Metabolomic Analysis of SCD during Goose Follicular Development: Implications for Lipid Metabolism

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Goose Follicle Collection and Primary GC Culture
2.3. RNA Extraction and qRT-PCR
2.4. Modulation of SCD with Small Interfering RNA
2.5. Overexpression of SCD with Recombinant Vector
2.6. Determination of SCD Activity
2.7. Cell Sample Preparation for Extraction
2.8. LC-Electrospray Ionization (ESI)-QTRAP-MS/MS) Analysis
2.9. Qualitative and Quantitative Analysis of Metabolites
2.10. Data Processing and Analysis
3. Results
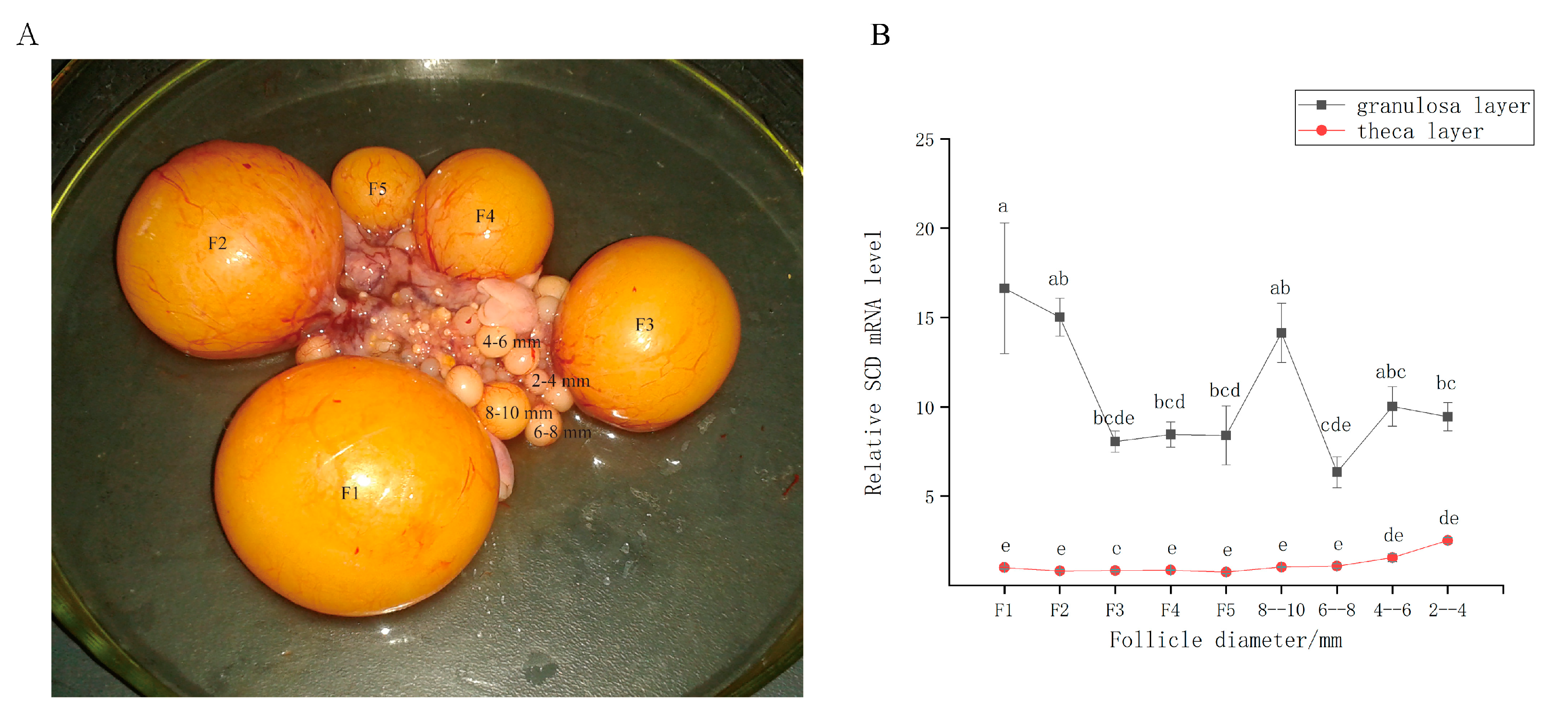
3.1. SCD is Expressed during Follicular Development In Vivo
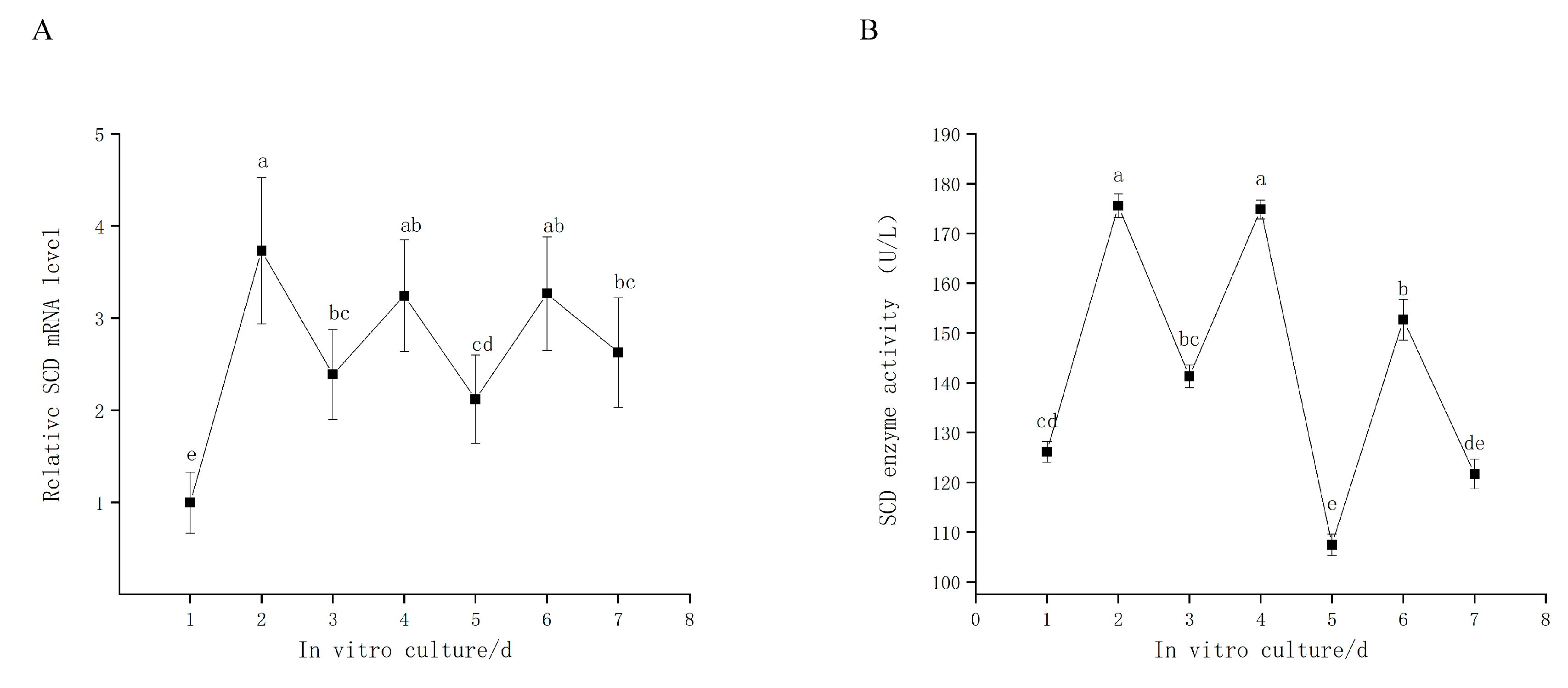
3.2. SCD is Expressed during GC In Vitro Culture
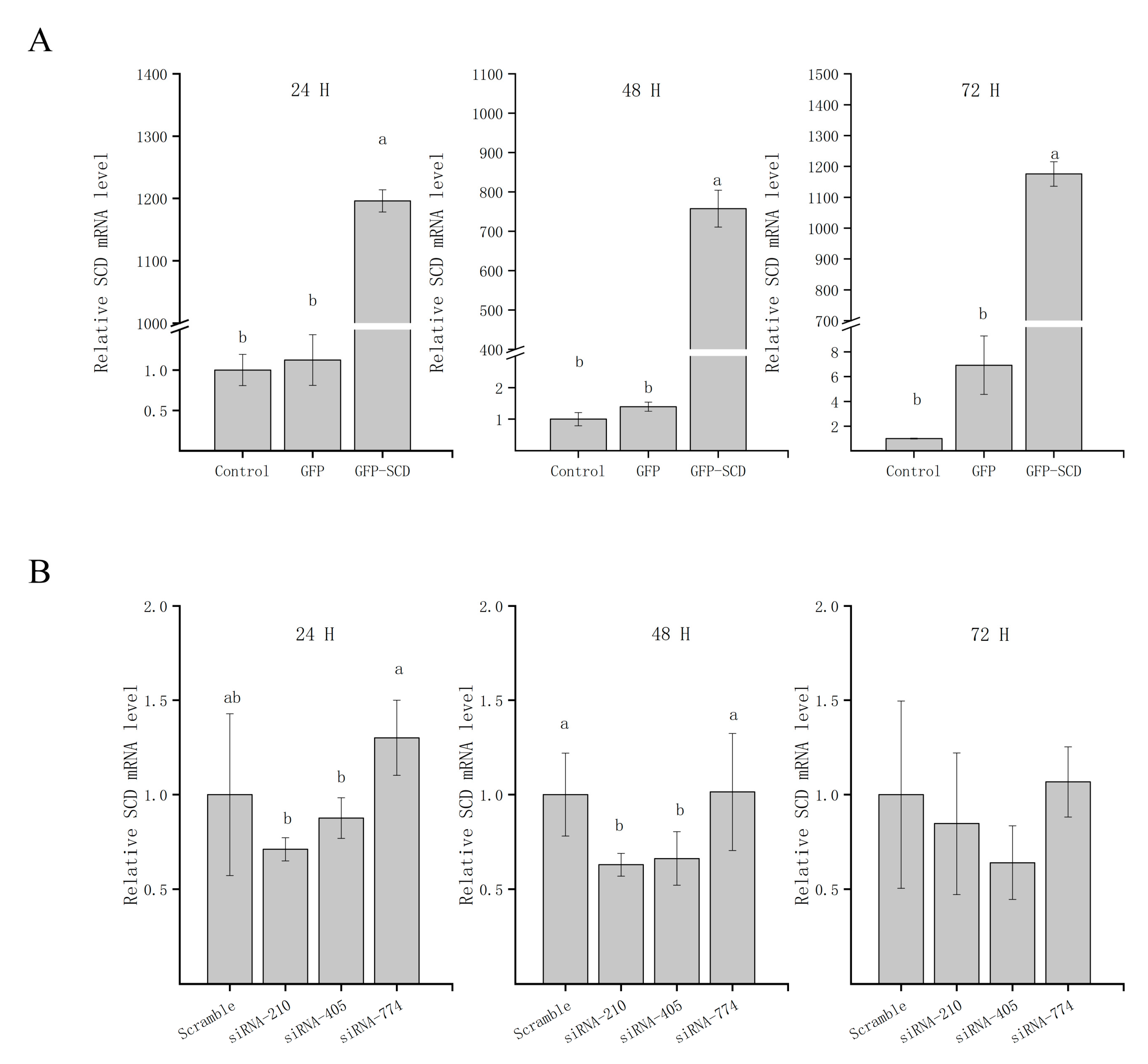
3.3. A GC Cellular Model of SCD Function
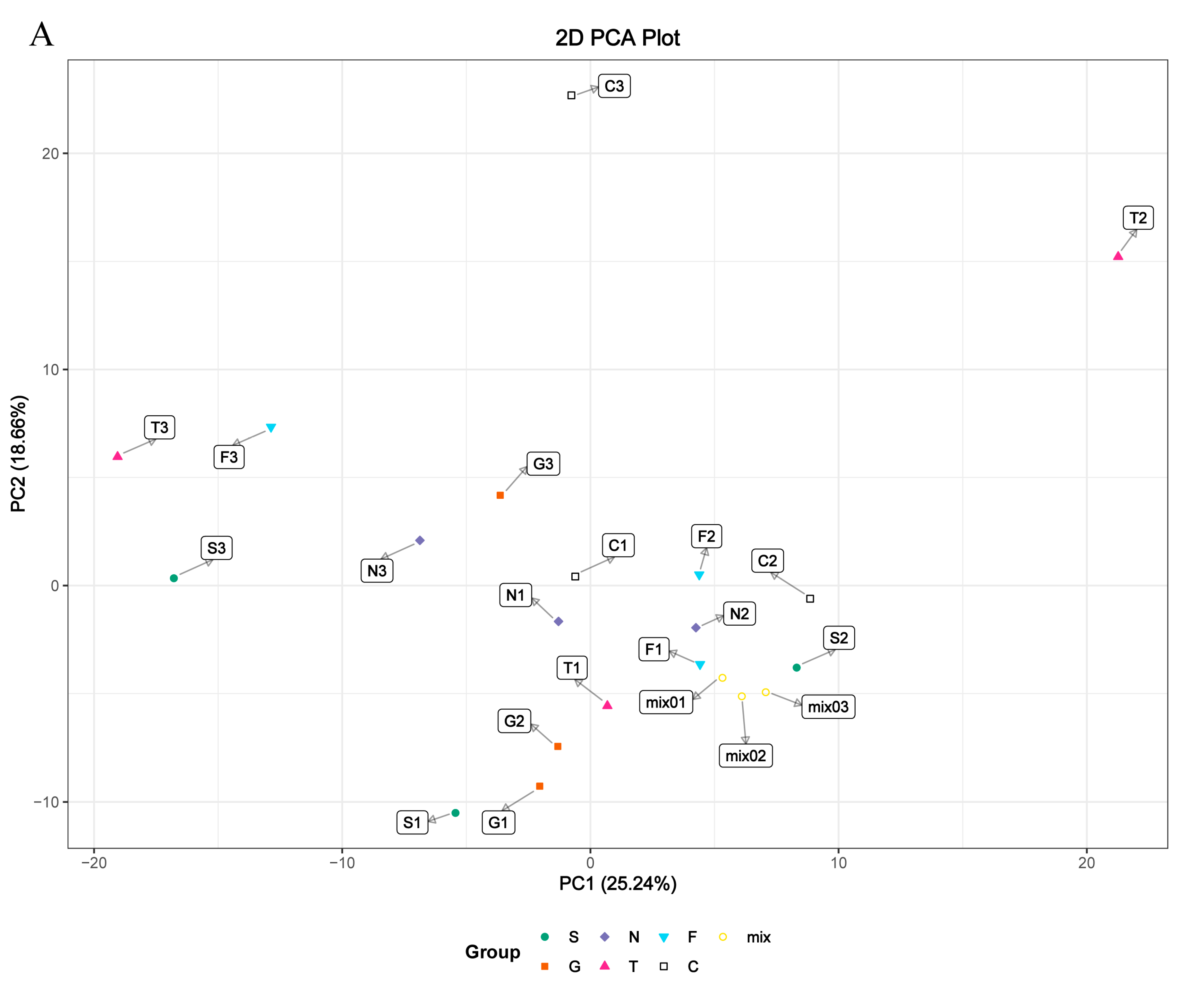
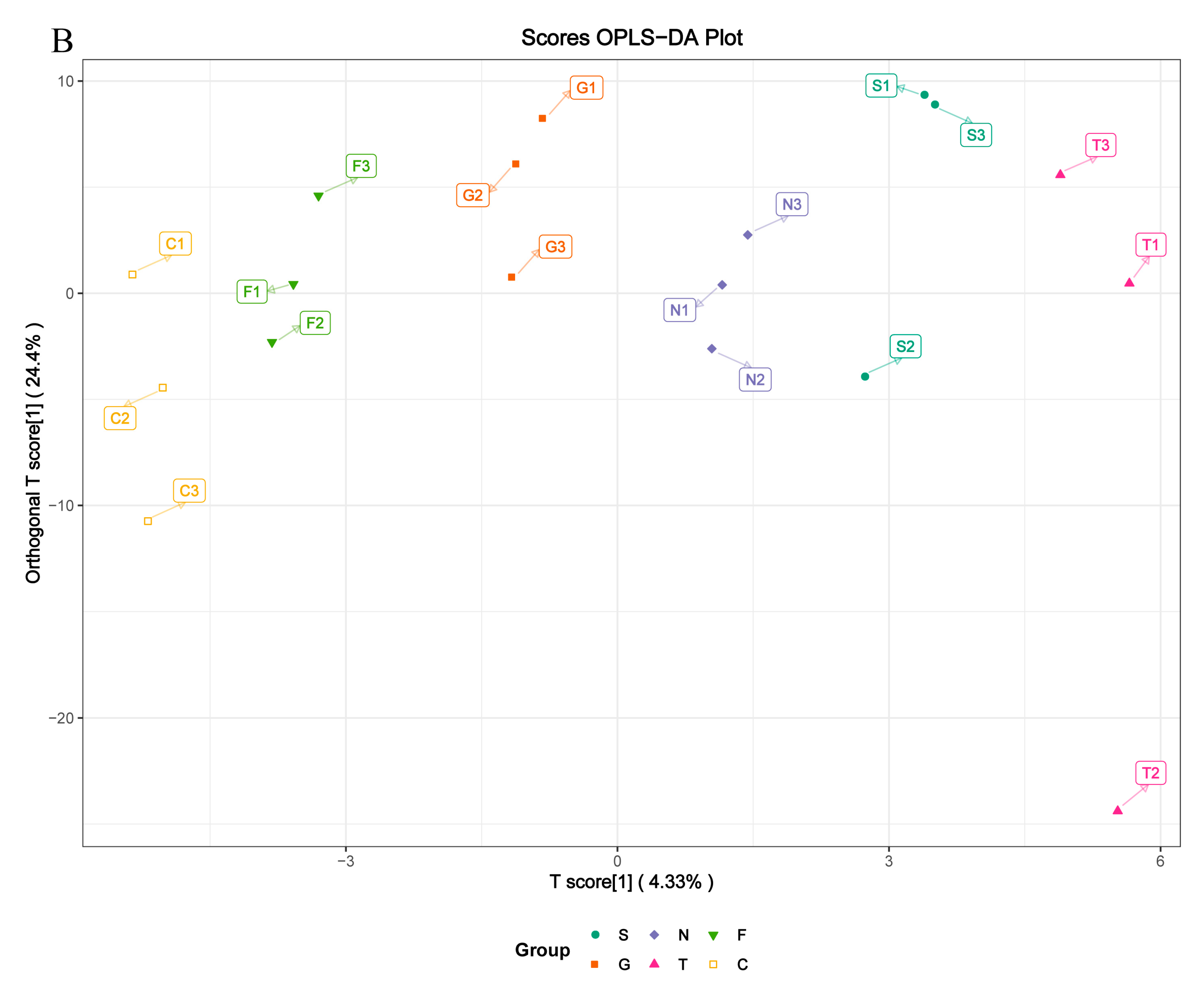
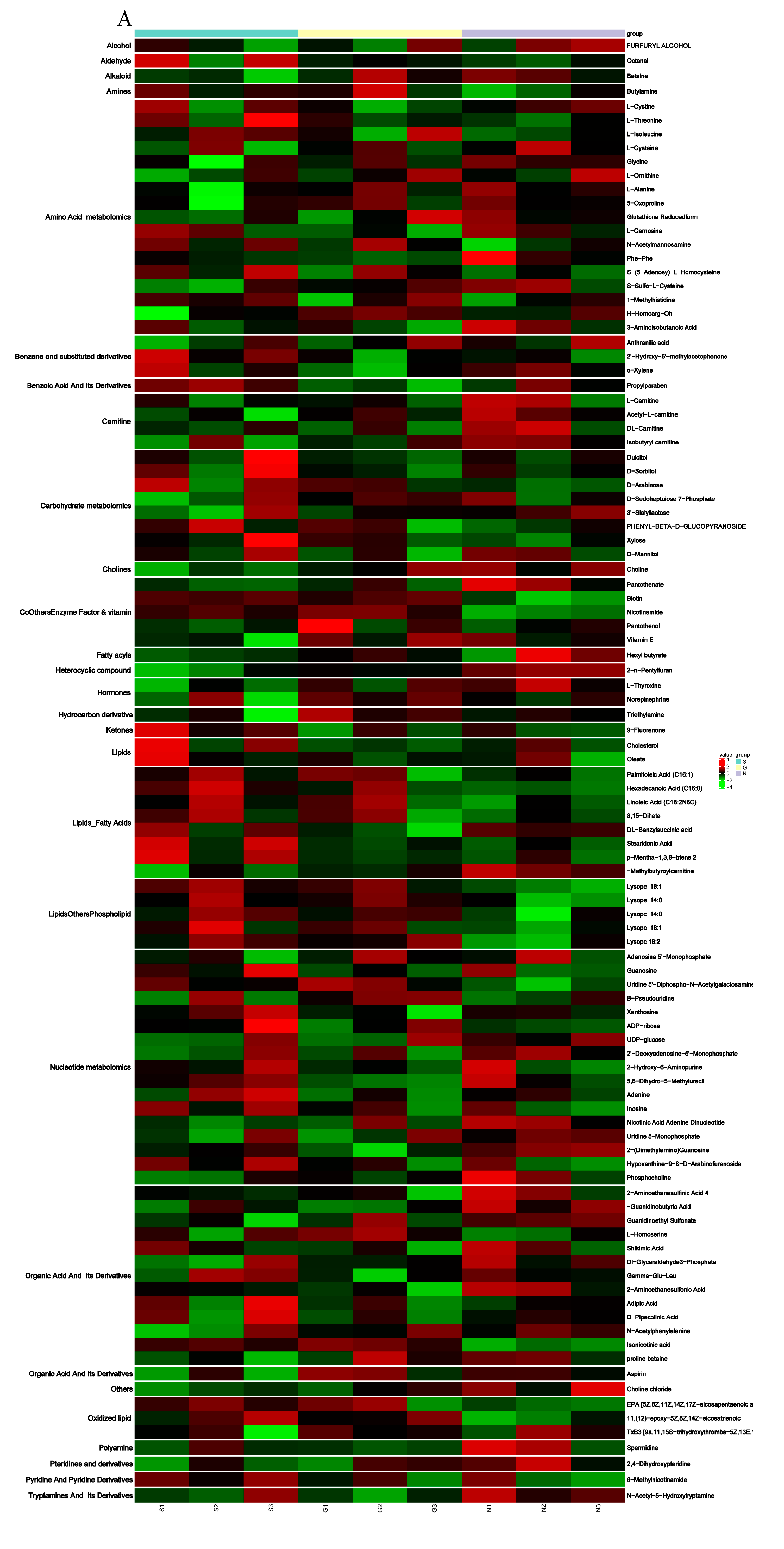
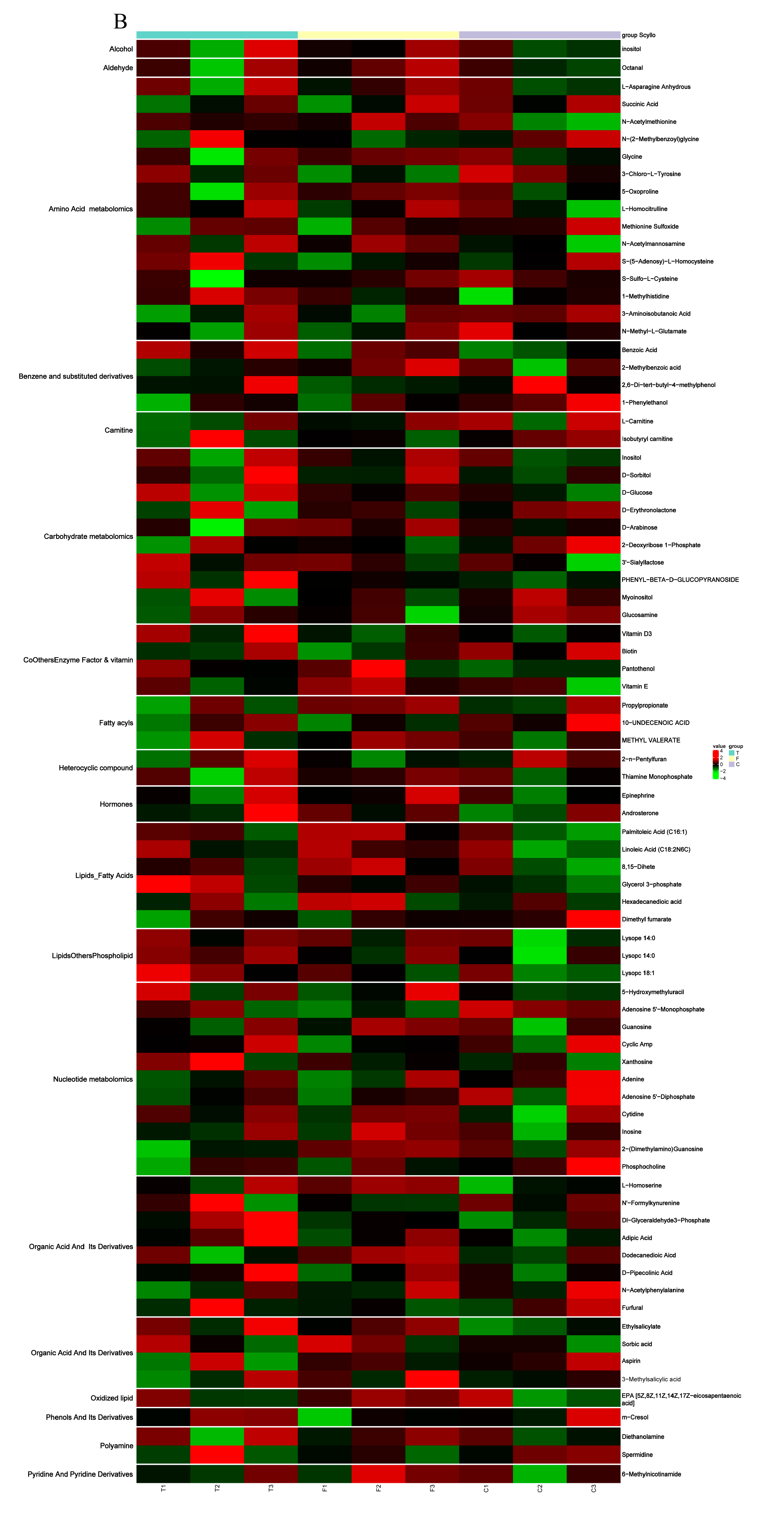
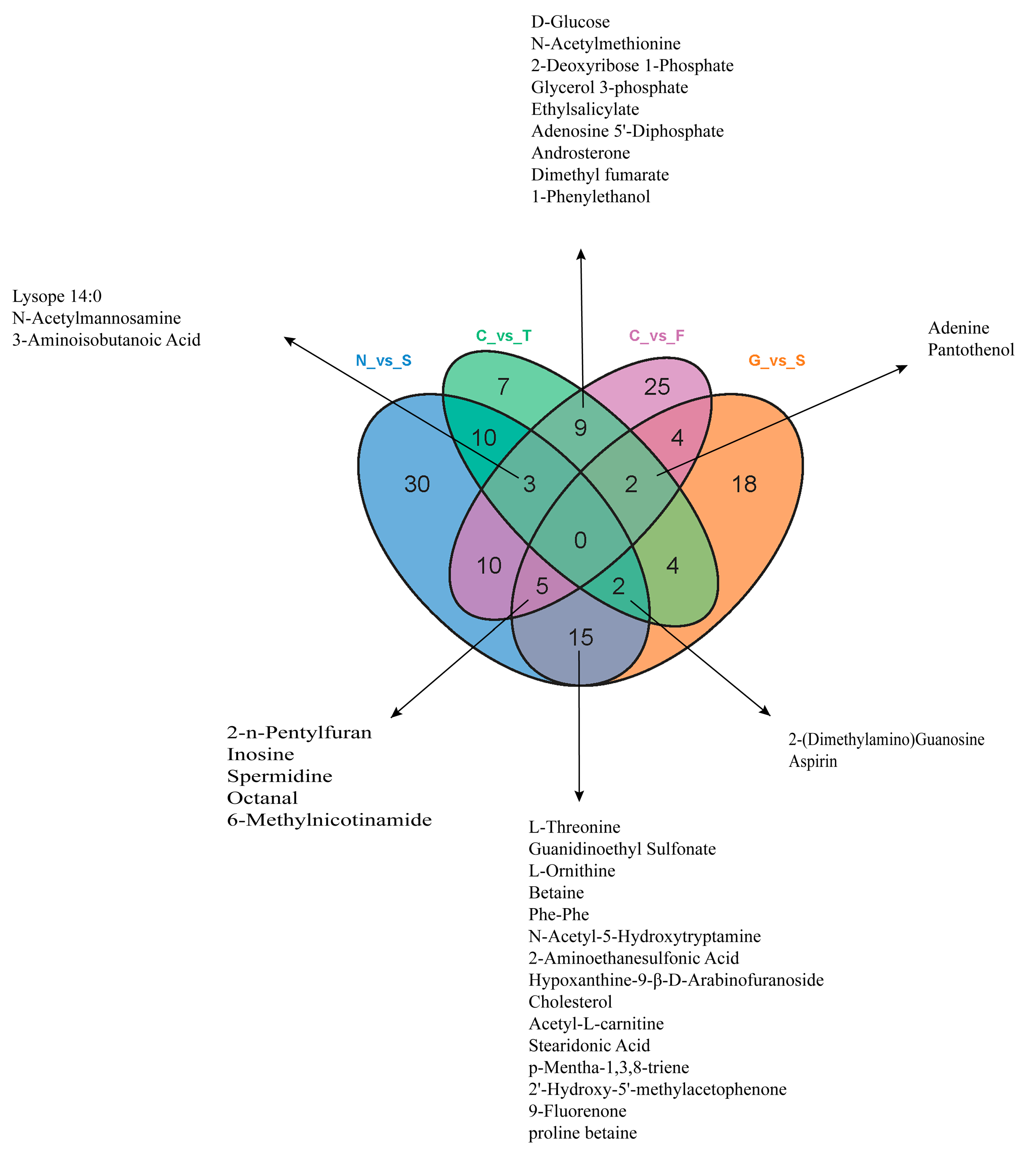
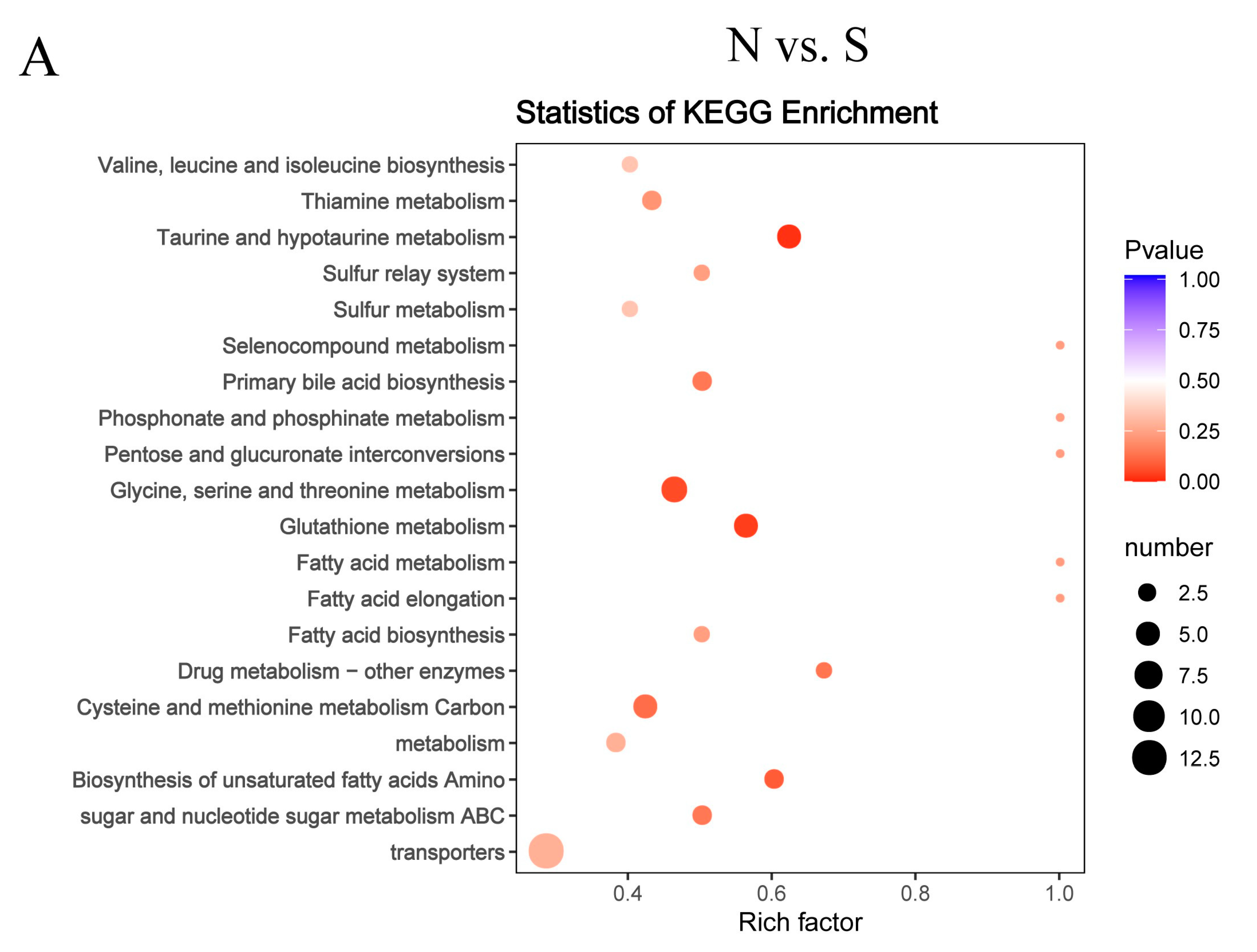
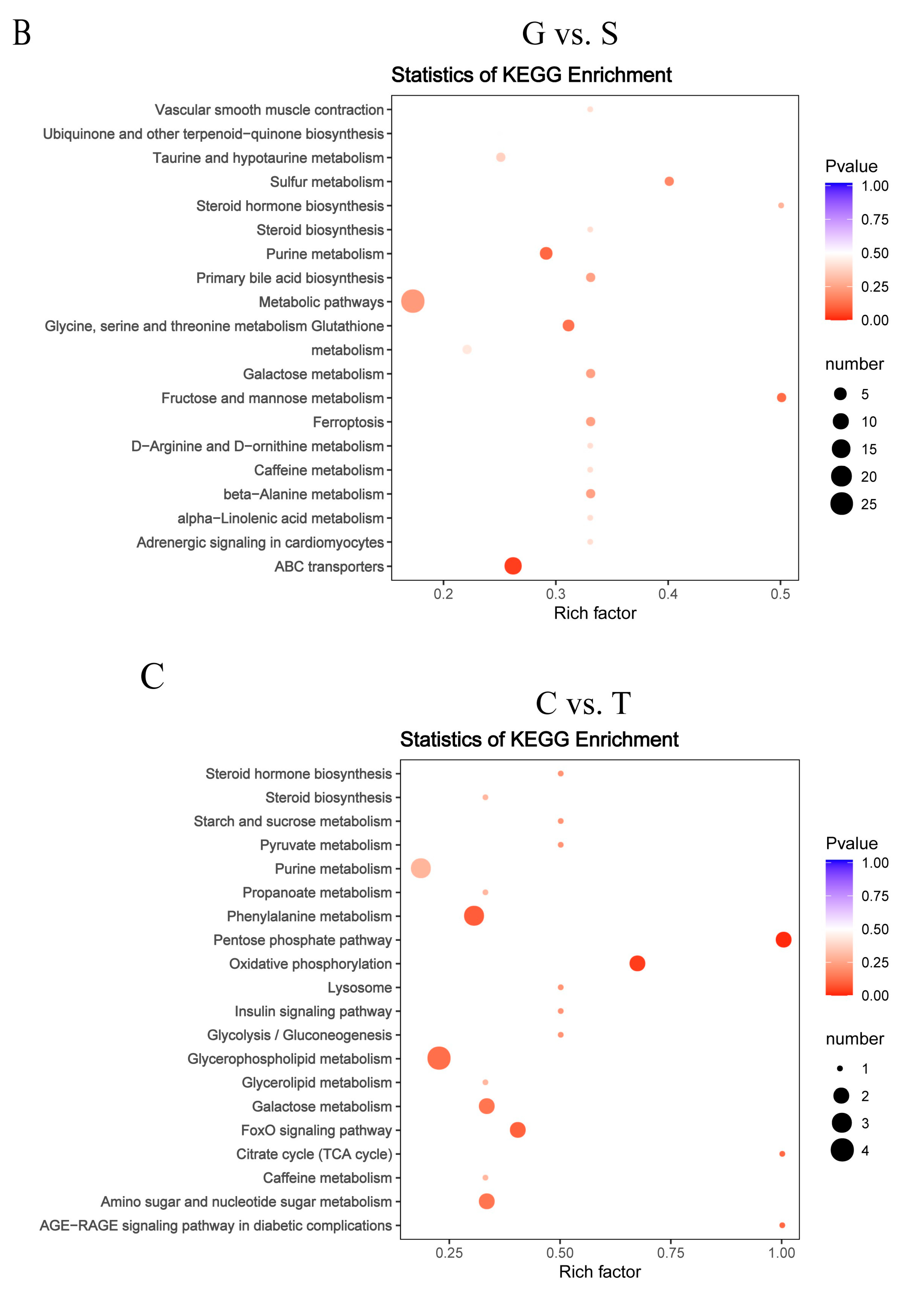
3.4. Metabolite Differences in N vs. S, G vs. S, C vs. T, and C vs. F Comparisons
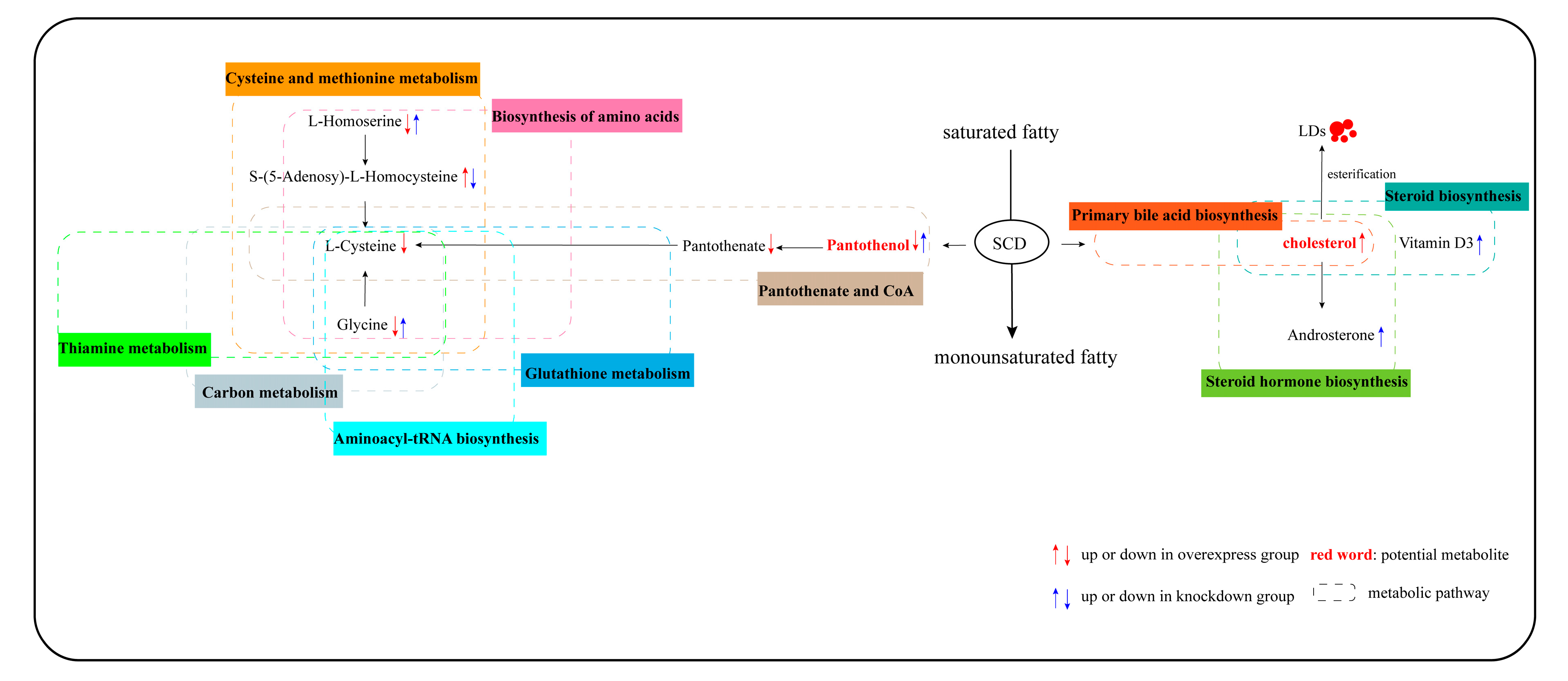
3.5. Pathway Analysis of Differentially Abundant Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CoA | coenzyme A |
| GC | granulosa cell |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LIT | linear ion trap |
| LC-ESI-MS/MS | liquid chromatography-electrospray ionization-tandem mass spectrometry |
| LC-MS/MS | liquid chromatography-tandem mass spectrometry |
| MUFA | monounsaturated fatty acid |
| MRM | multiple reaction monitoring |
| OPLS-DA | orthogonal correction partial least squares discriminant analysis |
| PUFA | polyunsaturated fatty acid |
| PCA | principal component analysis |
| QC | quality control |
| qRT-PCR | quantitative real-time PCR |
| SFA | saturated fatty acid |
| siRNA | small interfering RNA |
| SCD | stearoyl-CoA desaturase |
| QQQ | triple quadrupole |
| VIP | variable importance in projection |
| LD | lipid droplet |
References
- Bai, Y.; Mccoy, J.G.; Levin, E.J.; Sobrado, P.; Rajashankar, K.R.; Fox, B.G.; Zhou, M. X-ray structure of a mammalian stearoyl-CoA desaturase. Nature 2015, 524, 252–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Zou, X.; Chang, Q.; Zhang, Y.; Li, Y.; Zhang, L.; Huang, J.; Liang, B. The evolutionary pattern and the regulation of stearoyl-CoA desaturase genes. BioMed Res. Int. 2013, 2013, 856521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alarcon, G.D.J.; Roco, J.; Medina, A.; Van Nieuwenhove, C.; Medina, M.; Jerez, S. Stearoyl-CoA desaturase indexes and n-6/n-3 fatty acids ratio as biomarkers of cardiometabolic risk factors in normal-weight rabbits fed high fat diets. J. Biomed. Sci. 2016, 23, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamal, S.; Saleem, A.; Rehman, S.; Bibi, I.; Iqbal, H.M.N. Protein engineering: Regulatory perspectives of stearoyl CoA desaturase. Int. J. Bio. Macromol. 2018, 114, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Castro, L.F.C.; Wilson, J.M.; Goncalves, O.; Galanteoliveira, S.; Rocha, E.; Cunha, I. The evolutionary history of the stearoyl-CoA desaturase gene family in vertebrates. BMC Evol. Biol. 2011, 11, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aljohani, A.M.; Syed, D.N.; Ntambi, J.M. Insights into Stearoyl-CoA Desaturase-1 Regulation of Systemic Metabolism. Trends. Endocrin. Met. 2017, 28, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Park, W.J. The Biochemistry and Regulation of Fatty Acid Desaturases in Animals. In Polyunsaturated Fatty Acid Metabolism; AOCS Press: Urbana, IL, USA, 2018; pp. 87–100. [Google Scholar] [CrossRef]
- Shen, J.; Gang, W.; Tsai, A.L.; Ming, Z. Structure and Function of Mammalian Stearoyl-COA Desaturase. Biophys. J. 2018, 114, 426a. [Google Scholar] [CrossRef]
- Frigolet, M.E.; Gutierrezaguilar, R. The Role of the Novel Lipokine Palmitoleic Acid in Health and Disease. Adv. Nutr. 2017, 8, 173S–181S. [Google Scholar] [CrossRef] [PubMed]
- Ran, H.; Zhu, Y.; Deng, R.; Zhang, Q.; Liu, X.; Feng, M.; Zhong, J.; Lin, S.; Tong, X.; Su, Q. Stearoyl-CoA desaturase-1 promotes colorectal cancer metastasis in response to glucose by suppressing PTEN. J. Exp. Clin. Cancer Res. 2018, 37, 1–15. [Google Scholar] [CrossRef]
- Zhao, J.; Zhi, Z.; Wang, C.; Xing, H.; Song, G.; Yu, X.; Zhu, Y.; Wang, X.; Zhang, X.; Di, Y. Exogenous lipids promote the growth of breast cancer cells via CD36. Oncol. Rep. 2017, 38, 2105–2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.M. Fatty Acid and Phospholipid Biosynthesis in Prokaryotes. In Biochemistry of Lipids, Lipoproteins and Membranes; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Uzbekova, S.; Elis, S.; Teixeiragomes, A.; Desmarchais, A.; Maillard, V.; Labas, V. MALDI Mass Spectrometry Imaging of Lipids and Gene Expression Reveals Differences in Fatty Acid Metabolism between Follicular Compartments in Porcine Ovaries. Biology 2015, 4, 216–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vireque, A.A.; Tata, A.; Belaz, K.R.A.; Grázia, J.O.G.V.; Santos, F.N.; Arnold, D.R.; Basso, A.C.; Eberlin, M.N.; Silva De Sá, M.F.; Ferriani, R.A.; et al. MALDI mass spectrometry reveals that cumulus cells modulate the lipid profile of in vitro matured bovine oocytes. Syst. Biol. Reprod. Med. 2017, 63, 86–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suttonmcdowall, M.L.; Gilchrist, R.B.; Thompson, J.G. The pivotal role of glucose metabolism in determining oocyte developmental competence. Reproduction 2010, 139, 685–695. [Google Scholar] [CrossRef] [Green Version]
- Elis, S.; Desmarchais, A.; Maillard, V.; Uzbekova, S.; Monget, P.; Dupont, J. Cell proliferation and progesterone synthesis depend on lipid metabolism in bovine granulosa cells. Theriogenology 2015, 83, 840–853. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.K.; Onions, V.; Kendall, N.R.; Guo, L.; Scaramuzzi, R.J. The effect of monosaccharide sugars and pyruvate on the differentiation and metabolism of sheep granulosa cells in vitro. Reproduction 2010, 140, 541–550. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Qiao, J. Expression and regulation of adipocyte fatty acid binding protein in granulosa cells and its relation with clinical characteristics of polycystic ovary syndrome. Endocrine 2011, 40, 196–202. [Google Scholar] [CrossRef]
- Wen, R.; Gan, X.; Hu, S.Q.; Gao, S.Y.; Deng, Y.; Qiu, J.M.; Sun, W.Q.; Li, L.; Han, C.C.; Hu, J.W.; et al. Evidence for the existence of de novo lipogenesis in goose granulosa cells. Poult. Sci. 2018, 98, 1023–1030. [Google Scholar] [CrossRef]
- Li, Q.; Hu, S.; Wang, Y.; Deng, Y.; Yang, S.; Hu, J.; Li, L.; Wang, J. mRNA and miRNA Transcriptome Profiling of Granulosa and Theca Layers From Geese Ovarian Follicles Reveals the Crucial Pathways and Interaction Networks for Regulation of Follicle Selection. Front. Genet. 2019, 10, 988. [Google Scholar] [CrossRef] [Green Version]
- Bednarski, T.; Olichwier, A.; Opasinska, A.; Pyrkowska, A.; Gan, A.; Ntambi, J.M.; Dobrzyn, P. Stearoyl-CoA desaturase 1 deficiency reduces lipid accumulation in the heart by activating lipolysis independently of peroxisome proliferator-activated receptor α. Biochim. Biophys. Acta 2016, 1861, 2029–2037. [Google Scholar] [CrossRef]
- Dobrzyn, P.; Dobrzyn, A.; Miyazaki, M.; Cohen, P.; Asilmaz, E.; Hardie, D.G.; Friedman, J.M.; Ntambi, J.M. Stearoyl-CoA desaturase 1 deficiency increases fatty acid oxidation by activating AMP-activated protein kinase in liver. Proc. Nat. Acad. Sci. USA 2004, 101, 6409–6414. [Google Scholar] [CrossRef] [Green Version]
- Phillips, R.W.; Johnson, R.G.; Moyer, R.T. China’s Goose Industry. World Poult. Sci. J. 1948, 4, 34–35. [Google Scholar] [CrossRef]
- Johnson, A.L. Organization and Functional Dynamics of the Avian Ovary. In Hormones & Reproduction of Vertebrates; Chapter 3; Academic Press: Cambridge, MA, USA, 2011; Volume 40, pp. 71–90. [Google Scholar] [CrossRef] [Green Version]
- Johnson, P.A. Follicle Selection in the Avian Ovary. Reprod. Domest. Anim. 2012, 47, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, D.K.; Hollywood, K.A.; Goodacre, R. Metabolomics for the masses: The future of metabolomics in a personalized world. New Horiz. Transl. Med. 2017, 3, 294–305. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, J.K.; Lindon, J.C. Systems biology: Metabonomics. Nature 2008, 455, 1054–1056. [Google Scholar] [CrossRef] [PubMed]
- Bell, D.J.; Freeman, B.M. Physiology & Biochemistry of the Domestic Fowl; Academic Press: Cambridge, MA, USA, 1971. [Google Scholar]
- Gilbert, A.; Evans, A.; Perry, M.; Davidson, M. A method for separating the granulosa cells, the basal lamina and the theca of the preovulatory ovarian follicle of the domestic fowl (Gallus domesticus). Reproduction 1977, 50, 179–181. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−△△Ct Method. Methods 2000, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lucki, N.C.; Sewer, M.B. Multiple Roles for Sphingolipids in Steroid Hormone Biosynthesis. In Lipids in Health and Disease; Springer: Dordrecht, The Netherlands, 2008; Volume 49, pp. 387–412. [Google Scholar] [CrossRef] [Green Version]
- Campbell, B.K.; Kendall, N.R.; Onions, V.; Scaramuzzi, R.J. The effect of systemic and ovarian infusion of glucose, galactose and fructose on ovarian function in sheep. Reproduction 2010, 140, 721–732. [Google Scholar] [CrossRef] [Green Version]
- Holt, A.; De Almeida, R.F.M.; Nyholm, T.K.M.; Loura, L.M.S.; Daily, A.E.; Staffhorst, R.W.H.M.; Rijkers, D.T.S.; Koeppe, R.E.; Prieto, M.; Killian, J.A. Is there a preferential interaction between cholesterol and tryptophan residues in membrane proteins. Biochemistry 2008, 47, 2638–2649. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Jung, S.; Kim, N.; Shin, M.J.; Ryu, D.H.; Hwang, G. Myocardial metabolic alterations in mice with diet-induced atherosclerosis: Linking sulfur amino acid and lipid metabolism. Sci. Rep. 2017, 7, 13597. [Google Scholar] [CrossRef] [Green Version]
- Carter, R.N.; Morton, N.M. Cysteine and hydrogen sulphide in the regulation of metabolism: Insights from genetics and pharmacology. J. Pathol. 2015, 238, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; He, L.; Wan, D.; Yang, H.; Yao, K.; Wu, G.; Wu, X.; Yin, Y. Methionine restriction on lipid metabolism and its possible mechanisms. Amino Acids 2016, 48, 1533–1540. [Google Scholar] [CrossRef]
- Le, T.T.; Ziemba, A.; Urasaki, Y.; Hayes, E.; Brotman, S.; Pizzorno, G. Disruption of uridine homeostasis links liver pyrimidine metabolism to lipid accumulation. J. Lipid Res. 2013, 54, 1044–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, C.N.; LoVullo, E.D.; Kijek, T.M.; Fuller, J.R.; Brunton, J.C.; Steele, S.P.; Taft-Benz, S.A.; Richardson, A.R.; Kawula, T.H. PanG a New Ketopantoate Reductase Involved in Pantothenate Synthesis. J. Bacteriol. 2013, 195, 965–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maccallum, J.L.; Bennett, W.F.D.; Tieleman, D.P. Distribution of Amino Acids in a Lipid Bilayer from Computer Simulations. Biophys. J. 2008, 94, 3393–3404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarling, E.J.; Vallim, T.Q.D.A.; Edwards, P.A. Metabolism, Role of ABC transporters in lipid transport and human disease. Trend. Endocrin. Met. 2013, 24, 342–350. [Google Scholar] [CrossRef] [Green Version]
- Borroni, M.V.; Vallés, A.S.; Barrantes, F.J. The lipid habitats of neurotransmitter receptors in brain. BBA Biomembr. 2016, 1858, 2662–2670. [Google Scholar] [CrossRef]
- Dridi, S.; Taouis, M.; Gertler, A.; Decuypere, E.; Buyse, J. The regulation of stearoyl-CoA desaturase gene expression is tissue specific in chickens. J. Endocrin. 2007, 192, 229–236. [Google Scholar] [CrossRef] [Green Version]
- Mauvoisin, D.; Mounier, C. Hormonal and nutritional regulation of SCD1 gene expression. Biochimie 2011, 93, 78–86. [Google Scholar] [CrossRef]
- Matsuda, F.; Inoue, N.; Manabe, N.; Ohkura, S. Follicular Growth and Atresia in Mammalian Ovaries: Regulation by Survival and Death of Granulosa Cells. J. Reprod. Dev. 2012, 58, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Tesfay, L.; Paul, B.T.; Konstorum, A.; Deng, Z.; Research, S.V. Steroyl-CoA Desaturase 1 (SCD1) protects ovarian cancer cells from ferroptotic cell death. Cancer. Res. 2019, 79, 5355–5366. [Google Scholar] [CrossRef]
- Igal, R.A. Stearoyl CoA desaturase-1: New insights into a central regulator of cancer metabolism. BBA Mol. Cell Biol. Lipid 2016, 1861, 1865–1880. [Google Scholar] [CrossRef] [PubMed]
- Koeberle, A.; Loser, K.; Thurmer, M. Stearoyl-CoA desaturase-1 and adaptive stress signaling. BBA Mol. Cell Biol. Lipid 2016, 1861, 1719–1726. [Google Scholar] [CrossRef] [PubMed]
- Diot, C.; Lefevre, P.; Herve, C.; Belloir, B.; Narce, M.; Damon, M.; Poisson, J.; Mallard, J.; Douaire, M. Stearoyl-CoA desaturase 1 coding sequences and antisense RNA affect lipid secretion in transfected chicken LMH hepatoma cells. Arch. Biochem. Biophys. 2000, 380, 243–250. [Google Scholar] [CrossRef]
- Flowers, M.T.; Ntambi, J.M. Role of stearoyl-coenzyme A desaturase in regulating lipid metabolism. Curr. Opin. Lipidol. 2008, 19, 248–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, J.; Chang, T.Y. Further characterization of a Chinese hamster ovary cell mutant requiring cholesterol and unsaturated fatty acid for growth. Biochemistry 1982, 21, 3196–3202. [Google Scholar] [CrossRef]
- Gao, S.; Gan, X.; He, H.; Hu, S.; Deng, Y.; Chen, X.; Li, L.; Hu, J.; Li, L.; Wang, J. Dynamic characteristics of lipid metabolism in cultured granulosa cells from geese follicles at different developmental stages. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [PubMed]
- Dunning, K.R.; Russell, D.L.; Robker, R.L. Lipids and oocyte developmental competence: The role of fatty acids and β-oxidation. Reproduction 2014, 148, R15–R27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prates, E.G.; Nunes, J.T.; Pereira, R.M. A Role of Lipid Metabolism during Cumulus-Oocyte Complex Maturation: Impact of Lipid Modulators to Improve Embryo Production. Mediat. Inflamm. 2014, 692067. [Google Scholar] [CrossRef]
- Leonardi, R.; Zhang, Y.M.; Rock, C.O.; Jackowski, S. Coenzyme A: Back in action. Prog. Lipid Res. 2005, 44, 125–153. [Google Scholar] [CrossRef]
- Dansie, L.E.; Reeves, S.; Miller, K.; Zano, S.P.; Frank, M.; Pate, C.; Wang, J.; Jackowski, S. Physiological roles of the pantothenate kinases. Biochem. Soc. Trans. 2014, 42, 1033–1036. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sense Sequence (5′-3′) | Antisense Sequence (5′-3′) |
|---|---|---|
| siRNA210 | GCGAUACGUCUGGAGGAAUTT | AUUCCUCCAGACGUAUCGCTT |
| siRNA405 | GCGGAUCUUCUUGACUAUUTT | AAUAGUCAAGAAGAUCCGCTT |
| siRNA774 | GCUCAACGCCACUUGGCUATT | UAGCCAAGUGGCGUUGAGCTT |
| siRNA-scrambled | UUCUCCGAACGUGUCACGUTT | ACGUGACACGUUCGGAGAATT |
| primers for GFP-SCD | CCGCTCGAGATGGAGAAGGACTTACTCAGTCATG | CCCAAGCTTTCAGCCGCTCTTGTGACTCCC |
| Pathway | N vs. S | G vs. S | C vs. T | C vs. F | Lipid-Related Functions Reported | Reference |
|---|---|---|---|---|---|---|
| Steroid hormone biosynthesis/Steroid biosynthesis | Cholesterol | Cholesterol | Androsterone/Vitamin D3 | Androsterone | Steroid hormones are essential regulators of a vast number of physiological processes. | [31] |
| Galactose metabolism | UDP-glucose | D-Sorbitol; Dulcitol | D-Glucose; D-Sorbitol | D-Glucose; Myoinositol | Galactose exerts primarily suppressive effects of ovarian follicle number and steroid secretion by direct actions on the ovary. | [32] |
| Tryptophan metabolism | N-Acetyl-5-Hydroxytryptamine; Anthranilic acid | N-Acetyl-5-Hydroxytryptamine | Succinic Acid | Epinephrine | Molecular modeling studies suggested favorable stacking interactions between cholesterol and tryptophan, in which the face of the complex ring system of cholesterol and the indole ring of tryptophan build the interaction interface. | [33] |
| Sulfur metabolism | L-Cysteine; 2-Aminoethanesulfonic Acid | 2-Aminoethanesulfonic Acid; L-Homoserine | Succinic Acid | L-Homoserine | Recently, increasing attention has been paid to the role of sulfur amino acids in regulating lipid metabolism. | [34] |
| Cysteine and methionine metabolism | L-Cysteine; S-Sulfo-L-Cysteine; L-Alanine; Glutathione Reducedform; S-(5-Adenosy)-L-Homocysteine | L-Homoserine; L-Cystine | S-Sulfo-L-Cysteine | L-Homoserine; S-(5-Adenosy)-L-Homocysteine | Much attention has been recently focused on the effects of methionine restriction and cysteine on metabolic health, especially lipid metabolism. | [35,36] |
| pyrimidine metabolism | Uridine 5-Monophosphate; UDP-glucose | Β-Pseudouridine | Cytidine; 2-Deoxyribose 1-Phosphate | 2-Deoxyribose 1-Phosphate | Given the link between pyrimidine metabolism and liver lipid accumulation, there is a potential for the use of nucleosides and nucleoside analogs in the treatment of fatty liver conditions. | [37] |
| Pantothenate and CoA biosynthesis | Pantothenate; L-Cysteine | Pantothenol | Pantothenol | Pantothenol | Pantothenate forms the core of CoA and is a precursor to acyl carrier protein (ACP), making it essential in both energy and lipidmetabolism. | [38] |
| Biosynthesis of amino acids | Anthranilic acid; Glycine; S-Sulfo-L-Cysteine; S-(5-Adenosy)-L-Homocysteine; L-Threonine; L-Alanine; L-Cysteine; L-Ornithine; L-Isoleucine | L-Ornithine; L-Homoserine; L-Threonine; Shikimic Acid | S-Sulfo-L-Cysteine | L-Homoserine; Glycine; L-Asparagine Anhydrous; S-(5-Adenosy)-L-Homocysteine | Amino acids were distributed in a lipid bilayer. | [39] |
| ABC transporters | 2-Aminoethanesulfonic Acid; Spermidine; proline betaine; Glycine; Betaine; Glutathione Reducedform; L-Ornithine; L-Isoleucine; Biotin; Choline; L-Threonine; L-Alanine; Inosine | Betaine; D-Sorbitol; Inosine; L-Cystine; L-Threonine; L-Ornithine; D-Mannitol; Spermidine; Guanosine; proline betaine; 2-Aminoethanesulfonic Acid; Xanthosine | D-Sorbitol; Xanthosine; Glycerol 3-phosphate; D-Glucose; Cytidine | Biotin; Myoinositol; Spermidine; Glycerol 3-phosphate; Glycine; Inosine; Guanosine; D-Glucose | ATP binding cassette (ABC) transporter proteins are thought to facilitate the ATP-dependent translocation of lipids or lipid-related compounds—such substrates include cholesterol, plant sterols, bile acids, phospholipids and sphingolipids. | [40] |
| Neuroactive ligand-receptor interaction | 2-Aminoethanesulfonic Acid; L-Thyroxine; Glycine | 2-Aminoethanesulfonic Acid; Norepinephrine | Adenosine 5′-Diphosphate | Adenosine 5′-Diphosphate; Epinephrine; Glycine | Modulation of neurotransmitter receptors by lipids occurs at multiple levels, affecting a wide variety of activities, including their trafficking, sorting, stability, residence lifetime at the cell surface, endocytosis, and recycling, among other important functional properties at the synapse. | [41] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, X.; Hu, S.; Li, L.; Liu, H.; He, H.; Wang, J. Metabolomic Analysis of SCD during Goose Follicular Development: Implications for Lipid Metabolism. Genes 2020, 11, 1001. https://doi.org/10.3390/genes11091001
Yuan X, Hu S, Li L, Liu H, He H, Wang J. Metabolomic Analysis of SCD during Goose Follicular Development: Implications for Lipid Metabolism. Genes. 2020; 11(9):1001. https://doi.org/10.3390/genes11091001
Chicago/Turabian StyleYuan, Xin, Shenqiang Hu, Liang Li, Hehe Liu, Hua He, and Jiwen Wang. 2020. "Metabolomic Analysis of SCD during Goose Follicular Development: Implications for Lipid Metabolism" Genes 11, no. 9: 1001. https://doi.org/10.3390/genes11091001
APA StyleYuan, X., Hu, S., Li, L., Liu, H., He, H., & Wang, J. (2020). Metabolomic Analysis of SCD during Goose Follicular Development: Implications for Lipid Metabolism. Genes, 11(9), 1001. https://doi.org/10.3390/genes11091001