Novel Insights into the Protective Properties of ACTH(4-7)PGP (Semax) Peptide at the Transcriptome Level Following Cerebral Ischaemia–Reperfusion in Rats

 ,
,  , and
, and 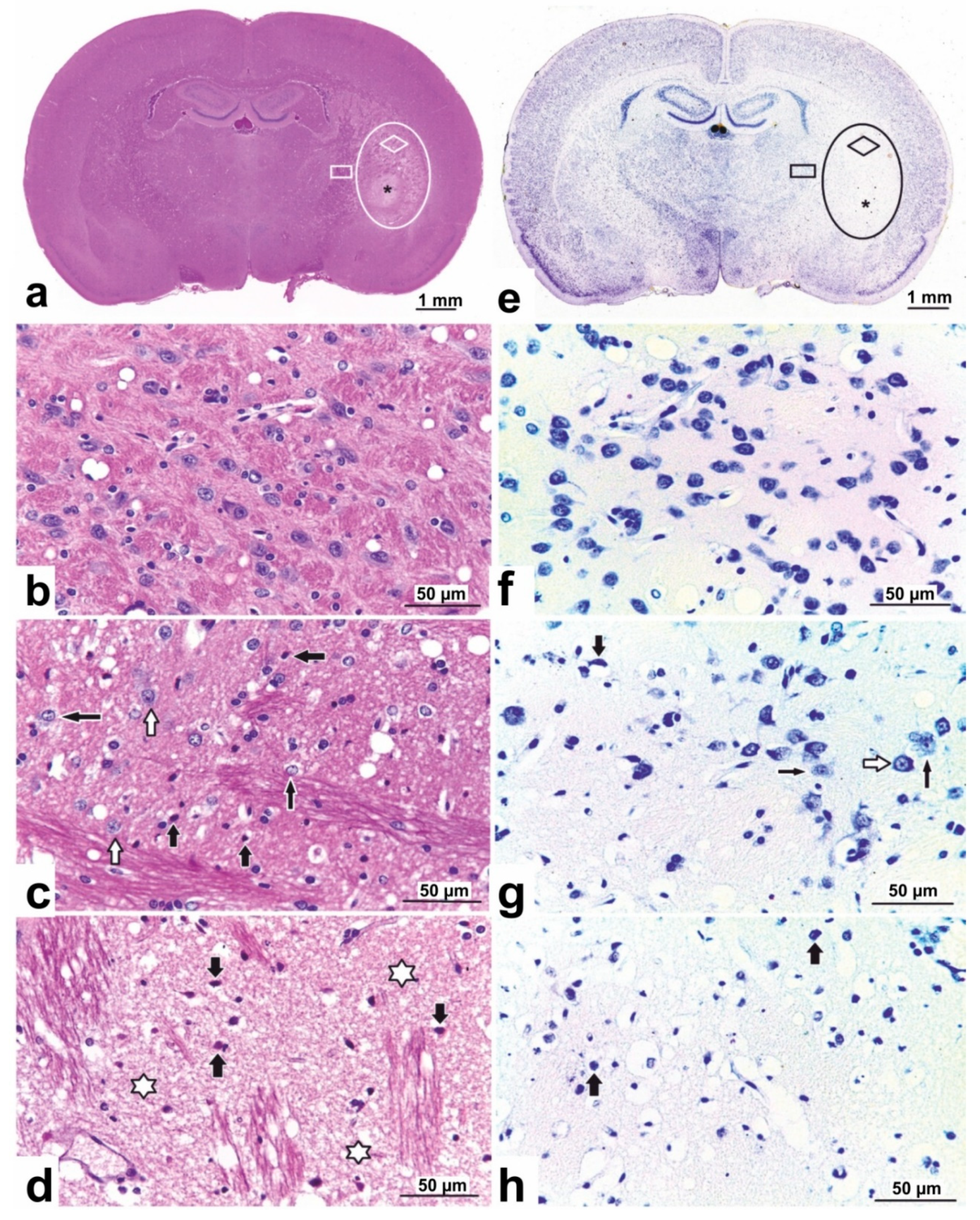{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Transient Cerebral Ischaemia Rat Model
2.2.1. Operation
2.2.2. Semax Administration
2.3. Magnetic Resonance Imaging
2.4. Histological Examination of Rat Brains
2.5. RNA Isolation
2.6. RNA-Seq
2.7. RNA-Seq Data Analysis
2.8. cDNA Synthesis
2.9. Real-Time Reverse Transcription Polymerase Chain Reaction (RT–PCR)
2.10. Data Analysis of Real-Time RT–PCR and Statistics
2.11. Functional Analysis
2.12. Availability of Data and Material
2.13. Ethics Approval and Consent to Participate
3. Results
3.1. Characterization of tMCAO Model Conditions Using MRI
3.2. Histopathological Characterization of Rat Brain
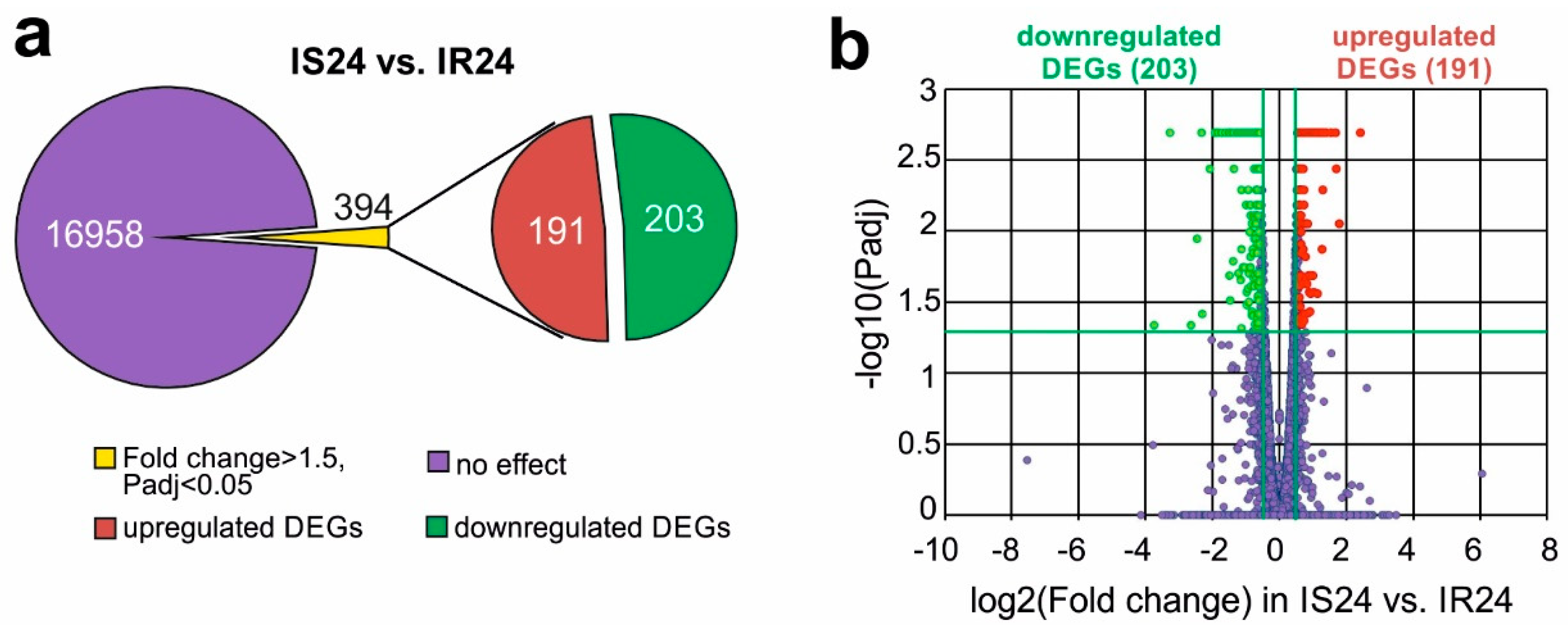
3.3. RNA-Seq Analysis of the Effect of Semax on the Transcriptome after tMCAO
3.4. Functional Annotation of Semax-Induced DEGs Identified in the Rat Brain Subcortex at 24 h after tMCAO
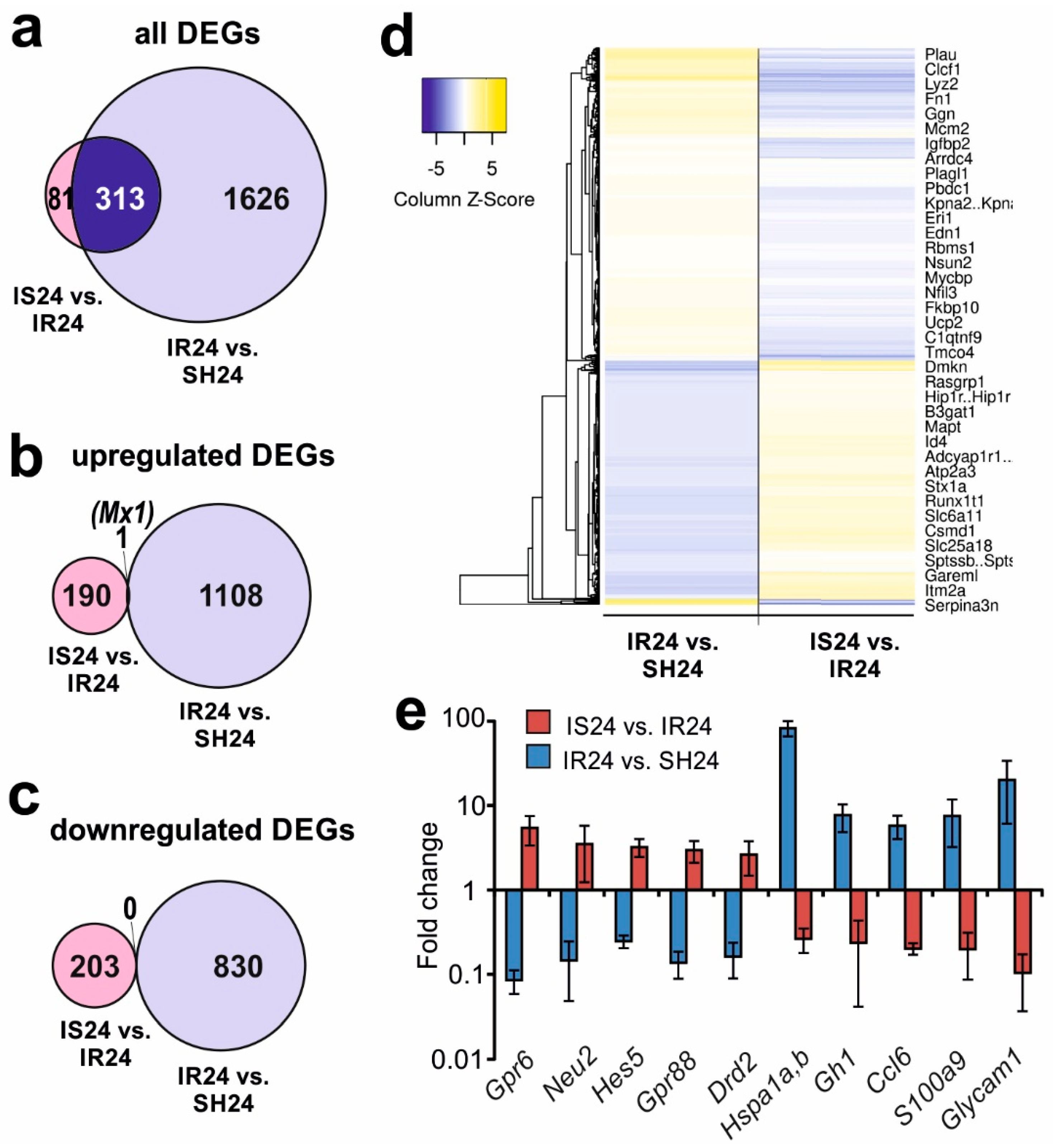
3.5. Differences in Rat Brain Transcriptomes Following Ischaemia and after Semax Administration
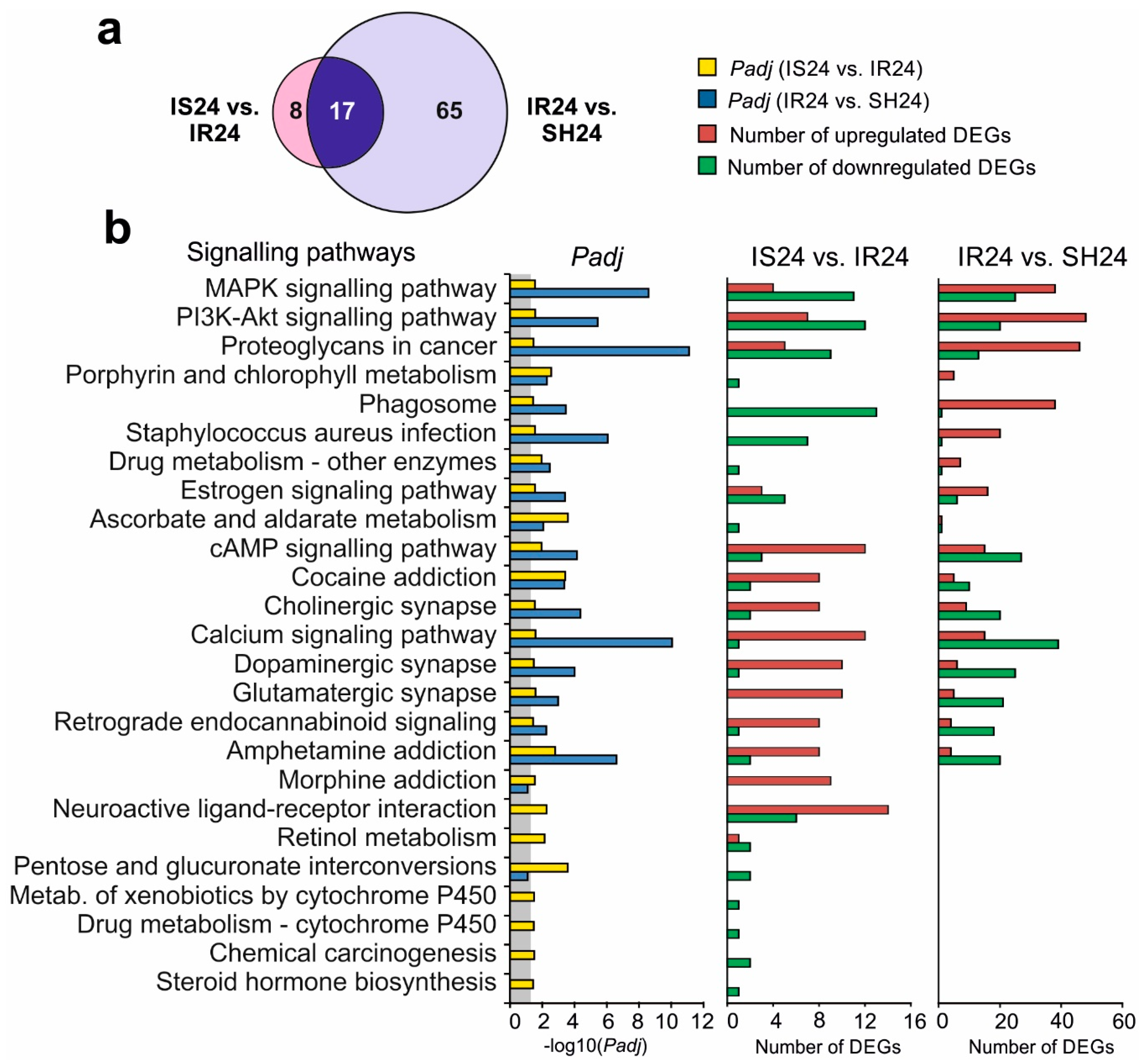
3.6. Signalling Pathways Associated with Semax-Induced DEGs under tMCAO Model Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Asmarin, I.P.; Nezavibat’ko, V.N.; Miasoedov, N.F.; Kamenskiĭ, A.A.; Grivennikov, I.A.; Ponomareva-Stepnaia, M.A.; Andreeva, L.A.; Kaplan, A.I.; Koshelev, V.B.; Riasina, T.V. A nootropic adrenocorticotropin analog 4-10-semax (l5 years experience in its design and study). Zh. Vyssh. Nerv. Deiat. Im. I P Pavlova 1997, 47, 420–430. [Google Scholar] [PubMed]
- de Wied, D. Neuropeptides in learning and memory processes. Behav. Brain Res. 1997, 83, 83–90. [Google Scholar] [CrossRef]
- Kaplan, A.I.; Koshelev, V.B.; Nezavibat’ko, V.N.; Ashmarin, I.P. [Increased resistance to hypoxia effected by the neuropeptide preparation SEMAX]. Fiziol. Cheloveka 1992, 18, 104–107. [Google Scholar] [PubMed]
- Storozhevykh, T.P.; Tukhbatova, G.R.; Senilova, Y.E.; Pinelis, V.G.; Andreeva, L.A.; Myasoyedov, N.F. Effects of semax and its Pro-Gly-Pro fragment on calcium homeostasis of neurons and their survival under conditions of glutamate toxicity. Bull. Exp. Biol. Med. 2007, 143, 601–604. [Google Scholar] [CrossRef]
- Inozemtsev, A.N.; Bokieva, S.B.; Karpukhina, O.V.; Gumargalieva, K.Z.; Kamensky, A.A.; Myasoedov, N.F. Semax prevents learning and memory inhibition by heavy metals. Dokl. Biol. Sci. Proc. Acad. Sci. USSR Biol. Sci. Sect. 2016, 468, 112–114. [Google Scholar] [CrossRef]
- Bashkatova, V.G.; Koshelev, V.B.; Fadyukova, O.E.; Alexeev, A.A.; Vanin, A.F.; Rayevsky, K.S.; Ashmarin, I.P.; Armstrong, D.M. Novel synthetic analogue of ACTH 4-10 (Semax) but not glycine prevents the enhanced nitric oxide generation in cerebral cortex of rats with incomplete global ischemia. Brain Res. 2001, 894, 145–149. [Google Scholar] [CrossRef]
- Gusev, E.; Skvortsova, V. Brain Ischaemia, 1st ed.; Meditsina Publishers: Moscow, Russia, 2001; p. 328. [Google Scholar]
- Tabbì, G.; Magrì, A.; Giuffrida, A.; Lanza, V.; Pappalardo, G.; Naletova, I.; Nicoletti, V.G.; Attanasio, F.; Rizzarelli, E. Semax, an ACTH4-10 peptide analog with high affinity for copper(II) ion and protective ability against metal induced cell toxicity. J. Inorg. Biochem. 2015, 142, 39–46. [Google Scholar] [CrossRef]
- Glazova, N.Y.; Sebentsova, E.A.; Manchenko, D.M.; Andreeva, L.A.; Dergunova, L.V.; Levitskaya, N.G.; Limborska, S.A.; Myasoedov, N.F. The Protective Effect of Semax in a Model of Stress-Induced Impairment of Memory and Behavior in White Rats. Biol. Bull. 2018, 45, 394–399. [Google Scholar] [CrossRef]
- Levitskaya, N.G.; Glazova, N.Y.; Sebentsova, E.A.; Manchenko, D.M.; Vilensky, D.A.; Andreeva, L.A.; Kamensky, A.A.; Myasoedov, N.F. Investigation of the Spectrum of Physiological Activities of the Heptapeptide Semax, an ACTH 4–10 Analogue. Neurochem. J. 2008, 2, 95–101. [Google Scholar]
- Ashmarin, I.; Nezavibatko, V.; Levitskaya, N.; Koshelev, V.; Kamensky, A. Design and Investigation of an ACTH(4-10) Analog Lacking D-Amino Acids and Hydrophobic Radicals. Neurosci. Res. Commun. 1995, 16, 105–112. [Google Scholar]
- Silachev, D.N.; Shram, S.I.; Shakova, F.M.; Romanova, G.A.; Myasoedov, N.F. Formation of spatial memory in rats with ischemic lesions to the prefrontal cortex; effects of a synthetic analog of ACTH(4-7). Neurosci. Behav. Physiol. 2009, 39, 749–756. [Google Scholar] [CrossRef]
- Romanova, G.A.; Silachev, D.N.; Shakova, F.M.; Kvashennikova, Y.N.; Viktorov, I.V.; Shram, S.I.; Myasoedov, N.F. Neuroprotective and antiamnesic effects of Semax during experimental ischemic infarction of the cerebral cortex. Bull. Exp. Biol. Med. 2006, 142, 663–666. [Google Scholar] [CrossRef]
- Gusev, E.I.; Martynov, M.Y.; Kostenko, E.V.; Petrova, L.V.; Bobyreva, S.N. [The efficacy of semax in the tretament of patients at different stages of ischemic stroke]. Zhurnal Nevrol. i psikhiatrii Im. S.S. Korsakova 2018, 118, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Gusev, E.I.; Skvortsova, V.I.; Miasoedov, N.F.; Nezavibat’ko, V.N.; Zhuravleva, E.I.; Vanichkin, A.V. [Effectiveness of semax in acute period of hemispheric ischemic stroke (a clinical and electrophysiological study)]. Zhurnal Nevrol. i psikhiatrii Im. S.S. Korsakova 1997, 97, 26–34. [Google Scholar]
- Hernandes, M.S.; Lassègue, B.; Hilenski, L.L.; Adams, J.; Gao, N.; Kuan, C.-Y.; Sun, Y.-Y.; Cheng, L.; Kikuchi, D.S.; Yepes, M.; et al. Polymerase delta-interacting protein 2 deficiency protects against blood-brain barrier permeability in the ischemic brain. J. Neuroinflamm. 2018, 15, 45. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Liu, H.; Zhang, L.; Wang, G.; Zhang, M.; Yu, Y. Neuroprotection of Dexmedetomidine against Cerebral Ischemia-Reperfusion Injury in Rats: Involved in Inhibition of NF-κB and Inflammation Response. Biomol. Ther. (Seoul) 2017, 25, 383–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghazavi, H.; Hoseini, S.J.; Ebrahimzadeh-Bideskan, A.; Mashkani, B.; Mehri, S.; Ghorbani, A.; Sadri, K.; Mahdipour, E.; Ghasemi, F.; Forouzanfar, F.; et al. Fibroblast Growth Factor Type 1 (FGF1)-Overexpressed Adipose-Derived Mesenchaymal Stem Cells (AD-MSCFGF1) Induce Neuroprotection and Functional Recovery in a Rat Stroke Model. Stem Cell Rev. 2017, 13, 670–685. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, Y.S.; Kim, H.Y.; Noh, M.-Y.; Kim, J.Y.; Lee, Y.-J.; Kim, J.; Park, J.; Kim, S.H. Early Treatment with Poly(ADP-Ribose) Polymerase-1 Inhibitor (JPI-289) Reduces Infarct Volume and Improves Long-Term Behavior in an Animal Model of Ischemic Stroke. Mol. Neurobiol. 2018, 55, 7153–7163. [Google Scholar] [CrossRef]
- Ismael, S.; Zhao, L.; Nasoohi, S.; Ishrat, T. Inhibition of the NLRP3-inflammasome as a potential approach for neuroprotection after stroke. Sci. Rep. 2018, 8, 5971. [Google Scholar] [CrossRef] [Green Version]
- Berger, C.; Stauder, A.; Xia, F.; Sommer, C.; Schwab, S. Neuroprotection and glutamate attenuation by acetylsalicylic acid in temporary but not in permanent cerebral ischemia. Exp. Neurol. 2008, 210, 543–548. [Google Scholar] [CrossRef]
- White, B.C.; Sullivan, J.M.; DeGracia, D.J.; O’Neil, B.J.; Neumar, R.W.; Grossman, L.I.; Rafols, J.A.; Krause, G.S. Brain ischemia and reperfusion: Molecular mechanisms of neuronal injury. J. Neurol. Sci. 2000, 179, 1–33. [Google Scholar] [CrossRef]
- Medvedeva, E.V.; Dmitrieva, V.G.; Povarova, O.V.; Limborska, S.A.; Skvortsova, V.I.; Myasoedov, N.F.; Dergunova, L.V. Effect of semax and its C-terminal fragment Pro-Gly-Pro on the expression of VEGF family genes and their receptors in experimental focal ischemia of the rat brain. J. Mol. Neurosci. 2013, 49, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Dmitrieva, V.G.; Povarova, O.V.; Skvortsova, V.I.; Limborska, S.A.; Myasoedov, N.F.; Dergunova, L.V. Semax and Pro-Gly-Pro activate the transcription of neurotrophins and their receptor genes after cerebral ischemia. Cell. Mol. Neurobiol. 2010, 30, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Stavchansky, V.V.; Yuzhakov, V.V.; Botsina, A.Y.; Skvortsova, V.I.; Bondurko, L.N.; Tsyganova, M.G.; Limborska, S.A.; Myasoedov, N.F.; Dergunova, L.V. The effect of Semax and its C-end peptide PGP on the morphology and proliferative activity of rat brain cells during experimental ischemia: A pilot study. J. Mol. Neurosci. 2011, 45, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Medvedeva, E.V.; Dmitrieva, V.G.; Povarova, O.V.; Limborska, S.A.; Skvortsova, V.I.; Myasoedov, N.F.; Dergunova, L.V. The peptide semax affects the expression of genes related to the immune and vascular systems in rat brain focal ischemia: Genome-wide transcriptional analysis. BMC Genomics 2014, 15, 228. [Google Scholar] [CrossRef] [Green Version]
- Medvedeva, E.V.; Dmitrieva, V.G.; Limborska, S.A.; Myasoedov, N.F.; Dergunova, L.V. Semax, an analog of ACTH(4-7), regulates expression of immune response genes during ischemic brain injury in rats. Mol. Genet. Genomics 2017, 292, 635–653. [Google Scholar] [CrossRef]
- Nieswandt, B.; Kleinschnitz, C.; Stoll, G. Ischaemic stroke: A thrombo-inflammatory disease? J. Physiol. 2011, 589, 4115–4123. [Google Scholar] [CrossRef]
- Ryang, Y.-M.; Dang, J.; Kipp, M.; Petersen, K.-U.; Fahlenkamp, A.V.; Gempt, J.; Wesp, D.; Rossaint, R.; Beyer, C.; Coburn, M. Solulin reduces infarct volume and regulates gene-expression in transient middle cerebral artery occlusion in rats. BMC Neurosci. 2011, 12, 113. [Google Scholar] [CrossRef] [Green Version]
- Lopes, R.D.; Piccini, J.P.; Hylek, E.M.; Granger, C.B.; Alexander, J.H. Antithrombotic therapy in atrial fibrillation: Guidelines translated for the clinician. J. Thromb. Thrombolysis 2008, 26, 167–174. [Google Scholar] [CrossRef]
- Alexandrov, A.V.; Hall, C.E.; Labiche, L.A.; Wojner, A.W.; Grotta, J.C. Ischemic stunning of the brain: Early recanalization without immediate clinical improvement in acute ischemic stroke. Stroke 2004, 35, 449–452. [Google Scholar] [CrossRef] [Green Version]
- Dergunova, L.V.; Filippenkov, I.B.; Stavchansky, V.V.; Denisova, A.E.; Yuzhakov, V.V.; Mozerov, S.A.; Gubsky, L.V.; Limborska, S.A. Genome-wide transcriptome analysis using RNA-Seq reveals a large number of differentially expressed genes in a transient MCAO rat model. BMC Genomics 2018, 19, 655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koizumi, J.; Yoshida, Y.; Nakazawa, T.; Ooneda, G. Experimental studies of ischemic brain edema. Nosotchu 1986, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Miasoedova, N.F.; Skvortsova, V.I.; Nasonov, E.L.; Zhuravleva, E.I.; Grivennikov, I.A.; Arsen’eva, E.L.; Sukhanov, I.I. [Investigation of mechanisms of neuro-protective effect of semax in acute period of ischemic stroke]. Zhurnal Nevrol. i psikhiatrii Im. S.S. Korsakova 1999, 99, 15–19. [Google Scholar]
- Li, Y.; Powers, C.; Jiang, N.; Chopp, M. Intact, injured, necrotic and apoptotic cells after focal cerebral ischemia in the rat. J. Neurol. Sci. 1998, 156, 119–132. [Google Scholar] [CrossRef]
- Lipton, P. Ischemic cell death in brain neurons. Physiol. Rev. 1999, 79, 1431–1568. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.H.; Liu, K.F.; Ho, K.L. Neuronal necrosis after middle cerebral artery occlusion in Wistar rats progresses at different time intervals in the caudoputamen and the cortex. Stroke 1995, 26, 636–642, discussion 643. [Google Scholar] [CrossRef] [PubMed]
- BUTTNER-ENNEVER, J. The Rat Brain in Stereotaxic Coordinates, 3rd edn. By George Paxinos and Charles Watson. (Pp. xxxiii+80; illustrated; f$69.95 paperback; ISBN 0 12 547623; comes with CD-ROM.) San Diego: Academic Press. 1996. J. Anat. 1997, 191, 315–317. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper–Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reimand, J.; Arak, T.; Adler, P.; Kolberg, L.; Reisberg, S.; Peterson, H.; Vilo, J. g:Profiler-a web server for functional interpretation of gene lists (2016 update). Nucleic Acids Res. 2016, 44, 83–89. [Google Scholar] [CrossRef]
- Mi, H.; Huang, X.; Muruganujan, A.; Tang, H.; Mills, C.; Kang, D.; Thomas, P.D. PANTHER version 11: Expanded annotation data from Gene Ontology and Reactome pathways, and data analysis tool enhancements. Nucleic Acids Res. 2017, 45, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/Traces/study/?acc=SRP148632 (accessed on 10 May 2020).
- National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/sra/PRJNA491404 (accessed on 10 May 2020).
- Wang, C.-M.; Pan, Y.-Y.; Liu, M.-H.; Cheng, B.-H.; Bai, B.; Chen, J. RNA-seq expression profiling of rat MCAO model following reperfusion Orexin-A. Oncotarget 2017, 8, 113066–113081. [Google Scholar] [CrossRef] [Green Version]
- Milani, D.; Cross, J.L.; Anderton, R.S.; Blacker, D.J.; Knuckey, N.W.; Meloni, B.P. Neuroprotective efficacy of poly-arginine R18 and NA-1 (TAT-NR2B9c) peptides following transient middle cerebral artery occlusion in the rat. Neurosci. Res. 2017, 114, 9–15. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.-M.; Cao, W.; Yao, K.-W.; Liu, Z.-Q.; Guo, J.-Y. Anti-inflammation and antioxidant effect of Cordymin, a peptide purified from the medicinal mushroom Cordyceps sinensis, in middle cerebral artery occlusion-induced focal cerebral ischemia in rats. Metab. Brain Dis. 2012, 27, 159–165. [Google Scholar] [CrossRef]
- Gusev, E.I.; Skvortsova, V.I.; Chukanova, E.I. [Semax in prevention of disease progress and development of exacerbations in patients with cerebrovascular insufficiency]. Zhurnal Nevrol. i psikhiatrii Im. S.S. Korsakova 2005, 105, 35–40. [Google Scholar]
- Hori, M.; Nakamachi, T.; Shibato, J.; Rakwal, R.; Shioda, S.; Numazawa, S. Unraveling the Specific Ischemic Core and Penumbra Transcriptome in the Permanent Middle Cerebral Artery Occlusion Mouse Model Brain Treated with the Neuropeptide PACAP38. Microarrays 2015, 4, 2–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.H.; Jobichen, C.; Chia, C.S.B.; Chan, T.H.M.; Tang, J.P.; Chung, T.X.Y.; Li, J.; Poulsen, A.; Hung, A.W.; Koh-Stenta, X.; et al. Targeting cancer addiction for SALL4 by shifting its transcriptome with a pharmacologic peptide. Proc. Natl. Acad. Sci. USA 2018, 115, 7119–7128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canazza, A.; Minati, L.; Boffano, C.; Parati, E.; Binks, S. Experimental models of brain ischemia: A review of techniques, magnetic resonance imaging, and investigational cell-based therapies. Front. Neurol. 2014, 5, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, G.; Xu, Z.; Gates, A.; Jiang, J.; Ford, B.D. Expression Analysis Systematic Explorer (EASE) analysis reveals differential gene expression in permanent and transient focal stroke rat models. Brain Res. 2006, 1071, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, G.A.; Estrada, E.Y.; Dencoff, J.E. Matrix metalloproteinases and TIMPs are associated with blood-brain barrier opening after reperfusion in rat brain. Stroke 1998, 29, 2189–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lochhead, J.J.; McCaffrey, G.; Quigley, C.E.; Finch, J.; DeMarco, K.M.; Nametz, N.; Davis, T.P. Oxidative stress increases blood-brain barrier permeability and induces alterations in occludin during hypoxia-reoxygenation. J. Cereb. Blood Flow Metab. 2010, 30, 1625–1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritter, L.S.; Orozco, J.A.; Coull, B.M.; McDonagh, P.F.; Rosenblum, W.I. Leukocyte accumulation and hemodynamic changes in the cerebral microcirculation during early reperfusion after stroke. Stroke 2000, 31, 1153–1161. [Google Scholar] [CrossRef] [Green Version]
- Nour, M.; Scalzo, F.; Liebeskind, D.S. Ischemia-reperfusion injury in stroke. Interv. Neurol. 2013, 1, 185–199. [Google Scholar] [CrossRef]
- Eremin, K.O.; Kudrin, V.S.; Saransaari, P.; Oja, S.S.; Grivennikov, I.A.; Myasoedov, N.F.; Rayevsky, K.S. Semax, an ACTH(4-10) analogue with nootropic properties, activates dopaminergic and serotoninergic brain systems in rodents. Neurochem. Res. 2005, 30, 1493–1500. [Google Scholar] [CrossRef]
- Sharonova, I.N.; Bukanova, Y.V.; Myasoedov, N.F.; Skrebitskii, V.G. Modulation of GABA- and Glycine-Activated Ionic Currents with Semax in Isolated Cerebral Neurons. Bull. Exp. Biol. Med. 2018, 164, 612–616. [Google Scholar] [CrossRef]
- Adan, R.A.H.; Oosterom, J.; Ludvigsdottir, G.; Brakkee, J.H.; Burbach, J.P.H.; Gispen, W.H. Identification of antagonists for melanocortin MC3, MC4 and MC5 receptors. Eur. J. Pharmacol. Mol. Pharmacol. 1994, 269, 331–337. [Google Scholar] [CrossRef] [Green Version]
- Giuliani, D.; Ottani, A.; Neri, L.; Zaffe, D.; Grieco, P.; Jochem, J.; Cavallini, G.M.; Catania, A.; Guarini, S. Multiple beneficial effects of melanocortin MC4 receptor agonists in experimental neurodegenerative disorders: Therapeutic perspectives. Prog. Neurobiol. 2017, 148, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Lisak, R.P.; Benjamins, J.A. Melanocortins, Melanocortin Receptors and Multiple Sclerosis. Brain Sci. 2017, 7, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mykicki, N.; Herrmann, A.M.; Schwab, N.; Deenen, R.; Sparwasser, T.; Limmer, A.; Wachsmuth, L.; Klotz, L.; Köhrer, K.; Faber, C.; et al. Melanocortin-1 receptor activation is neuroprotective in mouse models of neuroinflammatory disease. Sci. Transl. Med. 2016, 8, 362ra146. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filippenkov, I.B.; Stavchansky, V.V.; Denisova, A.E.; Yuzhakov, V.V.; Sevan’kaeva, L.E.; Sudarkina, O.Y.; Dmitrieva, V.G.; Gubsky, L.V.; Myasoedov, N.F.; Limborska, S.A.; et al. Novel Insights into the Protective Properties of ACTH(4-7)PGP (Semax) Peptide at the Transcriptome Level Following Cerebral Ischaemia–Reperfusion in Rats. Genes 2020, 11, 681. https://doi.org/10.3390/genes11060681
Filippenkov IB, Stavchansky VV, Denisova AE, Yuzhakov VV, Sevan’kaeva LE, Sudarkina OY, Dmitrieva VG, Gubsky LV, Myasoedov NF, Limborska SA, et al. Novel Insights into the Protective Properties of ACTH(4-7)PGP (Semax) Peptide at the Transcriptome Level Following Cerebral Ischaemia–Reperfusion in Rats. Genes. 2020; 11(6):681. https://doi.org/10.3390/genes11060681
Chicago/Turabian StyleFilippenkov, Ivan B., Vasily V. Stavchansky, Alina E. Denisova, Vadim V. Yuzhakov, Larisa E. Sevan’kaeva, Olga Y. Sudarkina, Veronika G. Dmitrieva, Leonid V. Gubsky, Nikolai F. Myasoedov, Svetlana A. Limborska, and et al. 2020. "Novel Insights into the Protective Properties of ACTH(4-7)PGP (Semax) Peptide at the Transcriptome Level Following Cerebral Ischaemia–Reperfusion in Rats" Genes 11, no. 6: 681. https://doi.org/10.3390/genes11060681
APA StyleFilippenkov, I. B., Stavchansky, V. V., Denisova, A. E., Yuzhakov, V. V., Sevan’kaeva, L. E., Sudarkina, O. Y., Dmitrieva, V. G., Gubsky, L. V., Myasoedov, N. F., Limborska, S. A., & Dergunova, L. V. (2020). Novel Insights into the Protective Properties of ACTH(4-7)PGP (Semax) Peptide at the Transcriptome Level Following Cerebral Ischaemia–Reperfusion in Rats. Genes, 11(6), 681. https://doi.org/10.3390/genes11060681