Determinants of Copper Resistance in Acidithiobacillus Ferrivorans ACH Isolated from the Chilean Altiplano

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Growth Conditions
2.2. Search for Copper Resistance Genetic Determinants in the ACH Strain Genome
2.3. Total RNA Extraction and cDNA Synthesis from Copper-Cultured A. Ferrivorans ACH
2.4. Relative Gene Expression Quantification
3. Results and Discussion
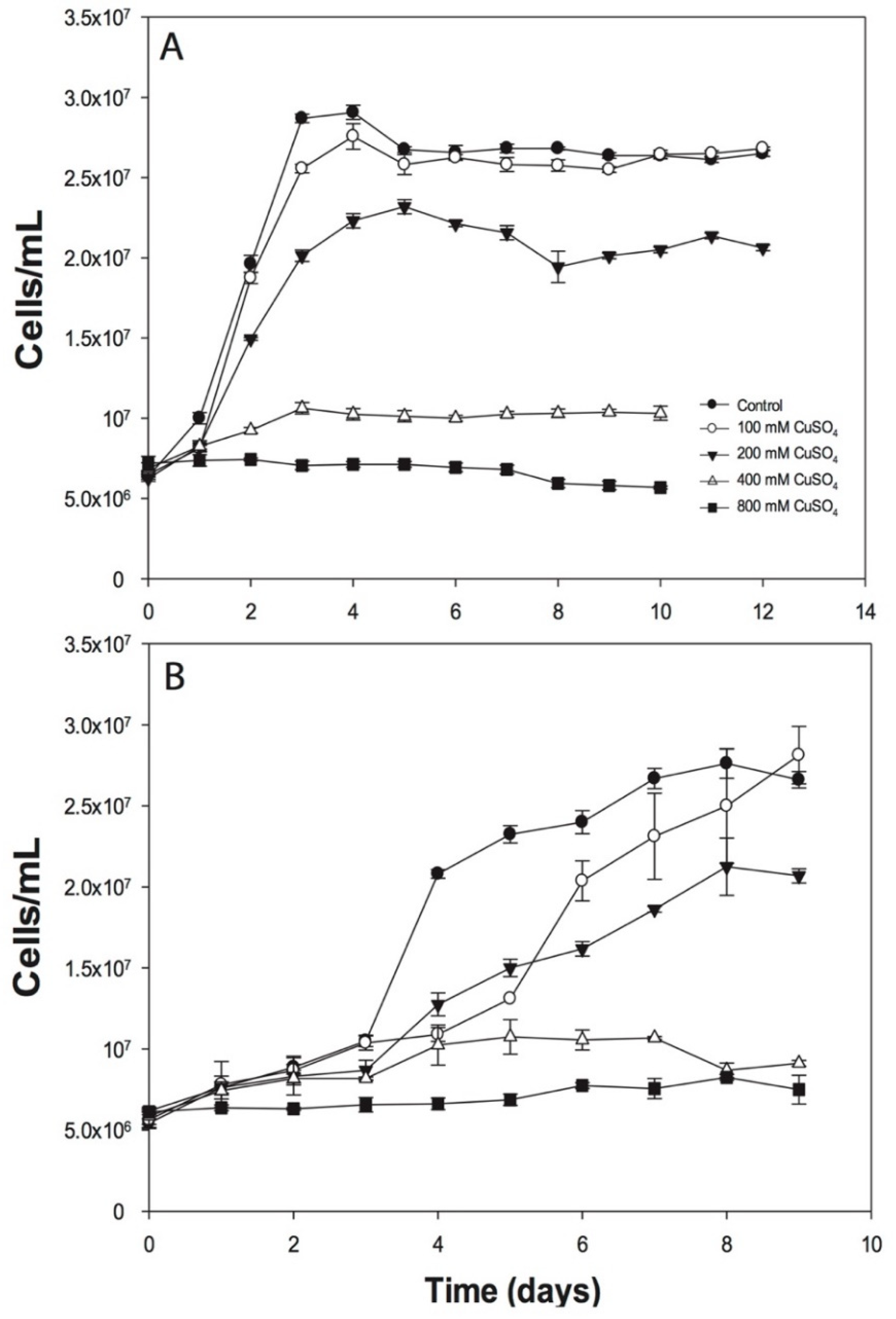
3.1. Effects of Copper on ACH Strain Growth
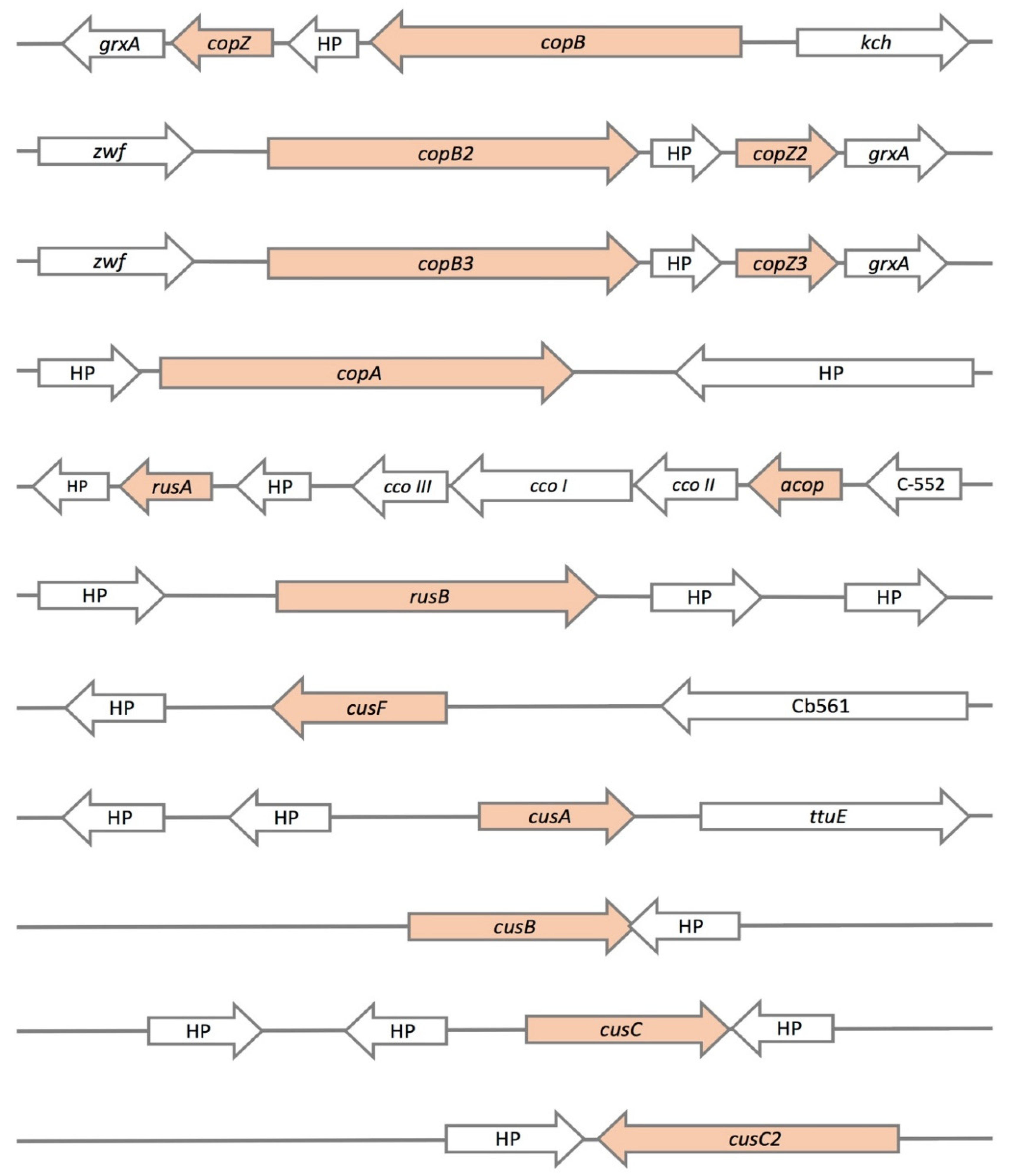
3.2. Genetic Determinants of Copper Resistance in ACH Strain
3.3. Comparison of Copper Resistance Genes Identified in A. Ferrivorans Strains
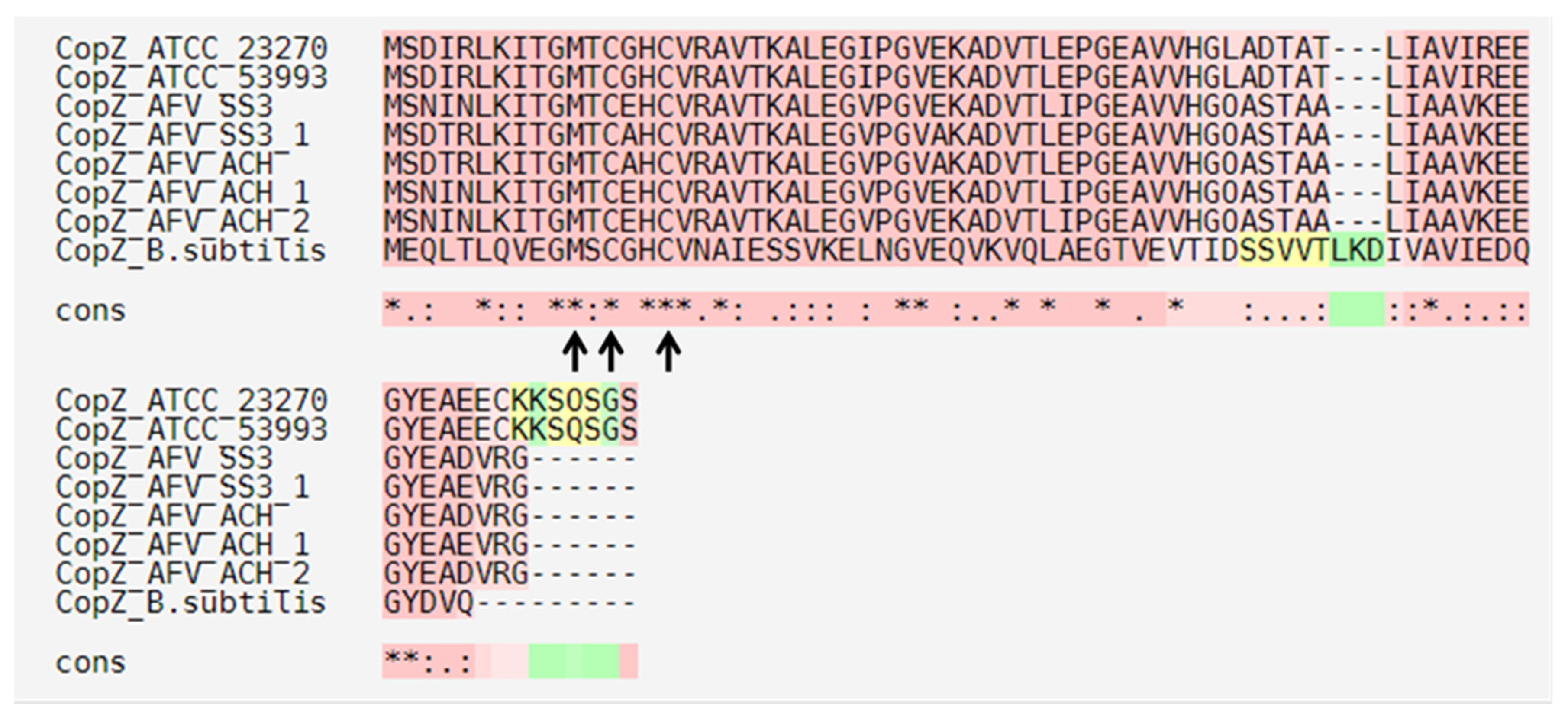
3.4. Conserved Amino Acid Motifs in ACH Strain Copper Resistance Proteins
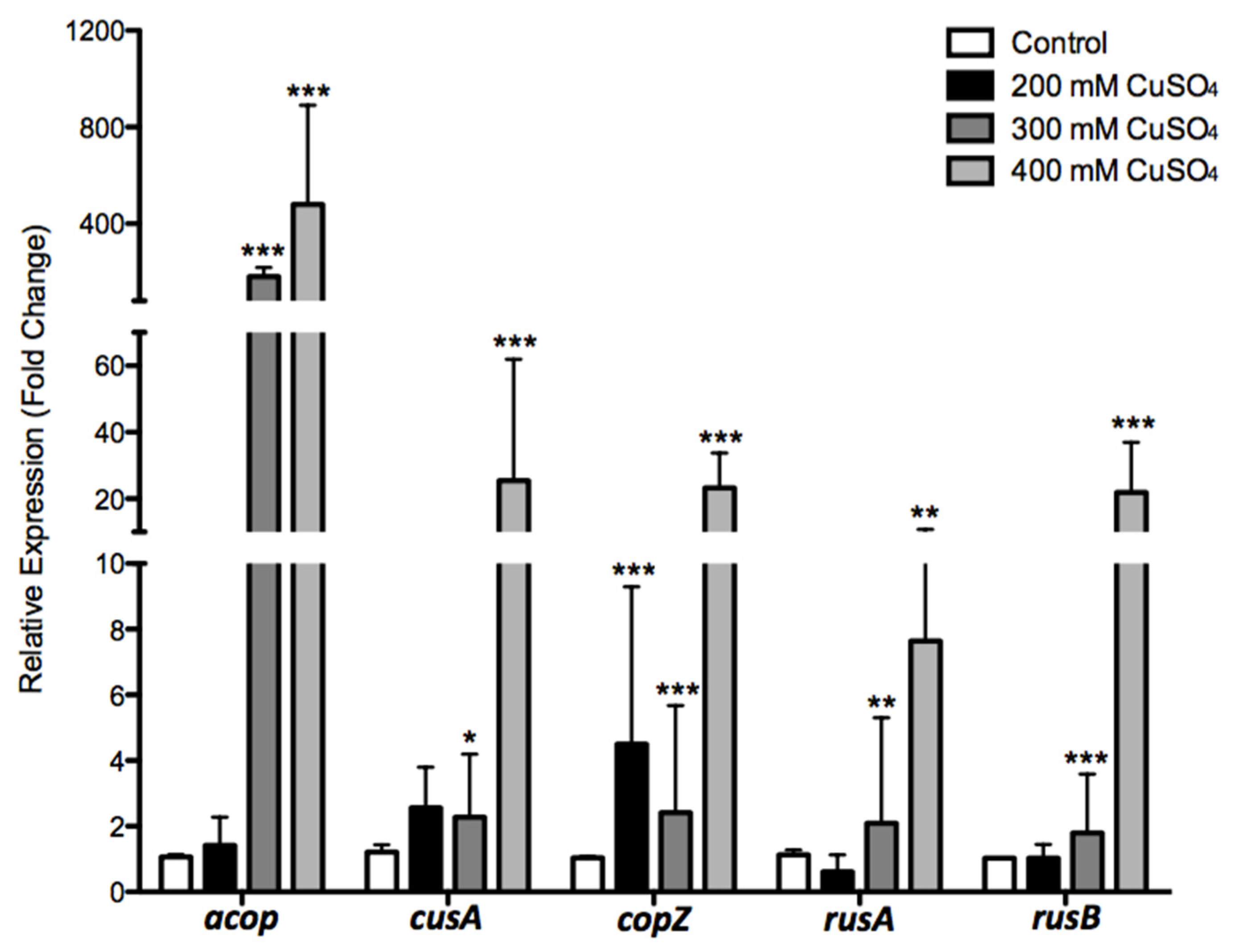
3.5. Effects of Copper in A. Ferrivorans ACH Gene Expression of Resistance Markers
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Olson, G.; Brierley, J.; Brierley, C. Bioleaching review part B: Progress in Bioleaching: Applications of microbial processes by the mineral industries. Appl. Microbiol. Biotechnol. 2003, 63, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Brierley, C. Chapter 6: Biological Processing: Biological Processing of Sulfidic Ores and Concentrates—Integrating Innovations. In Innovative Process Development in Metallurgical Industry; Lakshmanan, V., Roy, R., Ramachandran, V., Eds.; Springer: Cham, Switzerland, 2016; pp. 109–135. [Google Scholar]
- Dopson, M.; Baker-Austin, C.; Koppineedi, P.; Bond, P.L. Growth in sulfidic mineral environments: Metal resistance mechanisms in acidophilic microorganism. Microbiology 2003, 149, 1959–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watkin, E.; Keeling, S.; Perrot, F.; Shiers, D.; Palmer, M.; Watling, H. Metals tolerance in moderately thermophilic isolates from a spent copper sulfide heap, closely related to Acidithiobacillus caldus, Acidimicrobium ferrooxidans and Sulfobacillus thermosulfidooxidans. J. Ind. Microbiol. Biotechnol. 2008, 36, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Navarro, C.; von Bernath, D.; Jerez, C. Heavy Metal Resistance Strategies of Acidophilic Bacteria and Their Acquisition: Importance for Biomining and Bioremediation. Biol. Res. 2013, 46, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Nies, D. Microbial heavy-metal resistance. Appl. Microbiol. Biotechnol. 1999, 51, 730–750. [Google Scholar] [CrossRef]
- Franke, S.; Rensing, C. Chapters 21: Acidophiles: Mechanisms to tolerate metal and acid toxicity. In Physiology and Biochemistry of Extremophiles; Gerday, C., Glansdorff, N., Eds.; ASM Press: Washington, DC, USA, 2007; pp. 271–278. [Google Scholar]
- Orell, A.; Navarro, C.; Arancibia, R.; Mobarec, J.; Jerez, C. Life in blue: Copper resistance mechanisms of bacteria and Archaea used in industrial biomining of minerals. Biotechnol. Adv. 2010, 28, 839–848. [Google Scholar] [CrossRef]
- Dopson, M.; Holmes, D. Metal resistance in acidophilic microorganisms and its significance for biotechnologies. Appl. Microbiol. Biotechnol. 2014, 98, 8133–8144. [Google Scholar] [CrossRef]
- di Toro, D.; Allen, H.; Bergman, H.; Meyer, S.; Paquin, P.; Santore, R. Biotic ligand model of the acute toxicity of metals. Environ. Toxicol. Chem. 2001, 20, 2383–2396. [Google Scholar] [CrossRef]
- Harrison, J.; Ceri, H.; Turner, R. Multimetal resistance and tolerance in microbial biofilms. Nat. Rev. Microbiol. 2007, 5, 928–938. [Google Scholar] [CrossRef]
- Palmgren, M.; Nissen, P. P-type ATPasa. Annu. Rev. Biophys. 2011, 40, 243–266. [Google Scholar] [CrossRef]
- Tseng, T.; Gratwick, K.; Kollman, J.; Park, D.; Nies, D.; Goffeau, A.; Saier, M. The RND permease superfamily: An ancient ubiquitous and diverse family that includes human disease and development proteins. J. Mol. Microbiol. Biotechnol. 1999, 1, 107–125. [Google Scholar] [PubMed]
- Orell, A.; Navarro, C.; Rivero, M.; Aguilar, J.; Jerez, C. Inorganic polyphosphates in extremophiles and their possible functions. Extremophiles 2012, 16, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Bonnefoy, V.; Holmes, D. Minireview: Genomic insights into microbial iron oxidation and iron uptake strategies in extremely acidic environments. Environ. Microbiol. 2012, 14, 1597–1611. [Google Scholar] [CrossRef] [PubMed]
- Dew, D.; Muhlbauer, R.; van Buuren, C. Bioleaching of copper sulphide concentrates with mesophiles and thermophiles. In Biohydrometallurgy and the Environment Toward the Mining of the 21st Century; Elsevier Science: Madrid, Spain, 1999; ISBN 9780080527956. [Google Scholar]
- Watling, H. The bioleaching of sulphide minerals with emphasis on copper sulphides—A review. Hydrometallurgy 2006, 84, 81–108. [Google Scholar] [CrossRef]
- Martínez-Bussenius, C.; Navarro, C.; Jerez, C. Minireview: Microbial copper resistance: Importance in biohydrometallurgy. Microb. Biotechnol. 2016, 10, 279–295. [Google Scholar] [CrossRef] [Green Version]
- Navarro, C.; Orellana, L.; Mauriaca, C.; Jerez, C. Transcriptional and functional studies of Acidithiobacillus ferrooxidans genes related to survival in the presence of copper. Appl. Environ. Microbiol. 2009, 75, 6102–6109. [Google Scholar] [CrossRef] [Green Version]
- Chi, A.; Valenzuela, L.; Beard, S.; Mackey, A.; Shabanowitz, J.; Hunt, D.; Jerez, C. Periplasmic proteins of the extremophile Acidithiobacillus ferrooxidans: A high throughput proteomics analysis. Mol. Cell. Proteom. 2007, 6, 2239–2251. [Google Scholar] [CrossRef] [Green Version]
- Hallberg, K.; González-Toril, E.; Johnson, D. Acidithiobacillus ferrivorans, sp. nov.; facultatively anaerobic, psychrotolerant iron-, and sulfur-oxidizing acidophiles isolated from metal mine-impacted environments. Extremophiles 2010, 14, 9–19. [Google Scholar] [CrossRef]
- Talla, E.; Hedrich, S.; Mangenot, S.; Ji, B.; Johnson, D.; Barbe, V.; Bonnefoy, V. Insights into the pathways of iron-and-sulfur-oxidation, and biofilm formation from the chemolithotrophic acidophile Acidithiobacillus ferrivorans CF27. Res. Microbiol. 2014, 165, 753–760. [Google Scholar] [CrossRef]
- Peng, T.; Ma, L.; Feng, X.; Tao, J.; Nan, M.; Liu, Y.; Li, J.; Li, S.; Xu, X.; Yu, R.; et al. Genomic and transcriptomic analyses reveal adaptation mechanisms of an Acidithiobacillus ferrivorans strain YL15 to alpine acid mine drainage. PLoS ONE 2017, 12, e0178008. [Google Scholar] [CrossRef]
- Ccorahua-Santo, R.; Eca, A.; Abanto, M.; Guerra, G.; Ramirez, P. Physiological and comparative genomic analysis of Acidithiobacillus ferrivorans PQ33 provides psychrotolerant fitness evidence for oxidation at low temperature. Res. Microbiol. 2017, 168, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Barahona, S.; Dorador, C.; Remonsellez, F. Identification and characterization of a psychrotolerant Acidithiobacillus strain from Chilean Altiplano. Adv. Mater. Res. 2013, 825, 74–78. [Google Scholar] [CrossRef]
- González, C.; Yanquepe, M.; Cardenas, J.; Valdes, J.; Quatrini, R.; Holmes, D.; Dopson, M. Genetic variability of psychrotolerant Acidithiobacillus ferrivorans revealed by (meta)genomic analysis. Res. Microbiol. 2014, 165, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.; Mangenot, S.; Magdelenat, G.; Payen, E.; Rouy, Z.; Belahbib, H.; Grail, B.; Johnson, D.; Bonnefoy, V.; Talla, E. Comparative Genome Analysis Provides Insights into Both the Lifestyle of Acidithiobacillus ferrivorans Strain CF27 and the Chimeric Nature of the Iron-Oxidizing Acidithiobacilli Genomes. Front. Microbiol. 2017, 8, 1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dopson, M.; Halinen, A.; Rahunen, N.; Ozkaya, B.; Sahinkaya, E.; Kaksonen, A.; Lindstrom, E.; Puhakka, J. Mineral and Iron Oxidation at Low Temperatures by Pure and Mixed Cultures of Acidophilic Microorganisms. Biotechnol. Bioeng. 2007, 97, 1205–1215. [Google Scholar] [CrossRef]
- Escobar, B.; Buccicardi, S.; Morales, G.; Wiertz, J. Biooxidation of ferrous iron and sulphide at low temperatures: Implications on acid mine drainage and bioleaching of sulfide minerals. Hydrometallurgy 2010, 104, 454–458. [Google Scholar] [CrossRef]
- Kupka, D.; Liljeqvist, M.; Nurmi, P.; Puhakka, J.; Tuovinen, O.; Dopson, M. Oxidation of elemental sulfur, tetrathionate and ferrous iron by the psychrotolerant Acidithiobacillus strain SS3. Microbiology 2009, 160, 767–774. [Google Scholar] [CrossRef]
- Altschul, S.; Gish, W.; Miller, W.; Myers, E.; Lipman, D. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Jones, P.; Binns, D.; Chang, H.; Fraser, M.; Li, W.; Mcanulla, C.; Mcwilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [Green Version]
- Notredame, C.; Higgins, D.; Heringa, J. T-Coffee: A Novel Method for Fast and Accurate Multiple Sequence Alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.; Geer, R.; Gonzales, N.; Gwadz, M.; Hurwitz, D.; Marchler, G.; Song, J.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, 265–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passerini, M.; Lippi, A.; Frasconi, P. MetalDetector v2.0: Predicting the Geometry of Metal Binding Sites from Protein Sequence. Nucleic Acids Res. 2011, 39, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Vera, M.; Pagliai, F.; Guiliani, N.; Jerez, C. The chemolithoautotroph Acidithiobacillus ferrooxidans can survive under phosphate-limiting conditions by the expression of a C-P lyase operon that allows it to grow on phosphonates. Appl. Environ. Microbiol. 2008, 74, 1829–1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orellana, L.; Jerez, C. A genomic island provides Acidithiobacillus ferrooxidans ATCC 53993 additional copper resistance: A possible competitive advantage. Appl. Microbiol. Biotechnol. 2011, 92, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.; Remm, M.; Rozen, S. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, 115. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M. A new mathematical model for relative quantification in real-time RT PCR. Nucleic Acids Res. 2011, 29, 2002–2007. [Google Scholar] [CrossRef]
- Almárcegui, R.; Navarro, C.; Paradela, A.; Albar, J.; von Bernath, D.; Jerez, C. Response to copper of Acidithiobacillus ferrooxidans ATCC 23270 grown in elemental sulfur. Res. Microbiol. 2014, 165, 761–772. [Google Scholar] [CrossRef]
- D’ Amico, S.; Collins, T.; Marx, J.; Feller, G.; Gerday, C. Reviews: Psychrophilic microorganisms: Challenge for life. EMBO Rep. 2006, 7, 385–389. [Google Scholar] [CrossRef]
- Richard, D.; Ravigné, V.; Rieux, A.; Facon, B.; Boyer, C.; Boyer, K.; Grygiel, P.; Javegny, S.; Terville, M.; Canteros, B.; et al. Adaptation of genetically monomorphic bacteria: Evolution of copper resistance through multiple horizontal gene transfers of complex and versatile mobile genetic elements. Mol. Ecol. 2017, 26, 2131–2149. [Google Scholar] [CrossRef]
- Purves, J.; Thomas, J.; Riboldi, G.; Zapotoczna, M.; Tarrant, E.; Andrew, P.; Londoño, A.; Planet, P.; Geoghegan, J.; Waldron, K.; et al. A horizontally gene transferred copper resistance locus confers hyper-resistance to antibacterial copper toxicity and enables survival of community acquired methicillin resistant Staphylococcus aureus USA300 in macrophages. Environ. Microbiol. 2018, 20, 1576–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cárdenas, J.; Moya, F.; Covarrubias, P.; Shmaryahu, A.; Levicán, G.; Holmes, D.; Quatrini, R. Comparative genomics of the oxidative stress response in bioleaching microorgannisms. Hydrometallurgy 2012, 127, 162–167. [Google Scholar] [CrossRef]
- Si, M.; Zhang, L.; Yang, Z.; Xu, Y.; Liu, Y.; Jiang, C.; Wang, Y.; Shen, X.; Liu, S. NrdH Redoxin Enhances Resistance to Multiple Oxidative Stresses by Acting as a Peroxidase Cofactor in Corynebacterium glutamicum. Appl. Environ. Microbiol. 2014, 80, 1750–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imlay, J.; Chin, S.; Linn, S. Toxic DNA damage by hydrogen peroxide through the Fenton reaction in vivo and in vitro. Science 1988, 240, 640–642. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, A.; Orellana, O.; Levicán, G. Chapter 4: Oxidative stress and Metal Tolerance in Extreme Acidophiles. In Acidophiles, 1st ed.; Quatrini, R., Johnson, D., Eds.; Caister Academic Press: Santiago, Chile, 2016; pp. 63–76. [Google Scholar]
- Achour-Rokbani, A.; Cordi, A.; Poupin, P.; Bauda, P.; Billard, P. Characterization of the ars gene cluster from extremely arsenic-resistant Microbacterium sp. strain A33. Appl. Environ. Microbiol. 2010, 76, 948–955. [Google Scholar] [CrossRef] [Green Version]
- Castro-Severyn, J.; Pardo-Esté, C.; Bracho, S.; Noe, Y.; Cabezas, C.; Gariazzo, V.; Briones, A.; Morales, N.; Séveno, M.; Decourcelle, M.; et al. Arsenic response of three altiplanic Exiguobacterium strains with different tolerance levels against the metalloid species: A proteomics study. Front. Microbiol. 2019, 10, 2161. [Google Scholar] [CrossRef]
- Cox, M.; Battista, J. Deinococcus radiodurans—The consummate survivor. Nat. Rev. Microbiol. 2005, 3, 882–892. [Google Scholar] [CrossRef]
- Slade, D.; Radman, M. Oxidative stress resistance in Deinococcus radiodurans. Microbiol. Mol. Biol. Rev. 2011, 75, 133–191. [Google Scholar] [CrossRef] [Green Version]
- Castro-Severyn, J.; Remonsellez, F.; Valenzuela, S.; Salinas, C.; Fortt, J.; Aguilar, P.; Pardo-Esté, C.; Dorador, C.; Quatrini, R.; Molina, F.; et al. Comparative genomics analysis of a new Exiguobacterium strain from Salar de Huasco reveals a repertoire of stress-related genes and arsenic resistance. Front. Microbiol. 2017, 8, 456. [Google Scholar] [CrossRef] [Green Version]
- Rappas, M.; Niwa, H.; Zhang, X. Mechanisms of ATPases: A multidisciplinary approach. Curr. Protein Pept. Sci. 2004, 5, 89–105. [Google Scholar] [CrossRef]
- Solioz, M.; Vulpe, C. CPx-type ATPases: A class of P-type ATPase that pump heavy metal. Trends Biochem. Sci. 1996, 21, 237–241. [Google Scholar] [CrossRef]
- Bissig, K.; Wunderli-Ye, H.; Duda, P.; Solioz, M. Structure-function analysis of purified Enterococcus hirae CopB copper ATPase: Effect of Menkes/Wilson disease mutation homologues. Biochem. J. 2001, 357, 217–223. [Google Scholar] [PubMed] [Green Version]
- Fan, B.; Rosen, B. Biochemical Characterization of CopA, the Escherichia coli Cu(I)- translocating P-type ATPase. J. Biol. Chem. 2002, 277, 46987–46992. [Google Scholar] [CrossRef] [Green Version]
- Pufahl, R.; Singer, C.; Peariso, K.; Lin, S.; Schmidt, P.; Fahrni, C.; Cizewski, V.; Penner-Hahn, J.; O’halloran, T. Metal Ion Chaperone Function of the Soluble Cu (I) Receptor Atx1. Science 1997, 278, 853–856. [Google Scholar] [CrossRef] [PubMed]
- Banci, L.; Bertini, I.; del Conte, R.; Markey, J.; Ruiz-Dueñas, F. Copper Trafficking: The Solution Structure of Bacillus subtilis CopZ. Biochemistry 2001, 40, 15660–15668. [Google Scholar] [CrossRef] [PubMed]
- Navarro, C.; von Bernath, D.; Martínez-Bussenius, C.; Castillo, R.; Jerez, C. Cytoplasmic CopZ-Like Protein and Periplasmic Rusticyanin and AcoP Proteins as Possible Copper Resistance Determinants in Acidithiobacillus ferrooxidans ATCC 23270. Appl. Environ. Microbiol. 2016, 82, 1015–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennison, C. Investigating the structure and function of cupredoxins. Coord. Chem. Rev. 2005, 249, 3025–3054. [Google Scholar] [CrossRef]
- Arnesano, F.; Banci, L.; Bertini, I.; Thompsett, A. Solution structure of CopC: A cupredoxin-like protein involved in copper homeostasis. Structure 2002, 10, 1337–1347. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Tsui, H.; Bruce, K.; Sham, L.; Higgins, A.; Lisher, J.; Kazmierczak, K.; Maroney, M.; Dann, C.; Winkler, M.; et al. A new structural paradigm in copper resistance in Streptococcus pneumoniae. Nat. Chem. Biol. 2013, 9, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Castelle, C.; Ilbert, M.; Infossi, P.; Leroy, G.; Giudici-Orticoni, M. An unconventional copper protein required for cytochrome c oxidase respiratory function under extreme acidic conditions. J. Biol. Chem. 2010, 285, 21519–21525. [Google Scholar] [CrossRef] [Green Version]
- Rensing, C.; Grass, G. Escherichia coli mechanisms of copper homeostasis in a changing environment. FEMS Microbiol. Rev. 2003, 27, 197–213. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Rensing, C.; Mcevoy, M. Chaperone-mediated copper handling in the periplasm. Nat. Prod. Rep. 2010, 27, 711–719. [Google Scholar] [CrossRef]
- Franke, S.; Grass, G.; Rensing, C.; Nies, D. Molecular analysis of the copper transporting efflux system CusCFBA of Escherichia coli. J. Bacteriol. Res. 2003, 185, 3804–3812. [Google Scholar] [CrossRef] [Green Version]
- Munson, G.; Lam, D.; Outten, W.; O’halloran, V. Identification of a Copper-Responsive Two-Component System on the Chromosome of Escherichia coli K-12. J. Bacteriol. Res. 2000, 182, 5864–5871. [Google Scholar] [CrossRef] [Green Version]
- Magnani, D.; Solioz, M. How bacteria handle copper. In Molecular Microbiology of Heavy Metals; Springer: Berlin/Heidelberg, Germany, 2007; Volume 6, pp. 259–285. [Google Scholar]
- Martínez-Bussenius, C.; Navarro, C.; Orellana, L.; Paradela, A.; Jerez, C. Global response of Acidithiobacillus ferrooxidans ATCC 53993 to high concentrations of copper: A quantitative proteomics approach. J. Proteom. 2016, 145, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Roger, M.; Biaso, F.; Castelle, C.; Bauzan, M.; Chaspoul, F.; Lojou, E.; Sciara, G.; Caffarri, S.; Giudici-Orticoni, M.; Ilbert, M. Spectroscopic characterization of a green copper site in a single-domain cupredoxin. PLoS ONE 2014, 9, e98941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felicio, A.; Garcia, O.; Bertolini, M.; Ottoboni, L.; Novo, M. The effects of copper ions on the synthesis of periplasmic and membrane proteins in Acidithiobacillus ferrooxidans as analyzed by SDS-PAGE and 2D-PAGE. Hydrometallurgy 2003, 71, 165–171. [Google Scholar] [CrossRef]
- Zhang, L.; Koay, M.; Maher, M.; Xiao, Z.; Weed, A. Intermolecular Transfer of Copper Ions from the CopC Protein of Pseudomonas syringae. Crystal Structures of Fully Loaded CuI CuII Forms. J. Am. Chem. Soc. 2006, 128, 5834–5850. [Google Scholar] [CrossRef]
- Radford, D.; Kihlken, M.; Borrelly, G.; Harwood, C.; Le Brun, N.; Cavet, J. CopZ from Bacillus subtilis interacts in vivo with a copper exporting Cpx-type ATPase CopA. FEMS Microbiol. Lett. 2003, 220, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Cobine, P.; Wickramasinghe, W.; Harrison, M.; Weber, T.; Solioz, M.; Dameron, C. The Enterococcus hirae copper chaperone CopZ delivers copper(I) to the CopY repressor. FEBS Lett. 1999, 445, 27–30. [Google Scholar] [CrossRef] [Green Version]
- Toes, A.; Daleke, M.; Kuenen, J.; Muyzer, G. Expression of copA and cusA in Shewanella during copper stress. Microbiology 2008, 154, 2709–2718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stott, M.; Watling, H.; Franzmann, P.; Sutton, D. The role of iron-hydroxy precipitates in the passivation of chalcopyrite during bioleaching. Miner. Eng. 2000, 13, 1117–1127. [Google Scholar] [CrossRef]
- Leduc, L.; Trevors, J.; Ferroni, G. Thermal characterization of different isolates of Thiobacillus ferrooxidans. FEMS Microbiol. Lett. 1993, 108, 189–194. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
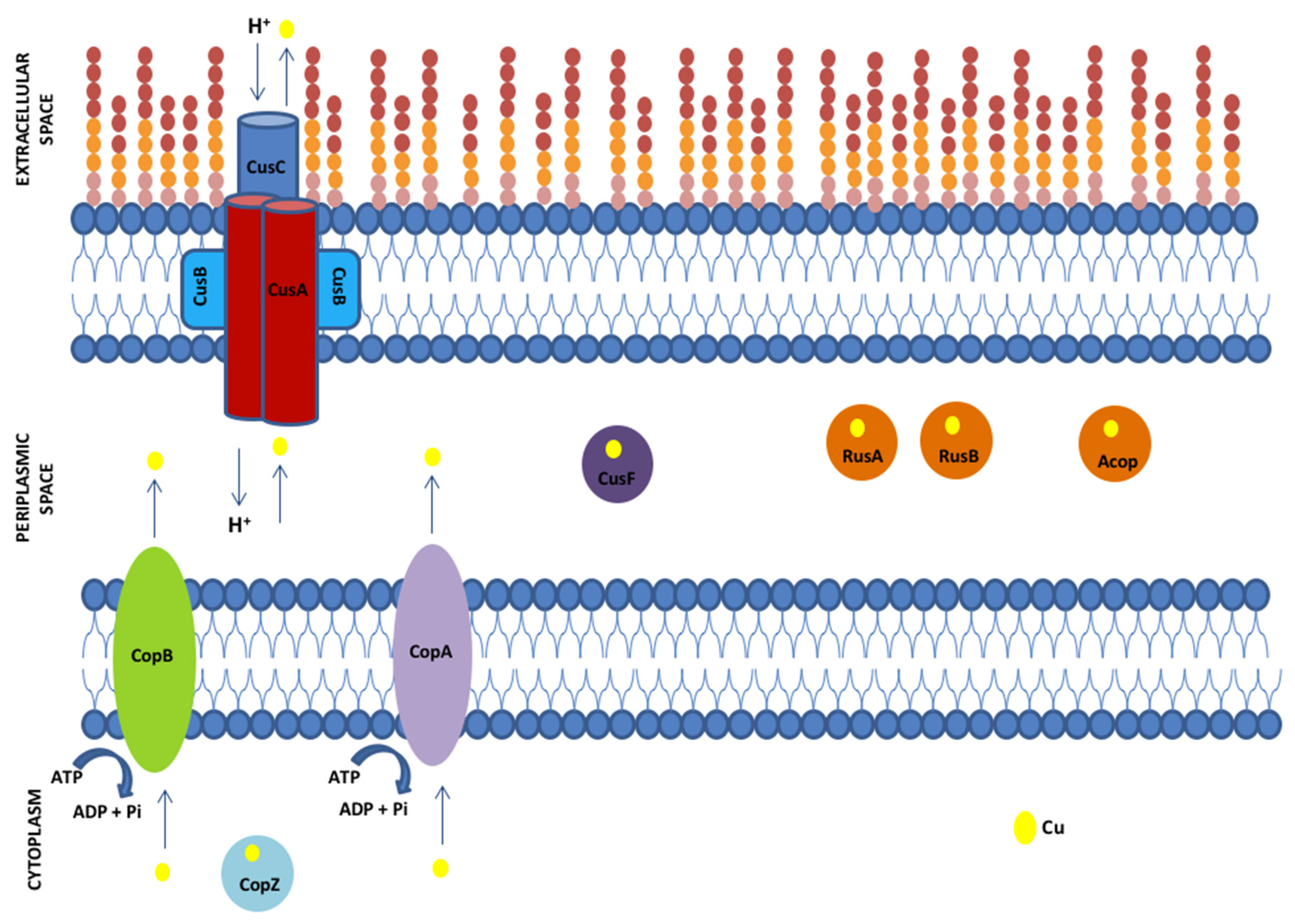
| Protein | Functional Description * |
|---|---|
| CopA | Copper-exporting P-type ATPase A, exports Cu+ from the cytoplasm to the periplasm; binds 2 Cu+ ions per monomer, which are transferred to periplasmic copper chaperone CusF upon ATP hydrolysis. |
| CopB | Copper-exporting P-type ATPase B, copper-translocating outer membrane protein. |
| CopZ | Copper chaperone, ion-binding protein delivering two Cu+ ions to the copper-transporting ATPase CopA. |
| CusA | Cation efflux system protein, copper efflux RND transporter permease. |
| CusB | Cation efflux system protein, copper efflux pump MFP component. |
| CusC | Cation efflux system protein RND transporter outer membrane channel component |
| CusF | Cation efflux system protein, periplasmic copper-binding chaperone component. |
| RusA | Rusticyanin type A, high potential iron sulfur protein, blue copper cupredoxin. |
| RusB | Rusticyanin type B, high potential iron sulfur protein, blue copper cupredoxin. |
| Acop | Acidophile cytochrome c oxidase partner, green copper cupredoxin [5]. |
| Protein | Metal Binding Motif | Phosphatase Domain | 6′ Translocation Motif | Phosphorylation Domain | Conserved GXGXXG/A Motif | TGDN Motif | GDGXNDXP Motif |
|---|---|---|---|---|---|---|---|
| CopA (E.coli) | CASC….CASC | TGEP | CPCALGLA | FDKTGTLT | GLGVSG | TGDN | GDGINAP |
| CopA (E.hirae) | CANC | TGES | CPCALGLA | LDKTGTLT | GAGISG | TGDN | GDGINAP |
| CopB (E.hirae) | No | TGES | CPHALGLA | LDKTGTLT | GVGLEA | TGDN | GDGINDAP |
| CopA1Af | No | TGES | CPHALGLA | FDKTGTLT | GKGAQA | TGDS | GDGVNDAP |
| CopA2Af | No | TGES | CPHALGLA | FDKTGTLT | GKGAQA | TGDS | GDGVNDAP |
| CopBAf | CASC….CASC | TGEP | CPCAMGLA | LDKTGTLT | GKGVRG | TGDL | GEGINDSP |
| CopA1ACH | No | TGES | CPHALGLA | FDKTGTLT | GKGAQA | TGDS | GDGVNDAP |
| CopBACH1 | CASC….CASC | TGEP | CPCAMGLA | FDKTGTLT | GYGVEG | TGDA | GDGINDAP |
| CopBACH2 | CASC….CASC | TGEP | CPCAMGLA | FDKTGTLT | GYGIEG | TGDG | GDGINDAP |
| CopBACH2 | CASC….CASC | TGEP | CPCAMGLA | FDKTGTLT | GYGIEG | TGDG | GDGINDAP |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barahona, S.; Castro-Severyn, J.; Dorador, C.; Saavedra, C.; Remonsellez, F. Determinants of Copper Resistance in Acidithiobacillus Ferrivorans ACH Isolated from the Chilean Altiplano. Genes 2020, 11, 844. https://doi.org/10.3390/genes11080844
Barahona S, Castro-Severyn J, Dorador C, Saavedra C, Remonsellez F. Determinants of Copper Resistance in Acidithiobacillus Ferrivorans ACH Isolated from the Chilean Altiplano. Genes. 2020; 11(8):844. https://doi.org/10.3390/genes11080844
Chicago/Turabian StyleBarahona, Sergio, Juan Castro-Severyn, Cristina Dorador, Claudia Saavedra, and Francisco Remonsellez. 2020. "Determinants of Copper Resistance in Acidithiobacillus Ferrivorans ACH Isolated from the Chilean Altiplano" Genes 11, no. 8: 844. https://doi.org/10.3390/genes11080844
APA StyleBarahona, S., Castro-Severyn, J., Dorador, C., Saavedra, C., & Remonsellez, F. (2020). Determinants of Copper Resistance in Acidithiobacillus Ferrivorans ACH Isolated from the Chilean Altiplano. Genes, 11(8), 844. https://doi.org/10.3390/genes11080844