Transcriptome Profiling and Differential Gene Expression in Canine Microdissected Anagen and Telogen Hair Follicles and Interfollicular Epidermis

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Skin Biopsies
2.3. Microdissection
2.4. Histological Analysis
2.5. RNA Extraction and Transcriptome Sequencing (RNA-seq)
2.6. Mapping to Reference Genome and Gene Expression Analysis
2.7. Differentially Gene Expression
2.8. Pathway and Gene Enrichment Analysis
3. Results
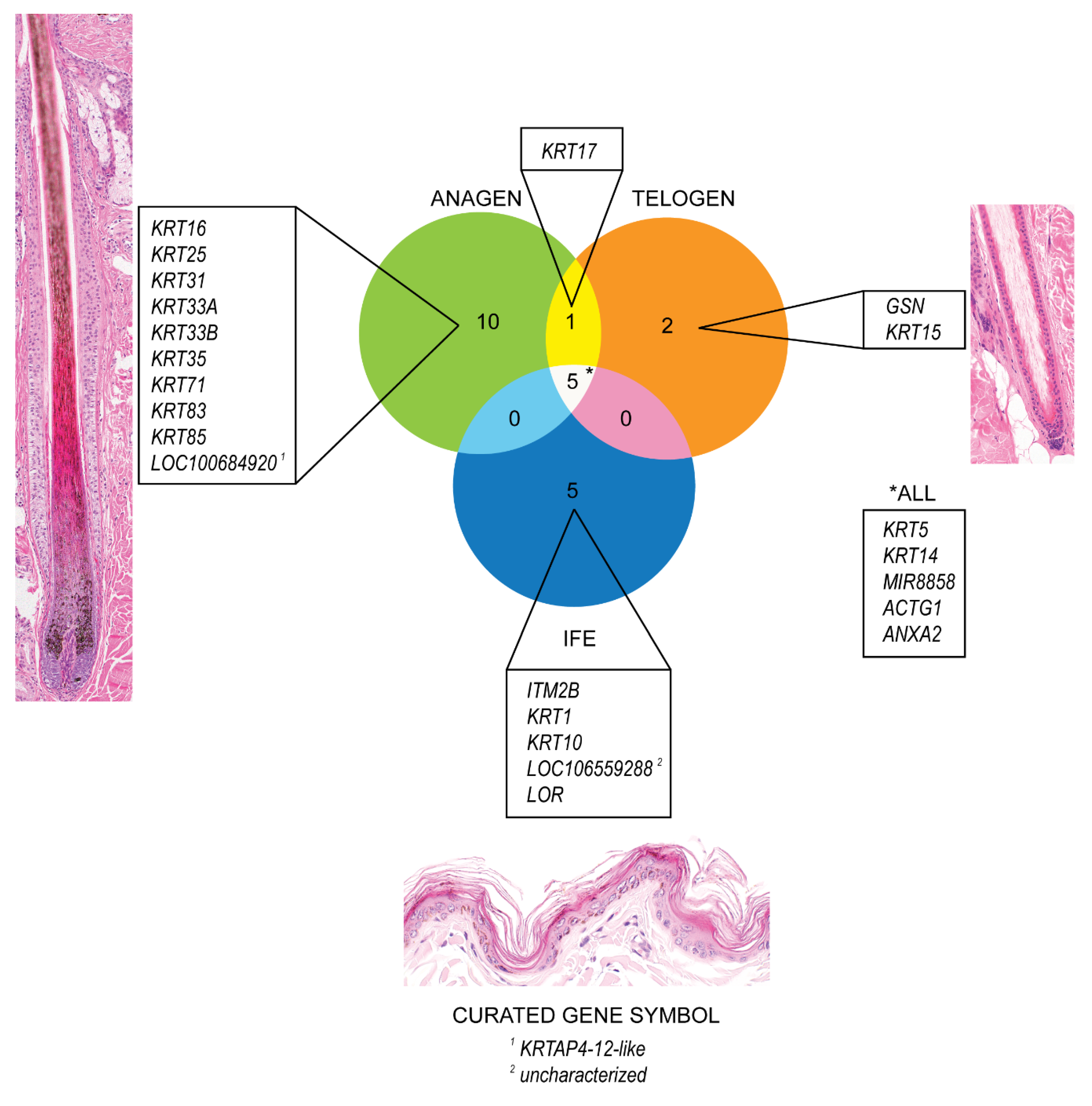
3.1. Microdissection
3.2. Histologic Findings
3.3. Gene Expression Profile in Anagen HF, Telogen HF and IFE
3.4. Highly Expressed Genes
3.5. Gene Expression Profile of Selected Genes
3.6. Expression of Genes Associated with Canine Genodermatoses
3.7. Differentially Expressed Genes in Anagen and Telogen Hair Cycle Stages
3.8. Functional Classification of Differentially Expressed Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fuchs, E. Scratching the surface of skin development. Nature 2007, 445, 834–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suter, M.M.; Schulze, K.; Bergman, W.; Welle, M.; Roosje, P.; Muller, E.J. The keratinocyte in epidermal renewal defence. Vet. Dermatol. 2009, 20, 515–532. [Google Scholar] [CrossRef] [PubMed]
- Halprin, K.M. Epidermal turnover time—A re-examination. Br. J. Dermatol. 1972, 86, 14–19. [Google Scholar] [CrossRef]
- Blanpain, C.; Fuchs, E. Epidermal stem cells of the skin. Annu. Rev. Cell Dev. Biol. 2006, 22, 339–373. [Google Scholar] [CrossRef] [Green Version]
- Watt, F.M. The stem cell compartment in human interfollicular epidermis. J. Dermatol. Sci. 2002, 28, 173–180. [Google Scholar] [CrossRef]
- Watt, F.M.; Fujiwara, H. Cell-Extracellular matrix interactions in normal and diseased skin. Cold Spring Harb. Perspect. Biol. 2011, 3, a005124. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.C.; Pasolli, H.A.; Fuchs, E. Dynamics between stem cells, niche, and progeny in the hair follicle. Cell 2011, 144, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.V.; Cheong, J.; Ciapurin, N.; McDermitt, D.J.; Tumbar, T. Distinct self-renewal and differentiation phases in the niche of infrequently dividing hair follicle stem cells. Cell Stem Cell 2009, 5, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Trempus, C.S.; Morris, R.J.; Bortner, C.D.; Cotsarelis, G.; Faircloth, R.S.; Reece, J.M.; Tennant, R.W. Enrichment for living murine keratinocytes from the hair follicle bulge with the cell surface marker CD34. J. Investig. Dermatol. 2003, 120, 501–511. [Google Scholar]
- Gerhards, N.M.; Sayar, B.S.; Origgi, F.C.; Galichet, A.; Müller, E.J.; Welle, M.M.; Wiener, D.J. Stem Cell-Associated Marker Expression in Canine Hair Follicles. J. Histochem. Cytochem. 2016, 64, 190–204. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Lyle, S.; Yang, Z.; Cotsarelis, G. Keratin 15 promoter targets putative epithelial stem cells in the hair follicle bulge. J. Investig. Dermatol. 2003, 121, 963–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaks, V.; Barker, N.; Kasper, M.; van Es, J.H.; Snippert, H.J.; Clevers, H.; Toftgård, R. Lgr5 marks cycling, yet long-lived, hair follicle stem cells. Nat. Genet. 2008, 40, 1291–1299. [Google Scholar] [CrossRef] [PubMed]
- Nowak, J.A.; Polak, L.; Pasolli, H.A.; Fuchs, E. Hair follicle stem cells are specified and function in early skin morphogenesis. Cell Stem Cell 2008, 3, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Snippert, H.J.; Haegebarth, A.; Kasper, M.; Jaks, V.; van Es, J.H.; Barker, N.; van de Wetering, M.; van den Born, M.; Begthel, H.; Vries, R.G.; et al. Lgr6 marks stem cells in the hair follicle that generate all cell lineages of the skin. Science 2010, 327, 1385–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, B.G.; Lee, C.; Kim, H.S.; Shin, M.S.; Cheon, M.S.; Kim, J.W.; Kim, W.H. Distinct expression profile of stem cell markers, LGR5 and LGR6, in basaloid skin tumors. Virchows Arch. 2017, 470, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Iwasaki, T.; Amagai, M.; Ohyama, M. Canine follicle stem cell candidates reside in the bulge and share characteristic features with human bulge cells. J. Investig. Dermatol. 2010, 130, 1988–1995. [Google Scholar] [CrossRef] [Green Version]
- Brunner, M.A.T.; Jagannathan, V.; Waluk, D.P.; Roosje, P.; Linek, M.; Panakova, L.; Leeb, T.; Wiener, D.J.; Welle, M.M. Novel insights into the pathways regulating the canine hair cycle and their deregulation in alopecia X. PLoS ONE 2017, 12, e0186469. [Google Scholar] [CrossRef] [Green Version]
- de Castro, R.V.G.; Tavares, M.R.; Bressan, F.F.; Pieri, N.C.G.; Hill, A.B.T.; Souza, A.F.; Cruz, N.d.R.N.; Martins, D.S.; Ambrosio, C.E.; Meirelles, F.V.; et al. In vitro identification of a stem cell population from canine hair follicle bulge region. Tissue Cell 2018, 50, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Jahoda, C.A.; Christiano, A.M. Niche crosstalk: Intercellular signals at the hair follicle. Cell 2011, 146, 678–681. [Google Scholar] [CrossRef] [Green Version]
- Plikus, M.V.; Chuong, C.M. Complex hair cycle domain patterns and regenerative hair waves in living rodents. J. Investig. Dermatol. 2008, 128, 1071–1080. [Google Scholar] [CrossRef] [Green Version]
- Festa, E.; Fretz, J.; Berry, R.; Schmidt, B.; Rodeheffer, M.; Horowitz, M.; Horsley, V. Adipocyte lineage cells contribute to the skin stem cell niche to drive hair cycling. Cell 2011, 146, 761–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Tumbar, T. Hairy tale of signaling in hair follicle development and cycling. Semin. Cell Dev. Biol. 2012, 23, 906–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rompolas, P.; Greco, V. Stem cell dynamics in the hair follicle niche. Semin. Cell Dev. Biol. 2014, 25, 34–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumigray, K.D.; Lechler, T. Cell adhesion in epidermal development and barrier formation. Curr. Top. Dev. Biol. 2015, 112, 383–414. [Google Scholar] [PubMed] [Green Version]
- Coulombe, P.A.; Kopan, R.; Fuchs, E. Expression of keratin K14 in the epidermis and hair follicle: Insights into complex programs of differentiation. J. Cell Biol. 1989, 109, 2295–2312. [Google Scholar] [CrossRef]
- Romani, N.; Ebner, S.; Tripp, C.H.; Flacher, V.; Koch, F.; Stoitzner, P. Epidermal Langerhans Cells—Changing views on their function in vivo. Immunol. Lett. 2006, 106, 119–125. [Google Scholar] [CrossRef]
- Klicznik, M.M.; Szenes-Nagy, A.B.; Campbell, D.J.; Gratz, I.K. Taking the lead-how keratinocytes orchestrate skin T cell immunity. Immunol. Lett. 2018, 200, 43–51. [Google Scholar] [CrossRef]
- Adachi, T.; Kobayashi, T.; Sugihara, E.; Yamada, T.; Ikuta, K.; Pittaluga, S.; Saya, H.; Amagai, M.; Nagao, K. Hair follicle-derived IL-7 and IL-15 mediate skin-resident memory T cell homeostasis and lymphoma. Nat. Med. 2015, 21, 1272–1279. [Google Scholar] [CrossRef]
- Nagao, K.; Kobayashi, T.; Moro, K.; Ohyama, M.; Adachi, T.; Kitashima, D.Y.; Ueha, S.; Horiuchi, K.; Tanizaki, H.; Kabashima, K.; et al. Stress-induced production of chemokines by hair follicles regulates the trafficking of dendritic cells in skin. Nat. Immunol. 2012, 13, 744–752. [Google Scholar] [CrossRef] [Green Version]
- Vahlquist, A.; Fischer, J.; Torma, H. Inherited Nonsyndromic Ichthyoses: An Update on Pathophysiology, Diagnosis and Treatment. Am. J. Clin. Dermatol. 2018, 19, 51–66. [Google Scholar] [CrossRef]
- Duverger, O.; Morasso, M.I. To grow or not to grow: Hair morphogenesis and human genetic hair disorders. Semin. Cell Dev. Biol. 2014, 25–26, 22–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.; Almohanna, H.; Griggs, J.; Tosti, A. Genetic Hair Disorders: A Review. Dermatol. Ther. (Heidelb.) 2019, 9, 421–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leeb, T.; Müller, E.J.; Roosje, P.; Welle, M. Genetic testing in veterinary dermatology. Vet. Dermatol. 2017, 28, 4-e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, A.; Nimmo, J.; Newman, R.; Brunner, M.; Welle, M.M.; Jagannathan, V.; Leeb, T. A splice site variant in the SUV39H2 gene in Greyhounds with nasal parakeratosis. Anim. Genet. 2018, 49, 137–140. [Google Scholar] [CrossRef] [Green Version]
- Drogemuller, M.; Jagannathan, V.; Becker, D.; Drogemuller, C.; Schelling, C.; Plassais, J.; Kaerle, C.; de Citres, C.D.; Thomas, A.; Muller, E.J.; et al. A mutation in the FAM83G gene in dogs with hereditary footpad hyperkeratosis (HFH). PLoS Genet. 2014, 10, e1004370. [Google Scholar] [CrossRef]
- Brunner, M.A.T.; Rufenacht, S.; Bauer, A.; Erpel, S.; Buchs, N.; Braga-Lagache, S.; Heller, M.; Leeb, T.; Jagannathan, V.; Wiener, D.J.; et al. Bald thigh syndrome in sighthounds-Revisiting the cause of a well-known disease. PLoS ONE 2019, 14, e0212645. [Google Scholar] [CrossRef]
- Joost, S.; Zeisel, A.; Jacob, T.; Sun, X.; La Manno, G.; Lonnerberg, P.; Linnarsson, S.; Kasper, M. Single-Cell Transcriptomics Reveals that Differentiation and Spatial Signatures Shape Epidermal and Hair Follicle Heterogeneity. Cell Syst. 2016, 3, 221–237.e9. [Google Scholar] [CrossRef] [Green Version]
- Acosta, J.R.; Joost, S.; Karlsson, K.; Ehrlund, A.; Li, X.; Aouadi, M.; Kasper, M.; Arner, P.; Ryden, M.; Laurencikiene, J. Single cell transcriptomics suggest that human adipocyte progenitor cells constitute a homogeneous cell population. Stem Cell Res. Ther. 2017, 8, 250. [Google Scholar] [CrossRef] [Green Version]
- Edqvist, P.H.; Fagerberg, L.; Hallstrom, B.M.; Danielsson, A.; Edlund, K.; Uhlen, M.; Ponten, F. Expression of human skin-specific genes defined by transcriptomics and antibody-based profiling. J. Histochem. Cytochem. 2015, 63, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, R.; Grzenda, A.; Allison, T.F.; Rawnsley, J.; Balin, S.J.; Sabri, S.; Plath, K.; Lowry, W.E. Defining Transcriptional Signatures of Human Hair Follicle Cell States. J. Investig. Dermatol. 2020, 140, 764–773.e4. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, X.; Yan, H.; Zeng, J.; Ma, S.; Niu, Y.; Zhou, G.; Jiang, Y.; Chen, Y. Comparative Transcriptome Analysis of Fetal Skin Reveals Key Genes Related to Hair Follicle Morphogenesis in Cashmere Goats. PLoS ONE 2016, 11, e0151118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, N.; Su, R.; Wang, Z.; Zhang, Y.; Li, J. Exploring differentially expressed genes between anagen and telogen secondary hair follicle stem cells from the Cashmere goat (Capra hircus) by RNA-Seq. PLoS ONE 2020, 15, e0231376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zheng, Y.; Guo, D.; Zhang, X.; Guo, S.; Hui, T.; Yue, C.; Sun, J.; Guo, S.; Bai, Z.; et al. m6A Methylation Analysis of Differentially Expressed Genes in Skin Tissues of Coarse and Fine Type Liaoning Cashmere Goats. Front. Genet. 2019, 10, 1318. [Google Scholar] [CrossRef] [PubMed]
- Su, R.; Fan, Y.; Qiao, X.; Li, X.; Zhang, L.; Li, C.; Li, J. Transcriptomic analysis reveals critical genes for the hair follicle of Inner Mongolia cashmere goat from catagen to telogen. PLoS ONE 2018, 13, e0204404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, J.M.; Creevy, K.E.; Franks, A.; O’Neill, D.G.; Promislow, D.E.L. The companion dog as a model for human aging and mortality. Aging Cell 2018, 17, e12737. [Google Scholar] [CrossRef]
- Martel, B.C.; Lovato, P.; Baumer, W.; Olivry, T. Translational Animal Models of Atopic Dermatitis for Preclinical Studies. Yale J. Biol. Med. 2017, 90, 389–402. [Google Scholar]
- Kobayashi, T.; Shimizu, A.; Nishifuji, K.; Amagai, M.; Iwasaki, T.; Ohyama, M. Canine hair-follicle keratinocytes enriched with bulge cells have the highly proliferative characteristic of stem cells. Vet. Dermatol. 2009, 20, 338–346. [Google Scholar] [CrossRef]
- Wiener, D.J.; Doherr, M.G.; Müller, E.J.; Welle, M.M. Spatial Distribution of Stem Cell-Like Keratinocytes in Dissected Compound Hair Follicles of the Dog. PLoS ONE 2016, 11, e0146937. [Google Scholar] [CrossRef] [Green Version]
- Müntener, T.; Doherr, M.G.; Guscetti, F.; Suter, M.M.; Welle, M.M. The canine hair cycle—A guide for the assessment of morphological and immunohistochemical criteria. Vet. Dermatol. 2011, 22, 383–395. [Google Scholar] [CrossRef] [Green Version]
- European Nucleotide Archive (ENA). Accession Number PRJEB21761. Available online: http://www.ebi.ac.uk/ena/data/view/PRJEB21761 (accessed on 26 June 2020).
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 19 July 2020).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Wickham, H. ggplot2—Elegant Graphics for Data Analysis, 2nd ed.; Springer International Publishing: New York, NY, USA, 2016. [Google Scholar]
- National Center for Biotechnology Information (NCBI). Gene. Available online: https://www.ncbi.nlm.nih.gov/gene (accessed on 19 July 2020).
- da Huang, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Database for Annotation Visualization and Integrated Discovery (DAVID). Available online: https://david.ncifcrf.gov/ (accessed on 19 June 2020).
- Pirooznia, M.; Nagarajan, V.; Deng, Y. GeneVenn—A web application for comparing gene lists using Venn diagrams. Bioinformation 2007, 1, 420–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, A.; De Lucia, M.; Jagannathan, V.; Mezzalira, G.; Casal, M.L.; Welle, M.M.; Leeb, T. A Large Deletion in the NSDHL Gene in Labrador Retrievers with a Congenital Cornification Disorder. G3 (Bethesda) 2017, 7, 3115–3121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leuthard, F.; Lehner, G.; Jagannathan, V.; Leeb, T.; Welle, M. A missense variant in the NSDHL gene in a Chihuahua with a congenital cornification disorder resembling inflammatory linear verrucous epidermal nevi. Anim. Genet. 2019, 50, 768–771. [Google Scholar] [CrossRef]
- Linek, M.; Doelle, M.; Leeb, T.; Bauer, A.; Leuthard, F.; Henkel, J.; Bannasch, D.; Jagannathan, V.; Welle, M.M. ATP2A2 SINE Insertion in an Irish Terrier with Darier Disease and Associated Infundibular Cyst Formation. Genes 2020, 11, 481. [Google Scholar] [CrossRef]
- Baldeschi, C.; Gache, Y.; Rattenholl, A.; Bouillé, P.; Danos, O.; Ortonne, J.P.; Bruckner-Tuderman, L.; Meneguzzi, G. Genetic correction of canine dystrophic epidermolysis bullosa mediated by retroviral vectors. Hum. Mol. Genet. 2003, 12, 1897–1905. [Google Scholar] [CrossRef] [Green Version]
- Niskanen, J.; Dillard, K.; Arumilli, M.; Salmela, E.; Anttila, M.; Lohi, H.; Hytönen, M.K. Nonsense variant in COL7A1 causes recessive dystrophic epidermolysis bullosa in Central Asian Shepherd dogs. PLoS ONE 2017, 12, e0177527. [Google Scholar] [CrossRef]
- Capt, A.; Spirito, F.; Guaguere, E.; Spadafora, A.; Ortonne, J.P.; Meneguzzi, G. Inherited junctional epidermolysis bullosa in the German Pointer: Establishment of a large animal model. J. Investig. Dermatol. 2005, 124, 530–535. [Google Scholar] [CrossRef] [Green Version]
- Olivry, T.; Linder, K.E.; Wang, P.; Bizikova, P.; Bernstein, J.A.; Dunston, S.M.; Paps, J.S.; Casal, M.L. Deficient plakophilin-1 expression due to a mutation in PKP1 causes ectodermal dysplasia-skin fragility syndrome in Chesapeake Bay retriever dogs. PLoS ONE 2012, 7, e32072. [Google Scholar] [CrossRef] [Green Version]
- Mauldin, E.A.; Wang, P.; Olivry, T.; Henthorn, P.S.; Casal, M.L. Epidermolysis bullosa simplex in sibling Eurasier dogs is caused by a PLEC non-sense variant. Vet. Dermatol. 2017, 28, 10–e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jagannathan, V.; Bannoehr, J.; Plattet, P.; Hauswirth, R.; Drogemuller, C.; Drogemuller, M.; Wiener, D.J.; Doherr, M.; Owczarek-Lipska, M.; Galichet, A.; et al. A mutation in the SUV39H2 gene in Labrador Retrievers with hereditary nasal parakeratosis (HNPK) provides insights into the epigenetics of keratinocyte differentiation. PLoS Genet. 2013, 9, e1003848. [Google Scholar] [CrossRef] [PubMed]
- Bannoehr, J.; Balmer, P.; Stoffel, M.H.; Jagannathan, V.; Gaschen, V.; Kühni, K.; Sayar, B.; Drögemüller, M.; Howald, D.; Wiener, D.J.; et al. Abnormal keratinocyte differentiation in the nasal planum of Labrador Retrievers with hereditary nasal parakeratosis (HNPK). PLoS ONE 2020, 15, e0225901. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.; Waluk, D.P.; Galichet, A.; Timm, K.; Jagannathan, V.; Sayar, B.S.; Wiener, D.J.; Dietschi, E.; Muller, E.J.; Roosje, P.; et al. A de novo variant in the ASPRV1 gene in a dog with ichthyosis. PLoS Genet. 2017, 13, e1006651. [Google Scholar] [CrossRef]
- Metzger, J.; Wöhlke, A.; Mischke, R.; Hoffmann, A.; Hewicker-Trautwein, M.; Küch, E.M.; Naim, H.Y.; Distl, O. A Novel SLC27A4 Splice Acceptor Site Mutation in Great Danes with Ichthyosis. PLoS ONE 2015, 10, e0141514. [Google Scholar] [CrossRef]
- Credille, K.M.; Barnhart, K.F.; Minor, J.S.; Dunstan, R.W. Mild recessive epidermolytic hyperkeratosis associated with a novel keratin 10 donor splice-site mutation in a family of Norfolk terrier dogs. Br. J. Dermatol. 2005, 153, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Mauldin, E.A.; Wang, P.; Evans, E.; Cantner, C.A.; Ferracone, J.D.; Credille, K.M.; Casal, M.L. Autosomal Recessive Congenital Ichthyosis in American Bulldogs Is Associated With NIPAL4 (ICHTHYIN) Deficiency. Vet. Pathol. 2015, 52, 654–662. [Google Scholar] [CrossRef] [Green Version]
- Casal, M.L.; Wang, P.; Mauldin, E.A.; Lin, G.; Henthorn, P.S. A Defect in NIPAL4 Is Associated with Autosomal Recessive Congenital Ichthyosis in American Bulldogs. PLoS ONE 2017, 12, e0170708. [Google Scholar] [CrossRef]
- Briand, A.; Cochet-Faivre, N.; Reyes-Gomez, E.; Jaraud-Darnault, A.; Tiret, L.; Chevallier, L. NIPAL4 deletion identified in an American Bully with autosomal recessive congenital ichthyosis and response to topical therapy. Vet. Med. Sci. 2019, 5, 112–117. [Google Scholar] [CrossRef] [Green Version]
- Grall, A.; Guaguère, E.; Planchais, S.; Grond, S.; Bourrat, E.; Hausser, I.; Hitte, C.; Le Gallo, M.; Derbois, C.; Kim, G.J.; et al. PNPLA1 mutations cause autosomal recessive congenital ichthyosis in golden retriever dogs and humans. Nat. Genet. 2012, 44, 140–147. [Google Scholar] [CrossRef]
- Credille, K.M.; Minor, J.S.; Barnhart, K.F.; Lee, E.; Cox, M.L.; Tucker, K.A.; Diegel, K.L.; Venta, P.J.; Hohl, D.; Huber, M.; et al. Transglutaminase 1-deficient recessive lamellar ichthyosis associated with a LINE-1 insertion in Jack Russell terrier dogs. Br. J. Dermatol. 2009, 161, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Drogemuller, C.; Karlsson, E.K.; Hytonen, M.K.; Perloski, M.; Dolf, G.; Sainio, K.; Lohi, H.; Lindblad-Toh, K.; Leeb, T. A mutation in hairless dogs implicates FOXI3 in ectodermal development. Science 2008, 321, 1462. [Google Scholar] [CrossRef]
- Casal, M.L.; Scheidt, J.L.; Rhodes, J.L.; Henthorn, P.S.; Werner, P. Mutation identification in a canine model of X-linked ectodermal dysplasia. Mamm. Genome 2005, 16, 524–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waluk, D.P.; Zur, G.; Kaufmann, R.; Welle, M.M.; Jagannathan, V.; Drogemuller, C.; Muller, E.J.; Leeb, T.; Galichet, A. A Splice Defect in the EDA Gene in Dogs with an X-Linked Hypohidrotic Ectodermal Dysplasia (XLHED) Phenotype. G3 2016, 6, 2949–2954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasouliha, S.H.; Bauer, A.; Dettwiler, M.; Welle, M.M.; Leeb, T. A frameshift variant in the EDA gene in Dachshunds with X-linked hypohidrotic ectodermal dysplasia. Anim. Genet. 2018, 49, 651–654. [Google Scholar] [CrossRef]
- Parker, H.G.; Whitaker, D.T.; Harris, A.C.; Ostrander, E.A. Whole Genome Analysis of a Single Scottish Deerhound Dog Family Provides Independent Corroboration That a SGK3 Coding Variant Leads to Hairlessness. G3 (Bethesda) 2020, 10, 293–297. [Google Scholar] [CrossRef] [Green Version]
- Hytönen, M.K.; Lohi, H. A frameshift insertion in SGK3 leads to recessive hairlessness in Scottish Deerhounds: A candidate gene for human alopecia conditions. Hum. Genet. 2019, 138, 535–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, H.G.; Chase, K.; Cadieu, E.; Lark, K.G.; Ostrander, E.A. An insertion in the RSPO2 gene correlates with improper coat in the Portuguese water dog. J. Hered. 2010, 101, 612–617. [Google Scholar] [CrossRef] [Green Version]
- Gerber, P.A.; Buhren, B.A.; Schrumpf, H.; Homey, B.; Zlotnik, A.; Hevezi, P. The top skin-associated genes: A comparative analysis of human and mouse skin transcriptomes. Biol. Chem. 2014, 395, 577–591. [Google Scholar] [CrossRef]
- Hawkshaw, N.J.; Hardman, J.A.; Alam, M.; Jimenez, F.; Paus, R. Deciphering the molecular morphology of the human hair cycle: Wnt signalling during the telogen-anagen transformation. Br. J. Dermatol. 2019, 182, 1184–1193. [Google Scholar] [CrossRef]
- Reddy, S.; Andl, T.; Bagasra, A.; Lu, M.M.; Epstein, D.J.; Morrisey, E.E.; Millar, S.E. Characterization of Wnt gene expression in developing and postnatal hair follicles and identification of Wnt5a as a target of Sonic hedgehog in hair follicle morphogenesis. Mech. Dev. 2001, 107, 69–82. [Google Scholar] [CrossRef]
- Panteleyev, A.A.; Paus, R.; Wanner, R.; Nurnberg, W.; Eichmuller, S.; Thiel, R.; Zhang, J.; Henz, B.M.; Rosenbach, T. Keratin 17 gene expression during the murine hair cycle. J. Investig. Dermatol. 1997, 108, 324–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plikus, M.V.; Mayer, J.A.; de la Cruz, D.; Baker, R.E.; Maini, P.K.; Maxson, R.; Chuong, C.M. Cyclic dermal BMP signalling regulates stem cell activation during hair regeneration. Nature 2008, 451, 340–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, R.; Zhang, W.; Sharma, R.; Chang, Z.; Yin, J.; Li, J. Characterization of BMP2 gene expression in embryonic and adult Inner Mongolia Cashmere goat (Capra hircus) hair follicles. Can. J. Anim. Sci. 2009, 89, 457–462. [Google Scholar] [CrossRef]
- McGowan, K.M.; Tong, X.; Colucci-Guyon, E.; Langa, F.; Babinet, C.; Coulombe, P.A. Keratin 17 null mice exhibit age- and strain-dependent alopecia. Genes Dev. 2002, 16, 1412–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Zhang, S.; Wang, G. Keratin 17 in disease pathogenesis: From cancer to dermatoses. J. Pathol. 2019, 247, 158–165. [Google Scholar] [CrossRef]
- Wang, Z.; Wong, P.; Langbein, L.; Schweizer, J.; Coulombe, P.A. Type II epithelial keratin 6hf (K6hf) is expressed in the companion layer, matrix, and medulla in anagen-stage hair follicles. J. Investig. Dermatol. 2003, 121, 1276–1282. [Google Scholar] [CrossRef] [Green Version]
- Lim, X.; Nusse, R. Wnt signaling in skin development, homeostasis, and disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008029. [Google Scholar] [CrossRef] [Green Version]
- Du, G.; Kataoka, K.; Sakaguchi, M.; Abarzua, F.; Than, S.S.; Sonegawa, H.; Makino, T.; Shimizu, T.; Huh, N.H. Expression of REIC/Dkk-3 in normal and hyperproliferative epidermis. Exp. Dermatol. 2011, 20, 273–277. [Google Scholar] [CrossRef]
- Prowse, D.M.; Lee, D.; Weiner, L.; Jiang, N.; Magro, C.M.; Baden, H.P.; Brissette, J.L. Ectopic expression of the nude gene induces hyperproliferation and defects in differentiation: Implications for the self-renewal of cutaneous epithelia. Dev. Biol. 1999, 212, 54–67. [Google Scholar] [CrossRef] [Green Version]
- Jave-Suarez, L.F.; Winter, H.; Langbein, L.; Rogers, M.A.; Schweizer, J. HOXC13 is involved in the regulation of human hair keratin gene expression. J. Biol. Chem. 2002, 277, 3718–3726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mecklenburg, L.; Tychsen, B.; Paus, R. Learning from nudity: Lessons from the nude phenotype. Exp. Dermatol. 2005, 14, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Meier, N.; Dear, T.N.; Boehm, T. Whn mHa3 are components of the genetic hierarchy controlling hair follicle differentiation. Mech. Dev. 1999, 89, 215–221. [Google Scholar] [CrossRef]
- Kawano, M.; Komi-Kuramochi, A.; Asada, M.; Suzuki, M.; Oki, J.; Jiang, J.; Imamura, T. Comprehensive analysis of FGF and FGFR expression in skin: FGF18 is highly expressed in hair follicles and capable of inducing anagen from telogen stage hair follicles. J. Investig. Dermatol. 2005, 124, 877–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plikus, M.V.; Baker, R.E.; Chen, C.C.; Fare, C.; de la Cruz, D.; Andl, T.; Maini, P.K.; Millar, S.E.; Widelitz, R.; Chuong, C.M. Self-organizing and stochastic behaviors during the regeneration of hair stem cells. Science 2011, 332, 586–589. [Google Scholar] [CrossRef] [Green Version]
- Hou, C.; Miao, Y.; Wang, X.; Chen, C.; Lin, B.; Hu, Z. Expression of matrix metalloproteinases and tissue inhibitor of matrix metalloproteinases in the hair cycle. Exp. Ther. Med. 2016, 12, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Paus, R.; Nickoloff, B.J.; Ito, T. A hairy privilege. Trends Immunol. 2005, 26, 32–40. [Google Scholar] [CrossRef]
- Paus, R.; Bertolini, M. The role of hair follicle immune privilege collapse in alopecia areata: Status and perspectives. J. Investig. Dermatol. Symp. Proc. 2013, 16, S25–S27. [Google Scholar] [CrossRef]
- Paus, R.; Slominski, A.; Czarnetzki, B.M. Is alopecia areata an autoimmune-response against melanogenesis-related proteins, exposed by abnormal MHC class I expression in the anagen hair bulb? Yale J. Biol. Med. 1993, 66, 541–554. [Google Scholar]
- Hoffmann, A.; Metzger, J.; Wöhlke, A.; Peters, M.; Junginger, J.; Mischke, R.; Distl, O.; Hewicker-Trautwein, M. Congenital Ichthyosis in 14 Great Dane Puppies With a New Presentation. Vet. Pathol. 2016, 53, 614–620. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Hair Cycle-Associated Genes | ||||
| Gene Symbol | Full Name of Gene | Anagen | Telogen | IFE |
| ADCY7 | Activin A receptor type 1 | 18.64 | 3.77 | 5.48 |
| BMP2 | Bone morphogenetic protein 2 | 44.37 | 281.79 | 66.23 |
| BMP4 | Bone morphogenetic protein 4 | 99.16 | 7.29 | 12.64 |
| BMP7 | Bone morphogenetic protein 7 | 25.65 | 7.97 | 22.11 |
| BNC1 | Basonuclin 1 | 70.47 | 59.68 | 83.80 |
| BNC2 | Basonuclin 2 | 48.34 | 40.38 | 4.74 |
| CCNA2 | Cyclin A2 | 29.04 | 6.11 | 9.87 |
| CYP1A1 | Cytochrome P450 family 1 subfamily A member 1 | 17.39 | 43.18 | 74.82 |
| CYP1B1 | Cytochrome P450 family 1 subfamily B member 1 | 96.94 | 214.59 | 129.88 |
| CYP24A1 | Cytochrome P450 family 24 subfamily A member 1 | 18.89 | 0.12 | 0.00 |
| CYP27B1 | Cytochrome P450 family 27 subfamily B member 1 | 57.64 | 1.45 | 0.96 |
| DKK3 | Dickkopf 3 | 102.28 | 129.90 | 218.67 |
| FGF18 | Fibroblast growth factor 18 | 8.70 | 133.54 | 4.28 |
| FGF5 | Fibroblast growth factor 5 | 53.66 | 4.65 | 1.78 |
| FOXN1 | Forkhead box protein N1 | 182.11 | 5.58 | 24.39 |
| FZD10 | Frizzled 10 | 15.66 | 3.34 | 3.48 |
| FZD3 | Frizzled 3 | 16.46 | 9.32 | 2.64 |
| HSD17B14 | Hydroxysteroid 17-beta dehydrogenase 14 | 396.58 | 17.20 | 25.22 |
| JAG1 | Jagged 1 | 30.05 | 22.36 | 13.30 |
| KISS1 | KISS-1 metastasis-suppressor | 60.79 | 0.17 | 0.00 |
| MMP7 | Matrix metalloproteinase 7 | 5.44 | 218.73 | 12.82 |
| MSX2 | Homeobox protein MSX2 | 771.09 | 49.38 | 64.14 |
| SFRP1 | Secreted frizzled related protein 1 | 107.40 | 37.31 | 18.18 |
| SFRP2 | Secreted frizzled related protein 2 | 39.53 | 14.07 | 0.26 |
| SOX10 | Sex determining region Y-box 10 | 54.93 | 59.27 | 12.94 |
| TCF7/TCF1 | Transcription factor 1 | 55.84 | 46.93 | 4.60 |
| TGFB2 | Transforming growth factor beta 2 | 2.53 | 11.32 | 5.62 |
| THBS2 | Thrombospondin 2 | 67.86 | 137.49 | 62.01 |
| VDR | Vitamin D receptor | 105.58 | 55.58 | 22.71 |
| WNT3 | Wnt family member 3 | 22.82 | 1.93 | 97.02 |
| WNT5a | Wnt family member 5A | 36.11 | 5.15 | 12.76 |
| WNT10A | Wnt family member 10A | 27.32 | 18.12 | 20.25 |
| Hair Follicle Structure-Associated Genes | ||||
| Gene Symbol | Full Name of Gene | Anagen | Telogen | IFE |
| CD133/PROM1 | Prominin | 15.86 | 3.44 | 0.86 |
| CD71/TFRC | Transferrin receptor | 61.61 | 31.88 | 23.85 |
| CDH3 | P-Cadherin 3 | 196.09 | 225.99 | 463.31 |
| GATA6 | GATA binding protein 6 | 14.80 | 59.76 | 4.07 |
| HOXC13 | Homeobox C3 | 374.22 | 41.39 | 13.79 |
| KRT16 | Keratin 16 | 5931.82 | 10,187.55 | 1790.56 |
| KRT17 | Keratin 17 | 15,476.37 | 77,099.96 | 8158.26 |
| KRT25 | Keratin 25 | 10,456.27 | 33.58 | 2.57 |
| KRT31 | Keratin 31 | 5422.80 | 2.05 | 1.89 |
| KRT33A | Keratin 33A | 10,416.91 | 2.24 | 7.39 |
| KRT33B | Keratin 33B | 10,528.67 | 1.77 | 2.25 |
| KRT34 | Keratin 34 | 3966.39 | 0.55 | 0.81 |
| KRT35 | Keratin 35 | 5365.72 | 1.11 | 0.36 |
| KRT6A | Keratin 6A | 1018.09 | 1513.19 | 37.94 |
| KRT6B | Keratin 6B | 3692.26 | 8621.62 | 80.91 |
| KRT71 | Keratin 71 | 17,621.04 | 11.18 | 7.17 |
| KRT75 | Keratin 5 | 2585.70 | 2895.72 | 2.59 |
| KRT79 | Keratin 79 | 279.52 | 3752.50 | 1216.86 |
| KRT83 | Keratin 83 | 5671.73 | 2.30 | 2.07 |
| KRT85 | Keratin 85 | 10,133.20 | 2.73 | 2.81 |
| KRT86 | Keratin 86 | 3068.60 | 1.20 | 1.06 |
| KRTAP16-1 | Keratin associated protein 16-1 | 2984.34 | 0.33 | 0.72 |
| LEF1 | Lymphoid enhancer binding factor 1 | 87.27 | 18.84 | 21.65 |
| PMEL | Premelanosome protein | 1907.90 | 21.59 | 135.56 |
| TCHH | Trichohyalin | 3997.42 | 5.24 | 8.35 |
| Stem Cell-Associated Genes | ||||
| Gene Symbol | Full Name of Gene | Anagen | Telogen | IFE |
| CD200 | CD200 Molecule | 23.69 | 33.44 | 6.71 |
| CD34 | CD34 Molecule | 111.79 | 131.55 | 47.56 |
| ITGA6 | Integrin subunit alpha 6 | 70.85 | 189.30 | 150.07 |
| ITGB1 | Integrin subunit beta 1 | 219.44 | 341.60 | 265.09 |
| KRT15 | Keratin 15 | 2485.12 | 17,673.04 | 4104.93 |
| KRT19 | Keratin 19 | 59.14 | 180.62 | 4.04 |
| LGR5 | Leucin rich repeat containing G protein-coupled receptor 5 | 9.43 | 10.24 | 0.02 |
| LHX2 | LIM homeobox 2 | 105.49 | 200.13 | 0.34 |
| NFATC1 | Nuclear factor of activated T cells | 19.16 | 76.54 | 21.90 |
| SOX9 | SRY-box transcription factor 9 | 563.03 | 817.93 | 104.80 |
| TCF3 | Transcription factor 3 | 127.36 | 105.95 | 82.50 |
| TCF4 | Transcription factor 4 | 42.90 | 104.53 | 40.37 |
| Epidermal Structure-Associated Genes | ||||
| Gene Symbol | Full Name of Gene | Anagen | Telogen | IFE |
| DSC1 | Desmocollin 1 | 328.67 | 71.79 | 1523.99 |
| DSG1 | Desmoglein 1 | 505.83 | 479.42 | 984.35 |
| DSP | Desmoplakin | 1406.58 | 1492.09 | 1610.92 |
| FLG | Filaggrin | 12.35 | 71.70 | 60.74 |
| IVL | Involucrin | 2244.92 | 480.55 | 85.91 |
| KRT1 | Keratin 1 | 203.64 | 988.44 | 21,712.07 |
| KRT5 | Keratin 5 | 13,982.27 | 18,257.94 | 12,733.06 |
| KRT10 | Keratin 10 | 339.13 | 2042.75 | 33,847.68 |
| KRT14 | Keratin 14 | 23,040.65 | 19,320.49 | 29,472.32 |
| LOR | Loricrin | 65.36 | 399.38 | 7229.10 |
| PLEC | Plectin | 55.78 | 87.56 | 95.43 |
| TP63 | Tumor protein p63 | 137.68 | 179.75 | 230.67 |
| Immune Function-Associated Genes | ||||
| Gene Symbol | Full Name of Gene | Anagen | Telogen | IFE |
| CCL20 | C-C motif chemokine ligand 20 | 0.78 | 3.94 | 14.63 |
| CD74 | CD74 molecule | 113.38 | 737.23 | 1498.77 |
| DLA88 | MHC class I DLA-88 | 116.10 | 439.94 | 584.87 |
| DLA-DQA1 | MHC class II DQ alpha 1 | 20.24 | 115.09 | 257.38 |
| HLA-DRB1 | MHC class II DR beta 1 | 34.90 | 211.64 | 455.74 |
| IL17B | Interleukin 17B | 15.01 | 11.49 | 4.35 |
| IL18 | Interleukin 18 | 18.55 | 62.51 | 94.44 |
| IL1A | Interleukin 1 alpha | 19.31 | 6.59 | 13.99 |
| IL33 | Interleukin 33 | 10.40 | 28.91 | 17.75 |
| TLR2 | Toll like receptor 2 | 1.79 | 15.28 | 2.43 |
| TLR4 | Toll like receptor 4 | 7.55 | 17.14 | 12.41 |
| Tissue Affected | Disease | Dog Breed | Gene Mutation | Anagen | Telogen | IFE | Ref. |
|---|---|---|---|---|---|---|---|
| IFE | CHILD’s like syndrome | Labrador retriever, Chihuahua | NSDHL | 41.2 | 36.5 | 21.2 | [60,61] |
| Darier disease | Irish Terrier | ATP2A2 | 14.3 | 19.3 | 18.9 | [62] | |
| Epidermolysis bullosa, dystrophic | Golden retriever | COL7A1 | 9.6 | 58.6 | 61.4 | [63,64] | |
| Epidermolysis bullosa simplex, junctional | German shorthaired pointer | LAMA3 | 20.6 | 58.6 | 41.4 | [65] | |
| Epidermolysis bullosa simplex, suprabasal | Chesapeake Bay retriever | PKP1 | 0.1 | 0.2 | 0.3 | [66] | |
| Epidermolysis bullosa simplex, basal | Eurasier | PLEC | 55.8 | 87.6 | 95.4 | [67] | |
| Hereditary nasal parakeratosis | Labrador retriever, greyhound | SUV39H2 | 9.8 | 13.9 | 12.5 | [34,68,69] | |
| Hereditary footpad hyperkeratosis | Irish terrier, Kromfohrländer | FAM83G | 174.2 | 190.6 | 158.6 | [35] | |
| Ichthyosis | German shepherd dog | ASPRV1 | 252.9 | 111.1 | 698.8 | [70] | |
| Great Dane | SLC27A4 (FATP4) | 178.9 | 125.9 | 147.6 | [71] | ||
| Norfolk terrier | KRT10 | 339.1 | 2042.8 | 33,847.7 | [72] | ||
| American bulldog | NIPAL4 | 60.9 | 68.2 | 250.1 | [73,74,75] | ||
| Golden retriever | PNPLA1 | 58.0 | 46.8 | 90.3 | [76] | ||
| Jack Russell terrier | TGM1 | 83.1 | 34.2 | 138.0 | [77] | ||
| HF | Canine ectodermal dysplasia | Chinese crested, Mexican hairless dogs, Peruvian hairless dog | FOXI3 | 0.4 | 7.9 | 0.1 | [78] |
| German shepherd dog, dachshund | EDA | 2.4 | 7.8 | 4.8 | [79,80,81] | ||
| Chesapeake Bay retriever | PKP1 | 0.1 | 0.2 | 0.3 | [66] | ||
| Hairlessness | American terrier, Scottish deerhound | SGK3 | 74.1 | 38.1 | 47.9 | [82,83] | |
| Improper coat | Portuguese water dog | RSPO | 1.2 | 0.6 | 0.0 | [84] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiener, D.J.; Groch, K.R.; Brunner, M.A.T.; Leeb, T.; Jagannathan, V.; Welle, M.M. Transcriptome Profiling and Differential Gene Expression in Canine Microdissected Anagen and Telogen Hair Follicles and Interfollicular Epidermis. Genes 2020, 11, 884. https://doi.org/10.3390/genes11080884
Wiener DJ, Groch KR, Brunner MAT, Leeb T, Jagannathan V, Welle MM. Transcriptome Profiling and Differential Gene Expression in Canine Microdissected Anagen and Telogen Hair Follicles and Interfollicular Epidermis. Genes. 2020; 11(8):884. https://doi.org/10.3390/genes11080884
Chicago/Turabian StyleWiener, Dominique J., Kátia R. Groch, Magdalena A.T. Brunner, Tosso Leeb, Vidhya Jagannathan, and Monika M. Welle. 2020. "Transcriptome Profiling and Differential Gene Expression in Canine Microdissected Anagen and Telogen Hair Follicles and Interfollicular Epidermis" Genes 11, no. 8: 884. https://doi.org/10.3390/genes11080884
APA StyleWiener, D. J., Groch, K. R., Brunner, M. A. T., Leeb, T., Jagannathan, V., & Welle, M. M. (2020). Transcriptome Profiling and Differential Gene Expression in Canine Microdissected Anagen and Telogen Hair Follicles and Interfollicular Epidermis. Genes, 11(8), 884. https://doi.org/10.3390/genes11080884