrossRef]
- Janko, K.; Kotusz, J.; Gelas, K.D.; Šlechtová, V.; Opoldusová, Z.; Drozd, P.; Choleva, L.; Popiołek, M.; Baláž, M. Dynamic Formation of Asexual Diploid and Polyploid Lineages: Multilocus Analysis of Cobitis Reveals the Mechanisms Maintaining the Diversity of Clones. PLoS ONE 2012, 7, e45384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choleva, L.; Musilova, Z.; Kohoutova-Sediva, A.; Paces, J.; Rab, P.; Janko, K. Distinguishing between Incomplete Lineage Sorting and Genomic Introgressions: Complete Fixation of Allospecific Mitochondrial DNA in a Sexually Reproducing Fish (Cobitis; Teleostei), despite Clonal Reproduction of Hybrids. PLoS ONE 2014, 9, e80641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotusz, J. Intra- and interpopulation morphological variability in diploid and varied-ploidy Cobitis from Poland. Folia Zool. 2000, 49, 219–226. [Google Scholar]
- Bohlen, J.; Ráb, P. Species and hybrid richness in spined loaches of the genus Cobitis (Teleostei: Cobitidae), with a checklist of European forms and suggestions for conservation. J. Fish Biol. 2001, 59, 75–89. [Google Scholar] [CrossRef]
- Grabowska, A.I.; Boroń, A.; Kirtiklis, L.; Spóz, A.; Juchno, D.; Kotusz, J. Chromosomal inheritance of parental rDNAs distribution pattern detected by FISH in diploid F1 hybrid progeny of Cobitis (Teleostei, Cobitidae) species has non-Mendelian character. J. Fish Biol. 2020, 96, 261–273. [Google Scholar] [CrossRef]
- Majtánová, Z.; Choleva, L.; Symonová, R.; Ráb, P.; Kotusz, J.; Pekárik, L.; Janko, K. Asexual Reproduction Does Not Apparently Increase the Rate of Chromosomal Evolution: Karyotype Stability in Diploid and Triploid Clonal Hybrid Fish (Cobitis, Cypriniformes, Teleostei). PLoS ONE 2016, 11, e0146872. [Google Scholar] [CrossRef] [Green Version]
- Boroń, A. Karyotypes and cytogenetic diversity of the genus Cobitis (Pisces, Cobitidae) in Poland: A review. Cytogenetic evidence for a hybrid origin of some Cobitis triploids. Folia Biol. (Krakow) 2003, 51, 49–54. [Google Scholar]
- Rábová, M.; Pelikánová, Š.; Choleva, L.; Ráb, P. Cytogenetics of bisexual species and their asexual hybrid clones in European spined loaches, genus Cobitis. II. Mapping of telomeric (TTAGGG)n sequences and DAPI-positive heterochromatins in four parental species. In ECI XII European Congress of Ichthyology, Book of Abstracts; Wiley: Cavtat, Croatia, 2007. [Google Scholar]
- Cioffi, M.B.; Bertollo, L.A.C. Chromosomal Distribution and Evolution of Repetitive DNAs in Fish. Repetit. DNA 2012, 7, 197–221. [Google Scholar] [CrossRef]
- Mehrotra, S.; Goyal, V. Repetitive Sequences in Plant Nuclear DNA: Types, Distribution, Evolution and Function. Genom. Proteom. Bioinform. 2014, 12, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Ramos, M. Satellite DNA: An evolving topic. Genes 2017, 8, 230. [Google Scholar] [CrossRef]
- Pezer, Ž.; Brajković, J.; Feliciello, I.; Ugarković, Đ. Transcription of Satellite DNAs in Insects. Prog. Mol. Subcell. Biol. 2011, 51, 161–178. [Google Scholar] [PubMed]
- Biscotti, M.A.; Olmo, E.; Heslop-Harrison, J.S. (Pat) Repetitive DNA in eukaryotic genomes. Chromosom. Res. 2015, 23, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.; Meles, S.; Escudeiro, A.; Mendes-da-Silva, A.; Adega, F.; Chaves, R. Satellite non-coding RNAs: The emerging players in cells, cellular pathways and cancer. Chromosom. Res. 2015, 23, 479–493. [Google Scholar] [CrossRef]
- Hemleben, V.; Kovarik, A.; Torres-Ruiz, R.A.; Volkov, R.A.; Beridze, T. Plant highly repeated satellite DNA: Molecular evolution, distribution and use for identification of hybrids. Syst. Biodivers. 2007, 5, 277–289. [Google Scholar] [CrossRef]
- Rayburn, A.L.; Gill, B.S. Isolation of a D-genome specific repeated DNA sequence from Aegilops squarrosa. Plant Mol. Biol. Rep. 1986, 4, 102–109. [Google Scholar] [CrossRef]
- Lorite, P.; Muñoz-López, M.; Carrillo, J.A.; Sanllorente, O.; Vela, J.; Mora, P.; Tinaut, A.; Torres, M.I.; Palomeque, T. Concerted evolution, a slow process for ant satellite DNA: Study of the satellite DNA in the Aphaenogaster genus (Hymenoptera, Formicidae). Org. Divers. Evol. 2017, 17, 595–606. [Google Scholar] [CrossRef]
- Dover, G.A. Molecular drive in multigene families: How biological novelties arise, spread and are assimilated. Trends Genet. 1986, 2, 159–165. [Google Scholar] [CrossRef]
- López-Flores, I.; Garrido-Ramos, M.A. The Repetitive DNA Content of Eukaryotic Genomes. Repetit. DNA 2012, 7, 1–28. [Google Scholar] [CrossRef]
- Novák, P.; Ávila Robledillo, L.; Koblížková, A.; Vrbová, I.; Neumann, P.; Macas, J. TAREAN: A computational tool for identification and characterization of satellite DNA from unassembled short reads. Nucleic Acids Res. 2017, 45, e111. [Google Scholar] [CrossRef] [PubMed]
- Janko, K.; Bohlen, J.; Lamatsch, D.; Flajšhans, M.; Epplen, J.T.; Ráb, P.; Kotlík, P.; Šlechtová, V. The gynogenetic reproduction of diploid and triploid hybrid spined loaches (Cobitis: Teleostei), and their ability to establish successful clonal lineages—on the evolution of polyploidy in asexual vertebrates. Genetica 2007, 131, 185–194. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple Sequence Alignment Using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2003, 00, 2–3. [Google Scholar] [CrossRef] [PubMed]
- R: A Language and Environment for Statistical Computing; Version 3.5.2; R Foundation for Statistical Computing: Vienna, Austria, 2018.
- Charif, D.; Lobry, J.R. SeqinR 1.0-2: A Contributed Package to the R Project for Statistical Computing Devoted to Biological Sequences Retrieval and Analysis. In Structural Approaches to Sequence Evolution; (Biological and Medical Physics, Biomedical Engineering); Bastolla, U., Porto, M., Roman, H.E., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 207–232. ISBN 978-3-540-35306-5. [Google Scholar]
- Bushnell, B. BBTools Software Package. Available online: http://sourceforge.net/projects/bbmap (accessed on 16 October 2019).
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Novák, P.; Neumann, P.; Pech, J.; Steinhaisl, J.; Macas, J. RepeatExplorer: A Galaxy-based web server for genome-wide characterization of eukaryotic repetitive elements from next-generation sequence reads. Bioinformatics 2013, 29, 792–793. [Google Scholar] [CrossRef] [Green Version]
- Treangen, T.J.; Salzberg, S.L. Repetitive DNA and next-generation sequencing: Computational challenges and solutions. Nat. Rev. Genet. 2012, 13, 36–46. [Google Scholar] [CrossRef]
- Mori, H.; Evans-Yamamoto, D.; Ishiguro, S.; Tomita, M.; Yachie, N. Fast and global detection of periodic sequence repeats in large genomic resources. Nucleic Acids Res. 2019, 47, e8. [Google Scholar] [CrossRef]
- Ráb, P. Cold-blooded vertebrates. Method. Chromosom. Anal. 1988, 115–124. [Google Scholar]
- Völker, M.; Ráb, P. Direct chromosome preparation from regenerating fish fin tissue. In Fish Cytogenetic Techniques: Ray-Fin Fishes and Chondrichthyans; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Callan, H.G. Lampbrush Chromosomes (Molecular Biology, Biochemistry and Biophysics Molekularbiologie, Biochemie und Biophysik); Springer: Berlin/Heidelberg, Germany, 1986; ISBN 978-3-642-82794-5. [Google Scholar]
- Gall, J.G.; Murphy, C.; Callan, H.G.; Wu, Z. Chapter 8 Lampbrush Chromosomes. In Methods in Cell Biology; Kay, B.K., Peng, H.B., Eds.; Academic Press: Cambridge, MA, USA, 1991; pp. 149–166. [Google Scholar]
- Dedukh, D.; Majtánová, Z.; Marta, A.; Pšenička, M.; Kotusz, J.; Klíma, J.; Juchno, D.; Boron, A.; Janko, K. Parthenogenesis as a solution to hybrid sterility: The mechanistic basis of meiotic distortions in clonal and sterile hybrids. bioRxiv 2019, 663112. [Google Scholar] [CrossRef] [Green Version]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Boron, A.; Kotusz, J. A preliminary report and karyotype of a new Cobitis species in the ichthyofauna of Poland. Cytobios 1999, 98, 59–64. [Google Scholar]
- Kohn, M.; Högel, J.; Vogel, W.; Minich, P.; Kehrer-Sawatzki, H.; Graves, J.A.M.; Hameister, H. Reconstruction of a 450-My-old ancestral vertebrate protokaryotype. Trends Genet. 2006, 22, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Mank, J.E.; Avise, J.C. Phylogenetic conservation of chromosome numbers in Actinopterygiian fishes. Genetica 2006, 127, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Arai, R. Fish Karyotypes: A Check List; Springer Science & Business Media: Tokyo, Japan, 2011; ISBN 978-4-431-53877-6. [Google Scholar]
- Janko, K.; Vasil’ev, V.P.; Ráb, P.; Rábová, M.; Šlechtová, V.; Vasil’eva, E.D. Genetic and morphological analyses of 50-chromosome spined loaches (Cobitis, Cobitidae, Pisces) from the Black Sea basin that are morphologically similar to C. taenia, with the description of a new species. Folia Zool. Praha 2005, 54, 405–420. [Google Scholar]
- Slijepcevic, P. Telomeres and mechanisms of Robertsonian fusion. Chromosoma 1998, 107, 136–140. [Google Scholar] [CrossRef]
- Bolzán, A.D. Interstitial telomeric sequences in vertebrate chromosomes: Origin, function, instability and evolution. Mutat. Res. Rev. Mutat. Res. 2017, 773, 51–65. [Google Scholar] [CrossRef]
- Lin, K.W.; Yan, J. Endings in the middle: Current knowledge of interstitial telomeric sequences. Mutat. Res./Rev. Mutat. Res. 2008, 658, 95–110. [Google Scholar] [CrossRef]
- Callan, H.G.; Lloyd, L.; Waddington, C.H. Lampbrush chromosomes of crested newts Triturus cristatus (Laurenti). Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1960, 243, 135–219. [Google Scholar] [CrossRef]
- Sims, S.H.; Macgregor, H.C.; Pellatt, P.S.; Horner, H.A. Chromosome 1 in crested and marbled newts (Triturus). Chromosoma 1984, 89, 169–185. [Google Scholar] [CrossRef]
- Yonenaga, Y. Karyotypes and Chromosome Polymorphism in Brazilian Rodents. Caryologia 1975, 28, 269–286. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, A.; Iwasa, M.A. Karyological characterization of the Japanese water shrew, Chimarrogale platycephala (Soricidae, Soricomorpha). Caryologia 2013, 66, 84–89. [Google Scholar] [CrossRef]
- Targino Valente, G.; de Andrade Vitorino, C.; Cabral-de-Mello, D.C.; Oliveira, C.; Lima Souza, I.; Martins, C.; Venere, P.C. Comparative cytogenetics of ten species of cichlid fishes (Teleostei, Cichlidae) from the Araguaia River system, Brazil, by conventional cytogenetic methods. Comp. Cytogenet. 2012, 6, 163–181. [Google Scholar] [CrossRef]
- Ruiz-Ruano, F.J.; López-León, M.D.; Cabrero, J.; Camacho, J.P.M. High-throughput analysis of the satellitome illuminates satellite DNA evolution. Sci. Rep. 2016, 6, 28333. [Google Scholar] [CrossRef] [Green Version]
- Louzada, S.; Lopes, M.; Ferreira, D.; Adega, F.; Escudeiro, A.; Gama-Carvalho, M.; Chaves, R. Decoding the Role of Satellite DNA in Genome Architecture and Plasticity—An Evolutionary and Clinical Affair. Genes 2020, 11, 72. [Google Scholar] [CrossRef] [Green Version]
- del Bosque, M.E.Q.; López-Flores, I.; Suárez-Santiago, V.N.; Garrido-Ramos, M.A. Satellite-DNA diversification and the evolution of major lineages in Cardueae (Carduoideae Asteraceae). J. Plant Res. 2014, 127, 575–583. [Google Scholar] [CrossRef]
- Bolsheva, N.L.; Melnikova, N.V.; Kirov, I.V.; Dmitriev, A.A.; Krasnov, G.S.; Amosova, А.V.; Samatadze, T.E.; Yurkevich, O.Y.; Zoshchuk, S.A.; Kudryavtseva, A.V.; et al. Characterization of repeated DNA sequences in genomes of blue-flowered flax. BMC Evol. Biol. 2019, 19, 49. [Google Scholar] [CrossRef] [Green Version]
- Fry, K.; Salser, W. Nucleotide sequences of HS-α satellite DNA from kangaroo rat Dipodomys ordii and characterization of similar sequences in other rodents. Cell 1977, 12, 1069–1084. [Google Scholar] [CrossRef]
- Ragghianti, M.; Guerrini, F.; Bucci, S.; Mancino, G.; Hotz, H.; Uzzell, T.; Guex, G.-D. Molecular characterization of a centromeric satellite DNA in the hemiclonal hybrid frog Rana esculenta and its parental species. Chromosom. Res. 1995, 3, 497–506. [Google Scholar] [CrossRef]
- Kuroda, M.; Fujimoto, T.; Murakami, M.; Yamaha, E.; Arai, K. Aberrant Meiotic Configurations Cause Sterility in Clone-Origin Triploid and Inter-Group Hybrid Males of the Dojo Loach, Misgurnus anguillicaudatus. Cytogenet. Genome Res. 2019, 158, 46–54. [Google Scholar] [CrossRef]
- Franck, J.P.C.; Kornfield, I.; Wright, J.M. The Utility of SATA Satellite DNA Sequences for Inferring Phylogenetic Relationships among the Three Major Genera of Tilapiine Cichlid Fishes. Mol. Phylogenet. Evol. 1994, 3, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Varley, J.M.; Macgregor, H.C.; Erba, H.P. Satellite DNA is transcribed on lampbrush chromosomes. Nature 1980, 283, 686–688. [Google Scholar] [CrossRef] [PubMed]
- Krasikova, A.V.; Gaginskaia, E.R. Organization of centromere regions of chromosomes in the lampbrush phase. Tsitologiia 2010, 52, 515–533. [Google Scholar] [PubMed]
- Eymery, A.; Callanan, M.; Vourc’h, C. The secret message of heterochromatin: New insights into the mechanisms and function of centromeric and pericentric repeat sequence transcription. Int. J. Dev. Biol. 2009, 53, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Gaginskaya, E.; Kulikova, T.; Krasikova, A. Avian Lampbrush Chromosomes: A Powerful Tool for Exploration of Genome Expression. Cytogenet. Genome Res. 2009, 124, 251–267. [Google Scholar] [CrossRef] [PubMed]
- Torres, G.A.; Gong, Z.; Iovene, M.; Hirsch, C.D.; Buell, C.R.; Bryan, G.J.; Novák, P.; Macas, J.; Jiang, J. Organization and Evolution of Subtelomeric Satellite Repeats in the Potato Genome. G3 2011, 1, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Ramos, M.A. Satellite DNA in Plants: More than Just Rubbish. Cytogenet. Genome Res. 2015, 146, 153–170. [Google Scholar] [CrossRef]
- Vicari, M.R.; Nogaroto, V.; Noleto, R.B.; Cestari, M.M.; Cioffi, M.B.; Almeida, M.C.; Moreira-Filho, O.; Bertollo, L.A.C.; Artoni, R.F. Satellite DNA and chromosomes in Neotropical fishes: Methods, applications and perspectives. J. Fish Biol. 2010, 76, 1094–1116. [Google Scholar] [CrossRef]
- Araújo, N.P.; de Lima, L.G.; Dias, G.B.; Kuhn, G.C.S.; de Melo, A.L.; Yonenaga-Yassuda, Y.; Stanyon, R.; Svartman, M. Identification and characterization of a subtelomeric satellite DNA in Callitrichini monkeys. DNA Res. 2017, 24, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Utsunomia, R.; Ruiz-Ruano, F.J.; Silva, D.M.Z.A.; Serrano, É.A.; Rosa, I.F.; Scudeler, P.E.S.; Hashimoto, D.T.; Oliveira, C.; Camacho, J.P.M.; Foresti, F. A Glimpse into the Satellite DNA Library in Characidae Fish (Teleostei, Characiformes). Front. Genet. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Louis, E.; Vershinin, A. Telomeres: A diversity of solutions to the problem of chromosome ends. Chromosom. Res. 2005, 13, 425–429. [Google Scholar] [CrossRef]
- Boroń, A.; Ozouf-Costaz, C.; Coutanceau, J.-P.; Woroniecka, K. Gene mapping of 28S and 5S rDNA sites in the spined loach Cobitis taenia (Pisces, Cobitidae) from a diploid population and a diploid–tetraploid population. Genetica 2006, 128, 71. [Google Scholar] [CrossRef] [PubMed]
- Culling, M.A.; Janko, K.; Boron, A.; Vasil’ev, V.P.; Côté, I.M.; Hewitt, G.M. European colonization by the spined loach (Cobitis taenia) from Ponto-Caspian refugia based on mitochondrial DNA variation. Mol. Ecol. 2006, 15, 173–190. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Satellite | Forward Primer 5′-3′ | Reverse Primer 5′-3′ |
|---|---|---|
| SatCE01 | TTTGGGGCAGTCTTGTTGGT | CGTGTGCCCATAGCTCTTCA |
| SatCE02 | ACAGTGTGGTTGGCAGTTGA | GCTGGGAACTAGATGCTTGGT |
| SatCE03 | TCACCCCTGTCCTGTACCAA | ACCTGATCCGGCACAGAAAG |
| SatCE04 | GCTCAGAGCAGCGTTTTACA | ACATCTGCATGTTGCTGTGAAC |
| SatCE05 | AAAGGACCTGT ACGTTGGGC | ACCTTTGAGCAGGGTCTTCG |
| SatCE06 | CCCTGCTCGTCCTACATGAAC | CGGGTGAAAAAGGCAATGGG |
| SatCE07 | GCCACCTCAGGTCAATCTCC | CGAAAGCAG GGGTTTGCTTC |
| Satellite | Length (bp) | Genome Proportion * | # of Signals in C. elongatoides | # of Signals in C. taenia | # of Signals in C. elongatoides × 2C. taenia |
|---|---|---|---|---|---|
| SatCE01 | 1010 | 0.07% | 2 SM | 2 SM | 3 SM |
| SatCE02 | 910 | 0.11% | 2 SM | 4 SM | 5 SM |
| SatCE03 | 1400 | 0.079% | 6 M | 2 A | 5 (3 M + 2 A) |
| SatCE04 | 128 | 0.6% | 40–42 | 46 | ~67 |
| SatCE05 | 628 | 0.049% | 2 SM | 2 SM | 3 SM |
| SatCE06 | 1820 | 0.077% | 4 (2 M + 2 SM) | 2 SM | 4 (1 M + 3 SM) |
| SatCE07 | 607 | 0.017% | 8–10 | 8–10 | 12–15 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marta, A.; Dedukh, D.; Bartoš, O.; Majtánová, Z.; Janko, K. Cytogenetic Characterization of Seven Novel satDNA Markers in Two Species of Spined Loaches (Cobitis) and Their Clonal Hybrids. Genes 2020, 11, 617. https://doi.org/10.3390/genes11060617
Marta A, Dedukh D, Bartoš O, Majtánová Z, Janko K. Cytogenetic Characterization of Seven Novel satDNA Markers in Two Species of Spined Loaches (Cobitis) and Their Clonal Hybrids. Genes. 2020; 11(6):617. https://doi.org/10.3390/genes11060617
Chicago/Turabian StyleMarta, Anatolie, Dmitry Dedukh, Oldřich Bartoš, Zuzana Majtánová, and Karel Janko. 2020. "Cytogenetic Characterization of Seven Novel satDNA Markers in Two Species of Spined Loaches (Cobitis) and Their Clonal Hybrids" Genes 11, no. 6: 617. https://doi.org/10.3390/genes11060617
APA StyleMarta, A., Dedukh, D., Bartoš, O., Majtánová, Z., & Janko, K. (2020). Cytogenetic Characterization of Seven Novel satDNA Markers in Two Species of Spined Loaches (Cobitis) and Their Clonal Hybrids. Genes, 11(6), 617. https://doi.org/10.3390/genes11060617