Identification of High Molecular Variation Loci in Complete Chloroplast Genomes of Mammillaria (Cactaceae, Caryophyllales)

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Source
2.2. Repeats Characterization and Short Sequence Repeats Identification
2.3. Sequence Divergence in Homologous Loci Among Species
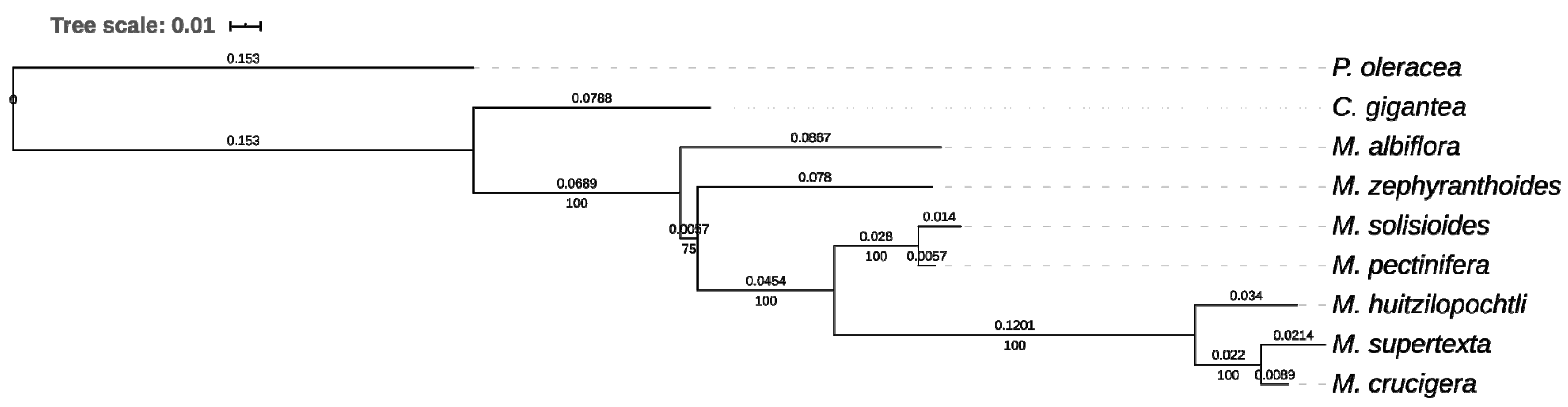
2.4. Phylogenetic Analysis
3. Results
3.1. Repeated Sequences
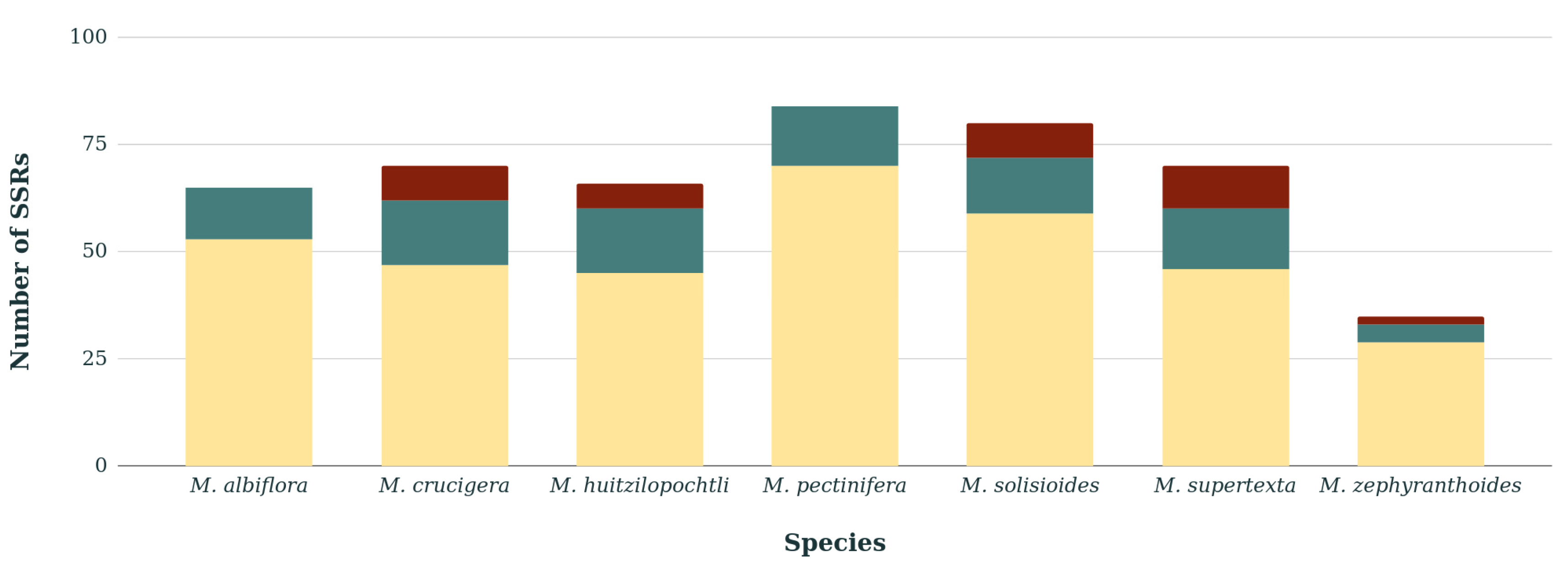
3.2. Variation and Localization of the Short Sequence Repeats
3.3. Divergent DNA Sequences in Homologous Loci
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lemieux, C.; Otis, C.; Turmel, M. Ancestral chloroplast genome in Mesostigma viride reveals an early branch of green plant evolution. Nature 2000, 403, 649–652. [Google Scholar] [CrossRef] [Green Version]
- Wicke, S.; Schneeweiss, G.M.; dePamphilis, C.W.; Müller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef] [Green Version]
- Lavin, M.; Doyle, J.J.; Palmer, J.D. Evolutionary significance of the loss of the chloroplast-DNA inverted repeat in the Leguminosae subfamily Papilionoideae. Evolution 1990, 44, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Downie, S.R.; Palmer, J.D. A chloroplast DNA phylogeny of the Caryophyllales based on structural and inverted repeat restriction site variation. Syst. Bot. 1994, 19, 236–252. [Google Scholar] [CrossRef]
- Cosner, M.E.; Raubeson, L.A.; Jansen, R.K. Chloroplast DNA rearrangements in Campanulaceae: Phylogenetic utility of highly rearranged genomes. BMC Evol. Biol. 2004, 4, 27. [Google Scholar] [CrossRef] [Green Version]
- Graham, S.W.; Reeves, P.A.; Burns, A.C.E.; Olmstead, R.G. Microstructural Changes in Noncoding Chloroplast DNA: Interpretation, Evolution, and Utility of Indels and Inversions in Basal Angiosperm Phylogenetic Inference. Int. J. Plant Sci. 2000, 161, S83–S96. [Google Scholar] [CrossRef] [Green Version]
- Kelchner, S.A. The Evolution of Non-Coding Chloroplast DNA and Its Application in Plant Systematics. Ann. Mo. Bot. Gard. 2000, 87, 482. [Google Scholar] [CrossRef]
- Borsch, T.; Quandt, D.; Koch, M. Molecular evolution and phylogenetic utility of non-coding DNA: Applications from species to deep level questions. Plant Syst. Evol. 2009, 282, 107–108. [Google Scholar] [CrossRef]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M.B. Four primer pairs for the amplification of chloroplast intergenic regions with intraspecific variation. Molec. Ecol. 1999, 8, 521–523. [Google Scholar]
- Steele, K.P.; Wojciechowski, M.F. Phylogenetic analyses of tribes Trifolieae and Vicieae, based on sequences of the plastid gene matK (Papilionoideae: Leguminosae). In Advances in Legume Systematics, Part 10, Higher Level Systematics; Klitgaard, B.B., Bruneau, A., Eds.; Royal Botanic Garden: Kew, UK, 2003; pp. 355–370. [Google Scholar]
- Kelchner, S.A.; Clark, L.G. Molecular evolution and phylogenetic utility of the chloroplast rpl16 intron in Chusquea and the Bambusoideae (Poaceae). Mol. Phylogenet. Evol. 1997, 8, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Prince, L.M.; Parks, C.R. Phylogenetic relationships of Theaceae inferred from chloroplast DNA sequence data. Am. J. Bot. 2001, 88, 2309–2320. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.T.; Bayer, R.J. Molecular phylogenetics of Acacia subgenera Acacia and Aculeiferum (Fabaceae: Mimosoideae), based on the chloroplast matK coding sequence and flanking trnK intron spacer regions. Aust. Syst. Bot. 2003, 16, 27–33. [Google Scholar] [CrossRef]
- Harpke, D.; Peterson, A.; Hoffmann, M.H.; Röser, M. Phylogenetic evaluation of chloroplast trnL–trnF DNA sequence variation in the genus Mammillaria (Cactaceae). Schlechtendalia 2006, 14, 7–16. [Google Scholar]
- Nyffeler, R. Phylogenetic relationships in the cactus family (Cactaceae) based on evidence from trnK/matK and trnL-trnF sequences. Am. J. Bot. 2002, 89, 312–326. [Google Scholar] [CrossRef] [Green Version]
- Mast, A.R.; Kelso, S.; Richards, A.J.; Lang, D.J.; Feller, D.M.S.; Conti, E. Phylogenetic Relationships in Primula L. and Related Genera (Primulaceae) Based on Noncoding Chloroplast DNA. Int. J. Plant Sci. 2001, 162, 1381–1400. [Google Scholar]
- Cuenoud, P.; Savolainen, V.; Chatrou, L.W.; Powell, M.; Grayer, R.J.; Chase, M.W. Molecular phylogenetics of Caryophyllales based on nuclear 18S rDNA and plastid rbcL, atpB, and matK DNA sequences. Am. J. Bot. 2002, 89, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Borsch, T.; Hilu, K.W.; Quandt, D.; Wilde, V.; Neinhuis, C.; Barthlott, W. Noncoding plastid trnT-trnF sequences reveal a well resolved phylogeny of basal angiosperms. J. Evol. Biol. 2003, 16, 558–576. [Google Scholar] [CrossRef] [Green Version]
- Solórzano, S.; Chincoya, D.A.; Sanchez-Flores, A.; Estrada, K.; Díaz-Velásquez, C.E.; González-Rodríguez, A.; Vaca-Paniagua, F.; Dávila, P.; Arias, S. De novo assembly discovered novel structures in genome of plastids and revealed divergent inverted repeats in Mammillaria (Cactaceae, Caryophyllales). Plants 2019, 8, 392. [Google Scholar] [CrossRef] [Green Version]
- Sanderson, M.J.; Copetti, D.; Búrquez, A.; Bustamante, E.; Charboneau, J.L.M.; Eguiarte, L.; Kumar, S.; Lee, H.O.; McMahon, M.; Steele, K.; et al. Exceptional reduction of the plastid genome of saguaro cactus (Carnegiea gigantea). Am. J. Bot. 2015, 102, 1115–1127. [Google Scholar] [CrossRef] [Green Version]
- Bárcenas, R.T.; Yesson, C.; Hawkins, J.A. Molecular systematics of the Cactaceae. Cladistics 2011, 27, 470–489. [Google Scholar] [CrossRef]
- Butterworth, C.A.; Wallace, R.S. Phylogenetic studies of Mammillaria (Cactaceae)—Insights from chloroplast sequence variation and hypothesis testing using the parametric bootstrap. Am. J. Bot. 2004, 91, 1086–1098. [Google Scholar] [CrossRef] [PubMed]
- Hunt, D.; Taylor, N.; Charles, G. The New Cactus Lexicon; DH Books: Milborne Port, UK, 2006. [Google Scholar]
- The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/search/grid?query=mammillaria&searchType=species (accessed on 15 January 2020).
- Arakaki, M.; Christin, P.-A.; Nyffeler, R.; Lendel, A.; Eggli, U.; Ogburn, R.M.; Spriggs, E.; Moore, M.J.; Edwards, E.J. Contemporaneous and recent radiations of the world’s major succulent plant lineages. Proc. Natl. Acad. Sci. USA 2011, 108, 8379–8384. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Hernández, T.; Brown, J.W.; Schlumpberger, B.O.; Eguiarte, L.E.; Magallón, S. Beyond aridification: Multiple explanations for the elevated diversification of cacti in the New World Succulent Biome. New Phytol. 2014, 202, 1382–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtz, S. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [Green Version]
- Thiel, T.; Michalek, W.; Varshney, R.; Graner, A. Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2003, 106, 411–422. [Google Scholar] [CrossRef]
- Katoh, K. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Yang, H.; Zhao, J.; Zhou, B.; Li, T.; Xiang, B. The complete chloroplast genome sequence of the folk medicinal and vegetable plant purslane (Portulaca oleracea L.). J. Hortic. Sci. Biotech. 2018, 93, 356–365. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Weng, M.-L.; Blazier, J.C.; Govindu, M.; Jansen, R.K. Reconstruction of the ancestral plastid genome in Geraniaceae reveals a correlation between genome rearrangements, repeats, and nucleotide substitution rates. Mol. Biol. Evol. 2014, 31, 645–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haberle, R.C.; Fourcade, H.M.; Boore, J.L.; Jansen, R.K. Extensive rearrangements in the chloroplast genome of Trachelium caeruleum are associated with repeats and tRNA genes. J. Mol. Evol. 2008, 66, 350–361. [Google Scholar]
- Blazier, J.C.; Jansen, R.K.; Mower, J.P.; Govindu, M.; Zhang, J.; Weng, M.-L.; Ruhlman, T.A. Variable presence of the inverted repeat and plastome stability in Erodium. Ann. Bot. 2016, 117, 1209–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunnucks, P. Efficient genetic markers for population biology. Trends Ecol. Evol. 2000, 15, 199–203. [Google Scholar] [CrossRef]
- Ellegren, H. Microsatellites: Simple sequences with complex evolution. Nat. Rev. Genet. 2004, 5, 435–445. [Google Scholar]
- Huang, H.; Shi, C.; Liu, Y.; Mao, S.-Y.; Gao, L.-Z. Thirteen Camellia chloroplast genome sequences determined by high-throughput sequencing: Genome structure and phylogenetic relationships. BMC Evol. Biol. 2014, 14, 151. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.-L.; Wang, R.-N.; Zhang, N.-Y.; Fan, W.-B.; Fang, M.-F.; Li, Z.-H. Molecular Evolution of chloroplast genomes of orchid species: Insights into phylogenetic relationship and adaptive evolution. Int. J. Mol. Sci. 2018, 19, 716. [Google Scholar] [CrossRef] [Green Version]
- Parks, M.; Cronn, R.; Liston, A. Increasing phylogenetic resolution at low taxonomic levels using massively parallel sequencing of chloroplast genomes. BMC Biol. 2009, 7, 84. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, J.B.; Lockhart, P.J. Deciphering ancient rapid radiations. Trends Ecol. Evol. 2007, 22, 258–265. [Google Scholar] [CrossRef]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA barcodes to identify flowering plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef] [Green Version]
- Wicke, S.; Quandt, D. Universal primers for the amplification of the plastid trnK/matK region in land plants. Anales Jard. Bot. Madrid 2009, 66, 285–288. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of SSRs | M. albiflora | M. crucigera | M. huitzilopochtli | M. pectinifera | M. solisioides | M. supertexta | M. zephyranthoides |
|---|---|---|---|---|---|---|---|
| 1. Homopolymer | |||||||
| A/T | 56 | 60 | 57 | 69 | 65 | 60 | 31 |
| C/G | 1 | 1 | 1 | 2 | 2 | 1 | 0 |
| 2. Dinucleotide | |||||||
| AT/AT | 5 | 6 | 7 | 9 | 9 | 6 | 3 |
| 3. Trinucleotide | |||||||
| AAG/CTT | 2 | 1 | 0 | 3 | 2 | 1 | 0 |
| AGG/CCT | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| AAT/ATT | 0 | 0 | 0 | 1 | 1 | 0 | 0 |
| 4. Tetranucleotide | |||||||
| AAAT/ATTT | 0 | 2 | 0 | 0 | 0 | 2 | 0 |
| 5. Pentanucleotide | |||||||
| AATAT/ATATT | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| 6. Hexanucleotide | |||||||
| ACGAGG/CCTCGT | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Total number of SSRs | |||||||
| 65 | 70 | 65 | 84 | 80 | 70 | 35 | |
| Species | Number of Loci | ||
|---|---|---|---|
| LSC Region | SSC Region | IRs Region | |
| Arrangement 1 (M. albiflora and M. pectinifera) | 144 | 132 | 142 |
| Arrangement 2 (M. crucigera, M. huitzilopochtli, M. solisioides and M. supertexta) | 34 | 34 | 17 |
| Arrangement 3 (M. zephyranthoides) | 2 | 14 | 21 |
| Locus Name | Locus Type | Location | Variation Percentage (%) | Average Length (bp) |
|---|---|---|---|---|
| 1. accD-trnM * | IGS | LSC | 31.37 | 2176 |
| 2. rrn5-rrn4.5 * | IGS | SSC | 18.56 | 721 |
| 3. infA-rps8 * | IGS | LSC | 11.52 | 103 |
| 4. trnL-trnF * | IGS | LSC | 10.64 | 325 |
| 5. ndhF-trnN | IGS | SSC | 10.63 | 455 |
| 6. trnS-clpP | IGS | LSC | 10.34 | 89 |
| 7. psbM * | Gene | LSC | 10.26 | 102 |
| 8. trnP-psaJ * | IGS | LSC | 9.82 | 399 |
| 9. accDψ | Pseudogene | LSC | 8.99 | 1574 |
| 10. trnN-trnR * | IGS | SSC/IRs | 8.74 | 360 |
| 11. trnF-psbJ | IGS | LSC | 8.68 | 619 |
| 12. clpP * | Gene | LSC | 7.61 | 616 |
| 13. ndhB-ycf2 | IGS | SSC/IRs | 7.34 | 354 |
| 14. trnI-ycf2 * | IGS | SSC/IRs | 7.02 | 109 |
| 15. psaC-ndhD * | IGS | SSC | 6.90 | 146 |
| 16. trnM-atpE * | IGS | LSC | 6.21 | 227 |
| 17. rpl22-rps19 * | IGS | LSC | 5.87 | 67 |
| 18. clpP-psbB * | IGS | LSC | 5.82 | 753 |
| 19. trnL-ycf1 | IGS | SSC | 5.34 | 256 |
| 20. rps4-trnT * | IGS | LSC | 5.14 | 277 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chincoya, D.A.; Sanchez-Flores, A.; Estrada, K.; Díaz-Velásquez, C.E.; González-Rodríguez, A.; Vaca-Paniagua, F.; Dávila, P.; Arias, S.; Solórzano, S. Identification of High Molecular Variation Loci in Complete Chloroplast Genomes of Mammillaria (Cactaceae, Caryophyllales). Genes 2020, 11, 830. https://doi.org/10.3390/genes11070830
Chincoya DA, Sanchez-Flores A, Estrada K, Díaz-Velásquez CE, González-Rodríguez A, Vaca-Paniagua F, Dávila P, Arias S, Solórzano S. Identification of High Molecular Variation Loci in Complete Chloroplast Genomes of Mammillaria (Cactaceae, Caryophyllales). Genes. 2020; 11(7):830. https://doi.org/10.3390/genes11070830
Chicago/Turabian StyleChincoya, Delil A., Alejandro Sanchez-Flores, Karel Estrada, Clara E. Díaz-Velásquez, Antonio González-Rodríguez, Felipe Vaca-Paniagua, Patricia Dávila, Salvador Arias, and Sofía Solórzano. 2020. "Identification of High Molecular Variation Loci in Complete Chloroplast Genomes of Mammillaria (Cactaceae, Caryophyllales)" Genes 11, no. 7: 830. https://doi.org/10.3390/genes11070830
APA StyleChincoya, D. A., Sanchez-Flores, A., Estrada, K., Díaz-Velásquez, C. E., González-Rodríguez, A., Vaca-Paniagua, F., Dávila, P., Arias, S., & Solórzano, S. (2020). Identification of High Molecular Variation Loci in Complete Chloroplast Genomes of Mammillaria (Cactaceae, Caryophyllales). Genes, 11(7), 830. https://doi.org/10.3390/genes11070830