Gene Loss and Evolution of the Plastome

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Retrieval and Annotation
2.2. Multiple Sequence Alignment and Creation of Phylogenetic Trees
2.3. Analysis of the Deletion and Duplication of Chloroplast-Encoded Genes
2.4. Recombination Events and Time Tree Construction of the Chloroplast Genome
2.5. Substitution Rate in Chloroplast Genomes
2.6. Statistical Analysis
3. Results
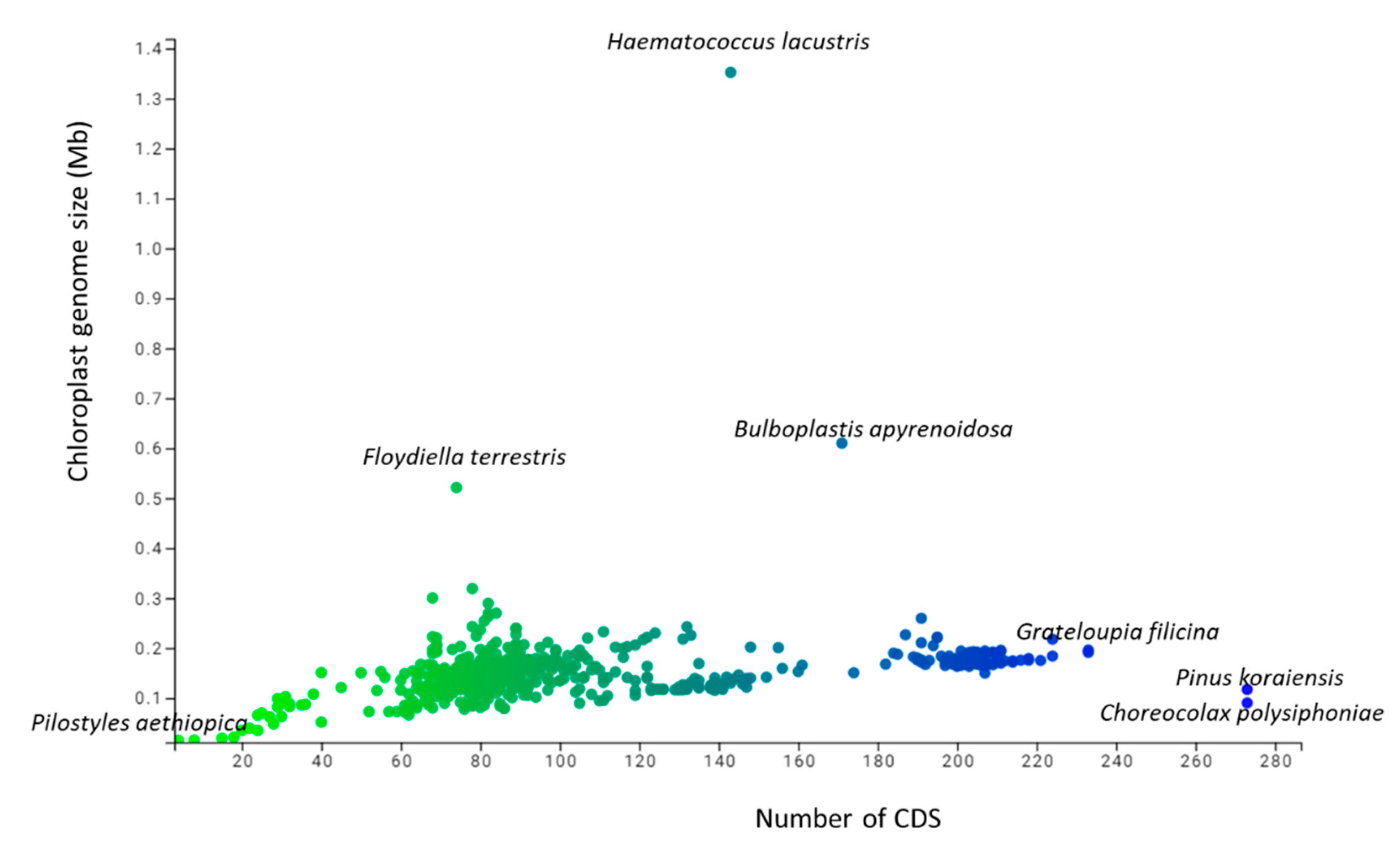
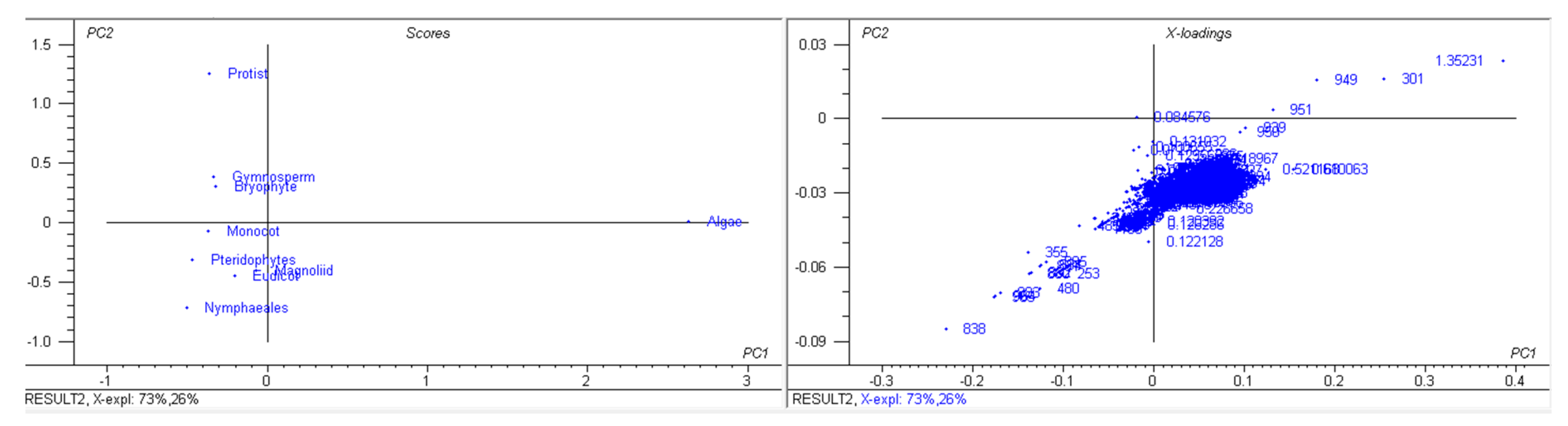
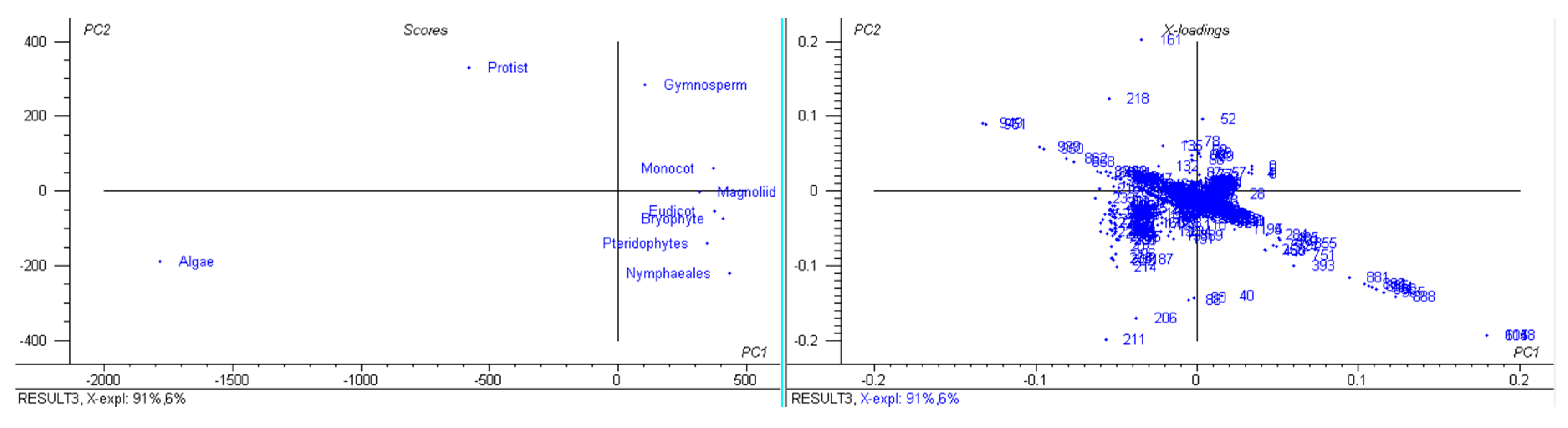
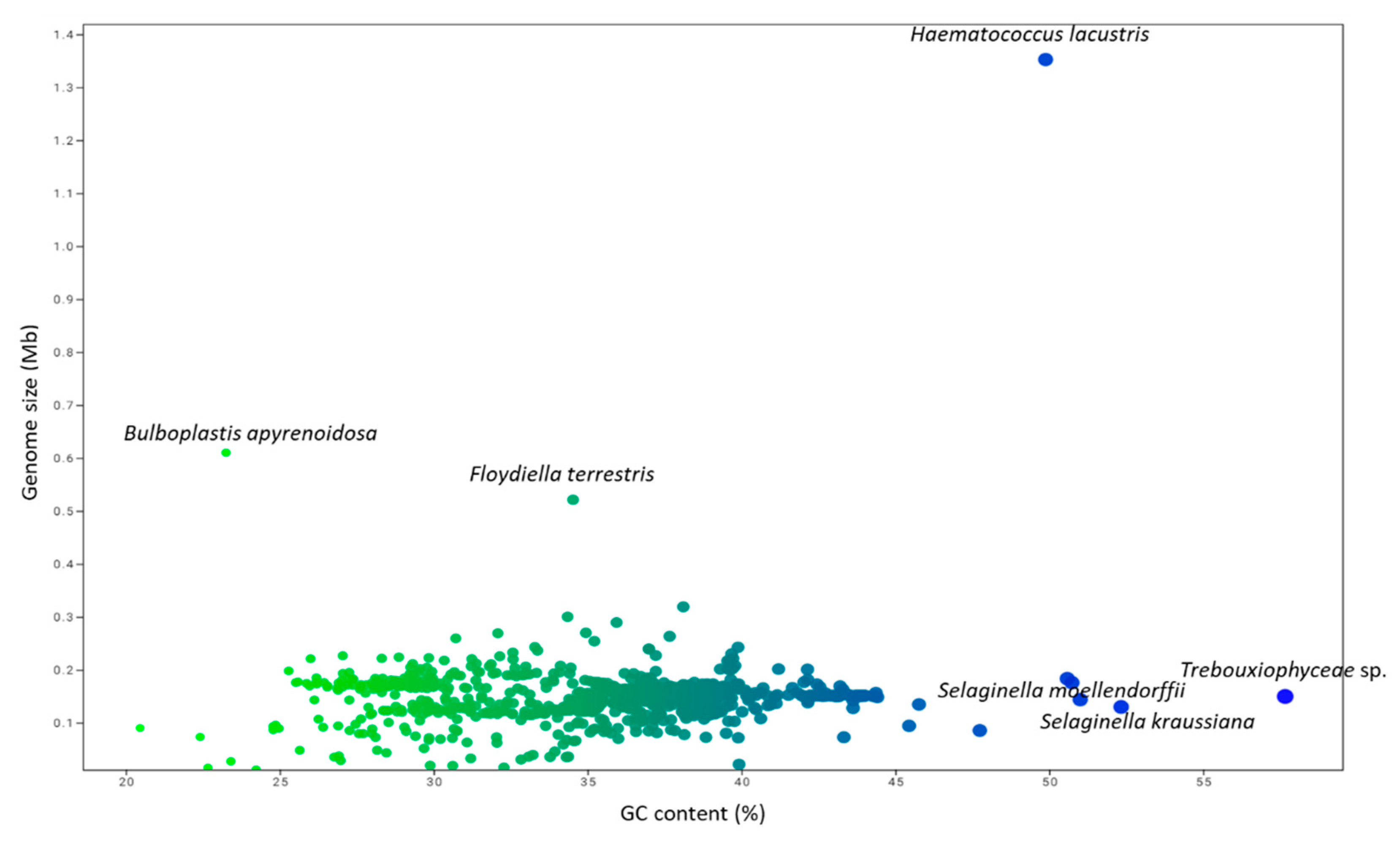
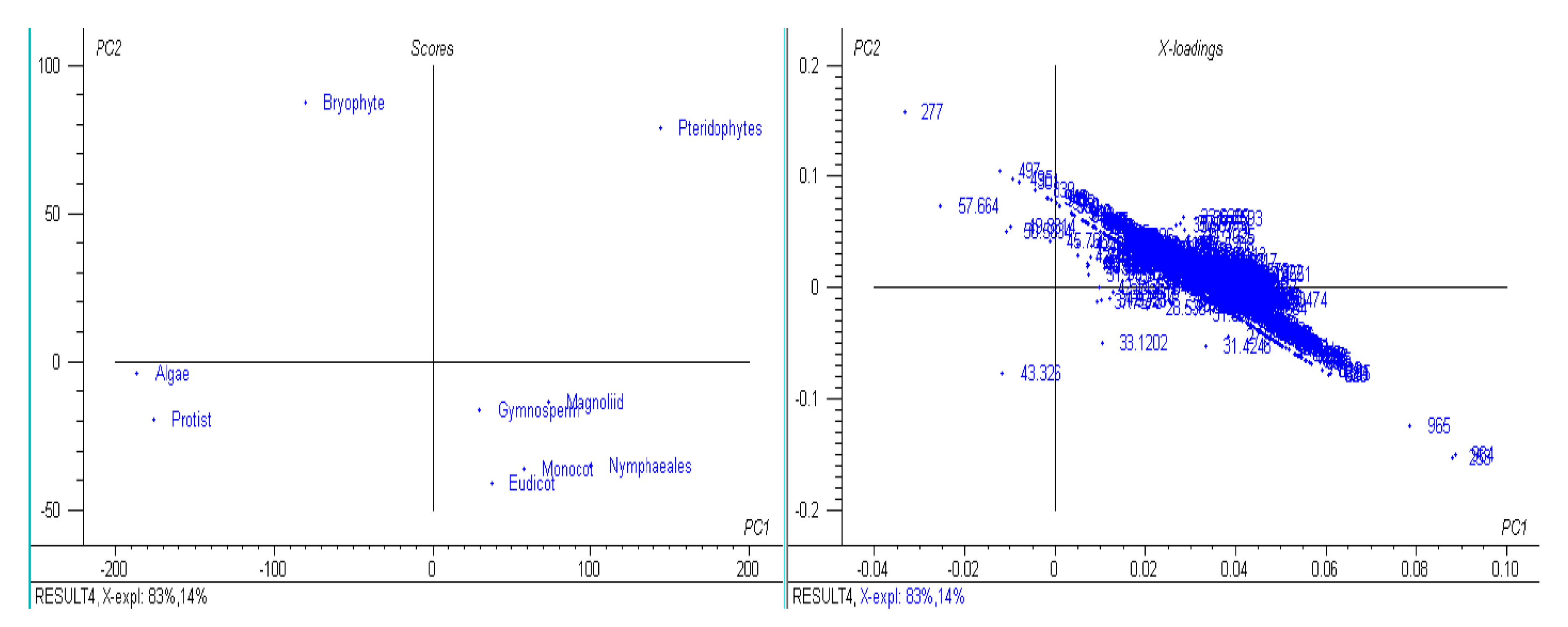
3.1. The Genomic Features of Chloroplast Genomes Are Diverse and Dynamic
3.2. PsaM, Psb30, ChlB, ChlL, ChlN, and RPL21 Are Chloroplast Genes Characteristic of Algae, Bryophytes, Pteridophytes, and Gymnosperms
3.3. The Rbcl Gene Has Been Lost in Parasitic and Heterotrophic Plant Species
3.4. Deletion of Inverted Repeats (IRs) Has Occurred across All Plastid Lineages
3.5. Several Genes in the Chloroplast Genome Have Been Lost
3.6. The Loss of Genes in Chloroplast Genomes is Dynamic
3.7. Chloroplast-Derived Genes Are Present in the Nuclear Genome
3.8. The Ratio of Nucleotide Substitution Is Highest in Pteridophytes and Lowest in Nymphaeales
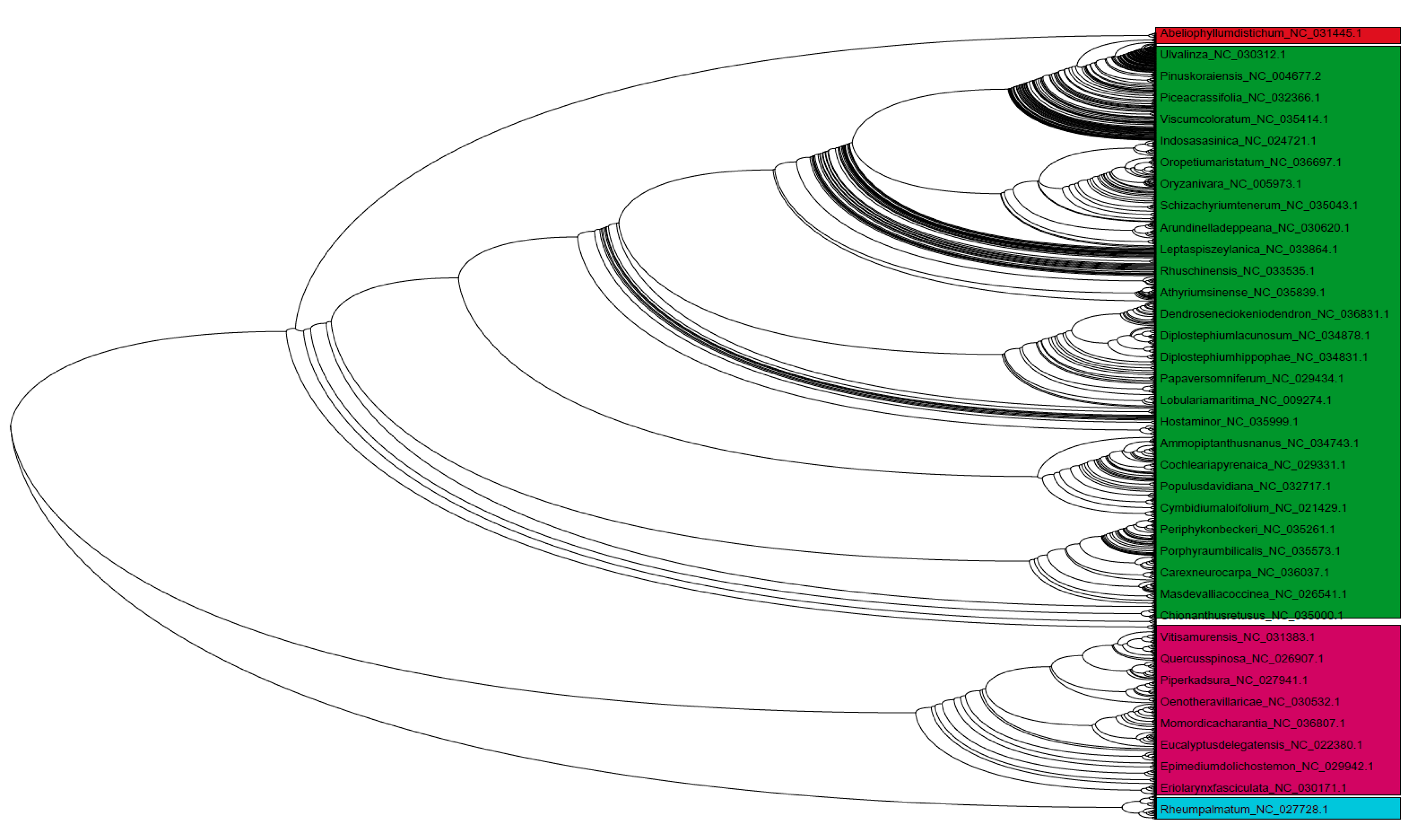
3.9. Chloroplast Genomes Have Evolved from Multiple Common Ancestral Nodes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barber, J.; Andersson, B. Revealing the blueprint of photosynthesis. Nature 1994, 370, 31–34. [Google Scholar] [CrossRef]
- Mishra, B.S.; Singh, M.; Aggrawal, P.; Laxmi, A. Glucose and Auxin Signaling Interaction in Controlling Arabidopsis thaliana Seedlings Root Growth and Development. PLoS ONE 2009, 4, e4502. [Google Scholar] [CrossRef] [PubMed]
- Moore, B.; Zhou, L.; Rolland, F.; Hall, Q.; Cheng, W.-H.; Liu, Y.-X.; Hwang, I.; Jones, T.; Sheen, J. Role of the Arabidopsis Glucose Sensor HXK1 in Nutrient, Light, and Hormonal Signaling. Science 2003, 300, 332–336. [Google Scholar] [CrossRef] [Green Version]
- Osteryoung, K.W.; Weber, A.P.M. Plastid Biology: Focus on the Defining Organelle of Plants. Plant Physiol. 2011, 155, 1475–1476. [Google Scholar] [CrossRef] [Green Version]
- Hedges, S.B.; Blair, J.E.; Venturi, M.L.; Shoe, J.L. A molecular timescale of eukaryote evolution and the rise of complex multicellular life. BMC Evol. Biol. 2004, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, H.S.; Hackett, J.D.; Ciniglia, C.; Pinto, G.; Bhattacharya, D. A Molecular Timeline for the Origin of Photosynthetic Eukaryotes. Mol. Biol. Evol. 2004, 21, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, S.P. The Chloroplasts of Some Algal Groups May Have Evolved from Endosymbiotic Eukaryotic Algae. Ann. N. Y. Acad. Sci. 1981, 361, 193–208. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Pudake, R.N.; Bae, H. Genome-wide identification of major protein families of cyanobacteria and genomic insight into the circadian rhythm. Eur. J. Phycol. 2017, 52. [Google Scholar] [CrossRef]
- Raven, J.A.; Allen, J.F. Genomics and chloroplast evolution: What did cyanobacteria do for plants? Gen. Biol. 2003, 4, 209. [Google Scholar] [CrossRef] [Green Version]
- Cavalier-Smith, T. Membrane heredity and early chloroplast evolution. Trends Plant Sci. 2000, 5, 174–182. [Google Scholar] [CrossRef]
- Kirk, P.R.; Leech, R.M. Amino Acid Biosynthesis by Isolated Chloroplasts during Photosynthesis. Plant Physiol. 1972, 50, 228–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noctor, G.; Arisi, A.-C.M.; Jouanin, L.; Foyer, C.H. Manipulation of Glutathione and Amino Acid Biosynthesis in the Chloroplast. Plant Physiol. 1998, 118, 471–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawke, J.C.; Rumsby, M.G.; Leech, R.M. Lipid biosynthesis by chloroplasts isolated from developing Zea mays. Phytochemistry 1974, 13, 403–413. [Google Scholar] [CrossRef]
- Britton, G. Biosynthesis of Chloroplast Carotenoids. In Current Research in Photosynthesis; Baltscheffsky, M., Ed.; Springer: Dordrecht, The Netherlands, 1990; pp. 2733–2740. ISBN 978-94-009-0511-5. [Google Scholar]
- García-Cerdán, J.G.; Schmid, E.M.; Takeuchi, T.; McRae, I.; McDonald, K.L.; Yordduangjun, N.; Hassan, A.M.; Grob, P.; Xu, C.S.; Hess, H.F.; et al. Chloroplast Sec14-like 1 (CPSFL1) is essential for normal chloroplast development and affects carotenoid accumulation in Chlamydomonas. Proc. Natl. Acad. Sci. USA 2020, 117, 12452–12463. [Google Scholar] [CrossRef] [PubMed]
- Ries, F.; Herkt, C.; Willmund, F. Co-translational protein folding and sorting in chloroplasts. Plants 2020, 9, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Liu, J.; Hao, G.; Zhang, L.; Mao, K.; Wang, X.; Zhang, D.; Ma, T.; Hu, Q.; Al-Shehbaz, I.A.; et al. Plastome phylogeny and early diversification of Brassicaceae. BMC Genom. 2017, 18, 176. [Google Scholar] [CrossRef] [Green Version]
- Graham, S.W.; Lam, V.K.Y.; Merckx, V.S.F.T. Plastomes on the edge: The evolutionary breakdown of mycoheterotroph plastid genomes. New Phytol. 2017, 214, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Gruenstaeudl, M.; Jenke, N. PACVr: Plastome assembly coverage visualization in R. BMC Bioinform. 2020, 21, 1–21. [Google Scholar] [CrossRef]
- Darshetkar, A.M.; Datar, M.N.; Tamhankar, S.; Li, P.; Choudhary, R.K. Understanding evolution in Poales: Insights from Eriocaulaceae plastome. PLoS ONE 2019, 14, e0221423. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, M. The chloroplast genome. In 10 Years Plant Molecular Biology; Schilperoort, R.A., Dure, L., Eds.; Springer: Dordrecht, The Netherlands, 1992; pp. 149–168. ISBN 978-94-011-2656-4. [Google Scholar]
- Yu, Q.-B.; Huang, C.; Yang, Z.-N. Nuclear-encoded factors associated with the chloroplast transcription machinery of higher plants. Front. Plant Sci. 2014, 5, 316. [Google Scholar] [CrossRef] [Green Version]
- Wicke, S.; Schneeweiss, G.M.; dePamphilis, C.W.; Müller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolodner, R.; Tewari, K.K. Inverted repeats in chloroplast DNA from higher plants. Proc. Natl. Acad. Sci. USA 1979, 76, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Wolf, P.G.; Der, J.P.; Duffy, A.M.; Davidson, J.B.; Grusz, A.L.; Pryer, K.M. The evolution of chloroplast genes and genomes in ferns. Plant Mol. Biol. 2011, 76, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Raubeson, L.; Jasen, R. Chloroplast genomes of plants. In Diversity and Evolution of Plants—Genotypic and Phenotypic Variation in Higher Plants; Henry, R., Ed.; CABI Publishing: Wallingford, UK, 2005; pp. 45–68. [Google Scholar]
- Wu, C.S.; Lai, Y.T.; Lin, C.P.; Wang, Y.N.; Chaw, S.M. Evolution of reduced and compact chloroplast genomes (cpDNAs) in gnetophytes: Selection toward a lower-cost strategy. Mol. Phylogenet. Evol. 2009, 52. [Google Scholar] [CrossRef]
- Daniell, H.; Lin, C.-S.; Yu, M.; Chang, W.-J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef] [Green Version]
- Rivas, J.D.L.; Lozano, J.J.; Ortiz, A.R. Comparative Analysis of Chloroplast Genomes: Functional Annotation, Genome-Based Phylogeny, and Deduced Evolutionary Patterns. Genome Res. 2002, 12, 567–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, D.B.; Goldschmidt-Clermont, M.; Hanson, M.R. Chloroplast RNA Metabolism. Ann. Rev. Plant Biol. 2010, 61, 125–155. [Google Scholar] [CrossRef]
- Cullis, C.A.; Vorster, B.J.; Van Der Vyver, C.; Kunert, K.J. Transfer of genetic material between the chloroplast and nucleus: How is it related to stress in plants? Ann. Bot. 2009, 103, 625–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckardt, N.A. Genomic Hopscotch: Gene Transfer from Plastid to Nucleus. Plant Cell 2006, 18, 2865–2867. [Google Scholar] [CrossRef] [Green Version]
- Turmel, M.; Otis, C.; Lemieux, C. The Chloroplast Genome Sequence of Chara vulgaris Sheds New Light into the Closest Green Algal Relatives of Land Plants. Mol. Biol. Evol. 2006, 23, 1324–1338. [Google Scholar] [CrossRef]
- Gao, L.; Su, Y.-J.; Wang, T. Plastid genome sequencing, comparative genomics, and phylogenomics: Current status and prospects. J. Syst. Evol. 2010, 48, 77–93. [Google Scholar] [CrossRef]
- Downie, S.R.; Palmer, J.D. Restriction Site Mapping of the Chloroplast DNA Inverted Repeat: A Molecular Phylogeny of the Asteridae. Ann. Mo. Bot. Gard. 1992, 79, 266–283. [Google Scholar] [CrossRef] [Green Version]
- Goulding, S.E.; Wolfe, K.H.; Olmstead, R.G.; Morden, C.W. Ebb and flow of the chloroplast inverted repeat. Mol. Gen. Genet. MGG 1996, 252, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Plunkett, G.M.; Downie, S.R. Expansion and Contraction of the Chloroplast Inverted Repeat in Apiaceae Subfamily Apioideae. Syst. Bot. 2000, 25, 648–667. [Google Scholar] [CrossRef]
- Guisinger, M.M.; Kuehl, J.V.; Boore, J.L.; Jansen, R.K. Extreme Reconfiguration of Plastid Genomes in the Angiosperm Family Geraniaceae: Rearrangements, Repeats, and Codon Usage. Mol. Biol. Evol. 2011, 28, 583–600. [Google Scholar] [CrossRef] [Green Version]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acid. Res. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq-versatile and accurate annotation of organelle genomes. Nucleic Acid. Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef] [PubMed]
- Stothard, P. The Sequence Manipulation Suite: JavaScript Programs for Analyzing and Formatting Protein and DNA Sequences. BioTechniques 2000, 28, 1102–1104. [Google Scholar] [CrossRef] [Green Version]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acid. Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef]
- Tamura, K.; Filipski, A.; Peterson, D.; Stecher, G.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef] [PubMed]
- Stolzer, M.; Lai, H.; Xu, M.; Sathaye, D.; Vernot, B.; Durand, D. Inferring duplications, losses, transfers and incomplete lineage sorting with nonbinary species trees. Bioinformatics 2012, 28, i409–i415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darby, C.A.; Stolzer, M.; Ropp, P.J.; Barker, D.; Durand, D. Xenolog classification. Bioinformatics 2017, 33, 640–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Durand, D.; Farach-Colton, M. NOTUNG: A Program for Dating Gene Duplications and Optimizing Gene Family Trees. J. Comput. Biol. 2000, 7, 429–447. [Google Scholar] [CrossRef]
- Vaughan, T.G. IcyTree: Rapid browser-based visualization for phylogenetic trees and networks. Bioinformatics 2017, 33, 2392–2394. [Google Scholar] [CrossRef]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Zhang, R.; Ge, F.; Li, H.; Chen, Y.; Zhao, Y.; Gao, Y.; Liu, Z.; Yang, L. PCIR: A database of Plant Chloroplast Inverted Repeats. Database J. Biol. Databases Curation 2019, 2019, baz127. [Google Scholar] [CrossRef]
- Ma, J.; Yang, B.; Zhu, W.; Sun, L.; Tian, J.; Wang, X. The complete chloroplast genome sequence of Mahonia bealei (Berberidaceae) reveals a significant expansion of the inverted repeat and phylogenetic relationship with other angiosperms. Gene 2013, 528, 120–131. [Google Scholar] [CrossRef]
- Bryant, N.; Lloyd, J.; Sweeney, C.; Myouga, F.; Meinke, D. Identification of Nuclear Genes Encoding Chloroplast-Localized Proteins Required for Embryo Development in Arabidopsis. Plant Physiol. 2011, 155, 1678. [Google Scholar] [CrossRef] [Green Version]
- Savage, L.J.; Imre, K.M.; Hall, D.A.; Last, R.L. Analysis of Essential Arabidopsis Nuclear Genes Encoding Plastid-Targeted Proteins. PLoS ONE 2013, 8, e73291. [Google Scholar] [CrossRef] [Green Version]
- Taylor, D.L. Chloroplasts as Symbiotic Organelles. In International Review of Cytology; Bourne, G.H., Danlelli, J.F., Eds.; JeonAcademic Press: New York, NY, USA; London, UK, 1970; Volume 27, pp. 29–64. ISBN 0074-7696. [Google Scholar]
- Trench, R.K. Chloroplasts: Presumptive and de facto organelles. Ann. N. Y. Acad. Sci. 1981, 361, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Stern, D.S.; Higgs, D.C.; Yang, J. Transcription and translation in chloroplasts. Trends Plant Sci. 1997, 2, 308–315. [Google Scholar] [CrossRef]
- Turmel, M.; Otis, C.; Lemieux, C. Divergent copies of the large inverted repeat in the chloroplast genomes of ulvophycean green algae. Sci. Rep. 2017, 7, 994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfe, K.H. The site of deletion of the inverted repeat in pea chloroplast DNA contains duplicated gene fragments. Curr. Genet. 1988, 13, 97–99. [Google Scholar] [CrossRef] [PubMed]
- Strauss, S.H.; Palmer, J.D.; Howe, G.T.; Doerksen, A.H. Chloroplast genomes of two conifers lack a large inverted repeat and are extensively rearranged. Proc. Natl. Acad. Sci. USA 1988, 85, 3898–3902. [Google Scholar] [CrossRef] [Green Version]
- Zhu, A.; Guo, W.; Gupta, S.; Fan, W.; Mower, J.P. Evolutionary dynamics of the plastid inverted repeat: The effects of expansion, contraction, and loss on substitution rates. New Phytol. 2016, 209, 1747–1756. [Google Scholar] [CrossRef] [Green Version]
- Palmer, J.; Osorio, B.; Aldrich, J.; Thompson, W. Chloroplast DNA evolution among legumes: Loss of a large inverted repeat occurred prior to other sequence rearrangements. Curr. Genet. 1987, 11, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Lemieux, C.; Otis, C.; Turmel, M. Comparative Chloroplast Genome Analyses of Streptophyte Green Algae Uncover Major Structural Alterations in the Klebsormidiophyceae, Coleochaetophyceae and Zygnematophyceae. Front. Plant Sci. 2016, 7, 697. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, C.; Kobayashi, Y.; Aoki, S.; Sugita, C.; Sugita, M. Complete chloroplast DNA sequence of the moss Physcomitrella patens: Evidence for the loss and relocation of rpoA from the chloroplast to the nucleus. Nucleic Acid. Res. 2003, 31, 5324–5331. [Google Scholar] [CrossRef]
- Sinn, B.T.; Sedmak, D.D.; Kelly, L.M.; Freudenstein, J.V. Total duplication of the small single copy region in the angiosperm plastome: Rearrangement and inverted repeat instability in Asarum. Am. J. Bot. 2018, 105, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Alverson, A.J.; Ruck, E.C.; Theriot, E.C.; Nakov, T.; Jansen, R.K. Serial Gene Losses and Foreign DNA Underlie Size and Sequence Variation in the Plastid Genomes of Diatoms. Genome Biol. Evol. 2014, 6, 644–654. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, A.D.; dePamphilis, C.W. The effect of relaxed functional constraints on the photosynthetic gene rbcL in photosynthetic and nonphotosynthetic parasitic plants. Mol. Biol. Evol. 1998, 15, 1243–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wicke, S.; Müller, K.F.; de Pamphilis, C.W.; Quandt, D.; Wickett, N.J.; Zhang, Y.; Renner, S.S.; Schneeweiss, G.M. Mechanisms of functional and physical genome reduction in photosynthetic and nonphotosynthetic parasitic plants of the broomrape family. Plant Cell 2013, 25, 3711–3725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, A.L.; Friedman, R. Genome Size Reduction in the Chicken Has Involved Massive Loss of Ancestral Protein-Coding Genes. Mol. Biol. Evol. 2008, 25, 2681–2688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, G.; Xu, Y.; Liu, H.; Sun, T.; Zhang, J.; Hettenhausen, C.; Shen, G.; Qi, J.; Qin, Y.; Li, J.; et al. Large-scale gene losses underlie the genome evolution of parasitic plant Cuscuta australis. Nat. Commun. 2018, 9, 2683. [Google Scholar] [CrossRef] [Green Version]
- Martín, M.; Sabater, B. Plastid ndh genes in plant evolution. Plant Physiol. Biochem. 2010, 48, 636–645. [Google Scholar] [CrossRef]
- Lin, C.-S.; Chen, J.J.W.; Huang, Y.-T.; Chan, M.-T.; Daniell, H.; Chang, W.-J.; Hsu, C.-T.; Liao, D.-C.; Wu, F.-H.; Lin, S.-Y.; et al. The location and translocation of ndh genes of chloroplast origin in the Orchidaceae family. Sci. Rep. 2015, 5, 9040. [Google Scholar] [CrossRef] [Green Version]
- Heckenhauer, J.; Barfuss, M.H.J.; Samuel, R. Universal multiplexable matK primers for DNA barcoding of angiosperms. Appl. Plant Sci. 2016, 4, apps.1500137. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Xue, J.H.; Zhou, S.L. New universal matK primers for DNA barcoding angiosperms. J. Syst. Evol. 2011, 49, 176–181. [Google Scholar] [CrossRef]
- Hollingsworth, P.; Forrest, L.L.; Spouge, J.L.; Hajibabaei, M.; Ratnasingham, S.; van der Bank, M.; Chase, M.W.; Cowan, R.S.; Erickson, D.L.; Fazekas, A.J.; et al. A DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar] [CrossRef] [Green Version]
- Li, F.-W.; Kuo, L.-Y.; Rothfels, C.J.; Ebihara, A.; Chiou, W.-L.; Windham, M.D.; Pryer, K.M. rbcL and matK Earn Two Thumbs Up as the Core DNA Barcode for Ferns. PLoS ONE 2011, 6, e26597. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Xu, C.; Li, C.; Sun, J.; Zuo, Y.; Shi, S.; Cheng, T.; Guo, J.; Zhou, S. ycf1, the most promising plastid DNA barcode of land plants. Sci. Rep. 2015, 5, 8348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goffinet, B.; Wickett, N.J.; Shaw, A.J.; Cox, C.J. Phylogenetic significance of the rpoA loss in the chloroplast genome of mosses. Taxon 2005, 54, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Gabaldón, T.; Koonin, E.V. Functional and evolutionary implications of gene orthology. Nat. Rev. Genet. 2013, 14, 360–366. [Google Scholar] [CrossRef] [Green Version]
- Jensen, R.A. Orthologs and paralogs—We need to get it right. Genome Biol. 2001, 2, interactions1002. [Google Scholar] [CrossRef] [Green Version]
- Sonnhammer, E.L.L.; Koonin, E.V. Orthology, paralogy and proposed classification for paralog subtypes. Trends Genet. 2002, 18, 619–620. [Google Scholar] [CrossRef]
- Palenik, B.; Grimwood, J.; Aerts, A.; Rouzé, P.; Salamov, A.; Putnam, N.; Dupont, C.; Jorgensen, R.; Derelle, E.; Rombauts, S.; et al. The tiny eukaryote Ostreococcus provides genomic insights into the paradox of plankton speciation. Proc. Natl. Acad. Sci. USA 2007, 104, 7705–7710. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, J.Y.; Bauer, C.E. Light-Independent Chlorophyll Biosynthesis: Involvement of the Chloroplast Gene chlL (frxC). Plant Cell 2007, 4, 929. [Google Scholar] [CrossRef]
- Fujita, Y.; Takagi, H.; Hase, T. Identification of the chlB gene and the gene product essential for the light-independent chlorophyll biosynthesis in the cyanobacterium Plectonema boryanum. Plant Cell Physiol. 1996, 37, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Yu, J.; Zhao, N. Partial recovery of light-independent chlorophyll biosynthesis in the chlL-deletion mutant of Synechocystis sp. PCC 6803. IUBMB Life 2001, 51, 289–293. [Google Scholar] [CrossRef]
- Burke, D.H.; Raubeson, L.A.; Alberti, M.; Hearst, J.E.; Jordan, E.T.; Kirch, S.A.; Valinski, A.E.C.; Conant, D.S.; Stein, D.B. The chlL (frxC) gene: Phylogenetic distribution in vascular plants and DNA sequence from Polystichum acrostichoides (Pteridophyta) and Synechococcus sp. 7002 (Cyanobacteria). Plant Syst. Evol. 1993, 187, 89–102. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, M.; Wakasugi, T.; Yoshinaga, K.; Sugiura, M. The chloroplast chlL gene of the green alga Chlorella vulgaris C-27 contains a self-splicing group I intron. Mol. Gener. Gen. 1996, 250, 655–664. [Google Scholar] [CrossRef]
- Karpinska, B.; Karpinski, S.; Hällgren, J.E. The chlB gene encoding a subunit of light-independent protochlorophyllide reductase is edited in chloroplasts of conifers. Curr. Gen. 1997, 31, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Stegemann, S.; Hartmann, S.; Ruf, S.; Bock, R. High-frequency gene transfer from the chloroplast genome to the nucleus. Proc. Natl. Acad. Sci. USA 2003, 100, 8828–8833. [Google Scholar] [CrossRef] [Green Version]
- Baldauf, S.L.; Palmer, J.D. Evolutionary transfer of the chloroplast tufA gene to the nucleus. Nature 1990, 344, 262–265. [Google Scholar] [CrossRef] [Green Version]
- Martin, W.; Herrmann, R.G. Gene Transfer from Organelles to the Nucleus: How Much, What Happens, and Why? Plant Physiol. 1998, 118, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of the Gene | Total No. of Sequences Studied | No. of Duplication | No. of Codivergence | No. of Losses | Transfer |
|---|---|---|---|---|---|
| PsaM | 84 | 12 (14.28%) | 37 (44.04%) | 34 (40.47%) | 0 |
| Psb30 | 157 | 39 (24.84) | 49 (31.21%) | 120 (76.43%) | 0 |
| ChlB | 288 | 35 (12.15%) | 116 (40.27%) | 126 (43.75%) | 0 |
| ChlL | 283 | 49 (17.31%) | 100 (35.33%) | 184 (65.01%) | 0 |
| ChlN | 83 | 8 (9.63%) | 34 (40.47%) | 46 (55.42%) | 0 |
| Rpl21 | 22 | 3 (13.63%) | 9 (40.90%) | 8 (36.36%) | 0 |
| rpoA | rpoB | rpoC1 | rpoC2 | ||||||||||||
| 26 | 19 | 21 | 13 | ||||||||||||
| atpA | atpB | atpE | atpF | atpH | atpI | ||||||||||
| 8 | 8 | 12 | 14 | 13 | 12 | ||||||||||
| accD | ccsA | cemA | chlB | chlL | ChlN | ||||||||||
| 387 | 29 | 29 | 2054 | 2062 | 2066 | ||||||||||
| ClpP | Rbcl | Ycf1 | Ycf2 | Ycf3 | Ycf4 | ||||||||||
| 142 | 19 | 161 | 219 | 30 | 39 | ||||||||||
| ndhA | ndhB | ndhC | ndhD | ndhE | ndhF | ndhG | NdhH | ndhI | ndhJ | ndhK | |||||
| 339 | 258 | 339 | 293 | 322 | 346 | 335 | 322 | 378 | 340 | 331 | |||||
| petA | petB | PetD | petG | petL | petN | ||||||||||
| 33 | 15 | 36 | 13 | 71 | 135 | ||||||||||
| psaA | psaB | psaC | psaI | psaJ | psaM | ||||||||||
| 16 | 10 | 19 | 72 | 24 | 2214 | ||||||||||
| psbA | psbB | psbC | psbD | psbE | psbF | psbH | psbI | psbJ | psbK | psbL | psbM | psbN | psbT | psbZ | Psb30 |
| 12 | 18 | 16 | 17 | 21 | 21 | 20 | 18 | 21 | 13 | 22 | 157 | 23 | 22 | 31 | 2126 |
| Rpl2 | Rpl14 | Rpl16 | Rpl20 | Rpl22 | Rpl23 | Rpl32 | Rpl33 | Rpl36 | |||||||
| 2 | 4 | 3 | 0 | 127 | 24 | 114 | 133 | 5 | |||||||
| Rps2 | Rps3 | Rps4 | Rps7 | Rps8 | Rps11 | Rps12 | Rps14 | Rps15 | Rps16 | Rps18 | Rps19 | ||||
| 3 | 3 | 4 | 3 | 3 | 2 | 2 | 7 | 249 | 284 | 5 | 5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohanta, T.K.; Mishra, A.K.; Khan, A.; Hashem, A.; Abd_Allah, E.F.; Al-Harrasi, A. Gene Loss and Evolution of the Plastome. Genes 2020, 11, 1133. https://doi.org/10.3390/genes11101133
Mohanta TK, Mishra AK, Khan A, Hashem A, Abd_Allah EF, Al-Harrasi A. Gene Loss and Evolution of the Plastome. Genes. 2020; 11(10):1133. https://doi.org/10.3390/genes11101133
Chicago/Turabian StyleMohanta, Tapan Kumar, Awdhesh Kumar Mishra, Adil Khan, Abeer Hashem, Elsayed Fathi Abd_Allah, and Ahmed Al-Harrasi. 2020. "Gene Loss and Evolution of the Plastome" Genes 11, no. 10: 1133. https://doi.org/10.3390/genes11101133
APA StyleMohanta, T. K., Mishra, A. K., Khan, A., Hashem, A., Abd_Allah, E. F., & Al-Harrasi, A. (2020). Gene Loss and Evolution of the Plastome. Genes, 11(10), 1133. https://doi.org/10.3390/genes11101133